0022-538X/86/100037-06$02.00/0
Copyright ©) 1986, American Society for Microbiology
Effect of Interferon
on
Replication of
Herpes
Simplex Virus
Types
1and 2 in
Human
Macrophages
INGRID DOMKE-OPITZ,* PETRA STRAUB, AND HOLGER KIRCHNER
Institiute ofViruis Research, German Cancer Research Center, Heidelberg, Feder-alReputblic
of
GermanyReceived 31 January 1986/Accepted 12 June 1986
Macrophages derived from human peripheral blood and cultured for 1 week were permissive for the
replicationof herpes simplex virus (HSV) types 1 and2. Low titers of interferon (IFN) were producedafter
virus infection. The yield of infectious virions was reduced by pretreatment of cells with natural and
recombinantIFN-otand naturalIFN-i.Recombinant and naturalIFN-yexhibitedverylow antiviral activity.
Treatment ofcells withIFN-ymixed with IFN-otorwith IFN-I didnotresult inasynergistic inhibition of virus
yield. We studied the synthesis of HSVtype 1- and HSVtype2-coded proteins in macrophages treated with
IFN-,. Induction of the HSV "-protein DNA polymerase was strongly inhibited in IFN-treated cells in a
dose-dependentmanner.As shown by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, otherI-and
-y-proteins of HSV were inhibited as well. Immunofluorescence studies revealed a strong inhibition of the
expression of immediate early a-protein ICP4. The results indicate that IFN acts early during the viral
replicationcycletoinhibitthe synthesis of HSVa-and IT-proteins.
Macrophages belong to the nonspecific defense
mecha-nisms and are known to be activated rapidly during a
primary infection with herpes simplex virus (HSV) (15). The role of mononuclear phagocytes in defense against HSV infections has been studied extensively in experimental
animals. Althoughmouse macrophages arecapable of virus
phagocytosis and of restricting virus infection by the
pro-duction of interferon (IFN) (16), they also replicate HSV
underappropriate culture conditions (4, 23).
HSV has been isolated from leukocytes of human patients
(12). In several studies,targetcells have been identified. The
virus has been shown to replicate in Epstein-Barr
virus-stimulated B cells (17) and in mitogen-stimulated T cells
expressing Ta antigen (3). HSV infection of fresh blood
monocytes leads to an abortive infection (28). However, after several days ofin vitro culture and differentiation into
macrophages, monocytes support virus replication (21). It
hasbeen shownby morphologicaland functional studies that invitro differentiation of human bloodmonocytesresembles in vivo differentiation of human peritoneal macrophages
(13). Thus,it appearsthat differentiated tissue macrophages
might replicateHSV andtherebycontributetodissemination
of the virus. Thispromptedusto testthe effect of IFNonthe
synthesis of infectious virions and viral proteins in human
macrophages.
MATERIALS AND METHODS
Cell cultures. Human peripheral blood mononuclear cells
were obtained by Ficoll-Hypaque density gradient
centrifu-gation (1) from buffycoats ofhealthy donors. Mononuclear
leukocytes were seeded at aconcentration of4 x 106/ml in
35-mmpetridishes(BectonDickinson andCo., Heidelberg,
Federal Republic of Germany) in medium consisting of RPMI 1640 (GIBCO Europe GmbH, Karlsruhe, Federal
Republic of Germany) supplemented with 10%
heat-inactivated, pooled human AB serum (Flow Laboratories,
Meckenheim, Federal Republic of Germany), minimal
es-sential medium vitamin solution (1:100), L-glutamine (2
* Correspondingauthor.
mM), sodiumpyruvate(1 mM), HEPES
(N-2-hydroxyethyl-piperazine-N'-2-ethanesulfonic acid) buffer (10 mM),
peni-cillin (100 IU/ml), and streptomycin (100 ,ug/ml). After incubationfor 16 hat 37°C, nonadherent cellswere washed
offby vigorouspipettingwith Hanksbalanced salt solution.
Theremainingadherentcellswerefurther cultured inRPMI
1640 medium containing 2% pooled human AB serum and thecomponents listed above. The mediumwasreplacedon alternate days. During in vitro culture, blood monocytes
were differentiated to macrophages. The cells increased in
size and assumed a spindle shape. After 6 days of culture, the numberof cellsperdishwasabout 5 x 105.The cultures
consisted of 100%macrophagesasshownby phagocytosisof latexparticles, byesterasestaining (18), and by
immunoflu-orescence assay with the monoclonal antibody OKM1
(Ortho Diagnostics, Inc., Heidelberg, Federal Republic of
Germany).
