0022-538X/82/010210-12$02.00/0
Interference
Among
Defective
Interfering
Particles
of
Vesicular Stomatitis
Virus
DONALDD.RAOt ANDALICE S. HUANG*Departmentof Microbiology and MolecularGenetics,HarvardMedical School, and DivisionofInfectious
Diseases, Children'sHospitalMedicalCenter, Boston, Massachusetts 02115
Received 11 May1981/Accepted 3September1981
Three defectiveinterfering(DI)particles of vesicular stomatitis virus
(VSV),
allderivedfrom thesameparentalstandard San Juan strain(Indianaserotype),were
used invarious combinations toinfect cells togetherwith theparentalvirus.The
replication of their RNA genomes in the presence of othercompetinggenomes
wasdescribed by the hierarchical sequence: DI 0.52particles>DI 0.45particles DI-T particles> standardVSV.TheadvantageofoneDIparticleoveranother was not due simply to multiplicity effects nor tothe irreversible occupation of
linmitedcellular sites. Interference, however, did correlate with a changein the
ratio of plus and minus RNAtemplatesthataccumulated
intracellularly
andwith the presence ofnewsequencesatthe 3' end of the DIgenomes. DI 0.52 particles containedsignificantlymorenucleotidesatthe 3' end thatwerecomplementaryto those at the 5' end of its RNA than did DI-Tor DI 0.45 particles. The first 45 nucleotides at the 3' ends of allofthe DI RNAswereidentical. VSV and its DIparticlescan be separated into three
classes,
depending
on their terminal RNAsequences. These sequences suggesttwomechanisms,onebasedontheaffinityof
polymerase binding and the otheron theaffinity ofN-protein binding, that may
account for interference by DI particles against standard VSV and among DI
particlesthemselves.
When viruses grow to high titer, deletion mutants arise. They become enriched in the
population, especially ifthey have thecapacity
tointerferewith the growth of the standard
wild-type parent (12,14, 45). To study the molecular
basis of thisinterference, we have used
vesicu-larstomatitis virus (VSV) because of its
asym-metric bullet shape, which permits the
separa-tion of shorter deletion mutants from the parental wild type(15). Thisseparation has led tothecharacterization of individual size classes
of defective interfering (DI) particles and the
reconstruction of experiments with known amounts of standard and DIparticles.
Theprimarycompetition between DI particles andstandardVSV occurs intracellularly during the step of RNA replication or genome amplifi-cation (17,
29,
30). Very little is known about this mechanism, because RNA replication is initiated early during infection, and all of the proteinsthat are involved have not been identi-fied. Itisknown, however, that inputmultiplic-itiesaffect this interference, that the small size
of theinterferingRNAalone cannotaccount for its ability to compete during replication, and that the derivation ofaDIparticle inrelation to the
tPresentaddress:Department of Microbiology,
Washing-tonUniversity, St. Louis, MO 63110.
strain ofstandardhelper virus determines in part the degree of competition and the subsequent ability of the DI RNAtobereplicated(14).
Continuous undiluted passages of the same VSV strain may result in the appearance of several different size classes of DI particles, withsome orone-sizeparticleseventually domi-nating the viral population (11, 21, 31, 37). These results show that interference occurs not only between DI particles and standard virus, but also among different DI particles. Byexamining DI particles,all derived from the sameparental standardvirus,itis suggested herethat, besides
RNA replication, there is another rate-limiting step during virusmaturation when interference
occurs. We found that the ability to interfere increased with the extent of complementary sequences foundat the termini of the RNA.
MATERIALS AND METHODS
Materials. The source of all materials has been
recently reported (35) except for thefollowing.
Cyti-dine 3',5'-bisphosphate and [,y-32P]ATP were
pur-chased from New England Nuclear Corp., Boston,
Mass. Actinomycin D was a kind gift from Merck
Sharp& Dohme, Rahway, N.J. Practical-grade
acryla-mide andelectrophoretic-grade
N,N'-methylene-bisa-crylamide were purchased from Eastman Kodak
Chemical Co., Rochester, N.Y. Aminobenzyloxy-methyl (ABM)-cellulose paper was purchased from
210
on November 10, 2019 by guest
http://jvi.asm.org/
Schleicher & Schuell Co., Keene, N.H.
Polynucleo-tide kinase andRNAligase fromT4-infected
Escheri-chia coliwerepurchased fromP-LBiochemicals,Inc.,
Milwaukee, Wisc. Calfintestine alkaline phosphatase
waspurchased from Boehringer Mannheim
Biochemi-cals, Indianapolis, Ind.RNasesTl,T2, A, andU2were
purchasedfrom Calbiochem-BoehringCorp., San
Die-go, Calif. RNase physarum Mwas akindgift from
Helen Donis-Keller.
Cells andviruses. BHK, CHO, andmouseL cells
were grown assuspensioncultures in Joklik modified
Eagle minimal essential medium supplemented with
5% bovine serum and nonessential amino acids.
Cloned andsucrosegradient-purified standardVSVof
theIndianaserotype(SanJuanstrain) and three size
classes of DIparticles derived from thissamestandard
strain were used. According to a recent agreement
(36), the threeDIparticlesaredesignated: VSIts+ SJ
DI-T/DI 0.33 (5') (15,19);VSIts+SJDI0.52 (5') (19,
34); and VSI ts+ SJDILT/DI0.45 (5') (5). DI 0.45was
kindly supplied by SueEmerson(University of
Virgin-ia,Charlottesville). All DI particles werepurified by
tworounds ofsucrosegradient centrifugation. Their
effectivemultiplicitywasdeterminedasdescribed by
Huangand Manders (17).
Infection ofceUsandextraction of cytoplasmicRNA.
Infection of4 x 106 cellsat34°C with the indicated
multiplicities for each experimentwasdone essentially
as described (9, 35). When RNAwas examined,the
cellsweretreated with actinomycinDat5 p.g/mlatthe
beginning of infection and0.1mCi of 32ppermlatthe
indicatedtimes. Cytoplasmicextractsweremade with
the nonionic detergent Nonidet P-40 (17).
Phenol-chloroform extractions of the RNA from the
cyto-plasmweredoneasdescribed previously(9,35).
Separation of RNA. For the separation of RNA
species ranging from 30 to 12,000 nucleotides, two
different gel systems (10%opolyacrylamide and1.5%
agarose)wereused as previously described(35). To
provide quantitative comparisons ofthe amounts of
RNA, either the autoradiogram was scanned orthe
bandswereexcised from the gels and theradioactivity
was counted. Conclusions basedontheincorporated
radioactivity(countsperminute)weresimilartothose
made whenmolaramountsof RNAwerecalculated.
