Immunodeficiency Virus Type
1Long
Terminal
Repeat-Directed
Gene
Expression by
tat:
Importance
of Base
Pairing, Loop
Sequence,
and
Bulges
in
the
tat-Responsive Sequence
SOPHIE ROY,'
NEIL T.PARKIN,' CRAIG
ROSEN,2 JOSEPH ITOVITCH,'
ANDNAHUMSONENBERGl.3*
Department of Biochemistry' and McGill Cancer
Center,3 McGill University, Montreal, Canada,
H3G 1Y6,
andDepartment
of Molecular
Oncology, Roche
Institutefor
MolecularBiology,
Nutley,
NewJersey
071102Received5 September1989/Accepted 17 November 1989
Inordertoelucidate the molecular mechanisms of action of the
tat-responsive
sequence,mutationalanalysis
of thetat-responsive
sequence wascarriedout.The most criticalregion comprised
nucleotides +18 to +44andincluded the
3-nucleotide bulge
atpositions +23
to+25,
theloop
sequence, andanintact stem. Inaddition,
base
pairing
up tonucleotide
+52 wasrequired
for the fullmagnitude
of the trans-activation response.Single-nucleotide bulges
atpositions
+5 to +17 weredispensable. Analysis
of truncated andfull-length
transcripts
demonstratedthat
atranscriptional antitermination
model does notfully
account for transactivation.
Two
viralgenesencode regulatory proteins
essentialfor
human
immunodeficiency virus
type 1(HIV-1) replication:
tat(1,
4, 9, 11, 28, 33, 34) and
rev(7, 28,
32, 35).
Bothgenestrans
activate HIV-1
geneexpression. The tat-responsive
sequence
(TAR)
wasoriginally mapped
to the virallong
terminal repeat betweennucleotides -17 and +80 and thusincludes
sequences present atthe 5' end of all HIV-1
mRNAs(27). Both nucleotide substitutions
and3' deletions
were
used
todelimit the TAR region
tonucleotides -17
to +44(15), +19
to +42(16), +18
to+44
(10),
or +14 to +45(30). The
mRNAtranscribed from the TAR region
canform astablestem-and-loop
structurein vitro(22), which
has beenproposed
tohave
functional significance (2, 8, 22, 24).
Toassess
the
requirements for intact base pairing
inthe
stem structure of
the
TARregion,
wemade several sets ofmutations in predicted
duplex regions, including and
extend-ing beyond the region previously shown
tobe
important for
TARfunction (see Fig. 1A). Three
setsof
3-
or4-base
mutations
wereintroducedby site-directed mutagenesis
(41)
oneither
side of the
stemandarepredicted
todecrease
thestability of
thestemwhen
presentalone.
Themutations
weredesigned
sothat
when
individual mutations
arecombined
with
those
atthe corresponding position
onthe opposite
side
of the stem,
complete
basepairing is
restored. Thisapproach
allowsacleardistinction
between changes in primary
nucle-otidesequenceand
secondary
structure.The mutations
weresubcloned
intheHIV-1
long terminal repeat-driven
reportergene
pU3RIII (34). The trans-activation activity,
measured
after transienttransfection by the DEAE-dextran method
(12),
wasdefined
asthe chloramphenicol
acetyltransferase
(CAT) activity
obtained inanestablished
HeLa-tat cell line
(26) compared with that in the parental cell line. Results,
expressed
asthe
averageof
two tofive
independent
exper-iments,
aresummarized
in Table
1.Mutations
predicted todisrupt base pairing
resulted in
amoderate (TARI, TARII),
severe
(TART'),
ornear-complete
(TARIII,
TARITI',
TARI,III, and TARI',III') loss of trans-activation activity,
*Correspondingauthor.
[image:1.612.318.557.451.699.2]with the
exception of the TARII' mutant, which had
aslight
stimulatory effect. These results indicate that nucleotides in
TAR
outside of the boundaries
previously
described
(10,
15,
16,
30)
areimportant
for full
activity.