Herpesviruses and virus titrations. A pool ofHSV type 1
(HSV-1) strain WAL was prepared as previously described
(33). Primary isolates of HSV-1 (numbers 214 and 337)and
HSV-2 (numbers 184 and 311) were kindly provided by K. E. Schneweis (Institut fur Medizinische Mikrobiologie
undImmunologie, Bonn, FederalRepublicofGermany)and
werepassaged onceonRITA cellsof Africangreenmonkey
kidney origin (RC-37 RITA; Italdiagnostics, Rome, Italy).
Virus titrations were carried out by a plaque assay with RITA cells. Titers in macrophages were determined after
freezingandthawing. Cellswere storedat -70°Cuntilassay
of virus yield.
IFNs and IFN assay. Recombinant
IFN-ot,
(9.2 x 106IU/mg) was a gift of C. Weissmann to H.K. Recombinant
IFN-aX2 (1 x 107 IU/mg) was kindly given to us by S.
Hiemstra (Essex Pharma, Munich, Federal Republic of
Germany). Natural IFN-ox(2.1 x 105 IU/mg)waspurchased
from Stratech Scientific Ltd. (London, United Kingdom).
Natural IFN-13 (5 x 106 IU/mg) was supplied by Serono
GmbH (Freiburg, Federal Republic of Germany). Native
IFN-y (8.3 x 106 IU/mg) was a gift of P. von Wussow
(Medizinische Hochschule, Federal Republic ofGermany)
toI. D.-O.Recombinant IFN-y (3.3x 107IU/mg)waskindly
provided by Biogen (Geneva, Switzerland). Forall
prepara-37
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Replicationof different strains of HSV in human macrophages
Virus titers Virus strain" (PFU/ml)b HSV-1 WAL... 6 x 10' HSV-1 214... 6 x 104
HSV-1 337... 7 x 105
HSV-2 184... 8 x 103
HSV-2 311... 1 x 105
aMOI, 0.002.
hCellswerefrozenat 5daysafterinfection.
tions, the titers weredeterminedin our laboratorybeforeuse
in experiments. Titers of the IFN preparations were
deter-mined against a reference standard (no. 23-902-530; National Institutes of Health, Bethesda, Md.). IFN titers were
deter-mined in a system of HEp-2 cells challenged with vesicular
stomatitis virusaspreviously described(8). Cells werefixed and stained 24 h after infection with vesicular stomatitis virus.
IFNtreatment and virus infection. Afterbeing cultured for
6days,themacrophages wereincubated with IFNfor18 h.
Before virus infection, IFN was removed by washing the
cells several times.The cellswere inoculatedwith0.2 mlof
virus for 1 h at different multiplicities of infection (MOI). After virus adsorption, the cells were washed, and 2 ml of
RPMI 1640 medium containing 20% fetal bovine serum
(GIBCO) was added. All experiments were performed in
duplicate cellcultures.
Assayof HSV DNA polymerase. Viral enzymeactivitywas
determined in postmitochondrial supernatants of cell
ex-tracts as described previously (27, 29). At indicated time
points, macrophages were washed once with Hanks
bal-anced saltsolutionand once withcold extraction buffer(10
mMTrishydrochloride [pH8.01, 2 mM2-mercaptoethanol,
20% glycerol). Cells were scraped with a rubberpoliceman
into 0.4 ml ofextraction buffer and stored at -70°C. After
thawing, cellextracts werecentrifugedat 12,000 x gfor15
min at4°C. HSV DNApolymerase activitywas assayedby
measuringtheincorporationof[3H]TTP (specific activity,50
Ci/mmol) into trichloroacetic acid-insoluble material with
activated DNA as the template primer. For suppressionof
cellular DNApolymerases, KCl(0.2 M) was included in the
assay mixtures. Residual cellular enzyme activities are
in-cluded in the data. All enzyme assays were performed in
triplicate in the linear range of enzyme activity. Protein
concentrations were determined by the Bio-Rad protein
assay(2).
