0022-538X/93/095487-09$02.00/0
CopyrightX 1993, American Society for Microbiology
Transformation by the fos
or
jun
Oncogene
Does Not
Increase AP-1
DNA-Binding Activity
KIML. HAWKER,' ALEX
PINTZAS,lt
ROBERT F. HENNIGAN,2 DAVID A. F.GILLESPIE,'ANDBRADFORD W.
OZANNE"*
Cancer Research Campaign Laboratories, BeatsonInstituteforCancerResearch, GarscubeEstate, SwitchbackRoad, Bearsden, Glasgow G61 IBD, United Kingdom,' and Department of Microbiology,
University of Texas Southwestern Medical Center, Dallas, Texcas 75235 Received 17 February 1993/Accepted 15 June 1993
The c-fos and c-jun proto-oncogenes encode components of the transcription factor AP-1. To determine whether transformation by the v-fosorv-jun oncogeneresultsinalterations inthe levelorregulation of this
factor, we have characterized AP-1 DNA-binding activity in nuclear extracts prepared from v-fos- and c-fos-transformed rat fibroblast cell lines and v-jun-transformed chicken embryo fibroblasts under various growth conditions. Durngproliferation, the level of AP-1 DNA-binding activity does notdiffer amongthe
v-fos-, c-fos-,orv-jun-transformed cells and their normal progenitors, despiteconstitutive overexpression of thecorresponding oncoproteins. Therefore, althoughnecessary,itisnot
likely
thatanincrease in DNA bindingis sufficient forfos orjun transformation. Normalratandchicken fibroblasts demonstrateverylowlevels of AP-1 DNA-bindingactivitywhenquiescent, and upon serumstimulation abiphasic increase is observed. A similarcyclical pattern is seeninv-fos-transformed cells, but in v-jun-transformed cellsAP-1 DNA-binding activitydoes not fluctuate inresponsetoserum stimulation,which suggeststhat this level ofcontrolmaybe exertedthroughtheJun componentof theAP-1 complex.
The FBJ (50) andFBR(16)mouse osteosarcomaviruses, which encodethefosoncogenes,transformratfibroblasts in culture and induce osteosarcomas in mice. The avian sar-comavirusASV17 retrovirus(37), encodingthev-jun onco-gene,induces focus formation in chickenembryofibroblasts (CEF) and fibrosarcomas in chickens. Thev-fos andv-jun
genes are retroviral homologs of the cellular
proto-onco-genes c-fos and c-jun, which encode components of the dimeric transcription factorAP-1 (52).AP-1 wasoriginally defined as a factor able to bind to the DNA consensus sequenceTGA(C/G)TCAandtransactivate asetofgenesin
responsetothephorbolester
12-O-tetradecanoylphorbol-13-acetate(2, 4, 26).
Linking the c-fos gene to a long terminal repeat allows constitutiveoverexpressionandconsequentlycellular trans-formation(38).Alongterminalrepeatisalsoresponsiblefor constitutivev-fos expressionuponinfection with either FBR
or FBJ mouse osteosarcoma virus (38). However, these
viruses transform more efficiently than p55c-Fos due to changesinthe amino acidsequencewhich contribute toward their increased oncogenic potential. The absence of C-ter-minal serine residues which are potential phosphorylation sites may change the DNA-binding activity and activation potentialof the viralproteins(42).Inparticular,
p75ag-fos-fa
demonstrates an immortalization potential and increased transactivation as aresult of the single amino acid substitu-tion ofglutamate138 tovaline (36).c-fos belongs to a multigene family (13, 14, 41, 55). The other members, fosB, fra-1, andfra-2, all show varying degrees of sequence similarity, with particularly high con-servation in the basic and leucinezipperdomains(32),which mediate DNAbinding anddimerization, respectively. Both
*Correspondingauthor.
tPresent address: Institut de Chimie Biologique, Faculte de
Medecine,67085Strasbourg Cedex,France.
FBJ p55-FOS and FBR p75vFOS also retain the basic and leucinezipper domains, permitting participation inthe for-mation of the AP-1transcriptionfactor incollaborationwith Jun(23, 29, 44).These domainsarenecessaryforoncogenic transformation becausemutantsof FBRp75VF0s lacking the leucinezipperfailtodimerize,bindDNA,ortransform cells
(36).
ASV17 was originally isolated from a chicken fibrosar-comabyCavalierietal.(11)and encodesapolypeptideof 65 kDain whichpartof theJunmoleculeis fusedtoviral Gag
sequences (37). Like the viral Fos proteins, p65Junis a morepotenttranscriptionalactivator thanits cellular
coun-terpart,p39cJun. This difference isthoughttoresult froma loss ofregulatory phosphorylation (7, 10, 43, 49) and the deletion ofadomain termed deltaadjacenttothe transacti-vationdomain(7, 8). c-junis alsoamember ofagenefamily
(24, 47).The othermembers, junBandjunD,also bind AP-1 siteseitherashomodimersorheterodimers witheach other orwithmembers of thec-fosgenefamily(3, 4, 9, 18, 45, 51). Fos-Jun heterodimers displaya higher affinityfor the AP-1 DNArecognitionsequencethan do Junhomodimers (11, 12,
20, 39, 45).
Retroviralexpressionof thev-fosandv-jungenesleadsto
alargeincrease in theexpressionofpotentialAP-1
compo-nents.It isgenerallyconsidered thatthis would result in an increase in the level of AP-1 activity and lead to the deregulation of AP-1-responsive genes and oncogenlesis. Thishypothesisissupported bytheobservationthat Fos and Jun cooperate in transformation (40). Furthermore, c-fos plays acrucial role in signal transduction, is necessaryfor cellcycle progression,and is involved in the transformation
process initiated by several oncogenes, including v-sis,
v-src, c-Ha-ras,v-mos, andv-raf (27, 28, 34, 46, 48, 54).
We considered that cells transformed by v-fos orv-jun maydemonstrateadifferencein AP-1DNA-binding activity whencomparedwith normal cells. However,mutantstudies ofFBRp75vFosrevealed that functions other than
transac-5487
on November 9, 2019 by guest
http://jvi.asm.org/
tivationareinvolved in the induction of transformation (36), suggesting that a qualitative change in the AP-1 activity of a transformed cell may be more important than a quantitative one. To explore this possibility, we studied in vitro AP-1 DNA-binding activity in normal cells, those transformed with the viral oncogenes, and a rat fibroblast cell line which has continuous expression of the c-fos proto-oncogene (33, 38).We report that AP-1DNA-binding activity is biphasic in the cell cycle and is not increased by the presence of the v-fos or thev-jun oncogene.
