Mutational Analysis of the 3' Open Reading
Frames
and the Splice
Junction
at
Nucleotide 3225 of Bovine Papillomavirus
Type
1PAUL L. HERMONAT ANDPETER M. HOWLEY*
Laboratory ofTumor VirusBiology,National Cancer Institute, Bethesda, Maryland 20892 Received 20 May 1987/Accepted 8 September1987
Functional analysis ofthe3'openreading frames (ORFs) of bovine papillomavirustype1 (BPV-1) has been
complicated by the organization ofthatpartofthegenome. A region between nucleotides(nt) 3173 and 3551
contains three overlapping ORFs (E2, E3,and E4), aswellas a3' splicejunctionatnt3225 which isused by
manyof the BPV-1 transcripts. Tomore clearly assign functionstospecific ORFsin this region, single-base substitutionmutationsweregenerated which introduced translational termination codons into each of the three
ORFs;afourth mutation substitutedanAwithaCatnt3223, altering the3'splice junctionconsensus sequence from AGtoCG. The E3-and the E4-specificmutantswerewildtypein their abilitiestotransformsusceptible
mouse C127 cells, to replicate as stable plasmids, and to trans-activate the E2 conditional enhancer. The E2-specific terminationmutantwasdefective for plasmid replication, transformation,and trans-activation and could not becomplemented for efficient transformation ofaflatcellline which expressed the full-length E2gene product. The splice junction mutant was defective for transformation of C127 cells and of a flat cell line expressingthefull-lengthE2geneproduct. These dataextend previous analyses of the3'ORFs andsuggestthat
a spliced E2 product is involved incellular transformation. The splice junctionmutantcould replicate asa stableplasmid, indicating that there isnoabsoluterequirementinplasmid replication foraviralgeneproduct expressed solely fromanmRNAusingthe3' splice junctionat nt3225.
The papillomaviruses are circular double-stranded DNA viruses whichcan causebenigntumorsinavariety of higher
vertebrates, including humans, and which have been
asso-ciated with carcinomas in some species. The host rangeof
these viruses is very specific, and their study has been
hampered by the lack ofanin vitrosystemfor their
propa-gation.Bovinepapillomavirustype1 (BPV-1) has provideda
usefulmodel for thestudy of the latent nonproductive phase of viral infection intransformed rodent tissue culture cells. The BPV-1 system haspermitted the analysis of viral func-tions involved in cellulartransformation, in viral transcrip-tionalregulation,and in viral DNAreplication (forareview,
seereference 11).
Sequence analysishasrevealedthat all of theopenreading
frames (ORFs) in the BPV-1 genome larger than 400 base pairs are located on one of the two DNA strands (3), and transcription studies have revealed that only one strand is
transcribedintransformed cells andinproductivelyinfected warts(1, 2, 7, 10). Viraltranscriptionin transformed cells is nonetheless complex. At least five putativepromoters have been identified by mapping the 5' ends of viral RNAs in transformedcells(2, 27). Therearemultiple speciesof RNA with complicated patterns ofsplicing (27, 28). Thegenetic mappingof BPV-1 functions involved in cellular transforma-tion and DNA plasmid replication has been difficult and complicated by the presence ofgenes whose products are
involved in the regulationoftranscription fromsomeof the viral promoters. Forinstance, the E2ORFhas been shown to encode a factor which can trans-activate conditional
enhancers located in the virallongcontrolregion (LCR) (25, 26). Afactor furthercomplicatingtheassignmentofspecific functions to specific ORFs is that there are regions of the
genome where ORFsoverlap.
*Correspondingauthor.
Inthis study, we have focusedouranalysison asegment of the BPV-1genomebetweennt3173 and3551,in which the E2, E3, andE4 ORFs overlap in each of the three potential translation frames. Just downstream from this region is the E5ORF, which partially overlapsthe3' end ofORFE2. E5 encodes oneofthetwotransforming proteins ofBPV-1 (9, 17, 22, 23, 29).
Thereare avariety ofways the ORFs within thisoverlap region can be expressed. There are two transcriptional
promoters, P2443 andP3080 located immediatelyupstreamof this region,anda spliceacceptor(SA)at nt3225 whichcan
be used to splice the 5' ORFs expressed from the LCR
promoters tothe 3' ORFsin thisregion (2, 27, 28). Several functions have been mapped to this overlap region. The productof the full E2 ORF isa trans-activator of enhancer elements located in the LCR(26). The 3' portion of the E2 ORF encodes a transcriptional repressor which can inhibit viral transformation andrepressE2-specifictrans-activation (13). Although the productfor the BPV-1 E4 ORFhas not yet been demonstrated, the E4 ORF of human papil-lomavirus type la encodes a late protein, as its product is present in high levels in warts (5). An additional function involved in cellular transformation has been suggested by the studies of Rabson et al. and mapped to this overlap region (17). Somemutantswith translation termination
link-ers (TTLs) inserted which affected the E2 and E4 ORFs could not be complemented for efficient transformation of YC-C59-3881 cells which express the full E2 gene product (17).
To further evaluate the viral functions encoded by this
portionof the BPV-1 genome,we have constructedaseries
ofORF-specificmutantsbysubstitution mutagenesis,
intro-ducing termination codons into each ofthe E2, E3, or E4
ORFs that shouldeliminatesynthesisoftheputative protein products. These ORF-specific mutants were then assessed
for their ability to transform susceptible C127 cells, to
3889 0022-538X/87/123889-07$02.00/0
CopyrightC 1987, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
3890 HERMONAT AND HOWLEY
replicate as stable plasmids, and to trans-activate the viral E2-responsive enhancer. An additional mutant was gener-ated with a basesubstitution at nt 3223,alteringthe 3' splice junction consensus sequence from AG to CG and, thereby, affecting the SA at nt 3225. This substitution creates a silent mutation in the overlapping E2 ORF and a
leucine-to-phenylalanine substitutionin the E4ORF.