Analyses of HSV proteins. Human macrophages cultured
on35-mmpetri dishes were infected with HSV. At 7.5 h after
infection, the medium was replaced in Eagle basal medium
TABLE 2. Production of IFN by HSV-infected humanmacrophages
IFN concn(IU/ml)
Hafterinfectiona
ExptI Expt II
4 13 21
9 16 21
12 13 35
24 13 56
48 16 21
72 <3 21
aCellswereinfectedwith HSV-1 at an MOI of 2.
without methionine, to which 40 ,Ci of [35S]methionine per
mlhad been added(Amersham Buchler, Brunswick,Federal
Republicof Germany). The cellswereharvestedafter 90 min
ofpulse-labeling. Labeled proteins were subjected to
elec-trophoresis on 8% sodium dodecyl sulfate-polyacrylamide
gels (19). For determination of molecular weights, protein
markers (Sigma Chemical Co., St. Louis, Mo.) were
in-cluded.
Immunofluorescence. For immunofluorescence studies,
macrophages grown on slides were infected with HSV-1.
Theywere fixed with methanol-acetone at -20°C 6 h after
virus infection. Cells were incubated for 30 min at room
temperature with a mixture containing phosphate-buffered
saline, 1% bovine serum albumin, and 10% fetal bovine
serum to preventunspecific binding of antibodiesto the Fc
receptorsof macrophages. Afterbeingwashed in
phosphate-buffered saline-1% bovine serum albumin, the cells were
coated for 30 minatroomtemperaturewitha1:10dilution of
amonoclonal antibody againstHSV-1 ot-protein ICP 4(30).
Afterbeing washed, slides were labeled for 30 minatroom
temperaturewith fluoresceinisothiocyanate-conjugated Fab
fragment (Dianova GmbH, Hamburg, Federal Republic of
Germany) diluted 1:25. The cellswere examinedand
photo-graphed under UV-optics with a microscope (Dialux 20;
Wild-Leitz GmbH, Wetzlar, Federal Republic of Germany).
RESULTS
Replication ofHSV andIFNproduction in human
macro-phages. Human blood monocytes cultured for 1 week
sup-portedthereplicationof HSV. As shownpreviously (20,28) susceptibilitytoHSVincreasesduringinvitrodifferentiation
ofblood monocytes to macrophages after several days of
culturing (datanotshown).ThecellswerereplicatingHSV-1
aswell as HSV-2 strains (Table 1). The yield of infectious
virusparticles after infectionat an MOIof 0.002wassimilar
with the four primary isolates tested when compared with
laboratory strain WAL.
Macrophages werecapable ofIFNproduction after infec-tion with HSV at high multiplicity. However, only small
':0
20~~~~
C 0~~~~~~~~
0~~~~~~~~~
0 0
2
-1 2 3 4
log10 lU/miIFN
FIG. 1. Inhibition of HSV-1 replication by different types of IFN.CellsweretreatedwithnativeIFN--y(A), recombinantIFN-y
(-), recombinant IFN-ot, (0), recombinant IFN-a2 (0), natural IFN-ox(H), or natural IFN-,B (A) for 18 h and infected with HSV-1
at an MOI of 2. Cells were stored at -70°C at 24 h after virus infection.Virus yieldin untreated cultures was 2 x 106PFU/ml.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.57.294.626.712.2]amounts of IFN were produced (Table 2). IFN was detected
by4haftervirus infection.The capacity for IFN production varied greatly among blood macrophages from different donors.
Effect of differentIFNs on HSV replication. Wecompared
theantiviralactivities ofdifferent types of native and recom-binant IFNs in human macrophages (Fig. 1). Inhibition of
HSV-1 replication was dose dependent with natural IFN-a and
IFN-P
as well as withrecombinantIFN-ao
andIFN-a2. However,the natural IFNsappeared to be more active thanthe recombinantIFNs. Inhibition was most pronounced with
native
IFN-P.
Pretreatment of macrophages with 10 IU ofIFN-P
per ml resulted in90% inhibition of virus growth. Incontrast,
IFN-y
exhibited very low antiviral activity inhumanmacrophages. This result was obtained with natural
IFN--y and with recombinant IFN-y. Log10reduction ofvirus
titerswas about 0.7, evenafter treatment of cells with IFN
dosesas high as
104
IU/ml.Additionally, we tested the effect of combinations of
different IFNs on the replication of HSV-1 (Table 3).
