JOURNAL OF VIROLOGY,JUlY1994,p.4377-4383 Vol. 68,No.7 0022-538X/94/$04.00+0
CopyrightC 1994,American Society forMicrobiology
Excision
of DNA Fragments
Corresponding to the Unit-Length
a
Sequence of Herpes Simplex Virus Type
1
and
Terminus
Variation Predominate
on
One
Side of the Excised Fragment
KENICHIUMENE*
DepartmentofVirology, Faculty of Medicine, Kyushu University 60, Fukuoka
812,
Japan Received 30November 1993/Accepted 18March 1994DNAfragmentscorresponding to the unit-length a sequence of herpes simplex virus
type
1 (HSV-1) were identified in HSV-1 DNA preparationsextracted by the method of Hirt. The DNA fragments weremolecularly
cloned, and nucleotide sequences were determined. Most termini of the fragments were at sites on DR1 corresponding to the termini of linear HSV-1 DNA generated by the cleavage-packagingsystem. In one-step growthexperiments, DNA fragments of the unit-length a sequence appeared
simultaneously
with the termini of linear HSV-1 DNAs produced by cleavage of circular and concatemeric DNAs. Therefore, excision of the unit-length asequence appeared closely related to thecleavage-packaging system. Termini of the excised DNA fragments of the variant a sequence with two DR2 arrays varied on theL-componentside,whileterminionthe S-component side were at the site on DRI corresponding to the authentic cleavage site. It is thus assumed that the cleavage-packaging system functionsadequately
on the DR1 second distal from the S component, and cleavages of other DRI are rare and less accurate. If this notion is tenable, then most termini on the S-component side of the excised DNA fragments are derived from the second DR1properly
cleaved and should be constant,whiletermini on the L-component side are from regions on and around the DR1 third distal from theS componentandmaybevariable.Cleavage of DRI islikely
tobeaffected by the topological relationship with theS component.The linear 155-kb genome of herpes simplex virus type 1 (HSV-1) iscomposed of two covalently linked components,L and S (19). The short sequence a is repeated directly at both endsof thegenomeandis present in inverse orientation at the L-Sjunction (Fig. la) (7, 15). One to several copies of the a sequence are present attheendofthe L component and at the L-S junction, but only one copy is present at the end of the S component. The a sequence encodes several cis-acting sites involved in (i)cleavage of the unit-length HSV-1 DNA from concatemeric formsgeneratedby rolling-circle replication and encapsidation of the excised molecules (cleavage-packaging system) (4, 8, 9, 16, 17, 21, 24, 35), (ii) thecircularization of viral DNA afterinfection(16),(iii) the L-S inversion (4, 15, 16, 20, 24, 35), and (iv) the expression of an mRNA extending fromthea sequence (5).
The a sequence contains unique (U) and directlyrepeated (DR) sequenceelements ofDR1,Ub, DR2n, a DR4 stretch, and Uc and is flankedbyDR1(Fig. lb,and2a) (7, 14,30,35). The lengths of a sequences ofvarious HSV-1 strains range from 220 to 570 bp and depend mainly on the number of reiterationsof DR2(25,30, 31, 33,34). Tandemly reiterateda sequences sharethe interveningDR1. Processing of the con-catemericform intounit-lengthviral DNAinvolves asymmet-ricsite-specific cleavage ofDR1sharedbythetwoadjacenta sequences
(authentic
cleavage by thecleavage-packaging
sys-tem) (16).
The pacl andpac2sequences,whichareregions
of stronghomology among theasequencesof diverseherpesvirus genomes, are likely candidates for signals that direct the site-specific cleavage(6,
8, 17,35).Thecleavageandpackaging
of viralDNA aretightlycoupledandmediatedbythe cleavage-packagingsystem(9, 16, 24,
35).
HSV-1 may have several recombination systems
(1,
2, 11,*Corresponding author.Phone:92-641-1151. Fax: 92-632-6402.
26-29,
32, 36, 37). The region spanning the L-S junction is inherentlyrecombinogenic, and thea sequencecouldbeahot spot for the recombination (3, 10, 15, 20, 21). The double-strand-break repair model for recombination has been pro-posed, and this double-strand break is assumed to act as anefficient initiator of recombination (12, 22). The free DNA ends generated by the cleavage-packaging system of HSV-1 possiblystimulate theinitiation ofrecombination,although the recombination does not invariably occur by cleavage and ligation at a single specific site (20). Virus-induced nuclear endonuclease, which cleaves the DR2 array and generates strand breaks necessary for the recombination, wasreported (39). Unusual DNAfragments derived from thea sequence, terminiofwhichweresituated in defined regions assumed to be recombinogenic, were identified in a previous study (31). Thus,itseemsclear that thestrand break is indeedassociated withrecombination.
Ifaregion onthea sequence is
preferentially
broken,then DNA fragments linked to the breakage should be more abundant than other DNAfragments,andanovel DNAband maybedetectableon agel.Inthisstudy,Isearched foraDNA band composed of DNA fragments derived from the a se-quence. Anovel DNAband, composedof excisedunit-length
asequence,appearedon anacrylamide
gelafterelectrophore-sis of the HSV-1 DNA extracted
by
the method of Hirt. Termini of the excisedDNAsofthestandardasequencewere at the authentic cleavage site on DR1 as a result of thecleavage-packaging
system. However, termini of the excised DNAsof variantasequence withtwoDR2 arrays variedonthe L-componentside,while theterminiontheS-component
side were attheauthenticcleavagesiteonDR1. Theseobservationsare interpreted to mean that the
cleavage-packaging
system functionsadequately
on the DR1 second distal from the S component,andcleavagesof otherDR1elementsare rareand 4377on November 9, 2019 by guest
http://jvi.asm.org/
4 L ,~0-*. s _
TRL U1 ~~~~~~IRLIRSU,TRi
() TRL UL
a a a
(b)
Z 1 5 tSinal Apal Eco471
I
-I .i
prepared from hybrid plasmid pUK340, containing the Dral fragment corresponding to unit-length a sequence of HSV-1 clone TW14at theSmaI site ofpUC18 (30,31).