MATERIALS ANDMETHODS
Cells. Mouse C127 cells(6), singlecell subclones of C127
cells, and YC-C59-3881 cells were maintained in Dulbecco modified Eagle medium with 10% fetal bovine serum, peni-cillin (100U/ml),andstreptomycin (100 ,ug/ml).The YC-C59-3881 cells have been previously described, are
non-transformed, and containaBPV-1 cDNA with anintact E2 ORF and a mutatedE5 ORF whichisexpressedfrom simian
virus40 early promoter (17).
DNA plasmid transfections. Transfections of C127 and
YC-C59-3881 cells were carried out by calcium phosphate coprecipitationaspreviously described (19) withthe
follow-ing modifications. Dishes (10 cm) of cells at approximately 80% confluence received 1to3.5,ugofplasmidDNAand 18 ,ugofherring spermcarrierDNA. This wasfollowed at 2 h
bytreatmentwith 10% glycerolinphosphate-buffered saline for1min. Thecellswerethenincubatedincomplete medium containing 4 mM sodium butyrate for approximately 12 h. When cells were selected withG418,mediumcontaining500 ,ugofthedrugper ml was added 72 hposttransfection. Cells were fixed with 10% Formalin at approximately 14 days
posttransfection and stained withmethylene blue.
Substitutionmutagenesis. Initially,the 1.3-kilobaseEcoRI
(2113)-to-KpnI (3456) fragment from the wild-type BPV-1
plasmid 142-6 was cloned into M13 mpl8 and mutagenized by the procedure used by Kunkel (12) with the following modifications. Briefly, a uracil-containing DNA
template
was grown in Escherichia coli BW313.
Oligonucleotide-primedDNAsynthesiswasallowedtoproceedat37°C for 1 handovernightat14WCwith 5 U of Klenow fragment of Poll and2 U ofligase. No uracilglycosylaseor alkalitreatment was used. The following antisense oligonucleotides were used as mismatch primers: 5' CGATCTCGAAAATCAG (SA, nt 3230 to 3215); 5' CCCAACCTAGGCCTG (E4, nt 3364 to 3350); 5' TGCGAGCTAGGACCG (E3, nt 3386 to 3372); and5' GTGCCCTACACCGGG (E2nt3455to3441).
The repairedDNA wastransfected intoJM107E. coli, and
bacteriophage wasscreened by selective hybridization with
5'-labeled oligonucleotides near thecalculated Tm. The se-quences of the base substitutions in positive phage were confirmed by the dideoxynucleotide method (18). The EcoRI-to-KpnIfragmentwasthenreligated back into the full BPV-1background and reprobedtoconfirm insertion ofthe substitution mutation. Inaddition,whenpossible, the DNAs were tested for newly created restriction sites (Fig. 1). Finally, C59-SA3225wasconstructedbyreplacing the BstEII-BstXI fragment of C59(29) with the analogous fragment from
p1039-1.
CAT assays. Chloramphenicol acetyl transferase (CAT)
assays wereperformedasdescribedpreviously (26) with the following modifications. C127 cellsweretransfected with1.0
jig
of the CAT reporterplasmid (pSV2CAT,pAjoCAT,
or p407-1), 1.0 ,ug of the plasmid being assayed for trans-activator function, and 18 ,ug of carrier DNA asdescribed above.Fiftypercentof the cell extract,prepared from 10-cmABPV-1
Mutant
1039-1 (SA)
908-1 (E4)
909-1 (E)
1038-1 (E2)
Sequence/A.A. Changes
C(3223) TTAGA
Arg
Leu-Phe TER Phe TER-Ser
A(3357) CCTZGG
Leu Ser-TER
Arg
A(3379)
CTZGC
Arg-Ser
Leu
Ser-TER
T(3448) TGZAG
Gln-*TER Cys
Ala-Val
ORF Restriction
Analysis
B
E2 new
E4 TaqI
none site
E2 new
E4 Avr II
E3 site
E2 new
E4 MaeI
E3 site
E2
E4 E3
p771-2581
2400
p772-1 p1038-1
I
I
I
MMMI I E2
L'~'-~~~~~~' ' ' '
2800
p908-1 337
rv
E4 |3173 p909-1 3526
3267 3551 3714 4010
p1039-1
P380 I
I .I . .
3200 3600 4000
none
FIG. 1. Mutant structure and genomic organization of theBPV-1E2 overlap region. (A) The recombinantplasmids are shown in the first columnontheleft.The second column indicates thenucleotide substitution effected in each particular mutant, the specific sequence number mutated in parentheses, and the resulting amino acid (A.A.) changes shown below. The crossed-out nucleotide represents the wild-type nucleotideremoved, and itssubstitution is indicated above. The resulting A.A. changes encoded for by the mutant are indicated byarrows. The ORFinwhich the A.A. substitution takes place is indicated in the third column. The fourth column indicates the presence of any newly createdrestriction sites.All mutations were transposed into the full-length BPV-1p142-6plasmid background. (B) Thegenomicorganization oftheregion. Theopenbars represent ORFs E2, E3, E4, andE5. The numbers below the ends of each ORF and M within the open bars indicate thepresenceof methionine codons of that ORF. The positions of the four mutants created by oligonucleotide-directed site-specific mutagenesisareindicatedby arrows. Two additional mutants p771-1andp772-1arealso shown atsequences 2878 and3235, respectively. Thesemutants are describedpreviously (17) and are linker insertion mutants containing a termination codon in alltranslationalframes. Over thesequencemarker at the bottom of the figure, the promoters P2443 and P3080 (2) are indicated, as well as the splice donor sited2505and the
majorSAsite
a3225-J. VIROL.
P2443
d2W5
Irl
.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.73.571.414.617.2]dishes 72 h after removal of sodium butyrate, was assayed
for CATactivity inan overnight incubation. Acetylated and
nonacetylated forms of ['4C]chloramphenicol were sepa-rated bythin-layer chromatography andquantitatedby scin-tillationcounting.