MATERIALS AND METHODS
Cells and cell culture.208Fratfibroblastcellsinfected with either the FBJ or FBR forms of the v-fos oncogene and transfected with the c-fos MMV construct were obtained from T. Curran (Hoffman-La Roche Inc., Nutley, N.J.).
All cell lines were grown in Dulbecco's modification of Eagle's minimal essential medium (DMEM) supplemented with 10% fetal bovine serum (FBS). Subconfluent cultures were arrested in
Go
by incubation for 64 h in DMEM containing 0.5%FBS.Quiescent cells were stimulated by the addition of FBS toafinal concentration of 10%.CEF were cultured in DMEM supplemented with 10% tryptose phosphate, 10% newborn calf serum, and 2% chickenserum. ASV17 virus was regenerated by transfect-ingCEFwithamolecular clone of ASV17 (37) together with RCAS (25) as a helper virus (7), and the culture was passaged until uniform transformation was evident. For serum stimulation experiments, CEF cultures were incu-bated in DMEM supplemented with 0.5% newborn calf serumfor 24 h and then stimulated by the addition of chicken serumto afinal concentration of 10% for the indicated times
(see
Fig. 4).Serum induction assays. Cells were seeded into 96-well tissue cultureplates at104cells per well in 10% serum. After 24h, the cells were washed twice with serum-free medium andplacedin0.5% serumfor anadditional 48 h. Serum was addedto cells for 12to 18 h, and then [3H]thymidine to 0.3 ,uCiperwellwasadded for 4 h. The cells were removed from the substrate by trypsinization, harvested, and washed on glass fiber filters by a multiwell harvester (Skatron Inc., Tranby,Norway). The incorporated [3H]thymidinewas de-terminedby scintillation counting.
RNAextraction and Northern (RNA) blotting.Total cellu-lar RNAwasprepared with the use of RNAzol B (Promega). A10-,ugaliquotwaselectrophoretically fractionated on a 1% formaldehyde denaturing agarose gel, blotted onto nylon membrane (Amersham), and hybridized according to stan-dard techniques. The EcoRI-HindIII murine c-fos insert of Sp65 was randomly primed and used as the probe. A randomly primed cDNA GAPDH fragment was used as a loading control.
Antibodies, immunoprecipitation, and Western immuno-blotting. The anti-Jun antiserum 948 is a polyclonal anti-Jun serumraisedagainst the carboxy-terminal 82 amino acids of avian c-Jun(17). Apolyclonal anti-Fos antiserum, 388, was generatedby immunizing New Zealand White rabbits with a bacterial TrpE-Fos fusion protein encoding a 77-amino-acid regionofthe FBJ mouse osteosarcoma virus v-Fos protein immediatelyupstream of and including the basic region (52). Reactivity of the immune serum against Fos proteins was verifiedbyimmunoprecipitation of the products of in vitro translation of a full-length mouse c-fos DNA and of mouse, rat, and chicken c-Fos proteins from metabolically labelled cell extracts (unpublished results). The anti-Gag antiserum
(5202) was prepared in rabbits against detergent-disrupted avian myeloblastosis virus particles and reacts primarily with gag-encoded proteins (15). Cells were labelled for a period of 2 h with
EXPRE35S35S-Dual
35S-protein
labelling mix(New England Nuclear)in methionine-free DMEM.Cell lysis and immunoprecipitation were performed in RIPA buffer (150 mM sodium chloride, 0.5% Nonidet P-40, 0.5% deoxycholate, 0.05% sodium dodecyl sulfate [SDS], 50 mM Tris [pH 7.5], and 0.5% aprotinin). Cell debri was removed bycentrifugation at 86,600 xgfor 30min, and samples were precleared with preimmune serum and protein A-Sepharose before immunoprecipitation. The immunoprecipitated com-plexes were collected on protein A-Sepharose beads, washed threetimes with RIPA buffer, and boiled for 10 min in sample loading buffer(57 mM Tris [pH 6.8], 10% glycerol, 20% SDS, and 0.1% 13-mercaptoethanol) before electro-phoretic separation.Nuclear protein extractions. All buffers were made fresh, and inhibitors were added to the following concentrations immediately before use: 5 mM dithiothreitol (DTT), 10 mM
3-glycerophosphate,
1 mMphenylmethylsulfonyl
fluoride,
0.5 mM benzamidine, 1 mM leupeptin, and 1 mM pepstatin. Cell pellets were resuspended in 5 ml of hypotonic solution (10 mM Tris [pH 7.4], 10 mM sodium chloride, 1.6 mM magnesium chloride, and 1 mM calcium chloride). Efficient cell lysis was achieved by the addition of Triton X-100 to 0.1% and 20 strokes with a homogenizer. Nuclear pellets were collected by centrifugation at 900 x g for 10 min, and then the pellets were washed in 10 ml of TMS (0.25 M sucrose, 5 mM magnesium chloride, and 10 mM Tris [pH 7.4])beforea second centrifugation for 10 min at 1,400 x g. The nuclei were resuspended in 100,ulof storage buffer (50 mM sodium chloride, 20 mM HEPES [N-2-hydroxyethylpi-perazine-N'-2-ethanesulfonic acid] [pH 7.9], 5 mM magne-sium chloride, and 20% glycerol). Sodium chloride was addedto 40 mM, and the nuclei were incubated on ice for 10 to 20min. After centrifugation at 164,000 x g for 60 min, the supernatant was removed and stored at -70°C.
Determination of protein concentration. The protein con-centration of the nuclear extracts was determined by an adaptation of the method of Lowry et al. (35). The extracts were incubated at 37°C for 30 min in a 50:1 mixture of bicinchoninic acid solution-copper II sulfate solution, and the resulting optical density was measured on a Dynatech enzyme-linked immunosorbent assay plate reader.