DNA sequencing. A restriction fragment from the hybrid plasmid was subcloned into both M13mplO and M13mpll. Sequences were determined by using the BcaBest dideoxy sequencing kit (Takara Shuzo Co., Kyoto, Japan) (31). The BcaBest DNA polymerase was obtained from thermophile Bacillus cardotenax and functions bestat65 to 75°C.
2B7
(c) Rd (DR2)5DR4n2 DRI
Cc)
ReCtSinai I 256 177
Apal I 237 170
ECO471 _ 7 287 138
FIG. 1. Mapsof HSV-1 DNA.(a) Structure of the HSV-1genome arranged in theprototypeorientation (19).HSV-1 DNAconsists of two covalently linked components, L and S, that constitute 82 and 18%,respectively,of thegenome.Eachcomponentconsists of unique sequences (ULandUs) bracketed by invertedrepeat sequences(TRL, IRL,IRS, andTRS). Ashortsequence, a,is repeated directlyatthe termini of the HSV-1 genome and is also present in the inverse orientationattheL-Sjunction. Onetoseveralcopiesoftheasequence arepresent atthe end of theL component andatthe L-Sjunction, but onlyonecopyispresent attheend oftheScomponent (7, 8, 15, 16, 35). (bandc)Schematicrepresentations oftheasequencesofHSV-1 clones GN28 (b) and GN29 (c). The a sequence of GN28 is the standard one.Theasequenceof GN29 isavariantonewithtwoDR2 arrays,generated by the recombination betweensitesonUb andUc (31).Thenovel recombinationsiteon theasequenceof GN29(Rec) is indicated byaclosedcircle. The authenticcleavage siteonDR1due tothecleavage-packagingsystemisindicatedbyaverticalarrow.SmaI, ApaI,andEco47Irestriction endonuclease cleavage sitesare shown. DNAfragments, whicharegeneratedbydigestion of the unit-lengtha
sequenceas aresultof authenticcleavage witheachof these restriction endonucleasesandaredetectable inSouthern hybridization analyses, areindicatedby double lines. Lengths of these DNAfragments are shown inbasepairs.
lessaccurate (16). Cleavage of DR1 islikelytobeaffected by thetopological relationshipwith the S component.
MATERIALSAND METHODS
Viruses and cells. HSV-1 strains GN28 and GN29 were singleclonal isolates of clinical strain K52(31). GN29 had the varianta sequencewith two DR2 arrays, and GN28 had the standardasequence(Fig.2).HSV-1 clinical strains K41, K56, K85, and Ty98 were as described elsewhere (34). HSV-1 was propagatedonVero cells inEagleminimum essential medium with2% fetal bovine serum (26).
ExtractionofHSV-1DNA by the method of Hirt. A Vero cell monolayer infected with an HSV-1 stock was collected by low-speed centrifugation. The pellet was lysed with a solution containing0.01MTris-HCl(pH 8.0), 0.01 MEDTA,and 0.6% sodiumdodecyl sulfate. NaCl was added to a final concentra-tion of 1.0 M, and the lysate was maintained overnight at 4°C. The supernatant was separated by centrifugation and extracted asdescribed previously (18, 26).
Southern hybridization. Southern hybridization was carried outon aBiodyneBtransfer membrane (Pall Ultrafine Corp.) as described previously (25). The 0.175-kb
SmaI
DNA frag-ment used as a probe for detection of a sequences wasRESULTS
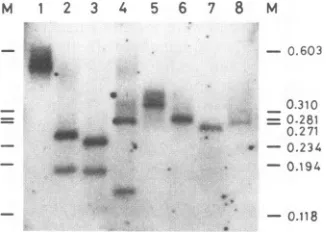
Detection of DNA fragments with the length of unit-lengtha sequence in the HSV-1 DNA preparations extracted by the method of Hirt. HSV-1 clone GN29, with the variant a sequence, was previously isolated (31) (Fig. lc and 2b). Un-usual DNAfragments containing parts of the a sequencewere present in DNA preparations derived mainly from viral nu-cleocapsids of GN29 (31). The unusual DNAfragments varied in length, and the presence of predominant unusual DNA fragments wasnotapparent inprevious work. In thisstudy, I searched for apredominant novel fragment in DNA prepara-tions of GN29 extractedbythe method of Hirt. The presence of a DNA band of 0.47 kb, corresponding to the length of unit-lengthasequence ofGN29,became evidentbySouthern hybridizationanalysis(Fig. 3, lane 1).
HSV-1 clone GN28 has the standard a-sequence structure (Fig. lb and 2a) (31). Two strains of GN28 and GN29 are apparently thesameexcept for thestructureof theasequence; both GN28 and GN29werederived from HSV-1 clinical strain K52. The DNA band of 0.33kb,correspondingtothelengthof unit-length a sequence of GN28, was detected in the DNA preparation of GN28 extractedby the method of Hirt (Fig. 3, lane 2). Therefore, generation of DNA fragments with the length of unit-length a sequence was notdue to DNA rear-rangementsuch as waspresent onthea sequence of GN29.