Replication analysis. C127 cellswere transfected with 0.5
pug of one of three plasmids expressing a neo-selectable
marker, 1.0 pug of the BPV-1 DNA, and 18 ,ug of carrier herring sperm DNA. The plasmids pMMTneo(302-3) and
pSV2neo have been described previously (14, 24). The plasmid p1042-1 (LCR-SVneo) was constructed by Carl
Baker by replacing the CAT sequences in p407-1
(HindIII-to-BamHIfragment) with the analogousdrugresistancegene
fragment from pSV2neo. Transfected cells were placed in
G418 forselectionat72 hafter sodium butyrate removal(8). Pooled resistantcells wereselected at3 weeks asdescribed
previously(17).
RESULTS
Construction of BPV-1mutants. Fourspecific base substi-tutionmutationsweregenerated byoligonucleotide-directed
site-specific mutagenesis. The mutations wereinitially made
intheEcoRI-to-KpnI fragment of BPV-1 and exchanged into the full-length wild-type BPV-1 plasmid as described in
Materials and Methods. Three of these mutations introduced
a termination codon specifically into the E2, E3, and E4
ORFs togenerate themutants p1038-1, p909-1, and p908-1, respectively (Fig. 1). An additional mutant, p1039-1, was
generated which alters the AG of the SA at nt 3225to CG (Fig. 1). The specific mutations generated, their locations in the BPV-1genome,andtheir effectoneachof thetranslation
framesareindicated in Fig. 1. Inaddition, the location of the
previously described TTL insertion mutants, p771-1 and p772-1, areindicated in Fig. 1 (17). Because of the
overlap-ping translation frames in this region of the viralgenome, it wasdifficult to create basesubstitution mutants resulting in
aterminationcodon inonetranslational frame thatwerealso
silent in theothertwoframes. Onlyoneof thefourmutants,
the E4ORF-specificterminationmutantp908-1,wassilent in
the other two reading frames. The E3 ORF termination
mutant, p909-1, created by changingaCat nt3379toan A,
also causes a missense mutation in E2 ORF, substituting a
serine for an arginine. The E2 ORF termination mutant,
p1038-1, generated by changinga C at nt3448 to aT, also
substitutesavalineforanalanine in theE3 ORF. The A-to-C
mutation in p1039-1 which alters the SA site at nt 3225 changes aterminationcodon upstream of the E3 ORFto a
serine residue. We feel this is unlikely to be a significant
alteration, however, since another termination signal
re-mainsintact 2codons furtherupstreamintheE3 translation frame. Inaddition, it results inamissense mutation in the E4
ORF, substitutingaphenylalanine for leucine.
Transformation of C127cellsby E2regionmutants.Eachof thesefourmutants wasassayed for its abilitytoinduce foci
on murine C127 cells after DNA transfection (Table 1). In this experiment, each ofthe BPV-1 DNAs was left intact
with the procaryotic pML2d plasmid sequences. The
wild-type complete BPV-1 plasmid (p142-6) induced approxi-mately 200 transformants per plate in thisexperiment. The E2-specific termination mutant (p1038-1) transformed at a
levelapproximately6% that of the wild type. TheE4-specific terminationmutant (p908-1)transformedatlevels
compara-ble with that of p142-6. The E3-specific mutant (p909-1)
[image:3.612.313.553.96.174.2]transformed with an efficiency slightly higherthan thewild type,andthefociappeared, onaverage, tobe slightly larger
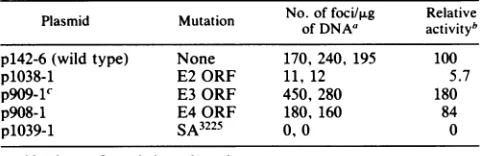
TABLE 1. Transformationof C127 cells by BPV-1 mutantplasmid DNAs
No.of
foci/p.g
Relative Plasmid Mutation of DNAa activity'p142-6 (wildtype) None 170, 240, 195 100
p1038-1 E2ORF 11, 12 5.7
p909-1 E3ORF 450, 280 180
p908-1 E4ORF 180, 160 84
p1039-1 SA3225 0,0 0
aNumbers refertoindependent plates.
bRelativetransforming activity determined from the average number of foci perplate,withp142-6defined as100.
cForthis mutant, the foci appeared, on the average, to be slightly larger thanthoseof the wild type induced byp142-6.
than those induced by the wild type. The SA mutant
(p1039-1)produced no foci. Thus, the E2 mutant behaved as manyotherdescribedE2 mutants, in thatit wassignificantly
impaired in its ability to transform mouse C127 cells when assayed in a wild-type genomicbackground (4, 9, 17). The data with the E3 and E4ORF-specificmutantsindicate that
theirgeneproducts arenotdirectly involved in
transforma-tion ofrodent cells. Whether the slight increase in
transfor-mation observed with p909-1 is aconsequenceof the
termi-nation codon inE3 or the amino acid alteration in E2 is not known atthistime. TheSA mutant
(pi039-1)
did not induce anyfociin this assay,indicatingthat theintegrity of the SA at 3225 is critical for the expression of the transformed phenotype.Todetermine whether mutation of the SA inp1039-1 has aneffectontheexpressionof the E5 gene,this mutationwas introduced into the viral cDNA, C59. The C59 cDNA
contains the BPV-1 3' ORFs intact and can efficiently transform C127 cells (28, 29). Mutational analysis of this
cDNAhas previously establishedthat the E5 ORF encodes thetransformingproteinand thatmutationsin the E2 ORFof this cDNAhadnodirect effecton transformingability (29). The structureof themRNAencodingthe E5 gene has not yet
beendefinitely identified,however. Two typesofviral RNAs canbepredicted fortheC59construct, onewith nosplicein the BPV-1 sequencesand the otherusingthe SA atnt3225 andpossiblytheviralsplicedonorat nt2505. Aswith studies
of the SA mutation in the full BPV-1 background, the
C59-SA recombinantwasmarkedly decreasedinitsabilityto induce transformation compared with the parental C59 clone. In independent plates with a subclone ofC127 cells selectedforhigh transformation susceptibility, C59plasmid produced83and 94fociper ,ugofDNA,whereas
C59-SA3225
produced 3 and 6 foci per ,ug of DNA. As expected, theC59-SA recombinantretained full E2 trans-activation activ-ity (datanotshown). This result suggests thatexpression of
theE5transforming proteininvolves anRNA(s) which uses the SA atnt 3225. A candidate for the mRNA which may
encodeE5 is the speciesderived from
P2443 using
thesplice
junctions at nt 2505 and 3225 (28). This
species
ofRNA isrelatively abundant in transformed mouse cells and in the dermal fibroblasts ofa bovine
fibropapilloma
(2). A cDNArepresentative ofthis mRNA species in an
Okayama-Berg
expression
vector wasoriginally
reported
tobe transforma-tionnegative (28)butcantransformasubclone of C127 cellsparticularly sensitive to BPV-1transformationbutat avery lowefficiency (P. L. Hermonatand P. M.