Possible
explanations
for this
discrepancy
arediscussed below. In all cases,
compensatory mutations
predicted
to restorebase
pairing
also restored TAR function
towild-type
levels. These
setsof
mutations
emphasize
the
requirement
for
secondary
struc-turein
the TAR
region
for
transactivation, especially
in
mutantTARI,III,I',III',
which has 16
nucleotide changes
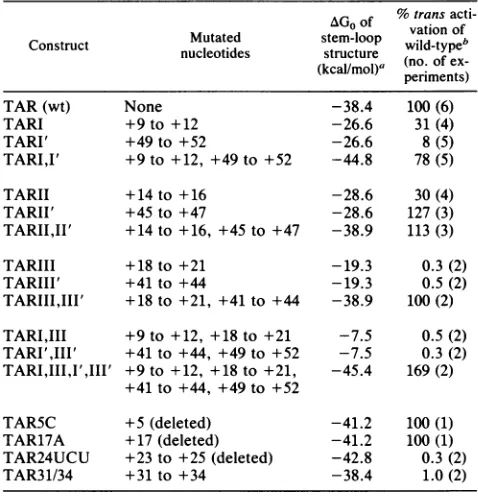
TABLE 1. Effects ofTARmutationson transactivation %trans
acti-Mutated stem-loop vation of
Construct nucleotides structure wild-type
(kcalImol)a
(no:
ofex-periments)
TAR(wt) None -38.4 100(6)
TARI +9to +12 -26.6 31(4)
TARI' +49to +52 -26.6 8(5)
TARI,I' +9to +12, +49to +52 -44.8 78(5)
TARII +14to+16 -28.6 30(4)
TARII' +45to+47 -28.6 127(3)
TARII,II' +14to+16, +45to+47 -38.9 113(3)
TARIII +18to+21 -19.3 0.3(2)
TARIII' +41 to+44 -19.3 0.5(2)
TARIII,III' +18to+21, +41to+44 -38.9 100(2)
TARI,III
+9to+12, +18to+21 -7.5 0.5(2) TARI',III' +41 to+44, +49to+52 -7.5 0.3(2) TARI,III,I',III' +9to+12, +18to+21, -45.4 169(2)+41 to +44, +49to +52
TAR5C +5 (deleted) -41.2 100(1)
TAR17A +17 (deleted) -41.2 100(1)
TAR24UCU +23to+25(deleted) -42.8 0.3(2)
TAR31/34 +31to+34 -38.4 1.0(2)
aCalculatedaccordingtotheenergyrulesofSalser(29).
bThe averagefoldtransactivationwas750.
1402
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES
1403
0
GU
G+30-C A
C G
G
C AU
CUG
C;-Ai
TARIII
TARI,III
TARIITARI
U
G
CG
cU
A U AG
CG
Cm7OpppO
C .. .TARIII'
TARII'
TARI'
COMPENSATORY
MUTANTS:
TARI,I'
TA12RII,II'
TARtIII,III'
TAItI,III,I',III'
TAR31/34
,
+30-C A
C O
a
C AU
TAR24UCU
I0
C(A)
uA
U-40
G C
.20-A U
C O
TAR17A
(C
0(A)
Q CA U U A U G OC
o
U
-.50o10-
U AC O
U O
C O
TAR5C
©UA
(A)
U
AOC OC
m7OpppO
C...
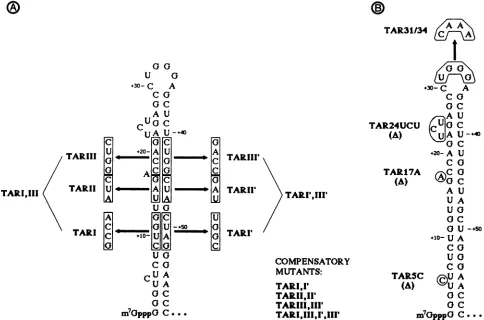
FIG. 1. (A) Secondarystructure of the TAR region and stem mutants. Nucleotides shown in boxes were changed to those indicated by site-directed mutagenesis. Mutationswere combined by further mutagenesis(TARI,III
andTARI',III')
or by subcloning, using theSacl site at+34(all other doublemutants). The namesofdouble mutants which restore base pairing in the stem areindicated (bottom, right). (B) Bulge andloopmutants. Bulged nucleotidesatpositions+5, +17, and +23 to +25 were deleted, and the sequenceofthe loop atpositions +31 to +34 was mutated to the indicated sequence.but
canstill be as well trans activated
by tat as the wild-type
construct.One
possible explanation
for the
discrepancy
between
this
report
and
others is the type of
transfection protocol used.