DNApreparations extractedbythemethod of Hirt of other HSV-1 strains of K41 (withasequenceof 0.25kb), K56(with
asequenceof 0.37kb),Ty98(withasequenceof 0.46 kbasthe major band and 0.38 kbasthe minorband), and K85 (witha sequence of 0.48 kb) were similarly analyzed by Southern hybridization(Fig. 3, lanes 3 to6) (34). DNAfragmentswith length ofunit-length a sequence of each HSV-1 strain were detected. Thus, the generation of DNAs with the length of unit-lengtha sequenceis common toall HSV-1strains exam-inedsofar andnot restricted to aparticular strain.
Structures oftheexcisedDNAfragmentswith thelengthof unit-length a sequence. To analyze structures of the DNA
fragments with the length of unit-length a sequence, DNA preparations of GN28 and GN29 extracted by the method of Hirtwere treated with the Klenow fragment and electropho-resedina5%acrylamide gel. Regions of the gel corresponding toDNAfragmentsof 0.3to0.4kb of GN28 and those of 0.4to 0.55 kb of GN29 were cut out. DNAs were extracted from the gels and cloned into the SmaI site of pUC18. Seven hybrid plasmids containing DNAs from GN28 (pUK365 series) and 11 hybrid plasmids containing DNAs from GN29 (pUK364 series) were constructed. Nucleotide sequences of the insert DNAs of these hybrid plasmids were determined and are summarized in Table 1.
The genomic termini of HSV-1 are generated by a single-base-pairstaggeredcleavage between nucleotides (nt) 18 and 20 of DR1 with a 3' single-base extension, leaving 1.5 bp of DR1at the S terminus and the remaining 18.5 bp of DR1 at theLterminus(authentic cleavage by the cleavage-packaging system) (16). Treatment of these termini with the Klenow
r
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.65.304.57.266.2]EXCISION OF UNIT-LENGTH a SEQUENCE OF HSV-1 4379
(a) GN28
1 30 60 90
CCGCGGGGGGCCCGGGCTGCCCGCCGCCGCGCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGGCCCGTTTTTGG
DR1 SmaI Ub DraI SmaI ApaI
ApaI
98 253 270 300 312
GGTCTGGG (CGCTCCTCCCC),4CGCCTTTTTCGGCCCCGCCCCCCACGCCCGCCGCGCGCGCGCACGCCGCCCGGACCGCCGCCCGCC
DR2 DR4n2 Uc Eco47I
330 338 TTTTTTGCGCGCGCGCACGC
(b) GN29
-31 -1
(Inverted repeat of L component) AGTGCTTGCCTGTCTAACTCGCTAGTCTCGG IR2
1 30 60 90
CCGCGGGGGGCCCGGGCTGCCCGCCGCCGCGCTTTAAAGGGCCGCGCGCGACCCCCGGGGGGTGTGTTTCGGGGGGGGGCCCGTTTTTGG
DR1 SmaI Ub DraI Smal ApaI
ApaI
98 253 2
GGTCTGGG (CGCTCCTCCCC),4CGCCTTTTTCGGCCCC
DR2 DR4n2
330 335 391
CGTTTTTGGGGTCTGGG (CGCTCCTCCCC)5,CGCCTTTT
DR2 DR4n2
450 476
CCGCCCGCCTTTTTTGCGCGCGCGCACGC
270 300 312
-GCCCCCCACGCCCGCCGCGCGCGCGCACGCCGCCCGGACCGCCCGGGGCC Uc
420
Eco47I Rec SmaIApaI rTCGGCCCCGCCCCCCACGCCCGCCGCGCGCGCGCACGCCGCCCCGGACCG
Uc Eco47I
FIG. 2. Nucleotide sequencesofasequencesof GN28 (a) and GN29(b), as determined in an earlier study (31). Nucleotide numbers start at theleftend of theleft DR1 andterminateatthe right end of Uc. The left end of each componentof theasequenceis indicated.DR1is underlined. Thecleavage sites of restrictionendonucleasesApaI,DraI,Eco47I, andSmaI areindicated.Thecytosine residue at nt 312,diagnosticof novel recombination of the a sequence of GN29, is underlined (Rec). Nucleotide sequences of major inverted repeats of the L component (RL) adjoining theasequence arealso showninpanelb andnumbered -1 to-31 (31). IR2 definedin strain Fis underlined(14).
fragment removesthe 3'extensiontomakeaflat end(at nt 18 and 20). Six of seven cloned DNAs from GN28 (pUK365 series)werethesameandcorrespondedtounitlength of thea sequence generated by the authentic cleavage of two DR1 elements encompassing one a sequence by the cleavage-packaging system(Table 1). Termini on the S-component side (right termini) of insert DNAs from GN29 (pUK364 series) were at the authentic cleavage site except in two cases, pUK364-9 andpUK364-14.TerminiontheL-component side (left termini)ofplasmidsofpUK364 seriesweremostlyonand around the left DR1 andwerevaried(Table 1).
To analyze the DNA fragments with the length of unit-length a sequence from each of GN28 and GN29, DNAs recovered from each region of the gel corresponding to the unit-length a sequence were cleaved with restriction endo-nucleasesSmaI, ApaI, andEco47I andanalyzed by Southern hybridization (Fig. 4). DNA fragments expected tobe gener-ated by digestion of the authentically cleaved unit-length
a sequence with each restriction endonuclease are shown in Fig. lb and c. Lengths of DNA fragments detected in the Southern hybridization analyses shown in
Fig.