Howley,
unpub-lishedobservation).Transformation ofE2 region transformation-defective
mu-tants in YC-C59-3881 cells. Several different classes ofE2
on November 10, 2019 by guest
http://jvi.asm.org/
3892 HERMONAT AND HOWLEY
TABLE 2. Complementationof BPV-1 3' ORFmutant
DNAs inYC-C59-3881 cells
No. offoci/,ug
Plasmid Mutation Mutation of DNA Relative
position
(nt) ExptA ExptBactivity"
cvtyp142-6
(wildtype)
None 200 210 100p771-1 2878 E2ORF 56 53 27
p772-1 3235 E2, E4 ORF 0 0 0
p1038-1 3448 E2ORF 6 16 5.4
p1039-1 3223 SA3225 0 0 0
aNumbersbased on comparison of average foci perplate, with p142-6
definedas100.
mutantshave been describedon the basisof their transfor-mation characteristics (17). One class consists of mutants mapping in the amino-terminal domain of E2 which are
defective in transformation of C127 cells but which can
efficiently transformaflat nontransformedcellline YC-C59-3881 expressing E2 from an integrated C59-3881 cDNA. A second class of E2 mutants is exemplified by the mutant, p772-1,which containsaTTL,insertedatnt3235.This class
of mutant is defective for transformation of C127 cells as well as of the YC-C59-3881 cell line. Since this mutant
(p772-1) contains a linker which introduces a nonsense codon in both the E2 and E4ORFs, the defectcouldnotbe
assigned to a specific ORF. Since the E4 ORF-specific
mutantp908-1 haswild-typetransforming activity,itseemed
likely that the function required for transformation of the
YC-C59-3881 linemust be encoded by the 3' portion ofthe E2 ORF. Totestthis, we assayedthe E2 and SA mutants,
p1038-1andp1039-1, fortheirabilitiestotransform YC-C59-3881 cells. In this experiment, two mutants, p771-1 and
p772-1, which have been previously described and which contain a TTL inserted at nt 2878 and 3235, respectively,
were also assayed (Table 2). As previously shown, the 5'
E2-specific mutantp771-1was capable oftransforming YC-C59-3881 cells, whereasp772-1 with the TTLinserted at nt 3235 downstream ofthe 3' splice junction at nt 3225 was
unabletotransform.TheE2-specific mutant
(pi038-1)
trans-formedtheYC-C59-3881cellline atthesamelowefficiency
as C127 cells, indicatinga lack ofcomplementation by the BPV-1 functions constitutively expressed in the cell line,
thereby confirmingthatthisadditionalfunction requiredfor
transformation maps to the 3' portion ofthe E2 ORF. The
SAmutant
(p1039-1)
producednofoci inYC-C59-3881cells,suggesting that access to the 3' portion ofthe E2 ORF is
through the splice junction at nt 3225. We have not yet
identified which upstream coding sequences are spliced to the E2 ORF through the 3' splice junction at nt 3225 to
provide such a function. By primer extension analysis of viral RNA in C127 cells containing p1039-1, we have
con-firmedthat the base substitution at nt 3223 does result in the loss of use of the 3' splice junction at nt 3225 (data not
shown).
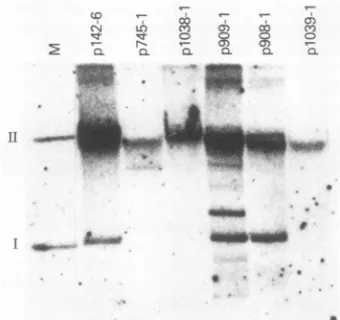
Plasmid maintenance by E2 overlap region mutants. To
determine if any of the base substitution mutants described inthis study were defective in plasmid maintenance, each of the mutant DNAs was transfected with the plasmid,
pM-MTneo(302-3), encoding a selectable drug resistance marker
into C127 cells. Total DNA was extracted from pooled
G418-resistant colonies and analyzed by Southern blot
hy-bridization 4 weeks after transfection. The ability of a mutant DNA to replicate as a stable extrachromosomal
plasmid was assayed by evaluating the cellular DNA from
the pooled drug-resistant colonies for supercoiled plasmid
DNA. In this analysis, the E3- and E4-specific mutants
(p909-1 and p908-1) were indistinguishable from the wild-type p142-6 in their abilities to replicate as stable plasmids (Fig. 2). Thus, the E3 and E4ORFsappear tobe
nonessen-tial for plasmid maintenance. The negative control for this
experimentwasp745-1,whichhasamutationatnt945in the 5' portionof the El ORF, andwas previously shown to be defective for stableplasmid replication (16). TheE2-specific
mutant plasmid, plO38-1, exhibited no form I DNA, in agreement with previous studies indicating that a gene product encoded by the full E2 ORF is required for stable plasmid maintenance (4, 17). Although noform I DNAwas
noted in thisexposureof thelanecontaining DNA from cells transfected with the SAmutant,P1039-1,asmallamountof form I DNAwasevident afterlongexposure(Fig. 2).