DNA-binding assays. The oligonucleotide used for the detection of AP-1 complexes has been previously described (17). Annealing and end labelling were performed by using standard procedures. DNA binding was done for 30min at 20°C in binding buffer (10 mM HEPES [pH 8.0], 0.5 mM EDTA, 100 mM sodium chloride, 10 mM magnesium chlo-ride, 1 mMDTT, 10% glycerol, and 0.1 mg of bovine serum albumin) with 20 jig of nuclear extract-5 mM DTT-9 jig poly(dI-dC)-poly(dI-dC). Bound and free oligonucleotides wereseparated electrophoretically on a 5% polyacrylamide gel at 180 V. Nuclear extracts prepared from HeLa cells were used as a positivecontrol for the determination of AP-1 DNA-binding activity. To determine Fos and Jun participa-tion in the AP-1complexes, 4 jil of antiserum specific for the twoproteins was added following normal equilibration of the AP-1 complex. Incubation proceeded for a further 15 min and the super-shift was observed by gel electrophoresis as described above.
Densitometry. Autoradiograms were scanned with the Molecular Dynamics scanning laser densitometer and
on November 9, 2019 by guest
http://jvi.asm.org/
N
B
v.<
I _
A
- > ._2;~WV
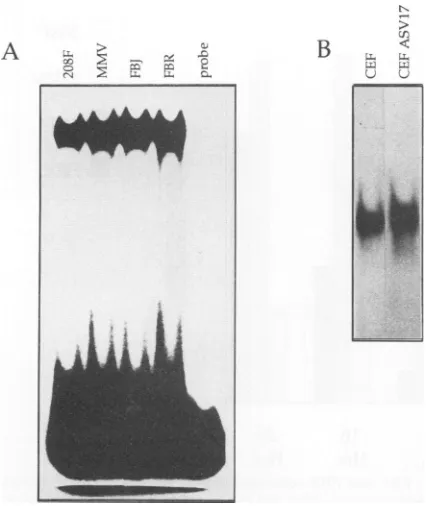
FIG. 1. Acomparison ofAP-1DNA-binding activityin logarith-mically growing, normal, and transformed cell lines. Nuclear ex-tracts werepreparedfrom each cell line andincubated for 30 min withanend-labelled E3AP-1oligonucleotide.Bound andfree probe wereresolved on a5%polyacrylamidegel.
lyzed on a SunSparc station with the Protein Database Incorporated imageanalysis software.
RESULTS
Cells transformed with v-Fos, c-Fos, or v-Jun do not demonstrate anincrease inAP-1 DNA-binding
activity
when compared with normal cells. Conventional gel retardation methodswereutilizedtoaddresswhether retroviral expres-sion of the fos orjun gene leads to a change in AP-1 DNA-binding activity that results in transformation and oncogenesis. Initially, the AP-1 DNA-binding activities ofnuclear extracts prepared from four logarithmicallygrowing rat fibroblast cell lines (208F, MMV, FBJ, and FBR) were compared. Equal quantities of nuclear protein from each cell line were incubated with32P-labelled AP-1-containing oligo-nucleotidederivedfrom the adenovirus E3 gene. The inten-sity of the resulting gel shift is a measure of the AP-1 DNA-binding activity of each nuclear protein extract. No increase in AP-1 activity was obvious in the v-Fos- or c-Fos-transformed cells when compared with normal cells (Fig. 1A). To obtain a more accurate measure of AP-1 DNA-binding activity, the intensity of each shift was mea-sured densitometrically. In the example shown, the absor-bance readings of 153, 185, 186, and 184 for 208F, MMV, FBJ, and FBR cells, respectively, indicate that AP-1 DNA-binding activity in each cell line is similar under these conditions; further experiments support this observation (Fig. 2A; see Fig. 4). The intensity of the retarded band decreases as an increasing quantity of cold E3 AP-1 oligo-nucleotide is included in the band-shift reaction (data not shown). This observation suggeststhat the retarded band is aconsequenceof the presence of the E3 AP-1 oligonucleo-tideand, again, that similarquantities of AP-1 DNA-binding activity exist in the four cell types. Identical results were obtainedwhen thisexperimentwas repeated with an oligo-nucleotide derived from the transin AP-1 recognition se-quence(data not shown).
In a similarexperiment, AP-1 DNA-binding activity was examined in nuclear extracts prepared from cultures of normal primary CEF infected with helper virus or CEF infected and uniformly transformed by ASV17, which ex-presshigh levels of the p65^-Jun oncoprotein (see below). As shown inFig. 1B, the overall level of AP-1-specific DNA-binding activityinASV17-transformed CEF is similartothat seeninnormal CEF. However, the ASV17-transformed cell extracts contain a novel AP-1 DNA-binding complex of slower mobility which is not seen innormal CEF extracts (Fig. 2B). As discussed below, antibody addition experi-ments suggested that the complex of slower mobility con-tains thep65vrJun oncoprotein.
Fos and Junareconstituents of theAP-1complex in normal and transformed cells. To confirm that thegel shift identified in the last experiment is indeed AP-1, we considered it important to demonstrate the presence of Fos- and Jun-related proteins in this complex. Preimmune serum and
208F MMV FBJ FBR
_- -- - - __ z
-- zor C.-It O- -,, st rt C.zt -c
- e, X et1 X, r e? Xl C" c X
B
CEF
[image:3.612.81.294.70.323.2]aml
CEFASV1 7FIG. 2. Anti-fosandanti-jun antisera demonstrate the participation of Fos and Jun family members in AP-1 complex formation.AP-1 DNA
bindingwascarriedoutasbefore, usingthesamenuclearextractsasthoseinFig. 1. Preimmuneserum(pl)andantiseratoFos(388),Jun
(948),andGag (5202)wereaddedtothe reaction for15min before gelelectrophoresis.
A
AR-I
r Zr ZC
C Zr Zr
-e Z - - Z
-,
U,:.
tqw.
.'.W
ffAmp-fm.m
L-
o,--'4
Ir :c 1,
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.112.525.537.688.2]8000.00
-7000.00
-6000.00
-a
m
4000.00-a 3000.00
-2000.00
-1000.00
-0.00
D
208F
El
MMV
E
FBJ
* FBR
n
ii
-24
0
4
8
12
16
20
24
[image:4.612.78.534.63.306.2]HMs.
Hrs.
Hrs.
HMs.
Hirs.
Hrs.
Hirs.
Hrs.
