Copyright © 2005, American Society for Microbiology. All Rights Reserved.
The B30.2(SPRY) Domain of the Retroviral Restriction Factor TRIM5
␣
Exhibits Lineage-Specific Length and Sequence Variation in Primates
Byeongwoon Song,
1Bert Gold,
2Colm O’hUigin,
3Hassan Javanbakht,
1Xing Li,
1Matthew Stremlau,
1Cheryl Winkler,
3Michael Dean,
2and Joseph Sodroski
1,4*
Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute, Department of Pathology, Division of AIDS,
Harvard Medical School, Boston, Massachusetts 021151; Laboratory of Genomic Diversity, National Cancer Institute,
Frederick, Maryland 21702-12012; SAIC-Frederick, Frederick, Maryland 21702-12013; and Department of
Immunology and Infectious Diseases, Harvard School of Public Health, Boston, Massachusetts 021154
Received 23 September 2004/Accepted 18 December 2004
Tripartite motif (TRIM) proteins are composed of RING, B-box 2, and coiled coil domains. Some TRIM pro-teins, such as TRIM5␣, also possess a carboxy-terminal B30.2(SPRY) domain and localize to cytoplasmic bodies. TRIM5␣has recently been shown to mediate innate intracellular resistance to retroviruses, an activity depen-dent on the integrity of the B30.2 domain, in particular primate species. An examination of the sequences of several TRIM proteins related to TRIM5 revealed the existence of four variable regions (v1, v2, v3, and v4) in the B30.2 domain. Species-specific variation in TRIM5␣was analyzed by amplifying, cloning, and sequencing nonhuman primateTRIM5orthologs. Lineage-specific expansion and sequential duplication occurred in the TRIM5␣B30.2 v1 region in Old World primates and in v3 in New World monkeys. We observed substitution patterns indicative of selection bordering these particular B30.2 domain variable elements. These results sug-gest that occasional, complex changes were incorporated into the TRIM5␣ B30.2 domain at discrete time points during the evolution of primates. Some of these time points correspond to periods during which pri-mates were exposed to retroviral infections, based on the appearance of particular endogenous retroviruses in primate genomes. The results are consistent with a role for TRIM5␣in innate immunity against retroviruses.
Following entry, retroviruses must negotiate a series of pro-cesses to establish a permanent infection of the host cell. These include uncoating of the viral core, reverse transcription, nu-clear access, and integration of the viral DNA into the host genome (1, 11, 60). Early postentry restrictions to retrovirus infection can determine tropism at the species level. Infection by N-tropic murine leukemia virus, for example, is inefficient in most human cells and in certain cell lines from African green monkeys (5, 53). Human immunodeficiency virus type 1 (HIV-1) encounters a postentry block in Old World monkeys, whereas simian immunodeficiency virus (SIVmac) is blocked in
most New World monkey cells (17, 18, 42). These species-specific restrictions share several features. First, the block oc-curs prior to or concurrent with reverse transcription, which occurs in the cytoplasm of the host cell. At most, low levels of early reverse transcripts are made in restricted cells (9, 17, 29, 42). Second, the viral determinant of susceptibility to the block is the capsid protein (9, 14, 22, 32, 33, 53). Other capsid-binding proteins, such as cyclophilin A in the case of HIV-1, can modify the degree of restriction (33, 34, 49, 55). Third, restriction is mediated by dominant host factors, the activity of which can be titrated by the introduction of virus-like particles containing proteolytically processed cap-sid proteins of the restricted viruses (4, 6, 9, 13, 29, 33, 46, 54).
These observations suggested a model in which host restric-tion factors interact, directly or indirectly, with the viral capsid
and prevent its progression along the infection pathway. A genetic screen identified a major restriction factor in monkey cells that acts on HIV-1, and to a lesser extent, on SIVmac(47).
The factor, TRIM5␣rh, was selected from a cDNA library
pre-pared from primary rhesus monkey lung fibroblasts. TRIM5␣rh
was shown to be sufficient to confer potent resistance to HIV-1 infection on otherwise susceptible cells. Moreover, TRIM5␣rh
was necessary for maintenance of the block to the early phase of HIV-1 infection in Old World monkey cells, as demon-strated by interference with the expression of the endogenous
TRIM5ortholog in these cells (47). HIV-1 infection in cells
expressing TRIM5␣rhwas blocked at the earliest stage of
re-verse transcription (47). Cells expressing TRIM5␣rhexhibited
a partial inhibition of SIVmacinfection but were as susceptible
as control cells to infection by Moloney murine leukemia virus vectors. Humans express a protein, TRIM5␣hu, that is 87%
identical in amino acid sequence to the rhesus monkey protein TRIM5␣rh(47). Even when expressed at comparable levels,
TRIM5␣huwas less potent in suppressing HIV-1 and SIVmac
infections than TRIM5␣rh (47). Recently, TRIM5␣hu was
shown to be responsible for the postentry restriction of N-tropic murine leukemia virus (N-MLV) in human cells (15, 20, 34, 63). TRIM5␣rhwas much less effective than TRIM5␣huat
blocking this murine leukemia virus. Thus, TRIM5␣hupotently
restricts N-MLV, specifies an intermediate level of resistance to HIV-1, and does not affect SIVmac infection. In contrast,
TRIM5␣rhpotently restricts HIV-1 and exhibits a modest
in-hibition of SIVmacand N-MLV infections. The TRIM5␣
pro-tein from African green monkeys, TRIM5␣agm, has recently
been shown to inhibit N-MLV, HIV-1, and SIVmacinfections
(15, 20, 43, 63). These observations underscore the importance * Corresponding author. Mailing address: Dana-Farber Cancer
In-stitute, 44 Binney Street, JFB 824, Boston, MA 02115. Phone: (617) 632-3371. Fax: (671) 632-4338. E-mail: [email protected] .edu.
6111
on November 8, 2019 by guest
http://jvi.asm.org/
of species-specific variations in TRIM5 orthologs for their abil-ity to restrict infections by particular retroviruses.
More than 50 genes encode tripartite motif (TRIM) proteins (36). The tripartite motif includes a RING domain, a B-box 2 domain, and a coiled coil (cc) domain; TRIM proteins have also been called RBCC proteins. Some TRIM proteins, includ-ing TRIM5, contain a C-terminal B30.2 or SPRY domain (16, 36). Differential splicing of the TRIM5 primary transcript gives rise to the expression of several isoforms of the protein product (36). The TRIM5␣isoform is the largest product (493 amino acid residues in humans) and contains the B30.2/SPRY do-main. The other TRIM5 isoforms (␥and␦are the best sub-stantiated of these) lack an intact B30.2/SPRY domain. The TRIM5␥rhisoform does not inhibit HIV-1 or SIVmacinfection
(47). In fact, TRIM5␥rh has been shown to exhibit a weak
dominant-negative activity, repressing the ability of wild-type TRIM5␣rh to inhibit HIV-1 infection (47). Thus, the B30.2
domain is critical for the ability of TRIM5␣to mediate anti-retroviral effects.
A common feature of TRIM proteins is the ability to assem-ble into large complexes in the cytoplasm or nucleus of the cell (36). The function of these cytoplasmic and nuclear bodies is largely unknown. The demonstration of a potent, specific an-tiretroviral activity for TRIM5␣raised the possibility that the major function of some TRIM proteins is the establishment of innate immunity to infectious agents. The localization of TRIM5 to cytoplasmic bodies (36, 62) is consistent with its ability to block retroviral infection shortly after entry of the viral capsid into the cytoplasm of the host cell.
Studies of endogenous retroviral sequences have indicated that vertebrates, including humans, have been exposed to ret-roviruses for many millions of years (2, 12, 19, 21, 25, 26, 28, 35, 44, 45, 56, 58, 59, 61). This long history of host-retrovirus coevolution might favor the selection of particular TRIM5␣ proteins that effectively suppress lethal retrovirus infections. Since many viruses preferentially infect particular host species, the selection of molecules involved in antiviral immunity often occurs in a lineage-specific manner. In this report, we charac-terize the TRIM5␣ proteins of several primate species and provide evidence of lineage-specific variation in particular B30.2 domain elements.