Elution of RNA from 10%polyacrylamdegels. After
electrophoresis of RNAon apolyacrylamidegel, the
gel was covered with Saran Wrap and
autoradio-graphed. Using the autoradiogramas atemplate, the
region of thegel containing labeled RNAwasexcised
and slicedintopieces of 2mm3.Smallgel cubeswere
thentransferredtoasiliconized tubetogether with2
volumes ofelution buffer (0.4 M sodiumacetate,0.01
M Tris, pH 7.8, and 0.2% sodium dodecyl sulfate)
containing 50,ug ofyeastcarrier RNAperml.Elution
proceededovernightat22°C with intermittent mixing. The small gel pieces were rinsed with 1 additional
volumeof elution buffer. Thecombinedeluateswere
then filtered through a 0.45-p.m filter to retain any
remainingparticulatepolyacrylamide. The RNAwas
thenprecipitatedby2volumesofethanol.
Transfer ofRNAfrom agarosegels toDBMpaper
andhybridization to32P-labeledRNA.The procedure
of Alwine et al. (1) was used with the following
modifications. Potassium phosphate buffer, pH 6.5,
was used instead of sodium borate buffer, pH 8.0.
Unlabeled VSV-specificRNAextracted from virions
andfrom thecytoplasm was firstseparatedon a1.5%
agarose gel. After urea was diffused out of the gel, the
gel wasplacedin200ml of 50 mMNaOH and rocked
gently for 40 min at roomtemperature. The gel was
thentreated twice with 200 ml of 200 mMpotassium phosphate buffer,pH6.5, for 5 min at22°C.
Conver-sion of the ABM paper to diazobenzyloxymethyl
(DBM) paper, subsequent transferof RNA from the
gel to DBM paper, and hybridization conditions all
followedthe
procedures
describedbyAlwineetal.(1),exceptthat3P-labeledsmallRNA from cells
coinfect-ed withDI-T andstandard VSV was thehybridization
probe ratherthan 32P-labeled DNA. After the
unhy-bridizedlabeledRNA wasthoroughlywashed off, the
DBMpaper was placed inSeal-N-Saveboilable bags
(SearsRoebuck and Co.) and exposedto X-rayfilm
with anintensifying screenat-70°C.
Extractionandpurificationof DIparticleRNA.
Puri-fied DI particles were diluted two- tothree-foldwith
Earle saline and then pelleted by centrifugation at
89,300 x g for90 min. The pelleted particles were
suspendedinETS buffer (0.01 M EDTA, 0.01 M Tris,
pH 7.4, and0.2%sodiumdodecyl sulfate)and
repeat-edly extracted with phenol-chloroform. Virion RNA
wasprecipitatedwithethanol.The RNAresuspended
in ETS bufferwas loaded onto a 15 to 30o (wt/wt)
sucrosegradientmadeinNETS buffer (0.15 MNaCIin
ETS buffer).Centrifugationwasfor 17 h at 89,300xg.
Thegradient fractions werecollected andmonitored
by UV absorbance. Full-length DI-particle genomic
RNA wasthenpooledandethanolprecipitated.
Radioactive
labeling
of theterminiofthe RNA of DIpartices.
Purified RNAs from DI particles werela-beled at the 3'endbyligationof[32P]cytidine
3',5'bis-phosphate accordingtoEnglandandUhlenbeck(6) as
modified by Schubert et al. (40). Afterlabeling, the
RNA was again purified through sucrose gradients.
For labeling atthe 5' end, purifiedRNAs were first
suspendedin 10 mMTris,pH 8.4, boiledfor1.5min,
and thenrapidlychilled to4°C. Calf intestine alkaline
phosphatasewasaddedto aratio of16-3Uper Fg of
RNA. After 30 min at34°C,the RNAwasextracted
with phenol-chloroform repeatedly and thenethanol precipitated. The RNA was suspended in 20 Ill of
buffer consisting of 50 mM Tris, pH 8.4, 10 mM
MgCi2,and 5 mMdithiothreitol. [y-32P]ATP at 100,uCi
was added, and the whole sample was lyophilized.
Then 20,ulof steriledouble-distilledwatercontaining
10 Uofpolynucleotidekinasewasaddedfor 30 minat
370C. RNA was again extractedwith
phenol-chloro-formrepeatedlyand thenethanolprecipitated. solationof
self-complementary
terminal sequences.End-labeledRNAsat1to4,ug from DIparticleswere
mixed with 50 ,ug ofyeastcarrier RNA in 25 p.l of
distilled water. Equal volumes of 2x hybridization
buffer(1x = 0.4 MNaCl, 0.01 MTris, pH 7.4,and
0.01 MEDTA)wereadded,and then the RNAsample
wassealed inacapillarytube andincubatedat70°Cfor
2 h.Sampleswerepipettedinto 200p.lof
hybridization
buffer containing, per milliliter, RNase T. at 50 U,
RNaseT2at5U,and RNase Aat100 ,ug. Digestion
proceededat37°Cfor 30min. Afterdigestion,
samples
wereextractedrepeatedly withphenol-cjlloroform
in thepresenceof addedcarrierRNA(50to100p.g)
and precipitatedwithethanol.Todenaturedouble-strand-VOL. 41,1982
on November 10, 2019 by guest
http://jvi.asm.org/
ed RNA, the RNAwassuspendedin 10
7.4,boiled for 1.5min, andrapidlychill4
RESULTS
Interference among different D Three different DIparticles,allisola
San Juan strain ofVSV, were use(
combinations together with standa
infect cells in orderto testwhether
the DIparticles was particularly mc
than the others during interferenc4 measuredbycomparingthe amount
lular RNAspecifictoeach DIparti4 As shown by the absence of stai
specific 40S RNA, each of the DI ;
0.45, DI 0.52, and DI-T) was capa pletely inhibiting the replication
VSV without affecting primary tran
the L,G, N,NS, andMmRNA's(Fi
f, and g). These resultsmeasuringRI ed well with inhibition of infecti identical combinations were used
forming abilityof standardVSV was three DI particles, singly or in c
a
b
c
d
e f
g
2.:r
um-L
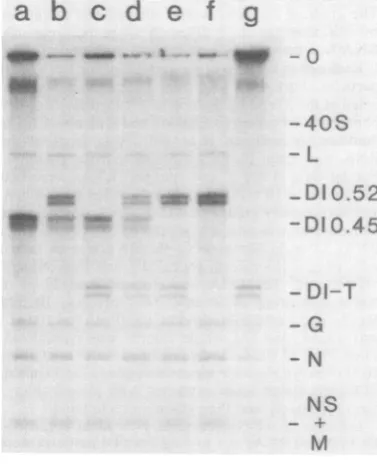
FIG. 1. Interference amongDIparti
BHK cells in five cultures were infec
indicated virusesatmultiplicitiesof 20(
addedtotheculturesat1to6 hpostini
plasmic RNAwasthenextracted, and e
were separated on a1.5%agarose gel.