4 corre-spondedtothoseexpected,asshown inFig.
lb andc.Thus,
the majority of the excised DNAfragments
with thelength
of unit-length a sequence were assumed to begenerated by
cleavageattheauthenticcleavagesiteonDR1or atothersites onand aroundDR1,in accordance with thestructures shown inTable 1.
Analyses of the appearance of excised unit-length a
se-quenceinsingle-step growthofGN28. To
analyze
the appear-ance of the unit-lengtha sequence, Vero cellswere infected with GN28at amultiplicityof 5 PFU per cell and harvested atvarious times upto16h
postadsorption
(Fig. 5).
Asingle-step
growthcurve wasconstructed(Fig.
5c).
HSV-1 DNAsfromtheM 1 2 3 4 5 6 M
(a)
-1.3 53 - 1.078
-0.872
_,x 0
- 0.603
La do
_.
0.310- ~~~~~~~=0.281
- 4 ~~~~~0.271 - ~~~~~~~~-0.2 34
-0.194
(b)
- 0.872
- 0.603 sop.
_ 0.310
=0.281
0.271
- 0.234
-0.194
FIG. 3. Southern hybridization profiles ofasequences of HSV-1 DNAsextractedbythe method of Hirt. HSV-1 DNAs extractedbythe method of Hirt were undigested (a) or digested with Dral (b),
electrophoresed in a 5% acrylamide gel, transferred to a nylon
membrane,andhybridizedwitha32P-labeled0.175-kbSmaIfragment ofpUK340(30).Lanes: 1,GN29; 2,GN28; 3, K41;4,K56; 5,Ty98; 6, K85(34).OneDralsiteis presentontheasequence,andapairofDral fragmentscorrespondingtothea sequencearegenerated(b) (7, 15,
16,30,31,35).One is thefragmentofunit-lengthasequence, andthe otheris shorterby16.5 bpas aresult ofauthenticcleavageonDR1.
Lane Misamarker mixtureofHaeIIIdigestsof
4XX174
phageDNA (25).Sizes offragmentsareshown in kilobasepairs.VOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.114.500.89.334.2] [image:3.612.364.509.429.624.2]TABLE 1. StructuresofclonedDNAfragmentscorrespondingtotheunit-lengthasequencepresentin the HSV-1 DNApreparations extractedby the method of Hirt
HSV-1 Lengthof
clone Hybrid plasmid insertDNA StructureofinsertDNAM
(bp)
GN28 20 338 18
pUK365-1, -2,-3,-6, -7,and -8 337 CCCGCCGCCGCGCTTTAAAG. . --GCCCGCGGGGGGCCCGGGCT
Ub DR1
4 20 338 18
pUK365-9 353 CGGGGGGCCCGGGCTGCCCG. ...GCCCGCGGGGGGCCCGGGCT
Ub DR1
GN29 20 476 18
pUK364-13and -17 475 CCCGCCGCCGCGCTTTAAAG.. . -GCCCGCGGGGGGCCCGGGCT
Ub DR1
21 476 18
pUK364-10and -15 474 CCGCCGCCGCGCTTTAAAGG. ... .GCCCGCGGGGGGCCCGGGCT
Ub DR1
18 476 18
pUK364-3,-4,and -5 477 TGCCCGCCGCCGCGCTTTAA. .-GCCCGCGGGGGGCCCGGGCT
Ub DRI
474 1 391 402
pUK364-9 405 CGCCCGCGGGGGGCCCGGGC - (CGCTCCTCCCC)5CGCCTTTTTCGG
DR1 DR2 DR4n2
451 476 20
pUK364-14 518 CCCGCCTTTTTTGCGCGCGCGCACGCCCGCGGGGGGCCCGGGCTGCCCG
DR1 Ub
476 16
.. ..ACGCCCGCGGGGGGCCCGGG
DR1
445 476 16
pUK364-2 526 CCGCCGCCCGCCTTTTTTGCGCGCGCGCACGCCCGCGGGGGGCCCGGG
DR1
20 476 18
CTGCCCG....GCCCGCGGGGGGCCCGGGCT
Ub DR1
-14 -1 6 476 18
pUK364-1 508 CTCGCTAGTCTCGGCCGCGG....GCCCGCGGGGGGCCCGGGCT
DR1 DR1
a Nucleotidenumbers are thoseofeachasequenceofGN28andGN29 andthemajor invertedrepeatof theLcomponentofGN29,asshowninFig.2(31).DR1
is underlined. Nucleotides 18and 20attermini of the insert DNA,whichcorrespondtotheflat ends of the clonedDNAfragmentderived fromauthenticallycleaved terminias aresultofthecleavage-packagingsystem andweregenerated byremoval of the 3' extensionat nt19as aresult of thetreatmentwith the Klenowfragment,
areunderlined(16).
infected cells harvested at various times were extracted by the method of Hirt and analyzed by Southern hybridization (Fig.5a and b). OneDral site is present on thea sequence, andapair ofDral fragments correspondingtothe a sequence were expected to be present when the linear HSV-1 DNAs were analyzed (7, 14-16, 30). One is the fragment of unit-length a sequence, and the other is shorter by 16.5 bp as a result of authentic cleavage on DR1. Linear HSV-1 DNAs packaged withinthe viralparticle arecircularized after infec-tion, and then the shorter Dral fragment generated by the authenticcleavage disappearsbecause of the absence of linear HSV-1 DNA termini as a result of circularization of linear DNAs. For up to 4 h postadsorption, one DraI fragment corresponding to the unit-length a sequence was detected; the shorteronewas notevident (Fig.Sb).TwoDraI fragments, including the shorter one due to authentic cleavage, were detected at and after 6 h postadsorption. The excised unit-length a sequence was first detected at 6 h postadsorption (Fig.Sa).The simultaneous appearance of both HSV-1 linear DNA termini and the excised unit-length a sequence is con-sistent with the hypothesis that production of the excised unit-length a sequences depends mainly on the cleavage-packagingsystem.