All other reports
investigating
the boundaries
of the TAR
region
(10, 15, 16, 30) have used
cotransfection of the
reporter gene
with a tat-expressing plasmid, often in systems
which
allow efficient replication of the transfected plasmids
(15,
30),
whereas
we
have
used transfection of the reporter
gene
into
cells
stably
expressing
tat.To
determine whether
there were major differences between the two protocols, we
cotransfected
the
wild-type
pU3RIII or mutant TARI,
TARI',
or
TARI,T'
with
a
simian virus
40-based
tatexpres-sion
vector,
in
COS
cells. In
this
system,
the
mutations
had
little
or noeffect
(data
notshown).
However, in
cotrans-fected
HeLa cells, in which the plasmid remains in low copy
number,
there were no
qualitative differences in the results
obtained
compared
with those described
above,
asevi-denced
by
the
inhibitory
effect of
TARIT'
and the
restoration
of
activity
in TARI,I'
(data
notshown).
Thus, it is
likely
that
the copy
number of the transfected plasmids has effects
onthe
phenotypes of some TAR mutants. Although unlikely,
wecannot
exclude the
possibility that the mutant
pheno-types
of TARI and TARI'
arespecies
dependent.
A
second
setof mutations in the
TARregion
wasmade
todetermine the
importance
of structural features other than
base
pairing
in the
stem(Fig.
1B). Several studies have
shown that
bulged
nucleotides in other RNA
stem-and-loop
structures are
required
for
protein binding (3,
14,
25, 39).
To
determine whether such
nucleotides
areimportant
for TAR
function, bulged
nucleotides
atpositions +5, +17,
and +23
to+25
weredeleted. In
addition,
the sequence
of the
loop
was
mutated
(positions
+31
to+34).
trans-Activation
activ-ity
of these mutants,
asassessed
by
transfection in the
HeLa-tat
cell
line,
is
summarized in Table
1.TAR5C
and
TAR17A, in which
single-nucleotide bulges
atpositions
+5
and
+17,
respectively,
weredeleted,
had
noeffect
on transactivation.
However, both TAR24UCU and
TAR31/34
had
very
low
activities
(less than
5% of
wild type) in HeLa-tat
cells. These results
indicate
that the
3-nucleotide
bulge
nearthe top
of
the
stemis also critical for TAR
function.
We
arecurrently
performing
additional
mutagenesis
in order
tobetter define the
requirement
for the 3-nucleotide
bulge, by
changing
the sequence
of these
nucleotides and
assaying
for
transactivation. In
this context, it is
important
that
conser-vation of the UCU
bulge
is
notabsolute among
different
isolates of
HIV-1,
since
HIV-lBRU
contains UUU instead
(38). Also, the
TARregion
of
HIV-2,
which
contains
loop
sequences
identical
tothose
in HIV-1
(CUGGG),
has
abulge
of
2uridine residues in
oneof the
proposed
stem-loop
structures
(8, 13).
Such differences may
reflect
amodulation
of the
ability
of these viruses
torespond
to transactivation
VOL.64, 1990
y
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.65.549.76.396.2]I0
il4
-180
*It -160
-4--a
tat:
- + -+
- +wt I?
I'
i I II'Ii.I1'
III III' 111I11W 1.111 ,1 1,Hlindll Pvuil EcoRl
...I IJ
-1
mII
CAT probeR
-.1~~PTW1~
24 31/34
ucu
eamHl
0~
[image:3.612.67.555.72.380.2]150Mt
FIG. 2.
Quantitative
RNaseprotection
analysis
of TAR-CAT mRNA in transfected cells. Cellsweretransfected and 20p.g
ofcytoplasmic
RNAwas
analyzed
asdescribed in thetext. Theunprotected probe migrated
as adoubletatapproximately
200nucleotides(MI- ).
LaneM,
DNAsize markers(end-labeled
pBR322
cutwithMspl);
odd-numberedlanes,
transfectedHeLacells(-);
even-numberedlanes,
transfected HeLa-tatcells(+).
Arrowsindicate theposition
of theprotected probe
fragment (150 nucleotides).
Anautoradiogram
froma3-day
exposure is shown. Theplasmid
usedfor transfection is indicated below theautoradiogram.