VSV plusDI 0.45 particles; (b)standard
0.45plus DI 0.52particles;(c) standard
0.45plus DI-T particles;(d) standard VSI
plusDI0.52 plusDI-Tparticles; (e) stand
DI0.52plusDI-T particles; (f) standard
0.52particles; (g) standardVSV plus Dl
mMTris, pH completely inhibited the growth ofVSV (D. D. ed onice. Rao and A. S. Huang, unpublished data).
Radio-active material between theoriginand40S RNA
probably represents contaminating DNA and
1I particles. some trapped RNA. Agarose gel patterns of
tedfromthe RNA obtained from cells infected with only d in various standard VSV have beenpublished (19, 34,35).
ird VSV to When cells weretriply infected with anytwo ranyone of of the DIparticlesandstandard VSV,40S RNA reeffective synthesis was still inhibited, but there were
e. This was varying degrees ofreplication of thegenomes of
tofintracel- DIparticles. DI0.45 and DI-Tparticles replicat-cle. ed about equally well in each other's presence
ndard virus- (Fig. 1, slot c), whereas DI 0.52particles hadan
particles
(DI advantage over the two smaller DI particles. Lble of com- When theindividual bandswereexcised and theof standard radioactivitywascompared by Cerenkov
count-iscription of ing, DI 0.52 particles synthesized between 1.2
ig. 1, slotsa, and 2.5 times the molar amount of plus and
NA correlat- minus strands synthesized by the smaller DI
ivity. When particles. The advantage increased with time
and plaque- (Fig. 1, slots b and e). When all three DI assayed, all particles were added to cells in the presence of
,ombination, standardvirus, therewasanoverall reductionin
the totalamountof RNAreplication for each of the genomes, but DI 0.52stillappearedto
repli-cate its RNA somewhat better than the other
twoDI particles (Fig. 1, slot d).
- O These results indicate that all three DI parti-cles interfered effectively against the parental standard VSV. Among the DI particles, the -40£)S
largest
oneappeared
tohaveareplicative
advan-tageoverthetwosmallerones.Inonlyonecycle- L of growth, interference against standard virus
was complete, whereas interference among DI
-DI 0.52 particles was onlypartial.
-DI0.4
Symmetry
ofreplication.
During
standardVSV genome replication, plus and minus RNA templates accumulate asymmetrically, with the bulk of the RNA being of the minus polarity(27,
- DI-T 44). It has been postulated thatdifferences
be-G tween plus and minus strands in their binding
affinities forpolymerases may account for this - N
asymmetry
and that ifbinding
affinities of DItemplates
arealtered,
the differentialaccumula-N S tion of the plus and minus strands may be +
changed
as well(16,
33).
Because theplus
andM minusstrands of RNA specific to thereplication of VSVDIparticleshavebeen recently separat-icles ofvsv. ed on gels (19), the two templates were
mea-:ted with the sured directly.
each. 32pwas To test the above hypothesis, the doublet
fection. Cyto- bands of DI RNAs from coinfected cells were
,qual
Portions
quantitated, and the ratio of plus strands(bot-VSV
plustDI
tomband)
to minus strands(top
band)
wasVSV plus DI determined
(Table 1).
DI 0.52particles
synthe-V plus DI0.45 sizedvirtually
equivalent
amounts ofplus
andlardVSVplus minus
strands,
whereas thetwosmallerDIparti-VSVplus DI cles made plus and minus RNA in the ratio of
I-Tparticles. 40:60.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.61.250.343.575.2]MECHANISMS INTERFERENCE
TABLE 1. Ratio of intracellular plus and minus
RNAspecific to DI particles'
DIparticles
Plusstrand Minusstrand
DI0.52 49±3 51±2
DI 0.45 40±1 60±1
DI-T 40±1 60±1
aBHK
cells
were coinfected with standard VSVand oneof the three DI particles, each at amultiplicity
of20, andexposed to32pfrom 0 to 6 h. Then the RNA
was extracted, separated on an agarose gel, and
ex-posedtoX-rayfilm. The bands representing
intracellu-lar RNAof the size of the genome of each DI particle
werequantitated by scanning thefilm.Theassumption
was made thatthefaster-migratingband is plus
strand-ed and theslower-migrating band is minus stranded
(19).
Thesedirect measurements confirm previous results for DI-T particles obtained by RNA hybridization studies (18). This is in contrastto
the ratio of20:80 for the intracellular plus and minusRNAstrands of standard VSV (27). These resultsindicate that as the interfering capability of the DIparticles increased, there appeared to be a moreequal accumulation of plus and minus RNAstrands.
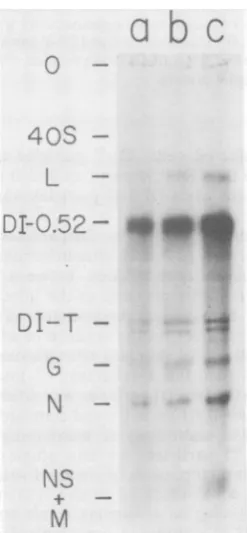
Interference by DI 0.52 particles against DI-T
particles and standard VSV in mouse L, CHO,
and BHKcells. Because DI-Tand DI 0.45
parti-cles appeared to be nearly equivalent in their
interfering potential and because DI 0.45
parti-cles have been studied elsewhere (40, 41), we chose tofocuson DI 0.52and DI-Tparticles for
detailed comparative studies. Initial attempts
were to select a cell line which showed the greatestdegree ofinterference. Three different cell lineswereeachtriply infectedwith standard
VSVand bothDIparticles,all atequal
multiplic-ities. When equal portions of the RNA from
each celltypewere separated on1.5% agarose
gels, RNA specific to DI 0.52 particles was
alwaysthepredominant
species
(Fig. 2).RNAofthesize ofstandard virus genomes,migratingat
40S, was undetectable, but some RNA
specific
toDI-Tparticleswasseen(Fig. 2).Theseresults
indicate that all three cell linessupported
inter-ference by DI 0.52 particles
against
standardVSV and DI-T particles. In
addition,
as wasseeninFig. 1, interference among the DI
parti-cles wasquantitatively
different frominterfer-ence withstandard virusbyDI
particles.