M 1 2 3 4 5 6 7 8 M
-U4.
-~~~~~~~~~~~i~~~i
- 4IkAM1
- 0.603
0.310
alb _.l.=0.281
-N* 0.271
* -0.234
-0.194
-0.118
FIG. 4. Southern hybridization profilesof DNA fragments corre-spondingtounit-length a sequence. HSV-1 DNAs of GN28 and GN29 extracted by the method of Hirt were electrophoresed in a 5% acrylamide gel. DNAs wererecovered from regions of the gel corre-spondingto0.3 to0.4 kbof GN28 and 0.4 to 0.55 kb of GN29. The recovered DNAs of GN29(lanes1to4) and GN28 (lanes 5 to 8) were electrophoresedina5%acrylamide gel after no digestion (lanes 1 and 5)ordigestion with SmaI (lanes2and6),ApaI (lanes3 and7), and Eco47I (lanes 4 and 8). The DNAs were transferred to a nylon membrane andhybridizedwith a32P-labeled0.175-kbSmaI fragment ofpUK340 (30). Lane M is a markermixture ofHaeIII digests of fX174 phage DNA (25). Sizes offragments are shown in kilobase pairs.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.360.523.500.616.2]EXCISION OF UNIT-LENGTH a SEQUENCE OF HSV-1 4381
(a) =
1 2 3 4 56 78
(b)
M
_1.353 -1.078
=0.281
-0.234
I.194
-0.603
-VW
(c)
LL
a-0
6
-5
-4
_0.310 =0.281
0.271
-0.234
-0.194
I I IS I
0 2 4 6 B 10 1216
Hours post-adsorption
FIG. 5. Detection of the excised unit-length a sequence in the single-step growth ofGN28. Vero cellswereinfected with GN28 ata
multiplicityof5PFUpercell. After adsorption for 2 hat370C, the cells
were washed three times and overlaid with Eagle minimum essential
medium containing 2% fetal bovine serum. Incubation at 370Cwas
continued until the infected cellswere harvestedat 0 h (lane 1), 2 h
(lane2), 4 h (lane3), 6 h (lane 4), 8 h (lane 5), 10 h (lane 6), 12 h (lane
7), and 16h (lane 8)postadsorption. HSV-1 DNAswereextractedby
the method of Hirt, digested with DraI (b) or undigested (a), and
electrophoresed ina5% acrylamide gel.The DNAsweretransferred
toanylonmembrane and hybridized witha32P-labeled0.175-kbSmaI
fragment of pUK340 (30). Lane M is a marker mixture of HaeIII
digests ofXX174 phage DNA (25). Sizesof fragments are shown in
kilobase pairs. The infectious virus yields from each sample were
titrated in Vero cells, and a one-step virus growth curve was
con-structed (c) (27).
DISCUSSION
This study is the first identification of the predominant
presenceof novelDNA fragments ofunit-lengthasequencein
HSV-1 DNA preparations (Fig. 3). These fragments were
found inall HSV-1 strainsexamined. Unusual DNA fragments containingpartsoftheasequenceof variouslengths have been
detected in DNA preparations derived largely from viral nucleocapsids of an HSV-1 clone, GN29, with the variant a
sequence (31); the predominant DNAband of unit-length a
sequence was not detected in that study. Therefore, the predominant novel DNAfragment of unit-lengtha sequence
was assumedtobe present in DNAs not packagedwithin the viral particle. Structures of DNA fragments forming bands with the length of the unit-length a sequence were studied by
determining the nucleotide sequences of individual DNA
fragments after molecular cloning and also by analyzing the restriction endonuclease cleavagepatterns ofDNAfragments correspondingtothe unit-lengthasequencewithoutmolecular
cloning (Table 1; Fig. 4). Alargemajority of DNAfragments correspondingto unit-lengthasequence werethosegenerated
by the authentic cleavage of two DR1 encompassing one a
sequence as aresult of the cleavage-packaging system.
The double-strand-break repair model for recombination has been proposed, and strand breaks are presumed to be produced on recombinogenic regions (8, 12, 20, 22).
DR1
is cleaved efficiently at the authentic cleavage site by the cleav-age-packaging system (9, 16, 17, 24). Although inter-arecom-bination does not occur at a single specific site, Smiley et al. hypothesized that site-specific DNA cleavages on DRI in-duced by the cleavage-packaging system stimulate the initia-tion of generalizing recombinainitia-tion by providing recombino-genic free DNA ends (20).