Syn-ergistic activitiesbymixturesof IFN--y and other IFNtypes
havebeendescribedforinhibition ofHSV in mouse
macro-phages (9) or cell growth inhibition (7, 10). However, we
could notfind apotentiation of antiviral activities ofIFN-a or IFN-1 by
IFN-y.
Effect of IFN on induction of HSV DNA polymerase. To
determine the stage of virus replication that is blocked by
IFN, we tested the effect ofIFN on the induction ofHSV
DNA polymerase. The virus-coded enzyme belongs to the
class of ,B-proteins and is expressed early during the viral
replication cycle. We tested the induction of the enzyme aftertreatmentofhumanmacrophages with different doses
ofnative
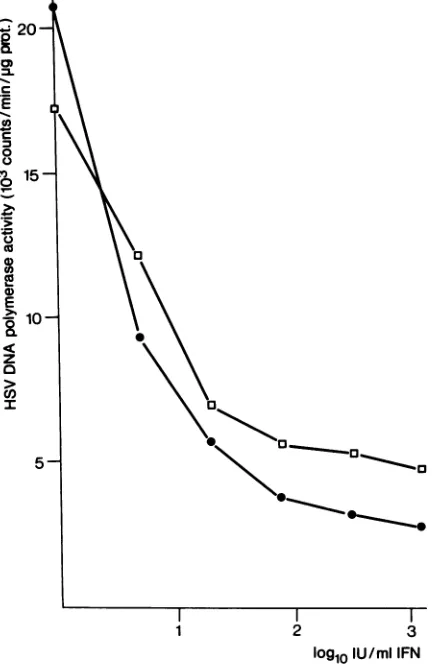
IFN-3
(Fig.2).HSVDNApolymeraseactivitywasstrongly reduced with IFN doses as low as 5 IU/ml. The
induction of HSV-2DNApolymerasewas moresensitiveto
IFN than was thatofHSV-1 DNA polymerase. After
treat-mentof cells withhighIFNdoses,theHSV-2-codedenzyme wasreduced nearlyto thecellular background level.
As described above, macrophages can be induced to
produce IFN. To analyze theeffect ofendogenous IFN on
theinduction of HSVDNApolymerase, wetreatedthecells
with
poly(I):poly(C).
Only moderate IFN titers weremea-sured in the supernatants of poly(I):poly(C)-treated cells
(Table 4). Subsequent infection with HSV-1 or HSV-2
re-sultedinamarked inhibition of HSVDNApolymerase. The
degree of inhibition by endogenous IFN wascomparable to
that observed with exogenous IFN. Again, inhibition of
HSV-2 DNApolymerase was morepronounced.
[image:3.612.331.548.75.407.2]The effect of IFN on the kinetics of induction ofHSV DNA polymerase was analyzed in human macrophages
TABLE 3. Inhibition of HSVreplication by combinations of differenttypesofIFN
a
~~~~~~~~~~~Logl0
reduction ofIFNtype" IFN dose(IU/ml) virusyieldb
a 300 1.2
,B 300 2.1
y 300 0.3
+ ,8 150 + 150 2.0
a+y 150 + 150 1.3
+ -Y 150+ 150 2.1
aCells were pretreated with natural IFN-a, natural
IFN-P,
and naturalIFN-,yasindicated.
bCellswereinfected with HSV-1at anMOIof2 andstoredat-70°Cat24 hafter infection. Virusyieldin untreated cultureswas9 x105PFU/ml.
0
0
D"
5.0
15E
z
5 0
1 2 3
10910
lU/miIFN FIG. 2. Dose response of IFN for inhibition of HSV DNA polymerase. Aftertreatment withIFN-P, cells were infected with HSV-1 (O) or HSV-2 (0) at an MOI of 6. Macrophages were harvestedforassayofHSVDNApolymeraseactivityat9hafter infection. Residual cellularenzyme activity inuninfected cultures was1.834cpm/,Lgof protein.infected with HSV-1 or -2 (Fig. 3). Kinetics of enzyme
inductionweresimilar with bothvirustypes. Enzyme
induc-tion wasdelayedin IFN-treated macrophages. Inuntreated
cultures, the first increase of viral enzyme activity was
observedat6 hafterinfection. Atthis time,DNA
polymer-ase activity in macrophages pretreated with IFN was
com-parable tocellularbackgroundlevels.