Parenthetically,the totalamountof32p
incor-poratedinto the RNA from
equivalent
numbersofcellswashighestfor BHK cells. This reflects
thedifferent abilities of thesecelltypesto incor-porate
32p
into totalvirus-specific
RNA,
eventhoughthe
production
ofinfectiousprogenywasequivalentfor each of thesethreecell lines
(G.
J.Freeman, Ph.D. thesis, Harvard University, Cambridge, Mass., A79). Because of theability ofsuspended BHKcellsto incorporate
32p
into VSV RNA, these cells were chosen for all subsequent experiments.Relative accumulation of RNA species during interference between DI 0.52 and DI-Tparticles. Tomeasure the rate of inhibition of synthesis of RNA at hourly intervals after infection,
32p-labeled RNAfrom cells coinfected with standard VSV and each orboth of the DI particles was separated and quantitated. Figure 3 shows the accumulation of the intracellular RNAs from each of the three sets of infected cells. The accumulation of total plus andminus strands of DIRNAs was compared with that of the mRNA for the N protein from the same cells. Whencellswere coinfected with either one of the DI
particles and standard VSV, DI RNA replication occurred to about the same extent (Fig. 3a,b). When cells were triply infected, total RNA synthesis was reduced to approximately 70% of that in cells coinfected with only one type of DI
particle and standard VSV (Fig. 3c). In these
c
040S
-
4K-DI-0.52
-DI-T
-
G-
N-.4.
NS
M
FIG. 2. Competition between DI-T and DI 0.52
particlesindifferent cell lines.Mouse L(a),CHO(b),
and BHK (c) cells were each triply infected with
standardVSV, DI0.52particles, and DI-Tparticles,
eachat amultiplicityof 20.32Pwasadded from 1to6 h
after infection. Equal amounts of the cytoplasmic
RNAobtained from these infected cellswere
separat-ed on1.5%agarosegels.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.298.422.335.602.2]214 RAO AND HUANG
re)
0 0
0
HOURS
POST
INFECTION
FIG. 3. Accumulationofintracellular RNAspeciesincells coinfected with standard VSV and DIparticles.
BHKcells werecoinfectedwithstandard VSV and either DI 0.52orDI-Tparticles.Another culture of cells was
triplyinfected with standard VSV and both DIparticles.Allmultiplicitieswereat20 each.32Pwasadded at the
time of infection together with actinomycin D. At hourly intervals, equal portions were removed, and
cytoplasmicRNA wasextracted andseparatedon1.5%agarosegels.Each of theVSV-specificRNA-containing
bands wasexcised,and theradioactivitywasquantitated byCerenkovcounting. (a)Standard VSV and DI 0.52
particles; (b)standard VSV and DI-Tparticles; (c)standardVSV and both DIparticles. Symbols: (@) plus-and
minus-strand RNA of DI 0.52particles; (0) plus-and minus-strand RNA of DI-Tparticles; (x)mRNA for the
nucleocapsid protein.
triply infected cells, DI-T particles replicated
less well than DI 0.52particlesand atonly 25%
theamountwhen DI 0.52particles were absent
(Fig. 3c).
Interference among the DI particles was de-tectedasearlyas2to3h afterinfection,withthe
difference in accumulation between the two
RNA genomes increasing as the infection
pro-gressed. Thisgradualaccumulation ofgenomes of DI-Tparticles, in the absence ofany detect-able synthesis ofstandard virusgenomes, again
suggests that the mechanism of interference
betweenthetwoDIparticleswasdifferentfrom thatbetweenDIparticles andstandardVSV.
Effectsofmultiplicity oninterference between the two DI partides. Previous studies showed thatinput multiplicitiesof standard virus and DI
particlescouldaffect the degree of interference, thereby giving an erroneous result concerning theabilitytointerfere byaparticularDIparticle (20,43). Toensurethat thepartial inhibition by DI0.52particles against DI-T particles wasnot
due simply to multiplicity, cells were triply infected with DI 0.52 particles together with standardVSV,eachataconstantmultiplicity of 20, and then with DI-T particlesover afivefold
range. As a measure ofinterference, the total
radioactivityinplus and minus RNA of thetwo
DIparticles wascompared.
In these triply infected cells, RNA of DI-T particles accumulated to the same reduced
amountdespite increased multiplicities of DI-T particles (Fig. 4). The lack of enrichment of RNA of DI-T particles at higher multiplicities indicated that the inhibitory effect of DI 0.52 particlesonDI-Tparticleswasmultiplicity
inde-pendent.
On the other hand, at the higher total multi-plicities, RNA of DI 0.52 particles was de-creased(Fig. 4). This reverse effect of increased
multiplicity of DI-Tparticles on the interfering
DI 0.52 particles relates to an expected
multi-plicity-dependent event where the competition
forproteins involved in RNA replicationis equal
between the two incoming DI genomes.
Effectofcycloheximide on the initial
competi-tion between DI-T andDI 0.52 particles.
Previ-ously, itwasfound that an advantage was recov-ered by DI-T particles over DI 0.52 particles if cellswereinfected with DI-T particles and stan-dard VSV for a period before superinfection with DI 0.52 particles (19). To determine
wheth-er this superinfection exclusion was due to the
ability ofinput DI-T particles to bind
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.97.412.71.295.2]4
0--
0-20:100 20:50 2.20
multiplicity DI-0.52 DI-T
FIG. 4. Effects of relative multiplicities on the
competition between DI-T and DI 0.52 particles.
Threecultures of BHKcellswereinfected with
stan-dardVSV and DI 0.52 particles, eachat amultiplicity
of 20, and with multiplicities of DI-T particles at20,
50, and 100. 32P was present from 1 to 6 h
post-infection. Cytoplasmic RNA was extracted and
sepa-rated on1.5% agarose gels. Bands containing
subgeno-mic-sized DI RNA, both plus and minus strands, were
excised, and the radioactivity was quantitated by
Cerenkov counting. Symbols: (d) RNA of DI 0.52 particles; (0) RNA of DI-T particles.
of DIparticlestocompeteagainst standard VSV (13, 23). It therefore becomes important to
ex-aminethesequences attheRNAterminiofboth DI genomes inorder todetermine whether any
differences could account for the ability of the
largerDIparticletoinhibit the smaller DI
parti-cle.Hybridization ofthe RNAgenomesin
solu-tiongave9%oself-annealing for both DI0.52and DI-Tparticles (19). Thissuggeststhatthelarger
genome mayhave moreself-complementary se-quences than the smaller genome. Oligonucleo-tide analysis indicates thatboth arederivedfrom the Lgeneandcontainsequencesspecifictothe 5' end of the VSV RNA (10, 19).