MATERIALS AND METHODS
Cells forTRIM5cDNA synthesis.SQMK-FP (Pindak) cells from a Bolivian squirrel monkey (Saimiri boliviensis boliviensis), Vero cells from an African green monkey (Cercopithecus aethiops pygerythrus), OMK cells from an owl monkey (Aotus trivirgatus), and human HeLa cells were obtained from the American Type Culture Collection. Primary rhesus lung fibroblasts were derived from a rhesus macaque (Macaca mulatta) (18). The Coriell Institute for Medical Re-search (Camden, N.J.) supplied the following cells: AG05352 cells from a black-handed spider monkey (Ateles geoffroyi), AG05308 cells from a red-chested mustached tamarin (Saguinus labiatus), AG06116 cells from a patas monkey (red guenon) (Erythrocebus patas), AG06209 cells from a Sumatran orangutan (Pongo pygmaeus abelii), AG05251 cells from a Western lowland gorilla (Gorilla gorilla), and GM03448 cells from a chimpanzee (Pan troglodytes verus).
Samples for DNA analysis.Nonhuman primate DNAs from a previously described collection of samples (2, 3) included two independent samples from squirrel monkeys (B263 [female] and B26) (Saimiri sciureus), samples from two baboon species (B21 [Papio cynocephalus] and B856 and B1542 [Papio anubis]), a patas monkey sample (B1530) (Erythrocebus patas), a grivet sample (B1532 [female]) (Cercopithecus sabaeus), two sooty mangabey samples (B400 [female] and B859) (Cercocebus atys), a gelada sample (B853) (Theropithecus gelada), two colobus samples (B130 [Colobus guereza] and B1527 [Colobus guereza caudatus]),
samples from two species of langur (B131 [female] [Presbytis obscurus] and B401 [male] [Presbytis senex]), samples from two species of gibbon (B23, B1533, and B837 [Hylobates lar] and B128 [Hylobates concolor]), and samples from gorillas (B27 [female], B461, and B1535 [female]) (Gorilla gorilla). Additional samples of patas monkey (Erythrocebus patas), white-cheeked gibbon (Hylobates concolor), and gorilla (Gorilla gorilla) DNA were obtained from the core collection of the Laboratory of Genomic Diversity, National Cancer Institute.
TRIM5cDNA cloning and sequencing.First- and second-strand cDNA syn-thesis was performed by use of a cDNA synsyn-thesis kit (Clontech), using RNAs prepared from cells derived from an African green monkey, a rhesus macaque, a Bolivian squirrel monkey, an owl monkey, a patas monkey, a tamarin, a spider monkey, an orangutan, a gorilla, and a chimpanzee (see above). Human and primateTRIM5cDNAs encoding TRIM5␣were amplified with the primers TRIMf2 (5⬘-GCGGAATTCGCCATGGCTTCTGGAATCCTGGTT-3⬘) and TRIMr2 (5⬘-GCGATCGATGCCTCAAGAGCTTGGTGAGCACAG-3⬘).
Amplification was carried out by use of a Clontech Advantage PCR kit, with thermocycling of the reactions at 95°C for 30 s, followed by 30 cycles of 95°C for
30 s, 55°C for 1 min, and 68°C for 3 min. Amplified cDNAs were inserted into the pCR-BluntII-TOPO plasmid (Invitrogen) and sequenced by use of the following primers: M13f (⫺20), GTAAAACGACGGCCAGT; M13r (⫺21), AACAGCT ATGACCATG; TRIMf3, GGAAGCTGACATCAGAGA; TRIMf4, GATAAG AGACAAGTGAGC; TRIMr3, TCTACCTCCCAGTAATG; and TRIMr4, TC CTTCTCCAGGTTTTGC.
PCR amplification of humanTRIM5exon 8 from genomic DNA.Thirty nano-grams of individual DNAs from the primates described above was amplified with 5 U of AmpliTaqGold enzyme in 3.5 mM MgCl2and 330M nucleoside
triphosphates, using 200 nanomoles each of the following primers containing an M13 forward or reverse tag: TRIM5Ex8SeqF, 5⬘-GTAAAACGACGGCCAGT TCCCTTAGCTGACCTGTTAATTT-3⬘; and TRIM5Ex8SeqR, 5⬘-GGAAACA GCTATGACCATGGCTGTACAGAAGGGGCTGAG-3⬘. The following cy-cling conditions were used: an initial 94°C, 5-min activation of AmpliTaqGold, followed by 20 cycles of 94°C for 20 seconds, 54°C for 20 seconds, and 68°C for 1 min and then an additional 20 cycles of 94°C for 20 seconds, 65°C for 20 seconds, and 68°C for 1 min, a final extension of 68°C for 7 min, and a hold overnight, if necessary, at 4°C.
DNA sequencing. Amplicons were treated with exonuclease I and shrimp alkaline phosphatase and subjected to Big Dye cycle sequencing in the presence of either an M13 forward or reverse primer tag (m13 forward, 5⬘-GTAAAACG ACGGCCAGT-3⬘; m13 reverse, 5⬘-GGAAACAGCTATGACCATG-3⬘). Se-quenced reactions were cleaned with Sephadex G-50, dried on a thermocycler (95°C without caps for 20 min), and resuspended in undiluted formamide (Ap-plied Biosystems) in preparation for capillary electrophoresis on an ABI 3730 HT apparatus. Subsequent to sequencing, forward and trace files were examined with Mutation Explorer software (Softgenetics) and sequences were visually determined.
Phylogenetic analysis.The data sets for the complete TRIM5␣protein and the B30.2(SPRY) domain were examined separately. Predicted amino acid se-quences were compiled and aligned for subsequent phylogenetic analyses by ClustalX (51) and were verified visually. Phylogenetic reconstruction was per-formed with MEGA, version 2.0, software (23) using the following methods: minimum evolution, neighbor joining, and maximum parsimony. A bootstrap analysis using 1,000 iterations was performed with each method (38).
Nonsynonymous/synonymous variation.The nonsynonymous/synonymous (Ka/Ks) ratios at various codon positions for pairwise comparisons ofTRIM5
cDNAs were calculated by the method of Li (24). TheKa/Ksratio was estimated
as a rolling average for a window of 150 codons, with the center of the window being moved codon-by-codon to produce a plot showing local variations in the degree of sequence conservation.
Nucleotide sequence accession numbers.The complete primateTRIM5coding sequences have been deposited in the GenBank database under accession num-bers AY740612 to AY740621. The primateTRIM5nucleotide sequences have been deposited in the GenBank database under accession numbers AY710295 to AY710304.
RESULTS
Variation among TRIM proteins related to TRIM5.TRIM proteins first appeared with the metazoans and dramatically expanded in number during vertebrate evolution (36). Thus, currentTRIMgenes presumably arose from a small number of ancestral genes. To gain insight into the conserved and variable elements of TRIM proteins, we aligned the primary amino acid
on November 8, 2019 by guest
http://jvi.asm.org/
sequences of several TRIM proteins. The TRIM proteins most closely related to TRIM5␣hu(as ascertained by BLAST
anal-ysis) were selected for study. Nearly all of these TRIM proteins contain a B30.2 domain, and many have been shown to local-ize, at least in part, to the cytoplasm of the cell (36). Several of these TRIM proteins are found in cytoplasmic bodies, as is TRIM5 (36).