Synthesisof HSVproteinsinIFN-treatedmacrophages. As
shown by the results of the induction ofHSV DNA
poly-merase, IFN acted on
p-protein
synthesis. Sodiumdodecylsulfate-polyacrylamide gelelectrophoresiswas
performed
toanalyzethepatternof
synthesis
ofadditionalHSVproteins.
Macrophageswerepulse-labeled
with[35S]methionine
at7.5 hafter infection. Proteinswereidentifiedby
comparing
theirmolecular weights with
published
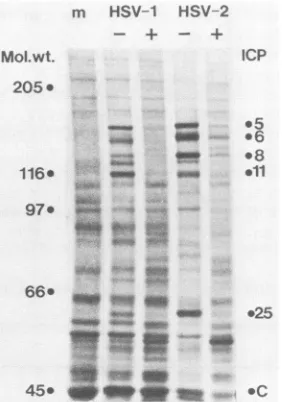
data (24). The resultspresented in Fig. 4 show a strong inhibition of HSV
P-proteins, namely ICP6 and ICP8. The synthesis of several
-y-proteins (ICP5, ICP11, and ICP25) were also inhibited in
IFN-treated cells. The resultsweresimilar for HSV-1 and -2.
We tried to analyze the effect of IFN on HSV
a-protein
synthesis by overinduction with
cycloheximide
andactinomycin D. However, it was not
possible
to detecta-proteins
ongels.
Inaddition,
detection of ICP4 wascomplicated bya
comigrating
cellularprotein.
Therefore,
westudieda-protein
synthesis by
immunofluorescenceby
usingon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.63.305.607.689.2]TABLE 4. Inhibition of HSV DNA polymerase by endogenous andexogenousIFN
DNApolymerase
IFNconcn activity(cpm/,ug
Treatment"l (IU/ml)b ofprotein)"
HSV-1 HSV-2
<3 15,140 16,067 Poly (I-C) (100 jig/ml) 60 11,432 6,039
Poly (I-C)(200 jig/ml) 100 6,958 3,298
IFN-P(100 lU/mI) 7,630 5,193
aPoly(I):poly(C)wasaddedfor 1.5 h, and the cells werewashedand further
culturedfor 16.5 h.
bSupernatantswereharvested beforeHSVinfection.
'Cellswereinfectedat anMOI of6and harvestedat8 hafterinfection.
Enzyme activity in uninfectedcultures was 1.235cpm/,lgof protein.
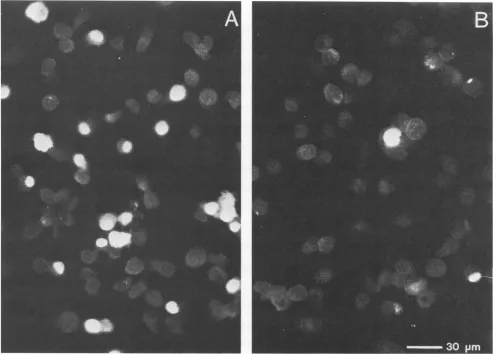
amonoclonal antibody against HSV-1 ICP4. About 40% of
thecells werepositive fortheexpression of ICP4at6hafter
infection(Fig. 5A). In contrast, after IFN treatmentonly4%
of the cells showed a fluorescent signal. All the results
_25
.',I
CL0
o20
SO
E 9-26 15 1
15
z
10
10~~~~~~~~~~~~~~
U-.-- ---o
3 6 9 12 15 18
Hoursafter infection
FIG. 3. Kineticsof induction of HSV-1- and HSV-2-coded DNA polymerase inIFN-treated macrophages. Untreated cells(0,*) or cellstreatedwith103IUof IFN-,B per ml(0,O) were infected with HSV-1(0, 0)orHSV-2 (-, O) at an MOI of 6. Cells were harvested
at the indicated time points for assay of HSV DNA polymerase activity. Residual cellular enzyme activity in uninfected cultures was204
cpm/Vjg
ofprotein.indicate that IFN inhibits HSVat anearly stage of the viral
replication cycle.
DISCUSSION
The present studywasperformed toanalyze the effect of
IFN onHSVreplication in human primary cells that might
play akeyrole in thecourseofanHSV infection. Linnavuori
and Hovi (21) have shownarestriction ofHSVreplicationin
undifferentiated blood monocytes by high titers ofIFN that
wereinducedafter virus infection. However, only low titers
ofIFN wereproducedafter HSV infection of differentiated
macrophages, and the cellswere permissivefor virus
repli-cation. Thus, the role of endogenous IFN induced by the
virus itself seems to be negligible in differentiated
macro-phages. However, if IFN was induced in macrophages
before HSV infection, it was veryeffective in
inhibiting
thevirus.