Totestforsequencedifferences atthe 3' end ofthe twoDI RNAs,wetook advantage of the
presence of a small RNA, 46 nucleotides in length,which is synthesized in largeamountsin cellscoinfected with standard VSV andDI
parti-a
bcde
ibly to limited resources or sites during the
period before superinfection, cycloheximide
was added during the period of superinfection. This prevents genome amplification without in-hibiting attachment and uncoating (17, 24). Cells coinfected with standard VSV and DI-T
parti-cles were treated or not with cycloheximide for 0.5 or 1 h and then superinfected with DI 0.52
particles, followedby the removal of
cyclohexi-mide0.5 h later.
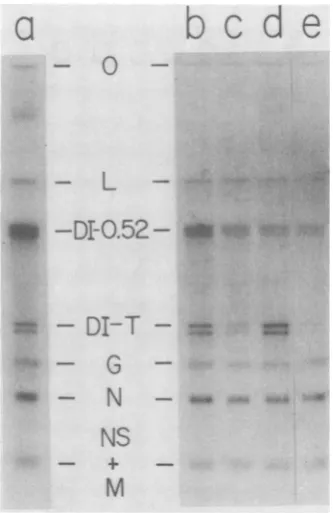
Figure 5a shows the usual predominance of
thelarger DI RNA when cells were triply infect-ed at the sametime.When DI-Tparticles had an
advantage of 0.5 or 1 h, there was a relative
decrease in theabilityof DI 0.52 toreplicate its
RNA (Fig. Sb,d). However, if cycloheximide
were present to prevent the amplification of
inputDI-Tgenomes, then the RNA of DI 0.52
was replicated to advantage over DI-T RNA
(Fig.
5c,e).
These results indicate that input DI-T parti-cles didnotirreversiblytieup limitedresources
of thecell, butthat superinfectionexclusion of
DI 0.52particlesrequired the initial increase in DI-Ttemplates. The end result ofpriorinfection by one DI particle was an increase in their
effectivemultiplicity.Therefore, prior infection, without RNAreplication,didnotirreversiblytie
up limitedresources oroccupy sites which con-trol RNAreplication.
Hybridization of small RNA to RNA from standardVSVand DIparticlesand from infected cells.Sequencedifferencesatthe3'end of RNA
genomesarepostulatedtoaccountfor theability
LJ
!
-DI-0.52
is
-DI-T -
= ::G2u'-
N
NS
M
FIG. 5. Reversal by cycloheximide of the competi-tive advantage of DI particles due to prior infection.
BHK cells at 20 x 106 cells/ml were infected with
standardVSV and DI-Tparticlesat amultiplicityof 20
each andthen divided into fiveequal portions.
Addi-tion of cycloheximide at 100 Fig/ml, superinfection
with DI 0.52 particles at a multiplicity of 20, and
exposureto32pwere asfollows. RNAwasharvested
at6 hafter infection andseparatedon a1.5%agarose
gel. (a)DI0.52particlesat0h,32Pat1to6h;(b)DI
0.52particlesat0.5h, 32Pat1.5to6h;(c) cyclohexi-mideat0to1h,DI0.52particlesat0.5h,32pat1.5to 6 h;(d) DI 0.52particles at1 h, 32P at2to6h; (e)
cycloheximideat0to1.5h,DI0.52particlesat1h, 32p
at2to6h.
- som 4:,::*:*4-a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.55.214.55.193.2] [image:6.504.272.439.281.538.2]216
cles(34). Because small RNA is co0
tothe 3' end of DI RNAs(23), itca
determine the identity or diverg quencesatthe 3' endby cross-hyb
To do so, 32P-labeled small Rls pared from cells coinfected with D and standard VSV (34, 35). Sma hybridized against unlabeled virio pared from standard VSV or DI
wellasagainst total unlabeled intra<
from singly or coinfected cells. TI
RNA was first separated on agar
then transferred topaper before hl Small RNA failed to anneal to t strand RNA from standard VSV bound to the negative-strand RNA DI 0.52orDI-Tparticles(Fig.6c,e
coinfectedcellswasrecognizedby
theywere DI genomes ortheircoI plus strand (Fig. 6d,f).
a
be
0-40S
-L
-DI-0.52
9
DI-T
-G
-N
-NS
[image:7.504.69.240.292.562.2]M
FIG. 6. Northern blot hybridization
toVSV-specificRNAspecies. Virion RI
,ug fromstandard VSVoritsDIparticle
lularRNAat3,Lgfromsinglyorcoinfec
separatedon a1.5%agarosegel, transfi
paper,andhybridizedto32P-labeledsm
(a) RNA fromstandard VSV;(b) RNA
VSV-infected cells; (c) RNA from DI(
(d) RNA fromcells coinfectedwith stan4
DI 0.52 particles; (e) RNA from DI-T
RNAfromcells coinfectedwithstandard
Tparticles.
mplementary Smallamounts ofhydridization occurred
be-an be used to tween small RNA and intracellular 40S RNA,
rence of se- presumably due to the complementary se-)ridization. quences onthelarge plusstrandwhich
comigrat-4A was pre- ed with virion 40S RNA (Fig. 6b). Annealingof
)I-Tparticles small RNA to the L mRNA from standard
VSV-,ll RNA was infected cells suggeststhat some L mRNAwas In RNA pre- not terminated, but contained run-through se-particles, as quences with 3' ends identical to those of the cellular RNA plus strand of 40S RNA (42). In Fig. 6f, the 'he unlabeled minor species whichbound small RNA was due ose gels and toacontaminatingDIparticlein ourDI-T prepa-iybridization. ration.
he negative- These hybridization results show that there
whereas it were sequences in common between the
ge-from either nomes of DI 0.52and DI-Tparticles whichwere
). RNA from not found on the negative-strand RNA of stan-small RNA if dard VSV. Thesesequenceswerepresumablyat
mplementary the 3' end. Therefore, if the two DI genomes
differed in their RNA sequenceatthe 3'
termi-nus,it would bedue to less than 20%divergence within the first 46 nucleotidesorinternaltothese
e nucleotides. Moreover, hybridization of small
RNA to both plus and minus strands of DI RNAs indicated sequence homology at the 3'
endsof theseRNAsandsuggested the presence
ofatleast 46nucleotidesthatwereterminal and
complementary at the 3' and 5' ends of both strands.
Isolation of self-hybridized duplex molecules
from the RNA of DI 0.52 and DI-T particles.