The predicted amino acid sequences of TRIM proteins re-lated to TRIM5␣huare aligned in Fig. 1. The sequences of
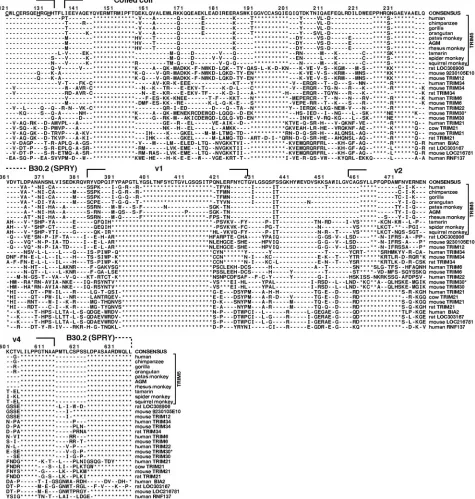
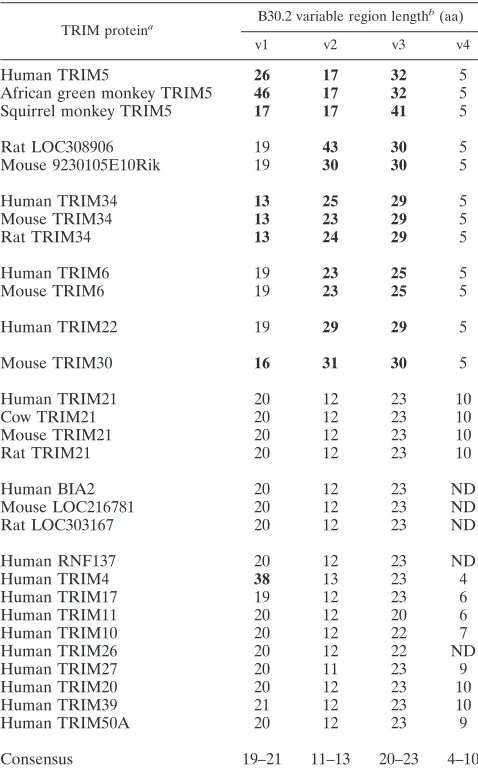
sev-eral TRIM5 orthologs from nonhuman primates that were determined in this study were included in the alignment (see below). A moderate degree of variation was observed in all of the domains of these selected TRIM proteins (Fig. 1). The existence of four regions within the B30.2 domain that exhib-ited substantial variation in length as well as extensive amino acid differences among the TRIM proteins was particularly noteworthy. We designated these variable regions v1, v2, v3, and v4 (Fig. 1). Because of the modest level of amino acid vari-ation among the TRIM B30.2 domains, the boundaries of these variable regions are somewhat arbitrary. We chose reasonably conserved sets of boundary residues to eliminate any ambiguity in defining the length of a particular variable region. This al-lowed us to examine the lengths of the B30.2 domain variable regions in different TRIM proteins, including those which are less closely related to TRIM5 (Table 1). Based on length variations in the B30.2 v1, v2, and v3 regions, two groups of TRIM proteins can be discerned. For one group, consisting of TRIM21, BIA2, mouse LOC216781, rat LOC303167, RNF137, TRIM17, TRIM11, TRIM10, TRIM26, TRIM27, TRIM20, TRIM39, and TRIM50A, each B30.2 variable region conforms to a narrow range of lengths. The v1 regions of these TRIM proteins are 19 to 21 residues long, the v2 regions are 11 to 13 residues long, and the v3 regions are 20 to 23 residues long. An examination of the available sequences of TRIM10, TRIM11, TRIM20, TRIM26, TRIM27, TRIM39, and TRIM50A from different mammalian species indicated that the B30.2 domain sequences and the variable region lengths are relatively well conserved (data not shown). The B30.2 domain v1, v2, and v3 regions of TRIM21 and the v3 region of TRIM17 exhibit more amino acid variability between humans and rodents (data not shown). Nonetheless, the lengths of the TRIM17 and TRIM21 variable regions are similar in humans and rodents (Table 1 and data not shown). Thus, one group of TRIM proteins ex-hibits a narrow range of B30.2 variable region lengths and, in many cases, minimal variation between human and rodent or-thologs. Based on the sequences of the TRIM proteins in this group, consensus TRIM B30.2 domain variable region lengths can be defined as 19 to 21 residues for v1, 11 to 13 residues for v2, and 20 to 23 residues for v3.
For the second group of TRIM proteins, the lengths of the B30.2 variable regions differ from those found for the group described above. This second group includes TRIM4, TRIM5, TRIM6, TRIM22, TRIM34, rat LOC308906, and mouse 9230105E10RiK. Compared with the consensus B30.2 variable loop lengths, TRIM4 exhibits a long v1 region, and TRIM6, TRIM22, TRIM30, TRIM34, rat LOC308906, and mouse 9230105E10RiK exhibit long v2 and v3 regions. Relative to the consensus, the primate TRIM5␣proteins exhibit differences in the lengths of v1, v2, and v3. It is noteworthy that a group of related TRIM proteins (TRIM5, TRIM6, TRIM22, TRIM30,
TRIM34, rat LOC308906, and mouse 9230105E10RiK) all share the property of increased B30.2 variable region lengths com-pared to the consensus. The humanTRIM5,TRIM6,TRIM22,
andTRIM34genes are located in a gene cluster at 11p15.4.
In rodents, TRIM6 and TRIM34 are adjacent to the rat LOC308903 and mouse 9230105E10RiK genes. This clustering may indicate recent paralogous relationships among these
TRIMgenes. In the available TRIM6 and TRIM34 sequences from other mammals, the B30.2 variable region lengths are similar to those found for the human proteins (Table 1), even though considerable amino acid variation is observed in these regions (data not shown). The human TRIM4 and TRIM22
genes do not have orthologs in the rodent genome, and con-versely,TRIM30is not found in the human genome (data not shown). Thus, in most cases for which data are available, the B30.2 variable region lengths are preserved in TRIM proteins from different species. TRIM5␣and the proteins encoded by the rat LOC308906 and mouse 9230105E10RiK genes are ex-ceptions (Table 1; see below).
The v4 variable regions of the B30.2 domains of the TRIM proteins examined are short, varying from 4 to 10 residues in length. The v4 variable region is five residues long in all of the TRIM proteins closely related to TRIM5 (Table 1).
Interspecies variation in the TRIM5␣ protein.Variations among TRIM5␣ proteins in primates affect the efficacy of restriction of particular retroviruses (15, 20, 34, 43, 47, 63). Ancient retroviral infections encountered by the members of specific primate lineages may have exerted selective pressure on the TRIM5␣ structure. To investigate this possibility, we cloned and sequenced theTRIM5cDNAs encoding the com-plete TRIM5␣proteins of several primate species. These in-cluded apes (chimpanzee, gorilla, and orangutan), Old World monkeys (rhesus macaque, African green monkey, and patas monkey), and New World monkeys (squirrel monkey, tamarin, and spider monkey). To ensure that these sequences were representative, we determined the sequences of severalTRIM5
cDNAs from individual humans, rhesus macaques, African green monkeys, and squirrel monkeys. The variation among the predicted TRIM5␣proteins of individuals within a species was⬍2.5% (data not shown).
We also examined the mouse and rat genomes for potential
TRIM5orthologs. The rodent proteins most closely related to
TRIM5 are encoded byTRIM12 (mouse), 9230105E10RiK (mouse), and LOC308906 (rat). Reciprocally, TRIM5 is the human protein most closely related to the protein prod-ucts of these rodent genes (data not shown). TRIM12 and 9230105E10RiK are essentially identical in their 5⬘ portions and have been assembled at adjacent loci on mouse chromo-some 7. It is possible that the current assembly will be revised and that TRIM12 and 9230105E10RiK are in fact the same gene. It is also possible that these genes are paralogs that arose from a duplication event after the divergence of mice and rats. An examination of current drafts of the chicken and dog ge-nomes failed to identify candidateTRIM5orthologs.
An alignment of the predicted amino acid sequences of the TRIM5-related proteins of rodents and several primates is shown in Fig. 1. The degree of variation among the TRIM5␣ -related proteins of these different species was larger than that observed within the individuals of a species (data not shown). The full-length protein product of the 9230105E10RiK gene is
on November 8, 2019 by guest
http://jvi.asm.org/
only 49% identical to human TRIM5␣. This is lower than the 80 and 67% identities seen when the human and rodent ver-sions of the TRIM6 and TRIM34 proteins, respectively, are compared. In fact, the degree of sequence identity between the
human TRIM5␣ and rodent 9230105E10RiK proteins is not higher than the similarity among TRIM5 and other TRIM pro-teins encoded by the humanTRIMcluster located at 11p15.4. Whether the mouse 9230105E10 and rat LOC308906 genes FIG. 1. Alignment of amino acid sequences of TRIM proteins related to TRIM5. The predicted amino acid sequences of several primate TRIM5␣proteins and other TRIM proteins related to TRIM5 were aligned. Those TRIM proteins located near the top of the alignment are more closely related to TRIM5␣than those near the bottom. Identity is indicated by dashes, and gaps are indicated by asterisks. A consensus sequence consisting of the most common residues at each position is shown at the top of the alignment. Residues important for zinc binding and folding of the RING and B-box 2 domains are underlined in the consensus sequence. The boundaries of the TRIM domains and the B30.2 variable regions (v1 to v4) are indicated. TRIM30* refers to the TRIM30-like protein encoded by mouse LOC209387. The arrow denotes the site of the sequence triplication in the v3 region of spider monkeys, which is not included in its entirety in this alignment. The complete sequence of the spider monkey v3 region is shown in Fig. 3.
on November 8, 2019 by guest
http://jvi.asm.org/
representTRIM5orthologs will be addressed elsewhere. At a minimum, the rodent TRIM5␣-related proteins exhibit sub-stantial differences from TRIM5␣hu. Thus, our analysis of
in-terspecies variation among TRIM5␣proteins will focus mainly on primate sequences.