FIG. 3. High-density thymidine uptakeassays in0.5%FBS.208F,MMV,FBJ,and FBR cells wereseededin0.5% FBSandpulsed with [3H]thymidine.Afterthe cells had becomequiescent, thecultureswereadjustedto10%FBS andtheincorporation of
[3H]thymidine
wasmonitored forafurther24h.
antisera specificfor Fos and Junwere used to analyze the observed AP-1 complexes inlogarithmically growing cells, initially from each of the four rat fibroblast cell lines. Anti-Fos antiserum 388 was raised against the basic and leucine zipper domains of
p55c-fos
and thereforewould be expected to cross-react with the other proteins of thefos genefamily.Anti-Jun antiserum 948 isrelatively specificfor c-Jun andcross-reactsweakly,ifatall,withJunB and JunD (24). Anti-Jun antiserum mediatesa supershiftofa propor-tion of the originalretarded complex,whereas the anti-Fos serum largely disrupts the retarded band and little or nosuper-shiftis apparent(Fig. 2).Both theseresults, supershift anddisruption, implythat the antiserarecognizeandinteract with components of thecomplex. Therefore, weconcluded that thegel-shiftrepresentsbonafide AP-1activity,since it is composed of Fos- and Jun-related proteins.
Similar anti-Jun and anti-Fos antibody addition
experi-ments were performed, using extracts from CEF and ASV17-transformed CEF. However, since v-Jun is ex-pressed as a fusion protein with the retroviral gag gene product, we also included a polyclonal anti-Gag antiserum (5202) in this analysis. Addition ofanti-Gag serum had no effectontheactivityof theretardedcomplexinnormalCEF extracts (Fig. 2) but completely disrupted the novel com-plex of slower mobility specific to the ASV17-transformed cell extract (Fig. 2), indicating that this complex contains
p65vJun.
Thiscomplexwasalsosupershiftedby 948anti-Junantiserum andpartiallydisrupted by 388 anti-Fosantiserum, suggesting that the novel complex contains Fos-reactive proteins in addition top65v-Jun. As with the rat cell extracts, the more rapidly migrating complexes present in both ASV17-transformed and normal CEF extracts were partially disrupted by both 948 anti-Jun and 388 anti-Fos antisera (Fig. 2), confirming the presence of both Fos- and Jun-relatedproteins in these complexes.
The increase in AP-1 DNA-binding activity upon serum stimulationisbiphasicinnormal and
v-fos-transformed
cells.Theexpression ofc-fosistightlyregulated in cells.Growing cells have low but detectable levels of
p55cFos,
while in quiescent cells theproteinisvirtually undetectable (see Fig. SA). After serum stimulation, there is a rapid increase in c-fos mRNA followed by a parallel increase in p55c-Fos (compare Fig. SF with SA). Since the v-fos genesareunder retroviral long terminal repeatregulation and therefore areconstitutively expressed,wewished todetermine the varia-tion of AP-1DNA-binding activityinrelationtothegrowth
stateof the cells. To examine thegrowthresponseto serum of the 208F cell line and its Fos transformants, cells were seeded at various concentrations and their growth was monitored by cellcounts. The FBR cellsrequired the least serum to maintain growth, whereas FBJ- and MMV-trans-formed cells did not differ significantly from the parental 208F cells (data not shown). After the cells had ceased growing in 0.5% serum for 2 days, theywere shifted into medium containing 10% serum. Each of the cell lines
re-sponded byproliferating. To determine that the cellswere
entering into a quiescent state, they were pulsed with [3H]thymidine to measureDNAsynthesisatthe restrictive serum concentrations. All of the Fos-transformed cells showed decreased DNA synthesiswhenmaintained in low serumconcentrations. If the cellswereshifted backto 10% serum, theybegan to synthesizeDNA, asmeasuredbyan increase in
[3H]thymidine
incorporation(Fig. 3).Nuclear extracts were prepared from logarithmically growing, quiescent and serum-stimulated cells, and AP-1 DNA-bindinglevelsweredetermined. The datapresented in Fig. 1 indicate that similar AP-1 DNA-binding activity is present in each cell line during logarithmic growth. When quiescent,all cell linesdisplayadecrease inAP-1activityin comparisontothe levels detected inlogarithmically growing cells(Fig. 4).Inparticular,AP-1activityisbarelydetectable in the FBR
v-fos-transformed
cell line under these condi-tions. Upon stimulation with serum for 1 h, all cell lines exhibited an increase in AP-1 DNA-binding activity whenon November 9, 2019 by guest
http://jvi.asm.org/
-I L-
L-:: z ;
208F1
N/I NI
v
F1BJ
FB1R
CEF
-4, --Mwpa - <A
a -
."
'tL-&.U"
L ^
C
FF
NWV17
FIG. 4. Timecoursesshowingthepatternofseruminduction of
AP-1DNA-bindingactivity inratandchickenfibroblasts.
Prepara-tionsof nuclear extractswereperformed aspreviouslydescribed.
HeLacell extractwasused as apositive control for AP-1
DNA-binding activity. The gel shift detected in the HeLa lane differs
slightly from that of the rat and chicken fibroblast cell lines due
possiblytoachange in the ratio of theproteinsavailable forAP-1
DNAbinding.
compared with the levels observed in both growing and serum-starved cells (Fig. 4). This increase inactivity paral-lels denovosynthesis ofc-fos (seebelow). Asinthecaseof
growing cells, the induced levels of AP-1 DNA-binding activityareindistinguishable amongthecell lines.
ThepatternofAP-1inductionover alongerperiodoftime
wasexamined inanattempt to identifyadifference between
AP-1DNA-bindingactivity innormalandtransformedcells.
We measuredAP-1 activity across aserum induction time
course in each of the four rat fibroblast cell lines. Four
individual induction experiments were performed on each
cellline, andtheresultswereconsistent. Thelevel ofAP-1
activity following serum induction remained high at least
until 3 h after stimulation. At5 h, the AP-1 DNA-binding activity of 208F, MMV,andFBRcells decreasedto alevel
similar to that observed when the cells were starved of serum. Asecond peak in AP-1
DNA-binding
activity
wasseen at 7 h after serum induction in these cells and was sustained at leastuntil 9 h. The FBJ cell line differs in this response from the other rat fibroblast cell
lines;
a minor decrease in AP-1DNA-binding
activity
was evident at 7 h after seruminduction, withasecondincrease inactivity
at9 h.A similar experiment conducted with CEF also demon-strated a biphasic increase in AP-1
DNA-binding
activity
after serum stimulation,
although
inthese cells the second phaseofstimulation occurredat9 h(like
FBJ)
incomparison
to7 h in eachof the
majority
of therat fibroblast cell lines (Fig.4). Thepattern ofseruminductionin ASV17cellswasunique to each of the cell lines studied here. After serum
induction, AP-1 activity was constant, not
decreasing
at either 5 or7h.Expression of v-fos, MMV
c-fos,
andv-jun
is in excesscompared with that of their normal cellular counterparts. Previouslypublisheddata demonstrate that the viral
fos
and jun oncogenes areconstitutively
expressed.