Theposition
of theprobe
relativetothestructureof the TAR-CAT mRNA is alsodiagrammed.
wt, Wildtype.or may
reflect reduced structural
orprimary
sequencere-quirements
within the
bulges.
To
define the level
atwhich
TARmutations affect
geneexpression,
weanalyzed
CAT
RNAlevels in
transfected
HeLa and HeLa-tat
cells,
using
anRNaseprotection
assay(40).
Cytoplasmic
RNA wasisolated
by
using
theNonidet
P-40
lysis
procedure
(20),
including
aDNase Idigestion
step.
The
probe
wasderived from the
coding
portion
of the CAT
gene(Hindlll
toPvull
fragment,
150 base
pairs)
and
sub-cloned in antisense orientation in
pSP65
(21)
and
is
expected
to
protect 150 nucleotides of the CAT
mRNAfrom
RNasedigestion (Fig.
2).
The 12P-labeledprobe (approximate
spe-cific
activity,
6 x108
cpm4[Lg)
wasgel purified
on a5%
polyacrylamide-urea gel,
eluted
overnight
at40C
in 0.5 Mammonium
acetate-i
mMEDTA-0.1% sodium
dodecyl
sul-fate,
extracted with
phenol-chloroform,
and ethanol
precip-itated.
Theoptimal hybridization
temperature for
the CATprobe
wasdetermined
tobe
550C.
Representative
results
areshown
inFig.
2(note
thatthe
undigested probe
and theprotected
CAT
band
aredoublets,
due tosomeheterogene-ity
in
theprobe).
Allprotection
assays werestandardizedby
including
aprobe
for
theendogenous
-y-actin
mRNA(6)
at the sametime
as theCAT
probe;
the
signal
for
-y-actin
wasobserved
to berelatively
constant betweensamples
(with
one
exception;
seebelow).
We were notable
todetect
theCAT
mRNAin the
absenceof
tat(Fig. 2,
odd-numberedlanes),
presumably
because of
the very low levelof
expres-sion. The
signal
obtained from
HeLa-tatcells transfected
with
mostplasmids (Fig.
2,
even-numbered
lanes)
roughly
(within
afactor of
two)
paralleled
thelevel of
transactiva-tion,
with
theexception
of TARII
(Table 1).
For
this
construct, the CAT
mRNA wasbarely
detectable
(Fig.
2,
lane
10)
although
it had
only
amoderate
inhibitory
effect
ontrans
activation. This
discrepancy
maybe accounted
for
by
a
reduction in the
amountof
RNAused in the
assay,since
the
signal
for
-y-actin
wasweaker in this
sample
than in
others
(data
notshown).
We
also examined the effect of
TARmutations
onthe
reported
antitermination
activity
of
tat(17, 30,
36).
This
activity
canbeassayed
by
comparing
the
cytoplasmic
levels
of short
RNAspecies
of 59to65nucleotides,
suggested
tobeprematurely
terminated
transcripts,
with those of
full-length
or
read-through
transcripts.
Figure
3shows
theresults of
an RNaseprotection
assay,using
cytoplasmic
RNAfrom
HeLa or HeLa-tatcells transfected with
wild-type
TAR(lanes
1 and2)
ormutants TARI(lanes
3 and4),
TARI'(lanes
5and
6),
andTARIJI'
(lanes
7and8)
andcorresponding
antisense
probes spanning
thefirst
85nucleotides
of
the HIV-1un-translated
region (PvuII-HindIII fragment
of
pU3RIII),
in-cluding
the TAR element. Theoptimal hybridization
temper-ature,for both short
andfull-length
transcripts,
wasfound
tobe
50 to550C.
Alow level of attenuated
transcripts
wasdetectable in
HeLa cellstransfected
withpU3RIII (wild
type;
Fig.
3,
lane1)
orwith
the doublemutantTARIJI'
(Fig.