Humanmacrophages weresensitivetothe antiviral
activ-ity ofexogenous IFN-o and
IFN-P,,
whereasIFN--y
exhib-itedonlyverylittleantiviral activity.Theeffectwas
macro-phage specific, because inhibition of HSV replication on
HEp-2 cellsbyIFN--ywascomparabletovirus inhibitionby
other IFN types(datanotshown). Inaddition, experiments
with human cell lines have revealed that similar
concentra-tions of
IFN-y
or IFN-ot or IFN-, were necessary forinduction of the antiviral state measured via induction of
(2'-5') oligoadenylate synthetase (32). A recent study
de-scribes differences in biological activitiesofIFN--yonhuman
monocytes and WISH cells; a comparison of the IFN-y
receptor on both cell typesrevealed different binding
char-acteristics (26). Thus, the lack of antiviral activity of IFN-Y
againstHSV on macrophages couldbe related to a different
receptor onthese cells.This is inagreementwith thefinding
that IFN-y is ineffective in potentiating the antiviral
activi-m HSV-1 HSV-2
ICP
Mol.wt. 205.
I 5
_ *6
,s
_s
*6
116 _ _ *11
97*
t *f-l..:
66-
_ _
mr.Tt>
._ mora
4
*25
FIG. 4. Analysis of HSV proteins in IFN-treated macrophages by sodium dodecyl sulfate-polyacrylamidegel electrophoresis. Un-treated cells (-) or cells Un-treated with1,000 IU of
IFN-P
per ml (+) wereinfectedwith HSV-1 or HSV-2 at an MOI of 6. m, The protein pattern ofmock-infected cultures. Cells were pulse-labeled for 90 min with[35S]methionine, and each lane received an equal amount ofcell protein extracts. Molecular-weight markers and ICPs are marked.C, Amajorcellularprotein.on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.56.297.94.182.2] [image:4.612.56.293.308.660.2] [image:4.612.369.510.445.646.2]FIG. 5. Effect of IFN on the expression of HSV-1 a-protein ICP4. Untreated cells (A) or cells pretreated with 1,000 IU ofIFN-Pper ml (B)wereinfected with HSV-1 at an MOI of 6. Cells were fixed at 6 h after infection and subjected to immunofluorescence with a monoclonal antibody against HSV-1 a-protein ICP4.
ties ofIFN-a or
IFN-P.
Weplan to analyze the induction ofenzymes supposed to be related to the antiviral state by
differenttypesofIFN inhuman macrophages.
Itwastheaim ofthis study to characterize the stage of the
viral replicationcycle that is mainly blocked by IFN. In our
experimental system, the synthesis of
P-
and -y-proteinscodedforby HSV-1or-2 wasclearlyinhibited. Inductionof
HSV DNA polymerase was delayed in IFN-treated cells.
This suggests that IFNmight effect HSV a-protein synthesis
becauseexpressionof,-proteins dependsonthe presenceof sufficient amounts of
at-proteins
(14). We have shown areduction in the a-protein level in IFN-treated cells by immunofluorescence with a monoclonal antibody against
HSV-1 a-protein ICP4. Similar findings were reached by
Gloger and Panet (11) withHeLa cells. Lipp and Brandner
(22) describe inhibition of HSVprotein synthesis inprimary
African green monkey kidney cells treated with poly(I): poly(C). Recent studies in ourlaboratory revealeda strong
inhibition of HSV-1
at-
and ,-proteinsynthesis
inIFN-treated splenic mousemacrophages (9, 31). Compared with
humanmacrophages, thesecells weremore sensitivetothe
antiviral activity of IFNagainst HSV. One reason mightbe
that the mouse macrophages werecultured in the presence
of colony-stimulating factor, which causes activation and
proliferation ofthecells.Nosimilargrowthfactor was used
in ourstudies of human macrophages.
Other studies have reached contradictory findings
con-cerningthe stageofHSVreplicationthatisblockedby IFN.