DR1
was found to be involved in the occurrence of illegitimate recombination, i.e., (i) the defective interfering DNA generated by recombination be-tweenDR1
and the unique sequence of the S component (14) and (ii) spontaneous deletions found in HSV-1 variant 1714 and HSV-2 variant JH2604 generated by the recombination betweenDR1
and the major inverted repeat of the L compo-nent (13, 23). Sites onDR1,
except for the authentic cleavage site, and the regions aroundDR1
were broken (Table 1). Thus, DR1 is assumed to be a hot spot for recombination of the a sequence.All but one termini of the molecularly cloned DNA frag-ments from the band corresponding to unit-length a sequence of GN28 were at the authentic cleavage site, while half of the termini of the cloned fragments of GN29 were at other sites (Table 1). The a sequence of GN28 is the standard one, and that of GN29 is the variant one with two DR2 arrays (Fig. lb and c). The DR2 array is a series of DR2 repeats with high G+C content and a strong purine-pyrimidine strand bias, and it can form non-B DNA structures (Fig. 2) (38-40). DNA has the capability of adopting several types of conformation, and unusual DNA structures are presumed to function in genetic regulation and other processes that involve DNA, e.g., recom-bination and chromosome packing (38). Nuclear extracts from tissue culture cell lines contained endonucleolytic activity cleaving sequences of non-B DNA conformation such as the DR2 array and is induced by HSV-1 infection (39). Target site recognition by the enzyme was assumed to be conformation specific rather than sequence specific. The presence of two DR2 arrays on the a sequence of GN29 may lead to a DNA conformation peculiar to the GN29 a sequence and differing from that of the standard a sequence. This conformational difference was assumed to affect actions of the cleavage-packaging system, leading to cleavage at sites other than the authentic one.
The termini located at sites except the authentic cleavage site were classified into five types. The first is that assumed to be made by removing several nucleotides from the authenti-cally cleaved termini, probably as a result of exonucleolytic activity (right terminus of pUK364-14, left termini of pUK364-10 and -15). The second is represented by the termini onDR1, leaving the authentic cleavage site within
DR1
(nt 19) (left termini of pUK365-9 and pUK364-3, -4, and -5). The third is represented by the termini on Uc (left termini of pUK364-9, -14, and -2). The authentic cleavage reaction is thought to be made at defined distances from thepacl
and pac2 signals located within the a sequence (8, 17, 35). Aberrant termini of the second type and that of pUK364-9 (third type) were on or near DR1 and might be due to an error in measuring the distance from thepacl
and pac2 signals in the cleavage reaction. The DNA conformation peculiar to the GN29 a sequence with two DR2 arrays may increase the occurrenceof such an error. The left termini of pUK364-14 andpUK364-2 on Uc were near the stretch involved in generation of thenovel recombination of GN29 (31). The fourth type is the terminus on the inverted repeat of the L component (left terminus of pUK364-1), and the fifth type is the terminus on the DR4VOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.97.256.83.341.2](a)RL DR1
D)R
DR1 DR1 Rs DRI DRI(a)
M o&^ t t
t
*t
RL DRI DRI DRI Rs
(
b)
IV(}1vAvtA t
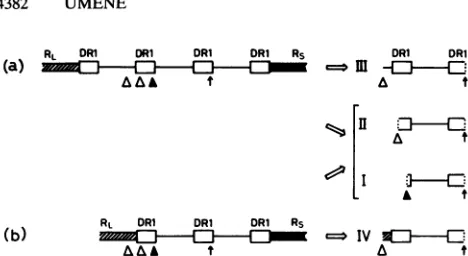
FIG. 6. Models for excision of theunit-lengtha sequence.These
modelsarebasedonthehypothesisthat thecleavage-packagingsystem
acts properlyon theDR1 second distal from the S component and
cleavages ofotherDR1arerareand lessaccurate (8, 16).Three(a) and two (b) copies of thea sequence are bracketed with the major invertedrepeatsof theL component(RL)and the S component(Rs). The authenticcleavage siteonDR1second distal fromRs,onwhich thecleavage-packaging systemactsproperly, is indicatedbyavertical
arrow.The authentic cleavage site on DR1 third distal from Rs is indicatedbyaclosedtriangle. Cleavagesites other than the authentic
cleavage site areindicated byopen triangles on DR1, Uc, and RL. Righttermini (Rsside) of the exciseda sequence are constantand
derived from theauthenticallycleaved endontheDR1second distal
fromRs,asindicatedbyaverticalarrow.However,the left termini(RL side)of the excised DNAsarederived from endson and around the
DR1third distal from Rsandvariable, i.e.,the authenticallycleaved end(I)asindicatedbyaclosedtriangleand other endsonDR1(II),
Uc (III), and RL (IV) as indicated by open triangles. The DNA
fragment ofIis theexactunitlengthof theasequence,both termini of whichareauthenticallycleaved ends duetothecleavage-packaging
system.The DNAfragmentof III is derived fromregionswith threeor more copies ofa sequence. The DNAfragment ofIV is from the
region withtwocopiesof thea sequence.
stretch, corresponding to DR4n2 of GN28 and GN29 (right terminus ofpUK364-9) (31).TheDR4stretchis assumed to be recombinogenic and islikely tobreak (4, 31). Therefore,the production of the rightterminus ofpUK364-9wasattributed to a strand break on DR4n2 and not to an error in the cleavage-packaging system.
Termini located at sites other than the authentic cleavage sitewere more frequentlyobservedontheL-component side
than on theS-component side of the cloned DNA fragment (Table 1). Onlyone aberrant terminus of cloned DNA frag-mentsfrom GN28was onthe left terminus ofpUK365-9but
not on the right. Of 18 right termini of analyzed DNA
fragmentsfromGN28 and GN29, onlytwowereatsites other than the authentic cleavage site (right termini of pUK364-9 and -14). The right termini of pUK364-9 was generated by
breakageonDR4n2, presumably independent of the
cleavage-packagingsystem.Therightterminus ofpUK364-14waslikely
produced byremoval oftwonucleotides fromanauthentically
cleaved terminus by exonucleolytic activity. Thus, cleavage
processesgeneratingtherighttermini seemedtobe accurate.