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES 1405
1 2 3 4 5 6 7 8
fulllength-_
85nt
short |
59-65nt
aW
M
-147
1-123
9,-90
aO
-7-67
46
.f
wo -6 ....
tat: - + - + - + - +
wt I 1
Pvull
5
RHindill
t
[image:4.612.72.296.68.411.2]I X ----
---n
FIG. 3. RNase protection analysis of 5' ends of TAR-CAT
mRNAs. Therelative abundance offull-length (85-nucleotide) and
short (59-to65-nucleotide) transcripts in cytoplasmic RNA
prepa-rations from HeLa (-) or HeLa-tat (+) cells transfected with pU3RIII (wildtype [wt])ormutantTARI, TARI',orTARI,I'was
determined by using homologous RNA probes spanning the tran-scriptional start site (see diagram at bottom). A 10-,ug amount of cytoplasmicRNAwasusedin theassay. Protectedfragmentswere
resolvedonan8% polyacrylamide-urea gel;anautoradiogram from
a7-day exposure is shown. Lane M, MspI-digested, end-labeled pBR322. The migration position of the undigested probe is indicated
3,
lane7). However,
these short RNAspecies
were not detected in cells transfected withTARI
(Fig. 3,
lane3)
orTARI'
(Fig. 3,
lane5), indicating
thatsecondary
structurein thisregion
isimportant
for thegeneration
and/orstability
of the shorttranscripts.
In HeLa-tatcells, full-length
tran-scripts,
whose relative amountsparallel
those obtainedby
using
the CATprobe,
arereadily
seen(Fig. 3,
even-num-bered
lanes).
It isimportant, however,
that there was anoverall increase in the total amount of RNA detected in HeLa-tat cells
compared
with that in HeLacells,
with apreferential up-regulation
in theexpression
offull-length
transcripts. Therefore,
unless there weremajor
differences inthestability of
the shorttranscripts
in HeLacells,
relative to that of thefull-length transcripts
in HeLa-tatcells,
the antitermination mechanism does not account for the full effect oftatin thissystem.Since short
transcripts
were not detected in cells trans-fected with mutantTARI
orTARI'
in the absence oftatandwere
barely detectable in the presence of tat, it is possible
that these
mutations affect the termination site, the
tran-scription
initiationefficiency,
orthe
stability of terminated
RNAs.One
interesting possibility, for which evidence is
being sought,
is based onthe
recentobservation that
induc-tion of beta interferon
transcription by double-stranded
RNA
is mediated in part
by
NF-KB(19, 37). If the
double-stranded
natureof the
TARregion could also induce NF-KB
activity,
perhaps via the double-stranded RNA-dependent
p68 kinase (5, 31, 37), then this mechanism may contribute
toHIV
long
terminal
repeat-directed
transcription,
since
the
long
terminal repeat
contains
twoNF-KB-binding
sites
(23).
The
mutantRNAs
TARI
orTARI', however, would be
expected
tobe less potent
activators of NF-KB and therefore
tobe transcribed less
efficiently (Fig.
2and
3).
This
study, designed
toanalyze
the structural
require-ments
for
TARfunction,
suggests
that
responsiveness
to tatrequires
the
bulge
atpositions
+23
to+25, the
loop
se-quence, and proper
base
pairing.
The
effects of
mutations
in
nucleotides +9
to +12and +49
to+52,
although
not asdramatic
asthose
within the upper
portion
of the stem,
arenevertheless
significant.
The
importance of
the
lower
region
may
nothave
been
previously
realized because it appears
tobe
dependent
onthe
transfection
protocol
and/or the cell
type
used
toexamine the
trans-activation response.
Since the
original
submission of the
manuscript, Laspia
etal.
reported
similar results with respect
tothe
contribution
of
antitermination
to transactivation, using
arecombinant
adenovirus vector(18).
Thefirsttwoauthors contributedequallytothis work. WethankBeverlyAkerman fortechnical assistance.
This research was supported by grants from The American Foundation for AIDS Research (grant 000946-7-RGR:AmFAR/ Drexel Burnham Lambert researchgrant)and TheCancer Research
Society
of MontrealtoN.S. N.S. isarecipient
ofanMRCScientist Award of the Medical Research Council of Canada. S.R. is arecipient ofaFondsde RechercheenSantedu
Quebec
studentship,
and N.P. isa
recipient
ofa Medical Research Council of Canadastudentship.
LITERATURE CITED
1. Arya,S. K., C. Guo, S.F. Josephs, andF. Wong-Staal. 1985. trans-activator gene ofhuman
T-lymphotropic
virus type III(HTLV-III).