During these experiments, itwasnoted that cotransfection ofp1039-1 with pMMTneo resulted in fewerG418-resistant
colonies thanwith other BPV-1mutants. Toinvestigate this
observation further, increasing amounts of p1039-1 DNA,
the E2 ORF mutant DNA (plO38-1), and wild-type p142-6
DNAweretransfectedwithpMMTneo(302-3)into C127 cells andselected with G418 (Table3).Transfectionofan increas-ingamountof either p142-6 DNAorp1038-1 DNA resulted inaslight increase in the number ofG418-resistant colonies. Incontrast, increasing theamountof p1039-1DNA resulted inadecrease ofG418-resistantcolonies. Itwaspossible that the 3' splice junction mutant somehow down-regulated
expression ofthe TNS neomycin resistance gene from the MMT promoter or directly expressed a factor which was cytotoxic. We were,therefore, concerned that assessment of the ability of p1039-1 to replicate as a stable plasmid in G418-resistant cells selected withpMMTneo(302-3)couldbe flawed.Therefore, the analysis was repeatedusingplasmids in which the drug resistance gene was under the control of other promoters. In one set of experiments, we used a
cD
2 n
. _
.W
II _.
&~'
"l C) C0 0 CD
r- - 0) ~~a),
.~~~~~~
FIG. 2. Southern blot analysis oftotal cellular DNA from pooled G418-selected C127 cells cotransfected with pMMTneo(302-3) and theindicatedBPV-1 mutant DNA. DNA was digested withApaI,a no-cut enzyme for the BPV-1 plasmids. Lane M contains 250 pg each offormIand formIIp142-6plasmid marker DNA with 10 ,ug ofherringsperm DNA.Allotherlanes contain 10 ,ug of the indicated totalcellularDNAfrompooledG418-selectedcells. The blot was hybridized to nick-translated P-labeled BPV-1 DNA and washed under stringent conditions. In this experiment, the p909-1-transfected cell line contains aberrant forms of plasmid DNA. A repeatexperiment showednoaberrantforms forp909-1.p745-1 is an Elmutant,previously described, defectiveforplasmid maintenance (16).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 3. G418-resistantcolonyformation of C127 cells by dual plasmid transfection"
DNA AmtofBPV-1 No.of
DNA (,ug) colonies/plate
pMMTneo(302-3) None 125
pMMTneo(302-3) + p142-6 0.1 360
pMMTneo(302-3) + p142-6 1.0 TNTCb
pMMTneo(302-3) + p142-6 3.0 TNTC
pMMTneo(302-3) + p1038-1 0.1 200
pMMTneo(302-3) + p1038-1 1.0 400
pMMTneo(302-3) + p1038-1 3.0 420
pMMTneo(302-3) + p1039-1 0.1 100
pMMTneo(302-3) + p1039-1 1.0 60
pMMTneo(302-3) + p1039-1 3.0 34
aEachplatereceived0.5 ,ugofpMMTneo(302-3)plus the indicated amount
ofBPV-1 DNA.
b Toonumerousto count(>500).
plasmid in which the neor gene might be activated by the BPV-1 E2 gene product. The mutant p1039-1 contains an
intactE2ORFand can express E2, as assayed by its ability totrans-activate an element in the viral LCR. We, therefore,
chose a plasmid (plO42-1) in which the TnS neomycin
resistance gene is expressedfrom the simian virus 40 early promoter enhanced by the BPV-1 LCR. Selection with
p1042-1 would presumably favor selection of those cells
expressing E2. Wild-type p142-6 DNA, the El mutant
p745-1, and the 3' splice junction mutant p1039-1 were
transfected into C127 cells along with the LCR-SVneo
plasmid, and G418-resistant colonies were selected. Total
cellular DNA was prepared from pooled G418-resistant
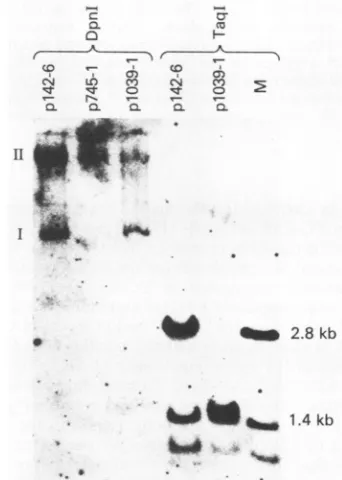
coloniesand analyzed forextrachromosomal DNA(Fig. 3). The restriction endonuclease DpnI was used to cleave the DNA before Southern blot analysis. This enzyme will only
cleave DNA methylated after replication in
dam'
bacteriaand does notcleave DNA which has replicated in
mamma-lian cells. The positive control p142-6 replicated in cells
cotransfectedwith pLCR-SVneo(1042-1)but the El replica-tion-defectivemutant,p745-1, didnot(Fig. 3). In contrast to the resultsobtained with coselectionwithpMMTneo(302-3), the SA mutant p1039-1 was present as a stable plasmid in cells selected withLCR-SVneo(1042-1). Toconfirmthat the DNA present in these drug selected cells was p1039-1, the DNAwas assayed by hybridization analysis after cleavage with TaqI(Fig. 3). The A-to-C substitution generates a new TaqI site which is in the middle ofa 2.8-kilobase fragment (Fig. 1). Analysis (Fig. 3) confirms that the BPV-1 DNA present in these cells is indeedp1039-1. Theseexperiments
werealsocarried out with pSV2-neo asaselectable marker. In theseexperiments, the
p1039-1
DNA also replicatedas a stableplasmid
(data not shown). These data, therefore, indicate that there is no absolute requirement inplasmid
replication foraviralgeneproduct which issolely
expressed
from anmRNAusingthe SA at nt3225.