Of 18 left terminioffragments from GN28 and GN29, 8were
attheauthenticcleavage site. Two termini ofpUK364-10 and -15wereassumedtobe produced byremoval ofonenucleotide
from the authentically cleaved termini. The other eight left terminicannotberegardedasbeing derived fromthe
authen-ticallycleaved termini, because DNA fragments havingthese eight left termini retained the authentic cleavage site on the
leftDR1(nt 19). Therefore, cleavageprocessesgenerating the
left termini were assumed to be less accurate than those generating right termini. The free S-component terminus of linearHSV-1 DNA consists ofonea sequencewithaterminal
DR1 containing 1 bp and one 3' nucleotide extension (16). Thus, cleavage ofcircular or concatemeric DNA due to the cleavage-packaging system has heretofore been assumed to occurwithin a DR1 second distal from the S component (8, 16). The difference in the accuracies of the
cleavage
events observed between the two termini of cloned DNAfragments favoredthehypothesis
that thecleavage-packaging
systemacts properlyonthe DR1 second distal from the S component and does so on other DR1 with a lesserfrequency
and accuracy (Fig.6).Theright
termini of the DNAfragments
found in this studyareassumedtoderivemostly
from theDR1second distal from the S component andareauthentically
cleavedproducts
linked to thecleavage-packagingsystem. The left termini are assumedtoderive
mostly
from theDR1third distal from the S component and are morefrequently
at sites apart from the authenticcleavage
site than theright termini,
because of a lesser degree of accuracy of actionof thecleavage-packaging
system. The cleavage-packaging system acts on the DR1 second distal from the Scomponent and also on other DR1
elements; therefore, the novel excised DNA fragments of
unit-length a sequence were
generated
to the extent that apredominantband became evident (Fig. 6).
ACKNOWLEDGMENTS
GratitudeisextendedtoM.Oharaforassistance withthe prepara-tionof this report.
Part of this studywas supported by grants from the Ministry of Education, Science,and CultureofJapan.
REFERENCES
1. Bortner,C.,T. R.Hernandez,I. R.Lehman,andJ.Griffith.1993. Herpessimplexvirus1single-strandDNA-bindingprotein (ICP8) willpromotehomologouspairingand strand transfer. J. Mol. Biol. 231:241-250.
2. Brown, S.M., J.H. Subak-Sharpe,J. Harland,and A. R. Mac-Lean. 1992. Analysisof intrastrain recombinationinherpes sim-plexvirus type 1 strain 17 andherpessimplexvirus type 2strain HG52usingrestrictionendonucleasesitesasunselectedmarkers and temperature-sensitive lesions as selected markers. J. Gen. Virol. 73:293-301.
3. Bruckner,R.C.,R.E.Dutch,B. V.Zemelman,E.S.Mocarski,and I. R. Lehman. 1992. Recombination in vitro between herpes simplex virus type 1 a sequences. Proc. Natl. Acad. Sci. USA 89:10950-10954.
4. Chou,J.,and B.Roizman. 1985. Isomerization ofherpessimplex virus1 genome:identificationof thecis-actingand recombination sites within the domain of theasequence.Cell 41:803-811. 5. Chou, J.,and B. Roizman.1986.Theterminalasequence of the
herpes simplex virus genome contains the promoter ofa gene located in the repeat sequences of the L component. J. Virol. 57:629-637.
6. Chou, J.,and B. Roizman. 1989. Characterization of DNA
se-quence-common and sequence-specific proteins binding to cis-actingsites forcleavageofthe terminalasequenceof theherpes simplexvirus 1 genome. J. Virol. 63:1059-1068.
7. Davison,A.J.,and N. M. Wilkie. 1981. Nucleotidesequencesof thejointbetween the L and S segments ofherpessimplexvirus types 1 and 2. J.Gen. Virol. 55:315-331.
8. Deiss,L.P., J. Chou,andN. Frenkel. 1986. Functional domains withintheasequenceinvolved in thecleavage-packagingofherpes simplexvirus DNA. J. Virol.59:605-618.
9. Desai, P., N. A. DeLuca, J. C. Glorioso, and S. Person. 1993. Mutations inherpes simplexvirus type1genesencodingVP5and VP23 abrogate capsid formation and cleavage of replicated DNA. J. Virol. 67:1357-1364.
10. Dutch,R.E.,R. C.Bruckner,E.S.Mocarski,andI.R. Lehman.
1992. Herpes simplex virus type 1 recombination: role of DNA replication andviralasequences. J.Virol.66:277-285.
11. Jones, D., R. Isfort, R. Witter, R.Kost, and H.-J. Kung. 1993. Retroviral insertions into a herpesvirus are clustered at the
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.61.295.65.193.2]EXCISION OF UNIT-LENGTH a SEQUENCE OF HSV-1 4383 junctions of the short repeat and short unique sequences. Proc.
Natl. Acad. Sci. USA 90:3855-3859.
12. Kobayashi,I. 1992. Mechanisms for gene conversion and homol-ogousrecombination: the double-strand break repair model and the successive half crossing-over model. Adv. Biophys. 28:81-133. 13. MacLean, A. R., M. Ul-Fareed, L. Robertson, J. Harland, and S. M.Brown. 1991. Herpessimplex virus type 1 deletion variants 1714and 1716 pinpoint neurovirulence-related sequences in Glas-gow strain 17+ between immediate early gene 1 and the 'a' sequence. J. Gen. Virol. 72:631-639.
14. Mocarski, E. S., L. P. Deiss, and N. Frenkel. 1985. Nucleotide sequence and structural features of a novelUs-ajunction present in adefective herpes simplex virus genome. J. Virol. 55:140-146. 15. Mocarski, E. S., and B. Roizman. 1981. Site-specific inversion sequence of the herpes simplex virus genome: domain and struc-tural features. Proc. Natl. Acad. Sci. USA 78:7047-7051. 16. Mocarski, E. S., and B. Roizman. 1982. Structure and role of the
herpessimplex virus DNA termini in inversion, circularization and generation of virion DNA. Cell 31:89-97.