Science229:69-73.2. Berkhout, B., R. H. Silverman, and K.-T. Jeang. 1989. TAT trans-activates the human
immunodeficiency
virusthrough
anascentRNAtarget. Cell 59:273-282.
3. Christiansen, J., S. R.
Douthwaite,
A.Chistensen,
and R.Gar-rett. 1985. Does
unpaired
adenosine-66fromhelixII ofEsche-richiacoli5SRNAbindto
protein
L18? EMBO J.4:1019-1024. 4. Dayton, A. I., J. G.Sodroski,
C. A.Rosen,
W. C.Goh,
andW. A.Haseltine.1986. Thetrans-activator geneofthe human T
cell
lymphotropic
virus type III isrequired
forreplication.
Cell 44:941-947.5. Edery, I.,R.Petryshyn,and N.
Sonenberg.
1989. Activation of double-stranded RNAdependent
kinase(dsl)
by
the TARregion
of HIV-1 mRNA: anovel translational control mecha-nism.Cell56:303-312.6. Enoch, T., K. Zinn, and T. Maniatis. 1986. Activation of the human
1-interferon
generequires
aninterferon-inducible factor. Mol. Cell. Biol. 6:801-810.7. Feinberg, M.B.,R. F.Jarrett,A.
Aldovini,
R.C.Gallo,
andF. Wong-Staal.1986.HTLV-IIIexpression
andproduction
involvecomplex
regulation
at the levels ofsplicing
andtranslation of viral RNA. Cell 46:807-817.8. Feng, S., and E. C. Holland. 1988. HIV-1 tat trans-activation requires the
loop
sequence within tar. Nature(London)
334: 165-168.9. Fisher, A. G., M. B. Feinberg, S. F.
Josephs,
M. E.Harper,
VOL. 64,1990
on November 10, 2019 by guest
http://jvi.asm.org/
LTR TATA and TARregion sequences required for
transcrip-tional regulation. EMBO J. 8:765-778.
11. Goh, W. C., C. Rosen, J. Sodroski, D. D. Ho, and W. A.
Haseltine.1986. Identification ofaprotein encoded by thetrans
activator gene tatIll of human T-cell lymphotropic retrovirus typeIII. J.Virol. 59:181-184.
12. Grosschedl, R., and D. Baltimore. 1985. Cell-typespecificity of
immunoglobulin geneexpression is regulated byat least three
DNAsequenceelements. Cell 41:885-897.
13. Guyader,M., M. Emerman, P. Sonigo, F. Clavel, L. Montagnier,
and M.Alizon. 1987. Genome organization and trans-activation ofthe human immunodeficiency virustype2. Nature(London) 326:662-669.
14. Haile, D. J., M. W. Hentze, T. A. Rouault, J. B. Harford,and
R. D. Klausner. 1989. Regulation of interaction of the iron-responsive element binding protein with iron-iron-responsive RNA
elements. Mol. Cell. Biol. 9:5055-5061.
15. Hauber,J., and B. R. Cullen. 1988. Mutational analysis of the trans-activation-responsive region of the human immunodefi-ciency virustypeIlong terminalrepeat. J. Virol. 62:673-679. 16. Jakobovits,A., D. H. Smith, E. B. Jakobovits, and D. J. Capon.
1988. A discrete element 3' of human immunodeficiency virus 1 (HIV-1) and HIV-2 mRNA initiation sites mediates transcrip-tional activation by an HIV trans activator. Mol. Cell. Biol. 8:2555-2561.
17. Kao, S.-Y.,A. F. Calman, P. A. Luciw, and B. M. Peterlin. 1987.
Anti-termination of transcription within the long terminalrepeat of HIV-1by tatgeneproduct. Nature (London) 330:489-493.
18. Laspia,M. F., A. P. Rice, and M. B. Mathews. 1989. HIV-1tat
protein increases transcriptional initiation and stabilizes elonga-tion. Cell59:283-292.
19. Lenardo, M.J.,C.-M.Fan, T. Maniatis, and D. Baltimore. 1989. The involvement of NF-KB in 13-interferon gene regulation reveals its role as awidely inducible mediator of signal trans-duction. Cell 57:287-294.
20. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory,
Cold Spring Harbor, N.Y.
21. Melton, D. A., P. A. Krieg, M. R. Rebagliati, T. Maniatis, K.
Zinn, and M. R. Green. 1984. Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids containing a bacteriophage SP6 promoter. Nucleic Acids Res.12:7035-7056.