Therefore,
weexpected that cells transformed with either of these
onco-geneswould showanincrease intheamountof
protein
ina cell available for AP-1 function.However,
we have shown that AP-1activity doesnotincrease incells transformedwith either the v-fos or the v-jun oncogene and we wished to determine any change in theavailability
of Fos and Jun protein in the cell lines studied.Immunoprecipitation
experiments
demonstrated thatp55c-Fos
cannotbedetected inquiescent
208F cells(Fig.
5A)
but is presentatverylowlevelsin
MMV,
FBJ,
and FBR cell lines(Fig. 5B,C, andD,respectively).
Serum stimulationfor 30 min increasedp55cFos levels inall cell lines(Fig. SA, B,
C, and D) andwas
particularly
striking
in MMV(Fig.
5B).
Both v-fos gene products are more abundant than their cellular counterpart, but FBR
p75-F0s
isconstitutively
ex-pressed (Fig.
5D),
whereas FBJp55V-Fs
isexpressed
in asimilar manner to
p55c-Fos
(Fig.
5C).
The observation thatp39c-Jun
andp55c-Fosarecoprecipitated
in each cell linewithanti-Fosantiserum388
(Fig. 5A, B, C,
andD)
and viceversa(Fig.
5A,
B,C,andD) provides
evidencefor c-Fos and c-Jun dimerization. p39c-Junisprecipitated
with the Fos anti-serumin the FBRcellline,
demonstrating
that FBRp75-Fos
iscapable ofdimerizationwith
p39c-Jun
(Fig.
SD).
Asimilar situation exists in the FBJ cell line.Immunoprecipitation
with anti-Jun antiserum 948 demonstrates that
p39c-Jun
can be detectedin eachgrowthcondition,
which suggests that it is synthesizedconstitutively(Fig.
5A,
B, C,
andD).
Western blotting
experiments,
using
nuclear extracts pre-pared fromlogarithmically
growing
CEF and ASV17-trans-formed CEF, indicate anapproximately
20-fold excess ofp65-Jun
overp39c-Jun
(Fig.
5E).
A similar difference in the level of expression of the twoproteins
was detected in quiescent and serum-inducedcells,
suggesting
that both proteins are constitutivelyexpressed
(results
notshown).
p39c-Jun
thus demonstrates aninduction mechanism whichis clearly responsive to serum, distinct from thatofp55cFos.
Northern blot analyses ofa serum induction time course
similartotheonediscussedaboverevealed thatthe expres-sion kinetics of the endogenous
c-fos
gene in the208F,
MMV, FBJ, and FBR cell lines are similar. That
is,
the mRNAsareonlydetectableat1hafterseruminduction(Fig.
5F). In contrast, the
c-fos
construct introduced into MMV cells isconstitutively
expressed,
butstillinducible,
reaching
a peak at 1 and 3 h after serum stimulation
(Fig.
SF).
Expression of the
v-fos
gene in the FBJ and FBR cell lineson November 9, 2019 by guest
http://jvi.asm.org/
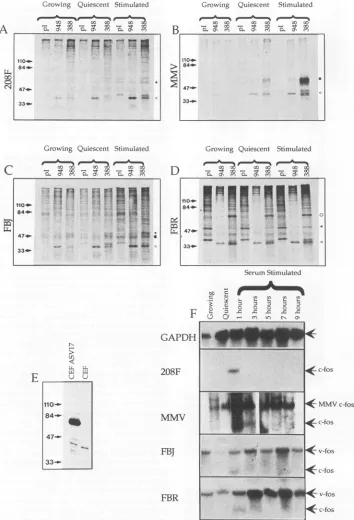
[image:5.612.76.287.75.476.2]5492 HAWKER ET AL.
Growinig Quiescenit Stimulated
0000 00 00 d0 a
- " 00 t0 00-eCZ 000
0. 0' CO 0E. 0' CO0 0' CO
Growing Quiescent Stimulated
00 00 0000 0
-.0'
,l-
C\ 0.0'C 0.F 00 - 00>
tif
84- 1Ms4*
47...
33_
-E
N
5;
L L.
VL
110-
84-.-*1
47.
33-_
Growinlg Quiescent Stimulated
At00a:-o # 0c0c 000'D
B
" m sm O
110_-84*- 0
$~~~~~~~~4a4
47- "
. -.>-.. jlLas
33*--Growinig Quiescent Stimulated
D
0.s0r
or
It00'D co,ON en CL0\ m -0 Com
SerumStimulated
[image:6.612.130.484.85.605.2]MMVc-fos
FIG. 5. Ananalysis offosand jun geneexpression. (A through D)Immunoprecipitations showingtheregulationand relativeabundance ofp55c-Fo, FBJp55W-FoS, and FBRp75'-F0. Cellswerelabelled for2 h and thenlysedinRIPAbuffer.Fos-andJun-reactiveproteinswere
sequestered bythe addition of antisera 388 and948,respectively,andcollectedonprotein A-Sepharosebeads. Proteinswere electrophoret-ically separatedon8% polyacrylamide gelsandautoradiographed.(E) Western blot showing therelativeabundanceof thep39c-Junandp65-J,n proteinsintheCEFandCEF ASV17cell lines. Nuclearextracts wereelectrophoretically separatedon an8%polyacrylamidegel,dry blotted
ontoSartoriousmembranes, and developed usingtheECLsystem(Amersham).(F) Northern blotanalysisshowing thepatternofexpression ofthec-fos and v-fosgenes over a9-h period inthe208F, MMV, FBJ,andFBRcelllines. TotalRNAprepared usingRNAzol(Promega)was electrophoretically separated on a 1% formaldehyde denaturing agarose gel and blotted onto a nylon membrane. Hybridization of random-primed p65murinec-foswasdonebystandardtechniques.Filterswerestripped and reprobed withaGAPDHfragmentas aloading control. Ineachcase,loadingwasequal, asexemplified by the reprobed 208F membrane
(top
row).Thefollowingsymbolsrepresentthe positions of the various immunoprecipitated proteins: 4,p55c-Fos;< p39c-Jun; U,MMVp55C OS;0,FBJp55-Fos; and 0,FBRp75v-Fos
A
0
~v."- -
Pn..-
SS 110_.84_.