LCR enhancer trans-activation byE2regionmutants.Each
of these E2
region
mutants was assessed for itsability
totrans-activate the LCR E2-responsive enhancer.
Trans-activationwas determined
measuring
CATexpression
from a BPV-1 LCR-CAT plasmid, p407-1, in which the LCR serves toenhance theenhancer-deletedsimian virus 40early
promoter(26). In these assays, C127 cellswere used, sinceexpression of the BPV-1 functions was from
homologous
viral promoters. Lysates wereprepared
72 h after sodiumbutyrate
removal,
and the reactions were allowed tocon-tinue overnight (see Materials and Methods)
(Fig.
4). In agreementwith theprevious
genetic analysis
of the BPV-1LCRtrans-activation function, only the E2-specific mutant,
p1038-1,wasdefective in thistrans-activation function, with levels approximately 8% that of the wild type. In contrast, the E3, E4, and SA mutants had trans-activation activities comparable with or slightly higher than that of the wild type.
DISCUSSION
Genetic dissection of the 3' ORFs of the BPV-1 genome has been complicated by the fact that the E2, E3, and E4 ORFs overlap and by thetranscriptional organization of the region. Our approach was to individually mutate each of the ORFsin this region as well as the SA so as to introduce silent or, hopefully, neutral changes in the other reading frames. Such mutants permit a more precise mapping of functions locatedin this region to specific ORFs.
Our data are consistent with previous published data that thetranscriptional trans-activator encoded by the full-length E2 ORF is important for both stable plasmid maintenance and cellular transformation. Its role, however, may be entirely indirect through the activation of the LCR promot-ersvia the E2-responsive elements (25, 26). The E2-specific mutant, p1038-1, was defective in plasmid maintenance,
transformation, and trans-activation functions (Table 1 and Fig. 4). Recently, Rabson et al. provided data which sug-gests that the 3' portion of the E2 ORF or overlapping
portion of the E4 ORF may encode an additional product
Q COa C
i- d
:t Irt C)_
111t
_.i
0.i's 0.
,--QC CO Cas 2.8 kb
%o%*
1.4 kbFIG. 3. Southernblotanalysisoftotal cellular DNA frompooled
G418-selected C127 cells cotransfected with LCR-SVneo(1042-1)
and the indicated BPV-1 mutant DNA. DNA was digested with DpnI and Taql where indicated. TaqI can distinguish between
p142-6 (wild type) and p1039-1 (SA) DNA by the presence ofan
additional TaqIsite inp1039atsequence 3223. Lane Mcontains 500 pgofp142-6 marker DNA with 10 ,ugofherring sperm DNA. All other lanes contain 10 ,ug ofthe indicatedtotalcellularDNAfrom
pooled G418-selectedcells.The blotwas
hybridized
asdone inFig.
1. p745-1is anEl mutant,previously described(16),defectivefor
plasmid maintenance. Although
p1039-1
wasfound tobedefective for plasmid maintenance when using MMTneo as a selectable marker (Fig. 1), when the LCR-SVneoplasmid
was used, thismutant was positiveforplasmid maintenance.kb, Kilobase.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.345.516.353.593.2]3894 HERMONAT AND HOWLEY
CM
0
0
.
e
AcCf
i:
*
C.4
*
.4l
FIG. 4. Trans-acti
withvarious BPV-1 c
p.geach of the constr
herring sperm carriei scribed in Materials
transfectedplates,an
columns are average
The relative convers
age percentconversic
nonacetylated and
spectively.
involved in cellul
analysis inYC-C59
E2-specifictermina
(plO39-1)
could beBPV-1 functions e
results strongly iml mapping5' of nt 23
expressed in the c downstream of the ORF encodes at
trans-activator, tht from P3080, and th junction at nt3225
possibility that thi
might have a dirt downstream E5-tra Thegeneproduci
for both transforn culture.Theseresu have also foundth, nonessential forph
vitro(15). The E4I identified; howeve
papillomavirus typi
dantcytoplasmicpi
the majorcapsid pi fibropapillomas sul
BPV-1encodesa lz
M Percent Relative
ORF
of the most abundant RNAspecies
transcribed fromConversion Activity the late promoter (PL) would be the E4ORF (2).
* pSV2 CAT 6.9 52.3 It has been suggested that the BPV-1 E3 ORF does not encodeaprotein, since it hasnomethionine initiation codon
pA,,J
CAT .02 0.2 and it is not highly conserved among otherpapillomaviruses.p407 1 .04 0.3 Our data does not provide any evidence to counter this
* ~ suggestion. The E3mutantp909-1 did have a slightly higher
p407-1 level of transformation than that of the
wild-type
plasmid
13.2 100.0
p142-6.
This difference may not be a result of the E3* j
J142
6 mutationbut, rather,becauseofthemissense mutationin E2 * )pp4O7-1
created in this mutant (Fig. 1) (i.e., serine replacing + 25.1 190.2 arginine). It isconceivable that this aminoacid substitutionJ
D908
-1
in the 3' domain of E2 produces aprotein
with altered* 1 p407-1 biological properties. Certainly, the E3 mutant,p909-1,was
+ 21.2 160.6 notdefective in transformationor
plasmid
replication,
per-p9091 mittingus toconcludethat the E3 ORF doesnotencodean
p4O7-1 essential protein for these processes. Whether it encodes a
+ 1.1 8.3 protein at all remains an open question.