Because theprevious hybridizationresultswere indirect, adirectapproachwas taken to isolate
these terminal sequences by annealing. To do
so, RNAs from purified DI particles were la-beled at either the 5' or 3' end, rapidly
self-annealed, and digested with RNase. The
result-ing
duplex
molecules (stems) were electrophoresedon a10%polyacrylamide geltodetermine theirrelative sizes.
Discrete self-hybridizable pieces of 45, 48, and 55nucleotides wereformed from theRNA
of
DI-T,
DI0.45,
and DI 0.52particles,
respec-tively (Fig. 7). These lengths were determined by comparison with tRNA markers and the small RNA as well as from the sequencing results to follow. Because radioactive RNA, labeled in vitroateitherterminus before hybrid-ization and RNase digestion, appeared in the
of smallRNA isolated stems from all three DI particles, the
NAsat0.5to1 particles must be formed by self-annealing of :s and intracel- RNAsequences at the 3' and 5'terminiandnot
ted
cetlls
were
by loop-back or otherinternal
complementaryerredtoDBM sequences. These results prove that terminal
from
O.52
standard complementarity existed in the RNAs of allparticles;
threeDIparticles.
Differences in the stemsizesdardVSV and correlated with their ability tocompete against r particles; (f) each other.
J VSVandDI- Sequencing ofend-labeled RNA from DI 0.52
and DI-Tparticles.Toensurethat thesequences
on November 10, 2019 by guest
http://jvi.asm.org/
MECHANISMS OF INTERFERENCE 217
a b c d e f incomplete when measured by RNAsynthesis,
tRN A - whereas inhibition of standard virus growth by
any one of the three DI particles resulted in complete inhibitionof standard genome
replica-XC 10- a* tion as well as infectious progeny. Moreover,
_m
inhibitionof one DIparticle by another could beshown to be multiplicity independent, resulting VW
do
in the same partial degree of RNA replication,abc de
abcd
e
FIG. 7. Terminal complementary stems obtained
from5'- or 3'-end-labeled DI RNAs. Purified RNAs
from DI 0.52, DI 0.45, and DI-T particles were
termi-nally labeled at either the 5' or 3' terminus. The
labeled RNAs were again purified on sucrose
gradi-ents,self-annealed, and digested with RNases.
RNase-resistant RNAs were then separated on a10%o
poly-acrylamide gel. tRNA and xylene cyanol (XC) dye
markers are indicated by arrows. (a) DI 0.52 RNA,
5'-endlabeled; (b) DI 0.45 RNA,5'-endlabeled;(c) DI-T
RNA, 5'-end labeled; (d) DI 0.52 RNA, 3'-endlabeled;
(e) DI 0.45 RNA,3'-endlabeled;(f)DI-T RNA,3'-end
labeled.
at the termini ofthe RNAs of the DI particles differed only in the extent of complementarity, sequence analysis was done on the 3'-end-la-beled stems obtained from DI 0.52 and
com-pared withDI-Tparticles.The 3' and 5'ends of
theRNA from DI-T and DI 0.45 particles have been sequenced (40).
Figure8shows thepatternof RNA fragments
from DI RNAslabeled at the 3' end obtained by
digestionwith alkali or several different RNases.
The overallpattern was identical for 45
nucleo-tides,indicatingthat theonly difference between
DI 0.52 and DI-Tparticleswas in theextentof
the self-complementarity at the termini. When
wholeRNAfromDI0.52 and DI-Tparticleswas
similarly sequenced, divergence at the 3' end
wasdetected from the first45 nucleotides. The
exact sequences in these first 45 nucleotides agree with previously published sequences for DI-T and DI 0.45particlesexceptfor nucleotide
positions35 and 36(40).Theseresults, however,
were notclearenough above the first45 nucleo-tides topermitfurthersequencedeterminations.
DISCUSSION
These studies relate sequences on the RNA
genomesof VSV and itsDIparticlestopossible mechanisms of interference. Interference among VSVDIparticles themselveswasquantitatively
differentfrom their inhibition of standard VSV
[image:8.504.77.223.81.214.2]growth. Interference among DI particles was
FIG. 8. Sequence comparisons of 3'-end-labeled
complementaryRNAfrom DI 0.52 and DI-Tparticles.
RNA from the two DI particles were prepared as
described inMaterials andMethods. RNA stems
la-beledatthe 3' endwerethendenaturedby boilingand
quickcoolingbefore partialnucleasedigestion bythe
method ofDonis-Kelleretal.(4). Separationwas ona
20% polyacrylamide gel. (a-e) 3'-end-labeled RNA
from DI 0.52 pa~rticles; (a'-e') 3'-end-labeled RNA
from DI-Tparticles; (a, a')notreatment;(b, b')RNase
T1;(c, c')RNase U2; (d,d')RNase physarumM; (e,
e')alkalinedenatured.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.257.446.189.572.2]218 RAO AND HUANG
irrespective of the initial concentration of the suppressed DIparticle.
When interference is multiplicity dependent,
the mechanism is likely to be an early event
involving input virions, whereas
multiplicity-independentinterference suggestsamechanism which occurs after the initial steps of viral multiplication. DIparticleshaveonlytwo
essen-tial functions: genome replication and
matura-tion intovirus particles. One is anearly event;
the other is a late event. The early event is multiplicitydependent,and the lateoneis multi-plicity independent.Therefore, itcanbe postu-lated that a competitive advantage might be gainedduringeachofthese functional steps.
How competitive advantages can be gained
during these steps may befoundinanevaluation ofthe RNA in VSV and in the DI
particles.
DI genomes contain additional complementaryse-quences aswellasthesingle-base
mutations
(8, 10, 40). Because single-base mutations are indifferent coding regions for the L
protein,
it isunlikely that they are involved in interference.
Complementary sequences atthe 3' end
of
theDI RNAs are suspected to be responsible for
interferenceby DIparticles with the growthof
standard VSV (13, 23).
The ability ofDI particlestointerfere among
themselves correlated with the extent of
addi-tional complementary sequences atthe 3' end.
The greater thecomplementarity, thebetterthe
abilitytointerfere. Similardatawith DIparticles
obtainedfromdifferent strains ofVSV support this
conclusion
(32). This, however is the first time suchstudies have been carried out with DI particles all derived from thesame parent..Oursequencing results and thosepublished by others on several DIparticles indicate that the first 45 to 47 nucleotides at the 3' terminus of the majority of DI RNAs are identical and comple-mentary to the 5' end (40). Normal asymmetry ofplus-andminus-strand synthesis during VSV RNA replication is thought to be due to the
differentialbinding affinity for polymerase at the
3' ends of standard RNA templates (16, 22, 33).