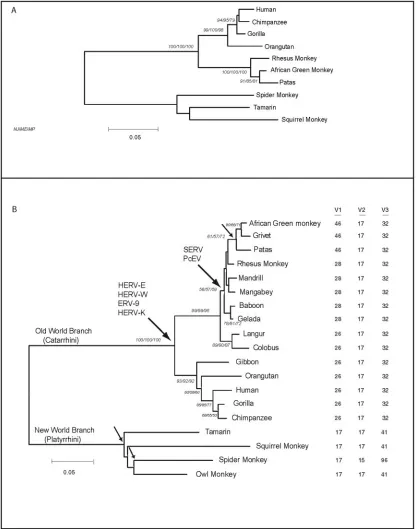
Full-lengthTRIM5cDNA sequences from 10 primate spe-cies were used to construct a phylogenetic tree (Fig. 2A). Bootstrap simulation using neighbor joining, minimum evolu-tion, and maximum parsimony algorithms all resolved the New Word/Old World dichotomy and the great ape/Old World monkey dichotomy with a high confidence. Patas monkeys
were differentiable from African green monkeys and rhesus monkeys in comparisons of the full-length TRIM5␣sequences; comparisons of the complete TRIM5␣sequences for this par-ticular branch differed somewhat from comparisons between rhesus macaques and related primates when only the B30.2 regions were compared (see below).
[image:5.585.55.530.74.573.2]Interspecies variation in TRIM5-like proteins was observed in all four of the defined TRIM5␣ domains, with the most variation occurring in the B30.2 domain (Table 2). To assess B30.2 domain variation more thoroughly, we amplified DNA fragments corresponding toTRIM5exon 8, which encodes the FIG. 1—Continued.
on November 8, 2019 by guest
http://jvi.asm.org/
TRIM5␣B30.2 domain, from several nonhuman primates and then sequenced the fragments. Attempts to amplifyTRIM5
exon 8 sequences from two prosimian species, lemurs and galagoes, were unsuccessful (data not shown). As expected from our analysis of TRIM proteins, the observed variation within the B30.2 domains of primate TRIM5␣proteins was not uniform, being concentrated within the v1, v2, and v3 variable regions defined above (Fig. 1). In the TRIM5␣proteins of the primates examined, all three of these variable B30.2 regions differed in length from the consensus TRIM variable region lengths (Fig. 2B). With the sole exception of the spider monkey TRIM5␣protein, in which v2 is 15 residues long, the primate TRIM5␣v2 regions are 17 residues in length. Thus, although the v2 region exhibits a high level of substitution of individual amino acids among primate TRIM5␣proteins, the length of
this region is well conserved. In contrast, a striking lineage-specific variation in the lengths of the v1 and v3 variable re-gions of the primate TRIM5␣proteins was observed (Fig. 2B). The v1 regions of the New World monkeys are 17 residues long, slightly shorter than the consensus v1 length. In contrast, the v1 regions of all of the Old World (catarrhine) primates are longer than that of the consensus. Since a v1 length of 26 res-idues is found in all hominoids and in the Colobinae subfamily of Old World monkeys, this may represent the v1 length of the TRIM5␣proteins of the shared catarrhine ancestor. In the Cer-copithecinae subfamily of Old World monkeys, the v1 length is expanded to 28 residues, and in the African green monkey/ grivet/patas monkey lineage, it is expanded to 46 residues. The 46-residue v1 region of TRIM5␣from these monkeys contains two tandem 20-residue repeats (Fig. 3, top). These are perfect repeats in some individual African green monkeys, but not in others (data not shown). Thus, the TRIM5␣B30.2 v1 region appears to have undergone some expansion in all Old World primates. In certain lineages, this expansion is dramatic and involves tandem sequence duplication.
The B30.2 v3 region is 32 residues long in the TRIM5␣ proteins of all Old World (catarrhine) primates. This is longer than the consensus v3 length and likely represents the v3 length of the TRIM5␣protein of the shared catarrhine ancestor. The TRIM5␣B30.2 v3 region is longer in the New World (platyr-rhine) monkeys than in the Old World monkeys. The v3 region is 41 residues long in all of the New World monkey TRIM5␣ sequences examined, except for that of the spider monkey. The B30.2 v3 region in spider monkeys has expanded to 96 residues in length. This remarkable expansion results from the presence of three imperfect tandem repeats, of 30, 31, and 24 residues (Fig. 3, bottom). Thus, the TRIM5␣B30.2 v3 region appears to have expanded in New World (platyrrhine) monkeys. In at least one lineage, that of spider monkeys, the expansion is dra-matic and involves tandem sequence duplication.
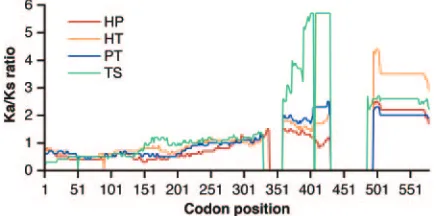
Analysis of nonsynonymous/synonymous variation inTRIM5
genes from different species. Analyses of nonsynonymous/ synonymous variations (Ka/Ksratios) can provide insight into selection for or against a change in the coding capacity of a gene (24). However, this approach cannot accommodate se-quences that exhibit large insertions or deletions (indels) rel-ative to one another. Therefore, for our comparison ofTRIM5
sequences from different primates, the indels involving the v1 and v3 regions of the B30.2 domain were excluded from the analysis. TheKa/Ksratios at various codon positions were cal-culated for pairwise comparisons of TRIM5 cDNAs from a hominoid (human), an Old World monkey (patas monkey), and two New World monkeys (tamarin and spider monkey), providing several phylogenetically independent paired com-parisons (Fig. 4). For the sequences encoding the N-terminal half of TRIM5␣(the RING, B-box 2, and coiled coil domains), theKa/Ksratio was generally⬍1, indicating selection against amino acid changes. For the sequences encoding the B30.2 domain, theKa/Ksratio increased dramatically, reaching values above 5 in some cases. Such high Ka/Ks ratios are strongly suggestive of selectively driven diversity in the B30.2 domain.
DISCUSSION
[image:6.585.43.282.89.476.2]The functions of most TRIM proteins are not understood (36). The TRIM genes arose in bilaterans and expanded in TABLE 1. Lengths of B30.2 domain variable regions
in TRIM proteins
TRIM proteina B30.2 variable region length b(aa)
v1 v2 v3 v4
Human TRIM5 26 17 32 5
African green monkey TRIM5 46 17 32 5
Squirrel monkey TRIM5 17 17 41 5
Rat LOC308906 19 43 30 5
Mouse 9230105E10Rik 19 30 30 5
Human TRIM34 13 25 29 5
Mouse TRIM34 13 23 29 5
Rat TRIM34 13 24 29 5
Human TRIM6 19 23 25 5
Mouse TRIM6 19 23 25 5
Human TRIM22 19 29 29 5
Mouse TRIM30 16 31 30 5
Human TRIM21 20 12 23 10
Cow TRIM21 20 12 23 10
Mouse TRIM21 20 12 23 10
Rat TRIM21 20 12 23 10
Human BIA2 20 12 23 ND
Mouse LOC216781 20 12 23 ND
Rat LOC303167 20 12 23 ND
Human RNF137 20 12 23 ND
Human TRIM4 38 13 23 4
Human TRIM17 19 12 23 6
Human TRIM11 20 12 20 6
Human TRIM10 20 12 22 7
Human TRIM26 20 12 22 ND
Human TRIM27 20 11 23 9
Human TRIM20 20 12 23 10
Human TRIM39 21 12 23 10
Human TRIM50A 20 12 23 9
Consensus 19–21 11–13 20–23 4–10
a
Proteins from different species that are likely to be orthologous are placed in a common section. TRIM proteins more closely related to TRIM5␣occupy the top half of the table.