22. Muesing, M. A., D. H. Smith,and D. J.Capon.1987. Regulation ofmRNA accumulation by a human immunodeficiency virus
trans-activator protein. Cell 48:691-701.
23. Nabel, G., and D.Baltimore. 1987. An inducible transcription factoractivates expression of human immunodeficiency virus in
Tcells. Nature (London) 326:711-713.
24. Okamoto, T., and F. Wong-Staal. 1986. Demonstration of virus-specific transcriptional activator(s) in cells infected with HTLV-IIIbyaninvitrocell-free system. Cell 47:29-35.
25. Peattie, D. A., S.Douthwaite, R. A. Garrett, and H. F. Noller.
Virol. 57:379-384.
27. Rosen, C. A., J. G. Sodroski, and W. A. Haseltine. 1985. The location ofcis-acting regulatory sequences in the human T cell lymphotropic virus type III (HTLV-III/LAV) long terminal repeat. Cell 41:813-823.
28. Sadaie, M. R., T. Betner, and F. Wong-Staal. 1988. Site-directed mutagenesis of two trans-regulatory genes (tat-III, trs) of HIV-1.Science 239:910-913.
29. Salser, W. 1977. Globin mRNA sequences: analysis of base-pairing and evolutionary implications. Cold Spring Harbor Symp. Quant. Biol. 42:985-1002.
30. Selby, M. J., E. S. Bain, P. Luciw, and B. M. Peterlin. 1989. Structure,sequence, andpositionof thestem-loopintar deter-minetranscriptional elongation by tatthrough the HIV-1long terminal repeat.Genes Dev.3:547-558.
31. SenGupta, D. N., and R. H. Silverman. 1989. Activation of interferon-regulated, dsRNA-dependentenzymesbyhuman im-munodeficiency virus-1 leader RNA. Nucleic Acids Res. 17: 969-978.
32. Sodroski, J., W. C. Goh, C.A.Rosen, A.Dayton,E.Terwilliger, and W. A. Haseltine. 1986. A secondpost-transcriptional
trans-activator generequiredforHTLV-III replication.Nature (Lon-don) 321:412-417.
33. Sodroski, J., R. Patarca, C. Rosen, F. Wong-Staal, and W. Haseltine. 1985. Location ofthe trans-activating regionon the genome of human T-cell lymphotropic virus type III. Science 229:74-77.
34. Sodroski, J., C. Rosen, F. Wong-Staal, S. Z. Salahuddin, M. Popovic, S. Arya, R. C. Gallo, and W. A. Haseltine. 1985. trans-actingtranscriptional regulationof human T-cell leukemia virus type IIIlong terminal repeat. Science 227:171-173. 35. Terwilliger, E., R. Burghoff, R. Sia, J.Sodroski,W.Haseltine,
andC. Rosen. 1988.The art geneproductofhuman immunode-ficiencyvirus isrequired for replication.J. Virol.62:655-658. 36. Toohey, M. G., and K. A. Jones. 1989. In vitroformation of
short RNApolymerase IItranscripts thatterminate within the HIV-1 and HIV-2 promoter-proximal downstream regions. Genes Dev. 3:265-282.
37. Visvanathan, K. V., and S. Goodburn. 1989. Double-stranded RNAactivatesbinding of NF-KB to aninducibleelement in the human ,-interferonpromoter. EMBO J. 8:1129-1138.
38. Wain-Hobson, S., S. P. Sonigo, 0. Danos, S. Cole, and M. Alizon. 1985. Nucleotide sequence of the AIDS virus, LAV. Cell40:9-17.
39. Wu, H.-N., and 0. C. Uhlenbeck. 1987. Role ofa bulged A residue in a specific RNA protein interaction. Biochemistry 26:8221-8227.
40. Zinn, K., D. DiMaio, and T. Maniatis. 1983. Identification of two distinctregulatory regions adjacent to the human
P-interferon
gene.Cell34:865-879.
41. Zoller, M. J., and M. Smith. 1982. Oligonucleotide-directed mutagenesis using M13-derivedvectors: anefficientandgeneral procedure for theproductionofpointmutations in anyfragment of DNA.Nucleic Acids Res. 10:6487-6500.