413
47-33_p
...
I--84-HO': 5.i w .,
47. .
33- . i _ u ,
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
can also be described as inducible but continuously
ex-pressed,
although neither message is as abundant as that ofthe MMV
c-fos
construct (Fig.5F).DISCUSSION
Thedatawepresent proffer two conclusions. First, AP-1
activity,
asmeasured by standard DNA-binding procedures,doesnot increase upon transformation of 208F cells by the
fos
oncogene orof CEF by the jun oncogene. Thisphenom-enon is observed despite an increase in the Fos or Jun
oncoprotein
in the transformed cells. Second, there is abiphasic
increase in AP-1 activity following serum stimula-tion ofquiescent
208F cells and CEF. Transformation of 208F cells eitherby the v-fos oncogene or by overexpression of thec-fos
gene does not alter thispattern, even thoughfos gene expression is constitutive. In contrast, v-jun-trans-formedCEF donothave abiphasic pattern of AP-1 activity. From these observations it can be inferred that the level ofFos
protein
in the cell is not the sole rate-limiting factor inthe
regulation
of AP-1 activity.Thelackofanincrease in AP-1 activity infos-transformed cells was contrary to our expectations on the basis of the
following
evidence. Normally,p55'F
is less abundant incellswhencompared with p39C un;the kinetics of induction
of
p55cFos parallels
the kinetics of AP-1 activity followingserum stimulation; a concomitant increase in p55c-Fos
ex-pression
and AP-1activity isobserved upon transformationby
other oncogenes, e.g., sis and ras (19, 27), and in ras-transformed cells there is evidence that AP-1 activity isrequired
for morphological transformation (27). Takento-gether,
theseresultssuggestedthat when there is an increase in theamountofp55cFOsin acellthere is an increase in AP-1DNA
binding.
This is clearly not the case for fostransfor-mation and
perhaps
the MMV-transformed cells are more indicative of this point. Although there is a significant increase in the levels ofp55c-Fos compared with 208F cells, there is noproportional
increase in AP-1 activity in theasynchronously
growingcells. Following serum stimulation of the MMV-transformed cells there is a significant increasein
p55cFOs,
but the increase in AP-1 activity is not as greatand
only slightly
higher than that observed in theserum-stimulated 208F cells. It is also of note that the MMV cells
donot
display
the transformedmorphology characteristic ofthe
v-/os-transformed
cells. Thisindicates that an increase inp55c-Fos
alone, at least to a level observed in our MMV-transformed cells, is not sufficient to induce morphologic transformation. This observation further suggests that mu-tations in thev-fos
oncogenesmediate morphological trans-formation without affecting the levels of AP-1 activity.We have demonstrated that serum deprivation prevents
growth
of thefos-transformed
cells and that the levels ofAP-1
activity
dramatically decrease in the serum-starvedcells. Inthecaseof the
v-fos-transformed
cells, the level ofFos
oncoprotein
detected byimmunoprecipitationdoes notvary
significantly
among quiescent, growing, andserum-induced cells. Furthermore, p39c-Jun isdetectable under all three
growth
states and its coprecipitation by the anti-Fosserum indicates that Fos-Juncomplexes exist in all growth states evenwhen there is littledetectable AP-1 activity. The variation of AP-1 levels with time,following serum
stimula-tion,
further suggests that AP-1 activity is regulated byfactors other than the level of Fosprotein in the cell. The
finding
thatc-fos
mRNAisundetectableat 3 h after induc-tion andfosB
mRNA is lost 2 h later may result in Fosprotein
being
limitingfor AP-1 activity 5 hfollowing serumstimulation. However, this seems unlikely since the
fra-1
and
fra-2
genes areboth expressed between3 and 9 h afterserum induction and have been shown to be the dominant Fos family proteins in the AP-1 complex in logarithmically
growing
cells(31).
In addition, the level offos oncogeneexpression remains elevated at all times in the
fos-trans-formed celllines. Thereareseveralpossible explanationsfor the observedbiphasicnatureof AP-1activity,but
phosphor-ylation of the carboxy terminus of
p39-J"',
which is known to result in a down regulation of DNA-binding, is thesimplest.
The constant level of AP-1activity
seeninv-jun-transformed cells is consistent with this theory, because v-Jun protein is not phosphorylated on its C terminus. A second possibility is that a specific inhibitor, IP-1, which may be regulated by protein kinases, becomes active 5 h after serum stimulation, reducingthe abilityof AP-1 tobind DNA (5, 6). The constant
AP-1
activity in ASV17-trans-formed cells could be explained if IP-1 was unable to recognize and regulatep65".Jun
Mutational analysis offos and jun oncogenes indicates thatDNA-binding is required fortransformation. However, recent reports suggest that transcriptional activation of the v-fos (30, 36)and v-jun(20,21) oncogenes isnotsufficient for transformation, even though the changes induced by the v-fos and v-jun genes result in increased transactivation. Indeed, one report suggests an inverse correlation between transformation and transactivation in v-jun-transformed cells(21), although DNA binding, as reported here, does not relate totransactivation.
Fos has recently been demonstrated to interact with proteinsother than Jun to effect transformation(53),making the processoffos-andjun-inducedoncogenesis increasingly complex. Clearly, much work remains to be done before a true understanding is established, but certainly the many recent reports would suggest that there is more tofosandjun transformation than AP-1 DNA-binding activity alone.
ACKNOWLEDGMENTS
The authors thank Tom Curran for providing the rat fibroblastcell lines, 208F, MMV, FBJ, and FBR; Natalie Teich for the Trp-E-Fos plasmid; and Bob Eisenman for the 5202 anti-Gag antiserum. Figure SEwas kindly provided by Anna Kilbey. Thanks to John Wyke and Margaret Frame for critical reading of the manuscript.
This work was supported by the Cancer Research Campaignand A.P. is grateful for an EMBO long-term fellowship.
REFERENCES
1. Allegretto, E. A., T. Smeal, P. Angel, B. M. Spielgelmen, and M. Karin. 1990. DNA-binding activity of Jun is increased through its interaction with Fos. J. Cell. Biochem. 42:193-206. 2. Angel, P., I.Baumann, B. Stein, H.