p10381 The
SA3225
mutantp1039-1
is interesting since itdisplays
analteredphenotype,yetthe mutation doesnotintroducean amino acidchangein the E2 ORF. Weconclude, therefore, that the effects manifestedareduetothe elimination of the 3' ^p407-1 29.0
219.6
splice junction
at nt3225.By
primer
extensionanalysis
with p1039-1 a5'-end-labeledoligomer complementary to the RNA span-ning the splice from nt 2505 to 3225, we have confirmedthat ivation ofp407-1(LCR-CAT) bycotransfection the SA3225 mutant is indeed defective for this splice (data not:onstructs. C127 cells were transfected with 1 shown). It ispossiblethat othercrypticSAsareused in this ucts indicated in the first columnplus10,ugof mutant. From analyses of cDNA libraries isolated in this
r DNA. The assays were carried out as de- laboratory, it is apparent that many BPV transcripts use the and Methods. Brackets indicate redundant 3' splice junction at nt 3225 (2, 28). One might expect this Id thepercentconversion and relativeactivity mutation to have pleiotropic effects by affecting genes
cre-es of the plates indicated by the brackets. ated
by
thesplicing
ofexons at thisjunction
as well asby
,ion is based on thep4O7-1
andp142-6
aver-affecting
exons locatedentirely
upstreamordownstream of rnbeing definedas 100. Lanes CM and AcCM afclgeoslctdetrl ptemo ontemoacetylated forms of chloramphenicol, re: this junction but expressed from RNAs using this splice
junction. Atthis time, we have notdefined the mechanism
underlying the transformation defect of p1039-1. BPV-1 contains two independent transforming genes, E5 and E6
(20, 21, 29). Analysis ofacDNAlibraryhasrevealedthatan ar transformation by complementation mRNA which could encode E6 and which has slight trans-)-3881 cells(17). Inthisstudy, neither the forming activity uses the 3' splice junction at nt3225 (28).
Ltion
mutant(pi038-1)
northeSAmutant Therefore, it ispossiblethatexpression of the E6protein iscomplemented for transformation bythe lower in this mutant. This, however, is not sufficient to
expressed in YC-C59-3881 cells. These explainthedefectbecause
p1039-1
cannotbecomplemented plicate afactorcontaininga codingexon towild-type transformationlevelsbythe cDNAencodingE6 60(i.e.,
the 5' endofthecDNA segment (data not shown). It seems more likely that this mutation ell line) and the portion ofthe E2 ORF affects the level ofE5 expression. Although the RNA thatsplice junction at nt3225. Thus the E2 serves asthe messageforE5has not beendetermined, it has least three factors: the full-length E2 been suggested that a spliced RNAfrom P2443using the 3'
e E2 transcriptional repressor promoted splice junctionat3225 maybethe E5 transcript (2). Our data ie new E2 gene requiring the 3' splice with the C59-SA mutantwhich contains the E5 ORF intact idiscussed here. We cannot rule outthe but which is still impaired in its abilityto transform would s translation terminator in the E2 ORF supportthis idea. Furthermore, we have analyzed the levels ect effect on the translatability of the of E5 protein in C127 cells in whichp1039-1 was introduced
nsforminggene. by selection with the LCR-SVneo construct p1042-1 and tof the E4 ORF appears to be dispensable found only low levels of the protein (data not shown). These
iation and plasmid replication in tissue data suggest that the transformation defect of the SA mutant ltsaresimilar to those of Neary et al. who (p1039-1) may be due to altered expression of at least two at theputative product ofthe E4 ORF is genes. First, through elimination of the splice junction at nt
asmid maintenance andtransformation in 3225, it would prevent the synthesis of the spliced message gene product forBPV-1has not yet been that may normally encode E5. Secondly, it would prevent
:r,
the analogous E4 ORF for human the synthesis of a spliced E2 product which might act e 1 has been shown to encode an abun- directly or indirectly to regulate expression of the viral rotein that is expressed coordinately with transforming factors.rotein (5). Analysis ofRNAfrom bovine Itis also possible that the transformation defect is not due
bstantiates the fact that the E4 ORF of tothedirect expression of one of the transforming genes but ate orwart-specific protein,in that the 5' maybeindirectly due to the expression of a nontransforming
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.79.298.73.321.2]gene involved in the regulation of gene expression. The
ability to express the E2 trans-activator is intact in this
plasmid (Fig. 4); however, it is possible that the SA3225 mutation could affect expression of other viral products which could indirectly affect transformation. We have been unable to complement p1039-1 for transformation with a
seriesof spliced cDNAs which used the 3' SAat nt3225 (28) either singlyor in combination (datanot shown).
Although the p1039-1 mutant was defective for
transfor-mation, it was able to replicate as astable plasmid in cells
cotransfected with the LCR-SVneo plasmid (p1042-1) and with pSV2neo. Thus, unlike transformation, there is no
absoluterequirement for products of RNAs using the m3225 SA for plasmid replication functions. These data contrast
with ourresultsin which cells takingup p1039-1 DNAwere
selected by cotransfection with p302-3 expressing the neoR
genefrom the mouse metallothionein promoter. The
major-ity of the 1039-1 DNAwas integrated in C127 cells selected
with pMMTneo(302-3). A titration experiment demonstrated thatincreasing amountsof p1039-1 DNA led tofewer drug-resistant colonies. This experiment suggested that p1039-1
was somehow inhibitory to the mouse metallothionein
pro-moter. Onepossibility is that the full E2 productmayaffect
expression of the metallothionein promoter. p1039-1
ex-pressed higher levels of the E2 trans-activator than the wild-type plasmid, p142-6 (Fig. 4). It is alsoaformal
possi-bility that the p1039-1 and the pMMTneo(302-3) DNAs might
competeforafactorrequired either for efficient transcription or for BPV-1 DNA replication. Experiments to further evaluate the effect of BPV-1-encoded factors on cellular
promotersincluding the MMTpromoterarein progress.
ACKNOWLEDGMENTS
We are grateful to C. R. Schlegel and C. C. Baker for critical
commentsonthis manuscriptandto Nan Freas forpreparation of this manuscript. We are grateful to C.C.B. for providing the
LCR-SVneoplasmidwhich he constructed.
P.L.H.wassupported byNationalResearch Service Awardgrant
3F32CA07830-02S2from the National Cancer Institute.