b
The lengths of the B30.2 variable regions, defined as shown in Fig. 1, are given in amino acid residues. Numbers in bold indicate variable loop lengths outside the range of the consensus length. ND, not determined due to ambiguity in the alignment of sequences flanking the variable regions.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 2. Phylogeny of primate TRIM5␣sequences. (A) Predicted amino acid sequences of the complete TRIM5␣proteins of the indicated primate species were used to construct phylogenetic trees with Clustal X (51). Phylogenetic reconstruction was performed by the neighbor-joining, minimum evolution, and maximum parsimony methods (38). A bootstrap analysis with 1,000 iterations was performed by using each method, and bootstrap values of⬎50% are indicated in the tree topology in the order: neighbor joining/minimum evolution/maximum parsimony. Branch distances (nucleotide substitutions) are indicated on the scale bar. (B) Nucleotide sequences ofTRIM5exon 8 were used to predict the sequence of the TRIM5␣B30.2 domain. The predicted B30.2 domain amino acid sequences were used to construct phylogenetic trees as described for panel A. The lengths of the B30.2 domain variable regions are listed to the right of the phylogenetic tree. Increases in the lengths of the variable regions with respect to those of the putative ancestral protein are highlighted. The TRIM5␣v4 region is five residues long in all of the species examined and therefore is not listed. The five arrows indicate points in the TRIM5␣phylogeny when expansions in specific B30.2 variable regions appeared. The two larger arrows indicate points during primate phylogeny associated with the appearance of the indicated endogenous retroviruses in the genome. Note that the owl monkey exon 8 sequences are probably vestigial, as theTRIM5gene in this species has been targeted by a retrotrans-poson and encodes a TRIM5-cyclophilin A chimera (30, 39).
on November 8, 2019 by guest
http://jvi.asm.org/
number during vertebrate evolution in response to as yet un-known selective forces. Several observations are consistent with the possibility that some of these proteins contribute to the es-tablishment of innate intracellular defense systems against in-fectious agents. The TRIM5␣proteins of rhesus monkeys and humans block infections by HIV-1 and murine leukemia vi-ruses, respectively (15, 20, 34, 47, 63). The African green mon-key TRIM5␣protein exhibits activity against a diverse group of retroviruses (15, 20, 43, 63). Some antiviral activity has also been reported for other TRIM family members (7, 8, 27, 57), al-though this activity lacks the potency and degree of specificity observed for TRIM5␣against particular retroviruses. Finally, someTRIMgenes are inducible by interferon (8, 31, 37, 52), sug-gesting a link to other elements of the innate immune system. The above considerations raise the possibility that infec-tious agents influenced the evolution of certain TRIM pro-teins. Length variation in particular segments of different TRIM B30.2 domains were examined as one potential indica-tor of such selection. Our observations are consistent with a parsimonious model in which ancestral TRIM proteins pos-sessed B30.2 domains with relatively short variable regions. In such a model, the ancestral TRIM B30.2 domain possessed a v1 region of 19 to 21 amino acids, a v2 region of 11 to 13 residues, and a v3 region of 20 to 23 residues, the variable region lengths found in many examples of extant TRIM pro-teins. For some cytoplasmic TRIM proteins such as TRIM21, amino acid variation, but not length variation, in the B30.2 variable regions occurred during the evolution of rodents and primates. For TRIM4 and the TRIM proteins encoded by
[image:8.585.41.540.82.180.2]the human 11p15.4 cluster (TRIM5, TRIM6, TRIM22, and TRIM34), expansion of the length of one or more B30.2 vari-able regions apparently occurred at various times in mam-malian evolution. For TRIM6 and TRIM34, the mutational events that caused the increased lengths of the v2 and v3 regions likely occurred before the divergence of rodents and primates. Once fixed, these variable region lengths were ap-parently maintained within narrow limits for at least 90 million years. In contrast, the evolution of different B30.2 variable region lengths in TRIM5␣continued during the diversification of primates (see below). SinceTRIM4andTRIM22are found in humans but not rodents, the emergence or loss of these genes must have occurred after the rodent-primate divergence. The TRIM5␣ B30.2 domain exhibits length variations in particular primate lineages. The shorter versions of the TRIM5␣ B30.2 domain v1, v2, and v3 regions among the primate species examined exhibit lengths of 17 residues, 15 to 17 residues, and 30 to 32 residues, respectively. These v2 and v3 regions are longer than the corresponding regions of the hypothetical an-cestral TRIM protein, suggesting that some expansion of these regions may have occurred in a TRIM5␣ancestor prior to the diversification of primates. Notably, expansion of the v2 and v3 regions characterizes all of the TRIM proteins encoded by the human 11p15.4 cluster; it seems plausible that these related TRIM proteins share a common ancestor that exhibited v2 and v3 regions which were longer than those of the putative ances-tral TRIM protein. Concomitant with the evolution of pri-mates, the TRIM5␣B30.2 domain apparently underwent epi-sodic expansions of either the v1 or v3 variable region in a
[image:8.585.80.503.579.686.2]FIG. 3. Repetitive elements in the TRIM5␣B30.2 domain variable regions. The tandem repeats within the v1 region of African green monkey TRIM5␣and the v3 region of spider monkey TRIM5␣were aligned with the corresponding sequences of human TRIM5␣. Identical amino acids between/among the repeats are shown in green; identical amino acids between the monkey and human TRIM5␣proteins are shown in red in the human sequence. Dashes represent gaps in the alignment.
TABLE 2. Interspecies variation in TRIM5␣-related proteins
Proteins for comparison
% aa difference in domaina
Overall
RING B-box 2 Coiled coil B30.2 (SPRY)
Human TRIM5␣vs chimpanzee TRIM5␣ 0 3 3 2 2
Human TRIM5␣vs rhesus monkey TRIM5␣ 13 3 12 17 13
Human TRIM5␣vs African green monkey TRIM5␣ 20 0 11 24 15
Human TRIM5␣vs squirrel monkey TRIM5␣ 16 12 28 45 32
Human TRIM5␣vs spider monkey TRIM5␣ 18 12 28 56 39
Human TRIM5␣vs mouse 9230105E10RiK 50 24 59 70 57
Human TRIM5␣vs rat LOC308906 41 21 56 72 58
aThe percentages of amino acid residues that differ between the TRIM5␣-related proteins of the indicated species within each domain and within the complete
protein are shown. The percentages were calculated as follows: number of different residues/total number of residues compared⫻100%. Insertions in one sequence were counted as differences and also contributed to the total number of residues compared.
on November 8, 2019 by guest
http://jvi.asm.org/
lineage-specific fashion. These v1 or v3 expansions occurred at discrete points in the TRIM5␣phylogeny (Fig. 2B, arrows). The evolution of a v1 length of 26 residues apparently occurred at the root of the Old World (catarrhine) branch of primates. A further expansion of v1 to 28 residues occurred with the evolution of the subfamily Cercopithecinae, beginning 9 to 13 million years ago. A final v1 expansion to 46 residues evolved with the African green and patas monkeys.
In the New World (platyrrhine) branch of monkeys, the TRIM5␣B30.2 domains evolved long v3 regions. At the root of this lineage, a v3 expansion to 41 residues occurred. A further expansion to 96 residues occurred in spider monkeys. Remark-ably, the longest B30.2 variable regions (v1 in African green monkeys and patas monkeys and v3 in spider monkeys) contain tandem duplications. Doublets, defined as short duplications of between 25 and 100 bp, are common in the genomes of mammals and other eukaryotes (50). Tandem doublets, such as those that encode the B30.2 variable region duplications ob-served in our study, tend to have arisen recently in evolution (50). It has been speculated that double-stranded breaks that are filled in and nonhomologously recombined might be the origins of adjacent doublets (50). Whatever the mechanism by which they arose, the resulting long variable regions in TRIM5␣were preserved and became prevalent in the respec-tive monkey species. TRIM5␣proteins with long B30.2 vari-able regions possibly provide better protection against retrovi-ral infections. Indeed, the TRIM5␣proteins of African green monkeys and spider monkeys exhibit antiviral activities against a diverse group of retroviruses (15, 20, 43, 63); it is possible that the tandem duplications in the B30.2 variable regions contribute to this breadth.