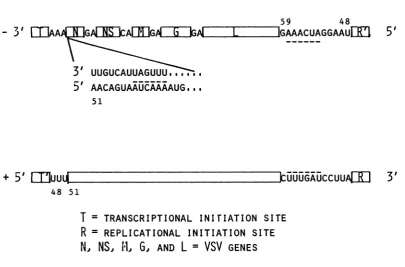
InFig. 9, these binding sequences are indicated by T and R, with R having the higher affinity for
polymerase.
With symmetrical terminal se-quences, the plus and minus DI RNAs would bothcontain R-like sequences at their 3' ends. Theresults should be equal production of both plus- and minus-strand templates.Measurement of RNA templates in thisstudy,
however, indicates approximately equal
plus-and minus-strplus-and synthesis for only DI 0.52 particles. The other two DI particles synthesize
minus-strand
RNA in excess over plus-strand RNA(60:40), but not at the same ratio (80:20) as standard VSV (27, 44). These results suggest that the asymmetry of RNA synthesis may bethe result of other factors beside polymerase binding.
If the first 45 to 47 nucleotides at either
terminus ofthegenomeaccounted for
polymer-asebinding, how could the next 12 nucleotides
be related to maturation? By examining pub-lished standard VSV sequences (summarized in
Fig.9),noncodingregionsaccountfor 50
nucle-otidesatthe3' end and 59 nucleotides atthe 5'
end (25, 39). Transcriptional initiation of the N genebeginsatposition51 from the 3' end(7, 26,
38), leaving only a triplet AAA without an
assigned function. However,atthe 5'end,after
termination ofthe coding regionfor the L
pro-tein, there are 12 nucleotidesin addition tothe terminal 47 nucleotides. SinceDI 0.52particles contain sequences at the 3' end of their RNA which are complementary to some of these
additionalnucleotides,thisregionbetween
posi-tions 48 and59,just internalto the terminal47, atthe 5' end may determine interactionsduring
morphogenesis. Becausetheinitial stepof
mor-phogenesisisencapsidationof the RNA with N
protein, the interactioncould be viewed as the
determinant forN-protein binding.
Apartof the sequencewithinthe 12
nucleo-tides, 5' AUCAAA 3', is also found in the
noncoding regionof the N mRNAjust beforethe
first AUGforinitiating translation(5'
m7AmA-CAGUAAUCAAAAUG 3'). Such ahexamer is
notfound in any othernoncodingregion of
VSV-specificRNAs. It is prematuretodecide
wheth-erthis sequenceatthe 5'endsoritscomplement
at the 3' ends determines the interaction be-tween VSV RNA and Nprotein. Nevertheless, if the hexamer and thenucleotides surrounding it affect the affinity of the association between RNA andN protein, then the same arguments
forthe polymeraseconcerning bindingaffinities
andsymmetryof sequences would hold for the
interactions of RNA and the N protein.
Al-thoughthe RNA of DI 0.52 particles has
termi-nalcomplementarityonlyforaportion of the 12
nucleotides,including the hexamer, the
extend-ed symmetrical sequence in its RNA may be enough to account for its ability to compete during encapsidation as well as during replica-tion. Blumberg et al. (B. M. Blumberg, M. Leppert, and D. Kolakofsky, Cell, in press), reasoning from a differenttack, conclude simi-larly that thesequencesresponsible for initiating N-protein binding are internal to the termini. Ourassignment of theinternalposition of these sequences differsfrom theirs; however, regula-tory sequences and binding sequences need not be the same.
Sincethe general understanding of VSV RNA
replication and encapsidation is scant, other hypotheses for interference mechanisms could be entertained. It is possible that the ability of
on November 10, 2019 by guest
http://jvi.asm.org/
59
48
- 3'
WIEAA
]AEGE]ECGAI
L
IGAAACUAGGAAUE
3'
UUGUCAUUAGUUU...
5
AACAGUAAUCAAAAUG...
51+
5'
LflUUU4
)CUUUGAUCCUEUACI
48 51
T
= TRANSCRIPTIONAL INITIATION SITER
=REPLICATIONAL
INITIATION SITEN, NS, [M, G,
ANDL
=VSV
GENESFIG. 9. Sequences at the termini of VSV RNA postulated to be responsible for RNA synthesis and
encapsidation. Sequencesaretakenfrom Schubertetal. (39), McGeoch and Dolan (25), McGeochetal. (26),and
Rose(38). T, Transcriptionalinitiation site;R,replicationalinitiation site;N,NS, M,G, and L, VSVgenes.
RNAtemplatestocircularize maycontribute in
some waytotheir abilitytobereplicated (28). If preferential replication dependsonthe stability
of circular template RNA, then the degree of
sequencecomplementarityatthetermini would correlate directly with such stability. On the otherhand, wehave made the assumptionthat all VSV particles, standard or defective, have
about the same particle-to-infectivity ratio. If the effective multiplicities for the DI particles
vary greater than threefold from our
assump-tions, thenany hypothesisbasedonthe degree
of sequence complementarity may have to be
revised.
Despitewhatevermechanisms turn out tobe
true,therearethree classes ofcontrolling
termi-nalsequencesontheRNAgenomesof VSVand
its DIparticles. Standardvirus and the DI 0.46
particle derived from the Orsay (HR) strain contain 18 nucleotides at the 3' and 5' termini
whicharecomplementaryexceptforafew base
mismatches(32).Other DIparticlescontain ter-minal RNA sequences which are completely
complementary for 45 to 48 nucleotides, as a secondgroup,and for 55to60nucleotides,asa thirdgroup (40). These three typesofterminal
sequences relate to a hierarchical interference
pattern where the extent of complementarity
results inagreaterabilitytocompeteduringthe
virus growth cycle; These results focus the
control of VSVRNAreplication and encapsida-tion onthese terminal sequences. Direct
com-petitive binding experiments with these se-quencesandtheirregulatoryproteinsareneeded
to understand how interference occurs during nucleicacidreplication and encapsidation.
ACKNOWLEDGMENTS
Thisworkwassupported by Public Health Service research
grantAI 16625 from the National Institutes of Health andby researchgrantMV-54 from the American CancerSociety.
We thankTrudy Lanman forexcellenttechnicalsupport andSuzanne Ress forthecomputer-aided preparationof this manuscript. Helen Donis-Keller, Peter Besmer,and Ian Ken-nedy kindlyprovidedtechnical adviceandmaterials.
LITERATURECITED
1. Alawne,J. C.,D.J. Kemp,andG. R. Stark. 1977. Method
fordetection ofspecificRNA inagarosegels bytransfer
to diazobenzyloxymethyl-paper and hybridization with
DNA probes. Proc. Natl. Acad. Sci. U.S.A.