Delius,
H. J. Rahmsdorf,andP. Herrlich. 1987. 12-O-tetradecanoyl-phorbol-13-acetate induc-tion of the human
collagenase
gene is mediated by an inducible enhancer element located in the 5' flanking region. Mol. Cell. Biol. 7:2256-2266.3. Angel, P.,K. Hattori, T. Smeal, and M. Karin. 1988. The jun proto-oncogene is positively autoregulated by its product,
Jun/
AP-1. Cell 55:875-885.4. Angel, P., M. Imagawa,R. Chui,B. Stein, R. J.
Imbra,
H. J. Rahmsdorf, C. Jonat, P.Herrlich, and M. Karin. 1987.Phorbol ester inducible genes containa common cis elementrecognised by a TPA-modulating transacting factor. Cell49:729-739. 5. Auwerx, J., and P. Sassone-Corsi. 1991. IP-1: a dominantinhibitor of Fos/Jun whose activity is modulated by phosphory-lation. Cell64:983-993.
6. Auwerx, J., and P. Sassone-Corsi. 1992. Regulation by IP-1: effect of signal transduction pathways andcell growth on AP-1 (Fos-Jun). Oncogene 7:2271-2280.
on November 9, 2019 by guest
http://jvi.asm.org/
7. Black, E. J., A. J. Street, and D. A. F. Gillespie. 1991. Protein phosphatase 2A reversesphosphorylation of cJun specified by the delta domain in vitro: correlation with oncogenic activity of vJun. Oncogene6:1949-1958.
8. Bohman, D., and R. Tjian. 1989. Biochemical analysis of transcriptional activation by Jun: differential activity of c- and v-Jun. Cell 59:709-717.
9. Bos, T., D.Bohmann, H. Tsuchie, R. Tjian, and P. K. Vogt. 1988. v-jun encodes a nuclear protein with enhancer binding properties of AP-1. Cell52:705-712.
10. Boyle, W. J., T. Smeal, L. H. K. Defize, P. Angel, J. R. Woodgett,M.Karin, andT.Hunter.1991.Activation of protein kinase C decreasesphosphorylation of cJun at sites that nega-tivelyregulate its DNA-binding activity. Cell64:573-584. 11. Cavalieri, F., T. Ruscio, R. Tinoco, S. Benedict, C. Davis, and
P. K.Vogt. 1985. Isolation of three newavian-sarcoma virus-es-ASV-9, ASV-17 and ASV-25. Virology 143:680-683. 12. Chui, R., W. J. Boyle, J. Meek, T. Smeal, T. Hunter, and M.
Karin. 1988. The cFos protein interacts with cJun/AP-1 to stimulate transcription of AP-1 responsive genes. Cell 54:541-552.
13. Cohen, D. R., and T. Curran. 1988.fra-l: a serum-inducible, cellular immediate-early gene that encodes a Fos-related anti-gen. Mol. Cell. Biol. 8:2063-2069.
14. Dobrzanski, P., T. Noguchi, K.Kovary,C. A. Rizzo, P.S. Lazo, and R.Bravo. 1991. Both products of the fosB gene, FosB and its short form, FosB/SF,are transcriptional activators in fibro-blasts. Mol. Cell. Biol. 11:5470-5478.
15. Eisenman, R. N., W. S. Mason, and M. Linial. 1980. Synthesis andprocessing of polymerase proteins of wild-type and mutant avian retroviruses. J. Virol.36:62-78.
16. Finkel, M. P., C. A.Reilly,and B.0.Biskis. 1975. Viral etiology of bone cancer. Front. Radiat. Ther. Oncol. 10:28-39. 17. Frame, M. C., N. M. Wilkie, A. J. Darling, A. Chudleigh, A.
Pintzas, J. C.Lang, and D. A. F. Gillespie. 1991. Regulation of AP-1/DNA complex in vitro. Oncogene 6:205-209.
18. Franza, B. R., F. J. Rauscher, S. F. Josephs, and T. Curran. 1988. The Fos complex and Fos-related antigens recognise sequence elements that contain AP-1 binding sites. Science 239:1150-1153.
19. Gilman, M. Z. 1988. The c-fos serum response element re-spondstoprotein kinase C-dependent and independent signals butnot tocAMP. Genes Dev. 2:394-402.
20. Halazonetis, T. D., K. Georgopoulos, M. E. Greenberg, and P. Leder. 1988. cJundimerises with itself and with cFos, forming complexes of different DNA-binding affinities. Cell 55:917-924. 21. Hard, M., and P. K. Vogt. 1992. Oncogenic transformation by Jun: role oftransactivation andhomodimerisation. Cell Growth Differ.3:899-908.
22. Havarstein,L.S., I. M. Morgan,W.-Y. Wong, and P. K. Vogt. 1992. Mutations in the Jun delta region suggest an inverse correlation between transformation and transcriptional activa-tion. Proc. Natl. Acad. Sci. USA 89:618-622.
23. Hirai, S.-I.,B. Bourachot, and M. Yaniv. 1990. Both Jun and Fos contribute to transcription activation by the heterodimer. Oncogene 5:39-46.
24. Hirai, S.-I., R. P. Ryseck, R. Bravo, and M. Yaniv. 1989. Characterisation of junD: a new member of the jun proto-oncogene family. EMBO J. 8:1433-1439.
25. Hughes, S. H., J. J. Greenhouse, C. J. Petropoulos, and P. Sutrave. 1987.Adapterplasmidssimplify the insertion of foreign DNA into helper-independent retroviral vectors. J. Virol. 61: 3004-3012.
26. Imbra,R.J., and M.Karin. 1987. Metallothionine gene expres-sion is regulated by serum response factors and activators of proteinkinaseC. Mol.Cell. Biol.7:1358-1468.
27. Imler,J. L., C.Schatz, C. Wasylyk, B.Chatton, and B.Wasylyk. 1988. A Harvey-ras responsive transcription element is also responsive to atumour-promoter and to serum. Nature (Lon-don)332:275-278.
28. Jamal,S., andE.Ziff. 1990.Transactivation of c-fos and
P-actin
genes by rafas a step in early response to transmembrane signals.Nature (London) 334:463-466.29. Jenuwein, T., and R.Muller. 1987. Structure-function analysis of Fosprotein: a single amino acid change activates the immor-talising potential ofv-fos. Cell 48:647-657.