Standard approaches to assess the selection of genes (e.g., comparisons of the ratios of nonsynonymous/synonymous changes) typically ignore insertions and deletions that result in length variations. An analysis of Ka/Ks ratios was useful for assessing the evolution of TRIM5␣ sequences outside the B30.2 v1 and v3 variable regions. Pairwise comparisons of
TRIM5cDNAs from four species, representing hominoids, Old
World monkeys, and New World monkeys, revealed a similar
pattern. For the 5⬘ portion of the TRIM5cDNA, which en-codes the RING, B-box 2, and coiled coil domains of TRIM5␣, theKa/Ksratios were generally⬍1. This indicates that purify-ing selection has operated on these TRIM5␣domains to pre-serve the amino acid sequences and, presumably, function. This conclusion is consistent with the conservation of amino acid residues known to be important for the integrity of these domains in the TRIM5␣proteins from the different primate species examined (Fig. 1). The demonstrated functional activ-ity of TRIM5␣proteins derived from different primate species in restricting particular retroviruses (15, 20, 43, 47, 63) also supports the conservation of TRIM5␣’s function during pri-mate evolution. Even in owl monkeys, in which a retrotrans-position has resulted in a loss of the ability to encode a com-plete TRIM5␣ protein and instead a TRIM5␣-cyclophilin A fusion protein is produced, the capacity to restrict some retro-viruses is preserved (30, 39).
In contrast to the results for the 5⬘portion ofTRIM5, the
Ka/Ksratios were⬎1 for the 3⬘end ofTRIM5, which encodes the B30.2 domain. Thus, even when the v1 and v3 variable regions, which exhibit significant variations in individual amino acid residues as well as in their lengths, were excluded from consideration, the analysis strongly suggested that diversity within primate TRIM5␣B30.2 domains has been driven by se-lection. In summary, evolutionary forces have operated to pre-serve the amino acids in the N-terminal portion and to diver-sify the amino acids in the B30.2 domain of primate TRIM5␣ proteins.
Although our analysis ofKa/Ksratios suggests that selection has influenced theTRIM5gene segments flanking the indels, the selective nature of the inserts themselves (i.e., whether an increased length is deleterious, neutral, or positively selected) requires further interpretation. The probability of fixation of an insert will be less than, equal to, or more than its creation rate through mutation when selection of the insert is negative, absent, or positive, respectively. The frequency of indels is known to be considerably lower than that of point substitutions (20A, 59A). Thus, the coincident fixation of several TRIM5
inserts during a short period of primate evolution is indicative of either an unusually high rate of indel creation or the pres-ence of positive selection favoring fixation of the insert. Be-cause of evidence of selection in flanking regions and the relative stability of the indel regions inTRIM5genes over long evolutionary periods, we consider positive selection to be a more likely explanation for the fixation of the inserts.
Beginning at discrete time points spaced over millions of years, TRIM5␣ B30.2 variable regions of particular lengths, and presumably with particular amino acid sequences as well, were sufficiently advantageous that the correspondingTRIM5
[image:9.585.54.272.71.179.2]genes became prevalent within the species. Episodic waves of lethal retrovirus infections may have contributed to this evo-lutionary pattern. The timing of some ancient retroviral epi-demics has been deduced from the presence of endogenous retroviruses in the germ lines of mammalian species (2, 12, 19, 21, 25, 26, 28, 35, 44, 45, 58, 59, 61). Notably, several endog-enous retroviruses appeared in primate germ lines during the same time periods in which two of the expansions of v1 length in the catarrhine TRIM5␣ proteins seem to have occurred. Between 25 and 40 million years ago, when Old World pri-mates diverged from New World monkeys, several endogenous FIG. 4. Nonsynonymous/synonymous ratios (Ka/Ks) for pairwise
comparisons ofTRIM5cDNAs. The plot shows theKa/Ksratios at
various codon positions for pairwise comparisons of human (H), patas monkey (P), tamarin (T), and spider monkey (S)TRIM5cDNAs. The alignment is the same as that shown in Fig. 1, with the gaps shared by allTRIM5genes removed. TheKa/Ksratio across the remaining gaps
was arbitrarily set to 0; thus, these gaps are indicated by sharp down-ward diversions of the plot lines. TheKa/Ks ratios, calculated by a
previously described method (24), were estimated as rolling averages for a window of 150 codons.
on November 8, 2019 by guest
http://jvi.asm.org/
retroviruses (human endogenous retrovirus type E [HERV-E], HERV-W, HERV-K, and ERV-9) were introduced into pri-mate genomes (21, 28, 35, 44, 45, 61). This corresponds to the period in which the TRIM5␣B30.2 v1 region length apparently expanded to 26 residues (Fig. 2B). About 9 to 13 million years ago, the simian endogenous retrovirus (SERV) and Papio
cynocephalusendogenous retrovirus were introduced into the
genomes of cercopithecine monkeys (26, 58, 59). The TRIM5␣ B30.2 v1 expansion to 28 residues occurred during this same period (Fig. 2B). Later TRIM5␣v1 expansions to 46 residues in the African green monkey/patas monkey lineage may also be related to retroviral infections. SIV infections are currently prevalent in these monkeys (10, 41), and SIV represents a can-didate retrovirus that could have applied selective pressure on
TRIM5. However, given the absence of endogenous SIV
pro-viruses, the extent of such infections in ancient times is un-known.
TRIM5␣ v3 expansion in New World monkeys could also involve ancient retroviral infections. The HERV-S and HERV-F endogenous retroviruses were introduced into primate germ lines between 32 and 56 million years ago, encompassing the period of divergence of the New World and Old World mon-keys (61). Further studies may reveal additional candidate ret-roviruses that potentially influenced platyrrhine TRIM5 pro-teins. Moreover, infectious agents beyond the retrovirus group may conceivably have influenced TRIM protein evolution.
The v1 region of the TRIM5␣B30.2 domain has undergone extensive changes during the evolution of Old World primates. In this light, it is noteworthy that the major determinant of anti-HIV-1 potency distinguishing the human and rhesus mon-key TRIM5␣ proteins was recently mapped to the B30.2 v1 region (48). Further studies will be needed to understand how the B30.2 variable regions might contribute to TRIM5␣ anti-retroviral activity. Length expansions of the degree observed in our survey of primate TRIM5␣sequences must be compatible with the fold of the B30.2 domain and thus are probably sur-face elements. The structure of the B30.2 domain is not known, although it has been speculated that it might be immunoglob-ulin-like (40). Surface-exposed variable elements of the B30.2 domain are reminiscent of the complementarity-determining region loops of immunoglobulins. Although we were not able to align the B30.2 variable segments and immunoglobulin CDRs in an instructive manner (data not shown), the TRIM B30.2 variable regions may play roles in the recognition of foreign ligands such as viral capsids.
The evolutionary relationships among TRIM genes in di-verse vertebrate species require additional investigation and may indicate functional interactions among the members of this interesting family of proteins. In parallel, the ability of TRIM proteins to modulate infections by viruses associated with these vertebrates should be investigated.
ACKNOWLEDGMENTS
We thank Yvette McLaughlin and Sheri Farnum for manuscript preparation and William Murphy and Jill Slattery for helpful discus-sions.
This work was supported by a grant from the National Institutes of Health (HL54785) and by a Center for AIDS Research award (P30 AI28691). We also acknowledge the support of the Bristol-Myers Squibb Foundation, the International AIDS Vaccine Initiative, and the late William F. McCarty Cooper. This work was funded in part with
federal funds from the Center for Cancer Research of the National Cancer Institute and the National Institutes of Health under contract no. NO1-CO-12400.
The content of this publication does not necessarily reflect the views of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorse-ment by the U.S. Governendorse-ment.