LITERATURE CITED
1. Amtmann, E., and G. Sauer. 1982. Bovine papilloma virus
transcription: polyadenylated RNA species and assessmentof
the direction oftranscription. J. Virol. 43:59-66.
2. Baker, C. C., and P. M. Howley. 1987. Differential promoter utilizationbythe bovinepapillomavirus in transformed cellsas productivelyinfectedwarttissues. EMBO J.6:1027-1035.
3. Chen,E.Y.,P. M. Howley,A.Levinson,and P. Seeburg.1982.
The primary structure andgenetic organization of the bovine
papillomavirustype1genome. Nature(London)29:529-534.
4. DiMaio,D. 1986.Nonsense mutation inopenreadingframe E2
of bovine papillomavirus DNA.J. Virol. 57:475-480.
5. Doorbar, J.,D.Campbell,R.J.A.Grand,and P. H. Gallimore.
1986. Identification of the human papillomavirus-la E4 gene
products. EMBO J.5:355-362.
6. Dvoretzky, I.,R.Shober,S.Chattopadhy,andD. R.Lowy.1980.
A quantitative in vitro focus forming assay for bovine papil-lomavirus. Virology103:369-375.
7. Engel,L. W., C.A. Heilman, and P. M.Howley. 1983.
Tran-scriptional organization of the bovinepapillomavirus type1. J.
Virol. 47:516-528.
8. Gorman,C.M.,and B. Howard. 1983. Expressionof recombi-nant plasmids in mammalian cells is enhanced by sodium
butyrate. Nucleic Acids Res. 11:7631-7648.
9. Groff,D.E.,and W. D. Lancaster.1986.Geneticanalysisof the 3' early region transformation and replication functions of bovinepapillomavirustype 1.Virology 150:221-230.
10. Heilman,C. A.,L.Engel, D.R.Lowy, and P. M. Howley. 1982.
Virus-specific transcription in bovine papillomavirus
trans-formed mousecells. Virology119:22-34.
11. Howley,P. M.,and R. Schlegel. 1987. Papillomavirus transfor-mation.p. 141-163. In N. P. Salzmanand P. M.Howley(ed.),
ThePapovaviridae. II.Thepapillomaviruses. Plenum Publish-ingCorp., NewYork.
12. Kunkel,T. A.1985.Rapidandefficientsite-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
13. Lambert, P. F., B. A. Spalholz, and P. M. Howley. 1987. A
transcriptionalrepressorencodedbyBPV-1 shares acommon
carboxy terminal domain with the E2 transactivator. Cell 50:69-78.
14. Law, M.-F., J. C. Byrne, and P. M. Howley. 1983. A stable bovinepapillomavirus hybridplasmidthat expressesadominant selective trait. Mol. Cell. Biol. 3:2110-2115.
15. Neary, K., B. H. Horwitz, and D. DiMaio. 1987. Mutational analysisofopenreadingframeE4ofbovinepapillomavirustype 1. J. Virol.61:1248-1252.
16. Rabson,M.S.,Y.-C.Yang,and P. M.Howley. 1986.Agenetic analysis of bovine papillomavirus type-1 transformation and plasmidmaintenancefunctions. Cancer Cells 4:235-243. 17. Rabson, M. S.,C. Yee, Y.-C.Yang, and P. M.Howley. 1986.
Bovinepapillomavirustype 1 3'earlyregiontransformation and plasmidmaintenance functions. J. Virol. 60:626-634.
18. Sanger, F.,S. Nicklen,and A. R.Coulson. 1977.DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
19. Sarver,N., J.C. Byrne,and P. M.Howley. 1982. Transforma-tion and replication in mouse cells of bovine papillomavirus/ pML2 plasmid vector that can be rescued in bacteria. Proc. Natl. Acad. Sci. USA79:7147-7151.
20. Sarver,N.,M. S. Rabson,Y.-C.Yang, J.C.Byrne, andP.M. Howley. 1984. Localization and analysis of bovine
papil-lomavirustype 1transformingfunctions. J. Virol.52:377-388. 21. Schiller,J.,W.C.Vass,andD. R.Lowy.1984.Identification of
asecond transforming regionof bovine papillomavirus. Proc. Natl.Acad. Sci. USA81:7880-7884.
22. Schiller, J. T., W. C. Vass, K. H. Vousden, andD. R. Lowy. 1986. E5open readingframe of bovine papillomavirus type 1 encodesatransforminggene.J. Virol. 57:1-6.
23. Schlegel, R., M. Wade-Glass, M. S. Rabson, and Y.-C. Yang. 1986. The E5transforming gene of bovine papillomavirus
en-codes synthesis ofa small hydrophobic protein. Science233: 464466.
24. Southern, P. J. J., and P. Berg. 1982. Transformation of mammalian cells toantibiotic resistance with abacterial gene undercontrol oftheSV40earlyregionpromoter.J. Mol.Appl.
Gen. 1:327-341.
25. Spalholz, B.A., P. F. Lambert,C. L.Yee, and P. M.Howley.
1987.Bovinepapillomavirustranscriptional regulation: localiza-tionoftheE2-responsiveelementsof thelongcontrolregion.J. Virol. 61:2128-2137.
26. Spalholz, B. A., Y.-C. Yang, and P. M. Howley. 1985. Transactivation ofabovine papillomavirus
transcriptional
reg-ulatoryelementbythe E2 gene product.Cell 42:183-191. 27. Stenlund, A.,J. Zabielski, H.Ahola, J. Moreno-Lopez, and U.
Pettersson. 1985. The messenger RNAs fromthe
transforming
region ofbovinepapillomavirus type 1. J. Mol. Biol. 182:541-544.
28. Yang, Y.-C., H. Okayama, and P. M. Howley. 1985. Bovine
papillomavirus contains multiple
transforming
genes. Proc. Natl. Acad. Sci. USA82:1030-1033.29. Yang,Y.-C.,B. A. Spalholz,M. S.Rabson,and P. M.Howley.
1985. Dissociation of transforming and transactivation func-tions for bovine