Munoz and Carrasco (25) as well as Chatterjeeet al. (5, 6)
claim that IFN inhibits alate stage duringthe viral replica-tion cycle.Whereas theformerauthors suggest that IFN acts
via the production of defective virions,the latterclaimthat
IFN inhibits the release of virus particles. Chatterjee etal.
(6) find noinhibition of HSVDNAsynthesisin IFN-treated
fibroblasts. Thesynthesis of normal levels of viral DNA in
IFN-treated cells should not be possible, given the strong
inhibition of HSVDNApolymerasedescribed in the present
studyand shown
previously
(9, 11). Comparingthe studies, it isunlikely
thatdifferentIFNdosesorpreparations
arethereasonfor thecontradictory findings.Moreover,Glogerand
Panet(11)aswellasMunozand Carrasco(25)used thesame
cell line. Nonetheless, it cannot be denied that HSV
repli-cationis inhibitedbyIFN in different cell types
by
differentmechanisms.
Furtherstudies remaintobe doneto
analyze
theantiviralactivity ofIFN
against
HSV. We willcontinue this workby
comparingthesynthesis
of viraltranscripts
as well asviral DNAsynthesis
in IFN-treatedmacrophages
and fibroblasts.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.68.564.70.424.2]The results of the prescnt study indicate that IFN blocks a
stepbefore HSV DNA synthesis. The mainblock during the
viral replication cycle occurs either during or before the
synthesis of immediate early(x-proteins.
ACKNOWLEDGMENTS
We thank M. Zweig for providing the antibody against HSV a-protein ICP4and H. Jacobsenfor helpful discussions.The excel-lent technical assistance of P. Hagendorn and the outstanding editorial work ofM. Kasamaschareacknowledged.
This research was supported by grant DO 301/1-1 from the DeutscheForschungsgemeinschaft.
LITERATURE CITED
1. Boyum, A. 1968.Separation of leucocytes from blood and bone marrow. Scand. J. Lab. Invest. 21(Suppl.97):77-89.
2. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities ofproteins utilizing the principle of the protein-dye-binding. Anal. Biochem. 72: 248-254.
3. Braun, R., H. Teute, H. Kirchner, and K. Munk. 1984. Repli-cation of herpes simplex virus inhuman Tlymphocytes: char-acterizationof the viraltargetcell. J. Immunol. 132:914-919. 4. Brticher,J., I.Domke, C. H.Schroder,and H. Kirchner. 1984.
Experimental infection of inbredmice withHSV. VI. Effect of interferon onin vitro virusreplication in macrophages. Arch. Virol. 82:83-93.
5. Chatterjee,S., E. Hunter, and R. Whitley.1985. Effect of cloned humaninterferons on protein synthesis and morphogenesis of herpessimplexvirus. J. Virol. 56:419-425.
6. Chatterjee, S., A. D. Lakeman, R. J. Whitley, and E. Hunter. 1984. Effect of cloned human interferons on thereplication of and cell fusion induced by herpes simplex virus. Virus Res. 1:81-87.
7. Czarniecki, C. W., C. W. Fennie, D. B. Powers, and D. A. Estell. 1984. Synergistic antiviral and antiproliferative activities of Escherichia coli-derived human alpha, beta, and gamma interferons.J. Virol. 49:490-496.
8. Doldi, K., M. Leroux, R. Augustin, H. Kirchner, and J. R. Kalden. 1985. Proliferation andinterferon production inwhole blood samples and isolated lymphocyte preparations. J. Inter-feronRes. 5:55-64.
9. Domke, I., P. Straub, H. Jacobsen, H. Kirchner, and A. Panet. 1985.Inhibitionofreplication of herpessimplexvirus inmouse macrophages by interferons. J. Gen. Virol. 66:2231-2236. 10. Fleischmann,W.R., Jr. 1982. Potentiation of the direct
anticel-lularactivity ofmouseinterferons: mutualsynergismand inter-feron concentrationdependence. Cancer Res. 42:869-875. 11. Gloger, I., and A. Panet. 1984. Synthesis of herpes simplex virus
proteins in interferon-treated human cells. J. Gen. Virol. 65:1107-1111.
12. Graig, C. P., and A. Nahmias. 1973. Different patterns of neurologic involvementwith herpes simplex virus types 1 and 2: isolation ofherpes simplex virus type 2 from the buffy coat of twoadults withmeningitis.J. Infect. Dis. 127:365-372. 13. Hammerstrom, J. 1979. Human macrophage differentiation in
vivo and in vitro. Acta Pathol. Microbiol. Scand. Sect. C 87:113-120.