REFERENCES
1.Artes, E. J., and M. A. Wainberg.1996. Human immunodeficiency type 1 reverse transcriptase and early events in reverse transcription. Adv. Virus Res.46:97–163.
2.Benveniste, R. E., R. Heinemann, G. L. Wilson, R. Callahan, and G. J. Todaro.1974. Detection of baboon type C viral sequences in various primate tissues by molecular hybridization. J. Virol.14:56–67.
3.Benveniste, R. E., L. Kuller, S. T. Roodman, S. L. Hu, and W. R. Morton. 1993. Long-term protection of macaques against high-dose type D retrovirus challenge after immunization with recombinant vaccinia virus expressing envelope glycoproteins. J. Med. Primatol.22:74–79.
4.Besnier, C., Y. Takeuchi, and G. Towers.2002. Restriction of lentivirus in monkeys. Proc. Natl. Acad. Sci. USA99:11920–11925.
5.Besnier, C., L. Ylinen, B. Strang, A. Lister, Y. Takeuchi, S. P. Goff, and G. J. Towers.2003. Characterization of murine leukemia virus restriction in mam-mals. J. Virol.77:13403–13406.
6.Bieniasz, P. D.2003. Restriction factors: a defense against retroviral infec-tion. Trends Microbiol.11:286–291.
7.Bonilla, W. V., D. D. Pinschewer, P. Klenerman, V. Rousson, M. Gamboli, P. P. Pandolfi, R. M. Zinkernagel, M. S. Salvato, and H. Hengartner.2002. Effects of promyelocytic leukemia protein on virus-host balance. J. Virol. 76:3810–3818.
8.Chee, A. V., P. Lopez, P. P. Pandolfi, and B. Roizman.2003. Promyelocytic leukemia protein mediates interferon-based anti-herpes simplex virus 1 ef-fects. J. Virol.77:7101–7105.
9.Cowan, S., T. Hatziioannou, T. Cunningham, M. A. Muesing, H. G. Gott-linger, and P. D. Bieniasz.2002. Cellular inhibitors with Fv1-like activity restrict human and simian immunodeficiency virus tropism. Proc. Natl. Acad. Sci. USA99:11914–11919.
10.Daniel, M. D., N. L. Letvin, N. W. King, M. Kannagi, P. K. Sehgal, R. D. Hunt, P. J. Kanki, M. Essex, and R. C. Desrosiers.1985. Isolation of T-cell tropic HTLV-II-like retrovirus from macaques. Science228:1201–1204. 11.Freed, E. O.1998. HIV-1 gag proteins: diverse functions in the virus life
cycle. Virology251:1–15.
12.Goodchild, N. L., D. A. Wilkinson, and D. L. Mager.1993. Recent evolu-tionary expansions of a subfamily of RTVL-H human endogenous retrovi-rus-like elements. Virology196:778–788.
13.Hatziioannou, T., S. Cowan, S. P. Goff, P. D. Bieniasz, and G. J. Towers. 2003. Restriction of multiple divergent retroviruses by Lv1 and Refl. EMBO J.22:385–394.
14.Hatziioannou, T., S. Cowan, U. K. von Schwedler, W. I. Sundquist, and P. Bieniasz.2004. Species-specific tropism determinants in the human immu-nodeficiency virus type 1 capsid. J. Virol.78:6005–6012.
15.Hatziioannou, T., D. Perez-Caballero, A. Yang, S. Cowan, and P. D. Bie-niasz.2004. Retrovirus resistance factors Ref1 and Lv1 are species-specific variants of TRIM5. Proc. Natl. Acad. Sci. USA101:10774–10779. 16.Henry, J., I. H. Mather, M. F. McDermott, and P. Pontarotti.1998.
B30.2-like domain proteins: update and new insights into a rapidly expanding family of proteins. Mol. Biol. Evol.15:1696–1705.
17.Himathongkham, S., and P. A. Luciw.1996. Restriction of HIV-1 (subtype B) replication at the entry step in rhesus macaque cells. Virology219:485– 488.
18.Hofmann, W., D. Schubert, J. LaBonte, L. Munson, S. Gibson, J. Scammell, P. Ferrigno, and J. Sodroski.1999. Species-specific, postentry barriers to primate immunodeficiency virus infection. J. Virol.73:10020–10028. 19.Johnson, W. E., and J. M. Coffin.1999. Constructing primate phylogenies
from ancient retrovirus sequences. Proc. Natl. Acad. Sci. USA96:10254– 10260.
20.Keckesova, Z., L. M. J. Ylinen, and G. J. Towers.2004. The human and African green monkey TRIM genes encode Ref1 and Lv1 retroviral restric-tion factor activities. Proc. Natl. Acad. Sci. USA101:10780–10785. 20a.Keightley, P. D., and D. J. Gaffney.2003. Functional constraints and
fre-quency of deleterious mutations in noncoding DNA of rodents. Proc. Natl. Acad. Sci. USA100:13402–13406.
21.Kim, H. S., O. Takenaka, and T. J. Crow.1999. Isolation and phylogeny of endogenous retrovirus sequences belonging to the HERV-W family in pri-mates. J. Gen. Virol.80:2613–2619.
22.Kootstra, N. A., C. Munk, N. Tonnu, N. R. Landau, and I. M. Verma.2003. Abrogation of postentry restriction of HIV-1-based lentiviral vector trans-duction in simian cells. Proc. Natl. Acad. Sci. USA100:1298–1303. 23.Kumar, S., K. Tamura, and M. Nei.1994. MEGA: molecular evolutionary
genetics analysis software for microcomputers. Comput. Appl. Biosci.10: 189–191.
on November 8, 2019 by guest
http://jvi.asm.org/
24.Li, W. H.1993. Unbiased estimation of the rates of synonymous and non-synonymous substitution. J. Mol. Evol.36:96–99.
25.Mager, D. L., and J. D. Freeman.1995. HERV.H endogenous retroviruses— presence in the New-World branch but amplification in the Old-World primate lineage. Virology213:395–404.
26.Mang, R., J. Maas, A. C. van der Kuyl, and J. Goudsmit. 2000.Papio cynocephalus endogenous retrovirusamong Old World monkeys: evidence for coevolution and ancient cross-species transmission. J. Virol.74:1578–1586. 27.Marcello, A., A. Ferrari, V. Pellegrini, G. Pegoraro, M. Lusic, F. Beltram, and M. Giacca.2003. Recruitment of human cyclin T1 to nuclear bodies through direct interaction with the PML protein. EMBO J.22:2156–2166. 28.Mariana-Costantini, R., T. M. Horn, and R. Callahan.1989. Ancestry of a
human endogenous retrovirus family. J. Virol.63:4982–4985.
29.Munk, C., S. M. Brandt, G. Luccero, and N. R. Landau.2002. A dominant block to HIV-1 replication at reverse transcription in simian cells. Proc. Natl. Acad. Sci. USA99:13843–13848.
30.Nisole, S., C. Lynch, J. P. Stoye, and M. W. Yap.2004. A Trim5-cyclophilin A fusion protein found in owl monkey kidney cells can restrict HIV-1. Proc. Natl. Acad. Sci. USA101:13324–13328.
31.Orimo, A., N. Tominaga, K. Yoshimura, Y. Yamauchi, M. Nomura, M. Sato, Y. Nogi, M. Suzuki, H. Suzuki, K. Ikeda, S. Inoue, and M. Mauramatsu. 2000. Molecular cloning of ring finger protein 21 (RNF21)/interferon-re-sponsive finger protein (ifp1), which possesses two RING-B box-coiled coil domains in tandem. Genomics69:143–149.
32.Owens, C. M., P. C. Yang, H. Gottlinger, and J. Sodroski.2003. Human and simian immunodeficiency virus capsid proteins are major viral determinants of early, postentry replication blocks in simian cells. J. Virol.77:726–731. 33.Owens, C. M., B. Song, M. J. Perron, P. C. Yang, M. Stremlau, and J.
Sodroski.2004. Binding and susceptibility to post-entry restriction factors in monkey cells are specified by distinct regions of the human immunodefi-ciency virus type 1 capsid. J. Virol.78:5423–5437.