30. Joos, K., and R. Muller. 1992. Analysis of temperature sensitive functions of Fos: lack of acorrelationbetween transformation and TRE-dependenttrans-activation. Oncogene 7:1933-1939. 31. Kovary,K., and R. Bravo. 1992.Existence of differentFos/Jun
complexes during theGO-to-G1transition and duringexponential growth in mouse fibroblasts: differential role of Fos proteins. Mol.Cell. Biol.12:5015-5023.
32. Landschulz, W. H., P. F. Johnson, and S. L. McKnight. 1988. The leucine zipper: ahypothetical structurecommon to a new class of DNA-binding proteins. Science 240:1759-1764. 33. Lee,W., A.Haslinger, M. Karin, and R. Tjian. 1987.Activation
of transcriptionby two factors that bind promoter andenhancer sequencesof the human metallothionine gene and SV40. Nature (London)325:368-372.
34. Uoyd, A., N. Yancheva, and B.Wasylyk 1991.Transformation suppressor activity of a Jun transcription factor lacking its activation domain.Nature(London) 352:635-638.
35. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem. 193:265-275.
36. Lucibello, F. C., M. Neuberg, T. Jenuwein, and R. Muller. 1991. Multiple regions of vFos protein involved in the activation of AP-1-dependent transcription: is transactivation crucial for transformation? NewBiol. 3:671-677.
37. Maki, Y., T. J. Bos, C. Davis, M. Starbuck, and P. K. Vogt. 1987. Avian sarcoma virus 17 carries the jun oncogene. Proc. Natl.Acad. Sci. USA84:2848-2852.
38. Miller, A. D., T. Curran, and I. M. Venna. 1984. cFos protein caninducecellulartransformation: a novel mechanism of acti-vation of a cellular oncogene. Cell36:51-60.
39. Morgan, I. M., L. J. Ransone, T. J. Bos, I. M. Verma, and P. K. Vogt. 1992. Transformation by Jun: requirement for leucine zipper, basic region and enhancement by Fos. Oncogene 7:1119-1125.
40. Neuberg, M., M.Schuermann, and R. Muller. 1991. Mutagene-sis of the DNA contact site in Fos protein: compatibility with the scissors grip model and requirement for transformation. Oncogene6:1325-1333.
41. Nishina, H., H. Sato, T. Suzuki, M. Sato, and H. Iba. 1990. Isolationand characterisation of ra-2, an additional member of the fosgenefamily. Proc. Natl. Acad. Sci. USA 87:3619-3623. 42. Ofir, R., V. J. Dwarki, D. Rashid, and I. M. Verma. 1990. Phosphorylationof the C-terminus of Fos protein is required for transcriptional transrepression of the c-fos promoter. Nature (London)348:80-82.
43. Pulverer, B. J., J. M.Kyriakis, J. Avruch, E. Nikolakaki, and J. R. Woodgett. 1991. Phosphorylation of cJun mediated by MAPkinases. Nature (London)353:670-674.
44. Ransone, L. J., J. Visvader, P. Sassone-Corsi, and I. M. Verma. 1989. Fos-Jun interaction: mutational analysis of the leucine zipperdomain ofboth proteins. Genes Dev.3:770-781. 45. Rauscher, F., D. Cohen, T. Curran, T. Bos, P. Vogt, D.
Bohmann, R. Tjian, and B. Franza. 1988. Fos associatedprotein p39 is the productof thejun proto-oncogene.Science 240:1010-1016.
46. Riva, P., and L. Larizza. 1992.Expression of c-sis and c-fos in human meningiomas and neurinomas. Int. J. Cancer 51:394-402.
47. Ryder, K., L. F.Lau,and D.Nathans. 1988. A geneactivated by growth factors is related to the oncogene v-jun. Proc. Natl. Acad.Sci.USA85:1487-1491.
48. Schontal, A., P. Herrlich, H. J. Rahmsdorf, and H. Ponta. 1988. Requirement for fos gene expression in the transcriptional activation ofcollagenase byother oncogenes and phorbol es-ters.Cell 54:325-334.
49. Smeal, T., B. Binetruy, D. A. Mercola, M. Birrer, and M. Karin. 1992. Oncogenesis and transcriptional co-operation with Ha-Ras requires phosphorylation of cJun on 63 and 73. Nature (London)354:494-496.
50. van Beveren, C., F. van Straatan, T. Curran, R. Muller, and
on November 9, 2019 by guest
http://jvi.asm.org/
I. M. Verma. 1983.Analysis of FBJ-MuSV provirus and c-fos
(mouse)genereveals that viral and cellular fos geneproducts
have different carboxy termini. Cell 32:1241-1255.
51. Vogt, P. K., T. J. Bos, and R. F. Doolittle. 1987. Homology between the DNA-binding domain of the GCN4 regulatory protein ofyeast and the carboxy terminal region ofa protein
coded for by the oncogenejun. Proc. Natl. Acad. Sci. USA
84:3316-3319.
52. Vosatka, R. J., A. Hermanowski-Vosatka, R. Metz, and E. B. Ziff. 1989. Dynamic interactions of cFos protein in serum
stimulated 3T3 cells. J. Cell. Physiol. 138:493-502.
53. Wick, M., F. C. Lucibello, and R. Muller.1992. Inhibition offos andrasinduced mutations with structural alterations in func-tionally different domains. Oncogene 7:859-867.
54. Zarbl, H., J. Latrelle, and P. Jolicoeur. 1987. Revertants of v-fos transformed fibroblasts have mutations in cellulargenes
essen-tial for transformation by otheroncogenes. Cell 51:357-369. 55. Zerial, M., L. Toschi, R.-P. Ryseck, M.Schuermann, R. Muller,
and R. Bravo. 1989. The product of a novel growth factor
activatedgene,fosB, interacts with Jun proteins enhancingtheir
DNAbinding activity. EMBO J. 8:805-813.
![FIG. 3.monitored[3H]thymidine. High-density thymidine uptake assays in 0.5% FBS. 208F, MMV, FBJ, and FBR cells were seeded in 0.5% FBS and pulsed with After the cells had become quiescent, the cultures were adjusted to 10% FBS and the incorporation of [3H]thymidine was for a further 24 h.](https://thumb-us.123doks.com/thumbv2/123dok_us/1299102.82984/4.612.78.534.63.306/monitored-thymidine-thymidine-quiescent-cultures-adjusted-incorporation-thymidine.webp)