14. Honess, R. W., and B. Roizman. 1974. Regulation of herpesvirus macromolecularsynthesis. I.Cascade regulation of the
synthe-sis ofthree groupsof viralproteins.J. Virol. 14:8-19. 15. Kirchner, H. 1982. Immunobiology of infection with HSV.
Monogr. Virol. 13:1-104.
16. Kirchner, H., H. Engler,C. H.Schroder, R.Zawatzky,andE. Storch. 1983. Herpes simplex virus type 1-induced interferon productionandactivationof natural killer cellsinmice.J.Gen. Virol. 64:437-441.
17. Kirchner, H., and C. H. Schroder. 1979. Replication of herpes simplex virus in human B lymphocytes stimulated by Ep-stein-Barr virus.Intervirology 11.61-65.
18. Koski, I. R., D. G. Poplack, and R. M. Blaese. 1976. A nonspecific esterase stain for the identification of monocytes and macrophages, p. 359-362. In B. R. Bloom (ed.), In vitro methods in cell-mediated andtumorimmunity. Academic Press, Inc.,NewYork.
19. Laemmli, U. K. 1970.Cleavageofstructuralproteinsduringthe assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
20. Linnavuori, K., and T.Hovi. 1981. Herpessimplex virus infec-tion inhuman monocyte cultures: dose-dependent inhibition of monocytedifferentiationresulting inabortiveinfection.J.Gen. Virol.52:381-385.
21. Linnavuori, K., and T. Hovi. 1983. Restricted replication of herpes simplex virus in human monocyte cultures: role of interferon. Virology 130:1-9.
22. Lipp, M., and G. Brandner. 1980. Inhibitionofherpessimplex virus type 1 specific translation in cells treated with poly(rI) poly(rC).J.Gen. Virol.47:97-111.
23. Lopez, C., and G. Dudas. 1979. Replication of herpes simplex virus type 1 in macrophages from resistant and susceptible mice. Infect. Immun.23:432-437.
24. Morse, L.S.,L.Pereira,B.Roizman, and P. A.Schaffer. 1978. Anatomyofherpessimplex virus (HSV)DNA. X. Mapping of viralgenesbyanalysis ofpolypeptides and functionsspecific by HSV-1 x HSV-2 recombinants.J. Virol. 26:389-410.
25. Munoz, A., and L. Carrasco. 1984. Formation of non-infective herpesvirus particles in cultures cells treated withhuman inter-feron.J. Gen. Virol. 65:1069-1078.
26. Orchansky, P., M. Rubinstein, and D. G. Fischer. 1986. The interferon-yreceptor in humanmonocytesis differentfromthe oneinnonhematopoietic cells. J. Immunol. 136:169-173. 27. Panet, A., and H. Falk. 1983. Inhibition by IFN of HSV
thymidine kinase and DNA polymerase in infected and bio-chemically transformed cells. J.Gen. Virol. 64:1999-2006. 28. Plaeger-Marshall, S., and J. W. Smith. 1978. Experimental
infection of subpopulations of human peripheral blood leucocytesby herpessimplex virus. Proc.Soc. Exp. Biol. Med. 158:263-268.
29. Purifoy, D. J. M., and M.Benyesh-Melnick. 1975. DNA
poly-merase induction by DNA-negative temperature-sensitive mu-tantsof HSV type 2. Virology68:374-386.
30. Showalter, S. D., M. Zweig, and B. Hampar.1981. Monoclonal antibodiestoherpes simplexvirus type 1proteins,includingthe immediateearlyprotein ICP4. Infect. Immun.34:684-692. 31. Straub, P., I. Domke, H.Kirchner,H.Jacobsen, and A. Panet.
1986. Synthesis ofherpes simplex virus proteins and nucleic acids ininterferon-treated macrophages. Virology 150:411-418. 32. Wallach, D., M. Fellous, and M. Revel. 1982.Preferential effect ofyIFNonthesynthesisof HLAantigensand their mRNAs in human cells. Nature (London)299:833-836.
33. Zawatzky, R.,I.Gresser,E.DeMaeyer, and H. Kirchner.1982. RoleofIFNin theresistanceof C57BL/6 micetodifferentdoses of HSV type 1. J. Infect. Dis. 146:405-410.