34.Perron, M., M. Stremlau, B. Song, W. Ulm, R. Mulligan, and J. Sodroski. 2004. TRIM5␣mediates the postentry block to N-tropic murine leukemia viruses in human cells. Proc. Natl. Acad. Sci. USA101:11827–11832. 35.Reus, K., J. Mayer, M. Sauter, H. Zischler, N. Mu¨ller-Lantzsch, and E.
Meese.2001. HERV-K (OLD): ancestor sequences of the human endoge-nous retrovirus family HERV-K (HML-2). J. Virol.75:8917–8926. 36.Reymond, A., G. Meroni, A. Fantozzi, G. Merla, S. Cairo, L. Luzi, D.
Riganelli, E. Zanaria, S. Messali, S. Cainarca, A. Guffanti, S. Minucci, P. G. Pelicci, and A. Ballabio. 2001. The tripartite motif family identifies cell compartments. EMBO J.20:2140–2151.
37.Rhodes, D. A., G. Ihrke, A. T. Reinicke, G. Malcherek, M. Towey, D. A. Isenberg, and J. Trowsdale.2002. The 52,000 MW Ro/SS-A autoantigen in Sjogren’s syndrome/systemic lupus erythematosus (Ro52) is an interferon-gamma inducible tripartite motif protein associated with membrane proxi-mal structures. Immunology106:246–256.
38.Rogers, J. S., and D. I. Swofford.1998. A fast method for approximating maximum likelihoods of phylogenetic trees from nucleotide sequences. Syst. Biol.47:77–89.
39.Sayah, D. M., E. Sokolskaja, L. Berthoux, and J. Luban.2004. Cyclophilin A retrotransposition into TRIM5 explains owl monkey resistance to HIV-1. Nature430:569–573.
40.Seto, M. H., H.-L. C. Liu, D. Zajchowski, and M. Whitlow.1999. Protein fold analysis of the B30.2-like domain. Proteins35:235–249.
41.Sharp, P. M., E. Bailes, R. R. Chaudhuri, C. M. Rodenburg, M. O. Santiago, and B. H. Hahn.2001. The origins of acquired immune deficiency syndrome viruses: where and when? Philos. Trans. R. Soc. Lond. B Biol. Sci.356:867– 876.
42.Shibata, R., H. Sakai, M. Kawamura, K. Tokunaga, and A. Adachi.1995. Early replication block of human immunodeficiency virus type 1 in monkey cells. J. Gen. Virol.76:2723–2730.
43.Song, B., H. Javanbakht, M. Perron, D. Park, M. Stremlau, and J. Sodroski. 2005. Retrovirus restriction by TRIM5␣variants from Old World and New World primates. J. Virol.79:3930–3937.
44.Steele, P. E., M. A. Martin, A. B. Rabson, T. Bryan, and S. J. O’Brien.1986. Amplification and chromosomal dispersion of human endogenous retroviral sequences. J. Virol.59:545–550.
45.Steinhuber, S., M. Brack, G. Hunsmann, H. Schwelberger, M. P. Dierich, and W. Vogetseder. 1995. Distribution of human endogenous retrovirus
HERV-K genomes in humans and different primates. Hum. Genet.96:188– 192.
46.Stoye, J. P.2002. An intracellular block to primate lentivirus replication. Proc. Natl. Acad. Sci. USA99:11549–11551.
47.Stremlau, M., C. M. Owens, M. J. Perron, M. Kiessling, P. Autissier, and J. Sodroski.2004. The cytoplasmic body component TRIM5␣restricts human immunodeficiency virus (HIV-1) infection in Old World monkeys. Nature 427:848–853.
48.Stremlau, M., M. Perron, S. Welikala, and J. Sodroski.2005. Species-specific variation in the B30.2(SPRY) domain of TRIM5␣determines the potency of human immunodeficiency virus type 1 restriction. J. Virol.79:3139–3145. 49.Thali, M., A. Bukovsky, E. Kondo, B. Rosenwirth, C. T. Walsh, J. Sodroski,
and H. G. Gottlinger.1994. Functional association of cyclophilin A with HIV-1 virions. Nature372:363–365.
50.Thomas, E. E., N. Srebro, J. Sebat, N. Navin, J. Healy, B. Mishra, and M. Wigler.2004. Distribution of short paired duplications in mammalian ge-nomes. Proc. Natl. Acad. Sci. USA101:10349–10354.
51.Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G. Higgins.1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res.25:4876–4882.
52.Toniato, E., X. P. Chen, J. Losman, V. Flati, L. Donahue, and P. Rothman. 2002. TRIM8/GERP RING finger protein interacts with SOCS-1. J. Biol. Chem.277:37315–37322.
53.Towers, G., M. Bock, S. Martin, Y. Takeuchi, J. P. Stoye, and O. Danos. 2000. A conserved mechanism of retrovirus restriction in mammals. Proc. Natl. Acad. Sci. USA97:12295–12299.
54.Towers, G., M. Collins, and Y. Takeuchi.2002. Abrogation of Ref1 retrovi-rus restriction in human cells. J. Virol.76:2548–2550.
55.Towers, G. J., T. Hatziioannou, S. Cowan, S. P. Goff, J. Luban, and P. D. Bieniasz.2003. Cyclophilin A modulates the sensitivity of HIV-1 to host restriction factors. Nat. Med.9:1138–1143.
56.Tristem, M.2000. Identification and characterization of novel human en-dogenous retrovirus families by phylogenetic screening of the human ge-nome mapping project database. J. Virol.74:3715–3730.
57.Turelli, P., V. Doucas, E. Craig, B. Mangeat, N. Klages, R. Evans, G. Kalpana, and D. Trono.2001. Cytoplasmic recruitment of INI1 and PML on incoming HIV preintegration complexes: interference with early steps of viral replication. Mol. Cell7:1245–1254.
58.van der Kuyl, A. C., J. T. Dekker, and J. Goudsmit.1995. Distribution of baboon endogenous virus among species of African monkeys suggests mul-tiple ancient cross-species transmissions in shared habitats. J. Virol.69:7877– 7887.
59.van der Kuyl, A. C., R. Mang, J. T. Dekker, and J. Goudsmit.1997. Complete nucleotide sequence of simian endogenous type D retrovirus with intact genome organization: evidence for ancestry to simian retrovirus and baboon endogenous virus. J. Virol.71:3666–3676.
59a.Watanabe, H., A. Fujiyama, M. Hattori, T. D. Taylor, A. Toyoda, Y. Kuroki, H. Noguchi, A. BenKahla, H. Lehrach, R. Sudbrak, M. Kube, S. Taenzer, P. Galgoczy, M. Platzer, M. Scharfe, G. Nordsiek, H. Blocker, I. Hellmann, P. Khaitovich, S. Paabo, R. Reinhardt, H. J. Zheng, X. L. Zhang, G. F. Zhu, B. F. Wang, G. Fu, S. X. Ren, G. P. Zhao, Z. Chen, Y. S. Lee, J. E. Cheong, S. H. Choi, K. M. Wu, T. T. Liu, K. J. Hsiao, S. F. Tsai, C. G. Kim, S. Oota, T. Kitano, Y. Kohara, N. Saitou, H. S. Park, S. Y. Wang, M. L. Yaspo, and Y. Sakaki.2004. DNA sequence and comparative analysis of chimpanzee chromosome 22. Nature429:382–388.
60.Whitcomb, J. M., and S. H. Hughes.1992. Retroviral reverse transcription and integration: progress and problems. Annu. Rev. Cell Biol.8:275–306. 61.Wilkinson, D. A., D. L. Mager, and J.-A. C. Leong.1994. Endogenous human
retroviruses, p. 465–535.InJ. A. Levy (ed.), The Retroviridae, vol. III. Plenum Press, New York, N.Y.
62.Xu, L., L. Yang, P. K. Moitra, K. Hashimoto, P. Rallabhandi, S. Kaul, G. Meroni, J. P. Jensen, A. M. Weissman, and P. D’Arpa.2003. BTBD1 and BTBD2 colocalize to cytoplasmic bodies with the RBCC/tripartite motif protein, TRIM5delta. Exp. Cell Res.288:84–93.
63.Yap, M. W., S. Nisole, C. Lynch, and J. P. Stoye.2004. Trim5 protein restricts both HIV-1 and murine leukemia virus. Proc. Natl. Acad. Sci. USA101: 10786–10791.