17. Nasseri, M., and E. S. Mocarski. 1988. The cleavage recognition signal is contained within sequences surrounding an a-a junction in herpes simplex virus DNA. Virology 167:25-30.
18. Pater,M. M., R. W. Hyman, and F. Rapp. 1976.Isolation of herpes simplex virus DNA from the "Hirt supernatant."Virology 75:481-483.
19. Roizman, B. 1979. The structure and isomerization of herpes simplexvirus genomes. Cell 16:481-494.
20. Smiley, J. R., J. Duncan, and M. Howes. 1990. Sequence require-ments for DNArearrangements induced by the terminal repeat of herpessimplex virus type 1 KOS DNA. J. Virol. 64:5036-5050. 21. Smiley, J. R., C. Lavery, and M. Howes. 1992. Theherpes simplex
virus type I (HSV-1) a sequence serves as acleavage/packaging signal but does not drive recombinational genome isomerization whenit is inserted into the HSV-2 genome. J. Virol. 66:7505-75 10. 22. Szostak, J. W., T. L. Orr-Weaver, R. J. Rothstein, and F. W. Stahl. 1983. The double-strand-break repair model for recombination. Cell33:25-35.
23. Taha, M. Y., G. B. Clements, and S. M. Brown. 1989. Theherpes simplex virus type 2(HG52)variant JH2604 hasa1488bpdeletion which eliminates neurovirulence in mice. J. Gen. Virol. 70:3073-3078.
24. Tengelsen, L. A., N. E.Pederson, P. R.Shaver,M. W.Wathen,and
F. L. Homa. 1993.Herpessimplexvirus type 1 DNAcleavageand encapsidation require the product of the UL28 gene: isolation and characterization oftwoUL28deletionmutants.J.Virol. 67:3470-3480.
25. Umene,K. 1985.Variabilityof the regionof the herpes simplex virus type 1 genome yielding defective DNA: SmaI fragment polymorphism. Intervirology23:131-139.
26. Umene, K. 1985. Intermolecular recombination of the herpes simplexvirus type Igenomeanalysedusingtwostrainsdifferingin
restriction enzymecleavage sites. J. Gen. Virol. 66:2659-2670. 27. Umene, K. 1986. Conversionof a fraction of the unique sequence
topartofthe inverted repeats in the S component of the herpes simplex virus type 1 genome. J. Gen. Virol. 67:1035-1048. 28. Umene,K.1987.Transition fromaheterozygousto ahomozygous
state ofa pairof loci in the inverted repeat sequences ofthe L component of the herpes simplex virus type 1 genome. J. Virol. 61:1187-1192.
29. Umene, K. 1989. Short, duplicated sequence indicative of the recombinogenicity of the junction between a unique and an inverted repeat sequence in the S component of the herpes simplex virus type 1 genome. J. Virol. 63:1877-1883.
30. Umene, K. 1991. Recombination of the internal direct repeat element DR2 responsible for the fluidity of the a sequence of herpessimplex virus type 1. J. Virol. 65:5410-5416.
31. Umene, K. 1993. Herpessimplex virus type I variant a sequence generated by recombination and breakage of the a sequence in defined regions,including the one involved in recombination. J. Virol. 67:5685-5691.
32. Umene, K., and L. W. Enquist. 1985. Isolation of novel herpes simplex virus type 1 derivatives with tandem duplications of DNA sequences encoding immediate-early mRNA-5 and anorigin of replication. J. Virol. 53:607-615.
33. Umene, K., and M. Yoshida. 1989. Reiterated sequences of herpes simplex virus type 1 (HSV-1) genome can serve as physical markers for the differentiation of HSV-1 strains. Arch. Virol. 106:281-299.
34. Umene, K., andM. Yoshida. 1993. Genomic characterization of twopredominant genotypes ofherpes simplex virus type 1. Arch. Virol. 131:29-46.
35. Varmuza, S. L., and J. R. Smiley. 1985. Signals for site-specific cleavage of HSV DNA: maturation involvestwoseparatecleavage events atsites distal to therecognition sequences. Cell 41:793-802. 36. Weber, P. C.,M.D.Challberg, N. J. Nelson,M.Levine, andJ. C. Glorioso.1988.Inversioneventsinthe HSV-1 genomearedirectly mediated by the viral DNA replication machinery and lack se-quencespecificity. Cell 54:369-381.
37. Weber, P. C., M. Levine, andJ. C. Glorioso. 1990. Recombino-genic properties of herpes simplexvirus type 1 DNA sequences resident in simian virus 40 minichromosomes. J. Virol. 64:300-306. 38. Wells, R. D., D. A. Collier, J. C. Hanvey, M. Shimizu, and F.
Wohlrab. 1988. The chemistry and biology of unusual DNA
structures adopted by oligopurine*oligopyrimidine sequences. FASEB J. 2:2939-2949.
39. Wohlrab, F., S. Chatterjee, and R. D. Wells. 1991. The herpes simplexvirus 1 segment inversion site isspecificallycleavedbya
virus-induced nuclear endonuclease. Proc. Natl. Acad. Sci. USA 88:6432-6436.
40. Wohlrab, F.,M.J. McLean,and R. D. Wells. 1987. Thesegment inversion site ofherpes simplexvirus type 1adoptsanovel DNA
structure.J. Biol. Chem. 262:6407-6416. VOL.68, 1994