74:5350-5354.
2. Colonno,R.J.,and A. K.Banedee.1976.AuniqueRNA
species involvedin initiation of vesicular stomatitisvirus
RNAtranscriptionin vitro. Cell 8:197-204.
3. Coonno, R. J., and A. K. Banerjee. 1978. Complete
5'
3'
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.504.52.458.76.343.2]220 RAO AND HUANG
nucleotide sequence of the leader RNA synthesized in vitro by vesicular stomatitis virus. Cell 15:93-101. 4. Donis-Keller, H., A. M. Maxam, and W. Gilbert. 1977.
Mapping adenines, guanines and pyrimidines in RNA. NucleicAcidsRes. 4:2527-2538.
5. Emerson, S. U., P. M. Dierks, and J. T. Parsons. 1977. In vitro synthesis of a unique RNA species by a T particlesof vesicular stomatitis virus. J. Virol. 23:708-716. 6. England, T. E., and0. C.Uhlenbeck. 1978. 3-Terminal
labeling of RNA with T4 RNAligase. Nature(London) 275:560-561.
7. Franze-Fernandez, M. T., and A, K. Banerjee. 1978. In vitro RNA transcription by the New Jersey serotype of vesicular stomatitis virus I. Characterization of the mRNA species.J. Virol.26:179-187.
8. Freeman, G. J., D. D. Rao, and A. S. Huang. 1978. Genome organization of vesicularstomatitis virus: map-ping of ts G41 and the defective interfering T particle, p. 261-270. In B. W. J. Mahy and R. D. Barry (ed.), Negative strand viruses and the host cell. Academic Press,Inc., New York.
9. Freeman, G. J., D. D. Rao, and A. S. Huang. 1979. RNA synthesis of vesicularstomatitisvirus. VIII. Oligonucleo-tides of the structural genes and mRNA. Gene 5:141-157. 10. Hagen, F. S., and A. S. Huang. 1981. Comparison of
ribonucleotide sequences from the genome of vesicular stomatitis virus and two of its defective interfering parti-cles. J. Virol. 37:363-371.
11. Holland, J. J., L. P. Villarreal, and M. Breindl. 1976. Factors involved in the generation and replication of rhabdovirus defective T particles. J.Virol. 17:805-815. 12. Huang, A.S. 1973. Defective interfering viruses. Annu.
Rev. Microbiol. 27:101-117.
13. Huang, A. S. 1977. Viral pathogenesis and molecular biology. Bacteriol. Rev. 41:811-821.
14. Huang, A. S., and D. Baltimore. 1977. Defective interfer-ing animal viruses, p.73-116. In H. Fraenkel-Conratand R. R. Wagner (ed.), Comprehensive Virology, vol. 10. Plenum Publishing Corp., New York.
15. Huang, A. S., J. W., Greenawalt, and R. R. Wagner. 1966. Defective T particles of vesicular stomatitis virus. I. Preparation, morphology, and some biologic properties. Virology 30:161-172.
16. Huang, A. S., S. P.Little,M. B. A. Oldstone, and D. D. Rao. 1978.Defectiveinterferingparticles:theireffecton gene expression and replication of vesicular stomatitis virus, p. 399-408.InG. Todaro, J. Stevens, and C. F. Fox (ed.), ICN-UCLA Symposium on Persistent Viruses. Ac-ademicPress,Inc.,New York.
17. Huang, A. S., and E. K. Manders. 1972.Ribonucleic acid synthesis ofvesicularstomatitis virus. IV. Transcription by standard virus in thepresence ofdefectiveinterfering particles.J. Virol. 9:909-916.
18. Huang, A. S., and E. L. Palma. 1974.Vesicularstomatitis virus: defectiveness and disease, p. 87-99. In W. S. Robinson and C. F. Fox (ed.), Mechanisms of virus disease. Benjamin,Inc.,Menlo Park, Calif.
19. Huang, A.S., D. D. Rao, and G.Lanman.1980.Defective
interferingparticles ofvesicular stomatitis virus:
struc-ture-function relationships. Ann. N.Y. Acad. Sci.
354:238-250.
20. Huang, A.S., and R. R. Wagner, 1966. Defective T particlesofvesicularstomatitis virus.II.Biologicrole in homologousinterference.Virology 30:173-181. 21. Kang,C. Y., T. Glimp, J. P.Clewley,and D. H. L.Bishop.
1978. Studies on the generation ofvesicular stomatitis virus (Indiana serotype) defective interfering particles.
Virology84:142-152.
22. Kolakofsky, D., M. Leppert, and L. Kort. 1978. Agenetic
map of Sendai virus DI RNAs, p. 539-553. In B. W. J. Mahy and R. D. Barry (ed.), Negative strandvirusesand the hostcell.AcademicPress,Inc., New York. 23. Leppert, M., L. Kort, and D. Kolakofsky. 1977. Further
characterization of Sendai virus DI-RNAs: a model for their generation. Cell 12:539-552.
J. VIROL.
24. Marcus, P.I.,D. L.Engelhardt, J.M.Hunt,andM. J. Sekellick.1971. Interferon action: inhibitionofvesicular stomatitis virus RNAsynthesisinducedbyvirion-bound
polymerase.Science 174:593-598.
25. McGeoch, D. J., and A. Dolan. 1979. Sequenceof200 nucleotides at the 3' terminus of the genome RNA of vesicular stomatitis virus. Nucleic Acids Res. 6:3199-3211.
26. McGeoch, D.,A.Dolan,andC. R.Pringle.1980.
Compari-sonsof nucleotide sequencesin the genomes of theNew
Jerseyand Indiana serotypes of vesicular stomatitisvirus.
J.Virol. 33:69-77.
27. Morrison, T. G., M. Stampfer, H. F. Lodish, and D. Baltimore.1975. Invitro translation of vesicular stomatitis virus messenger RNA'sandtheexistence ofa40 S"plus"
strand, p. 293-306. In B. W. J. Mahy and R. D. Barry
(ed.), Negative strand viruses. Academic Press, Inc.,
New York.
28. Naeve, C. W., C. M. Kolakofsky, and D. F. Summers. 1980.Comparisonof vesicularstomatitisvirus intracellu-lar andvirionribonucleoproteins. J. Virol.33:856-865. 29. Palma, E. L., S. M. Perlman, and A. S. Huang. 1974
Ribonucleic acid synthesis ofvesicular stomatitis virus. VI.Correlation of defectiveparticleRNAsynthesiswith standard RNAreplication.J. Mol. Biol.85:127-136. 30. Perrault, J.,andJ.J.Holland.1972.Absence of
transcrip-taseactivity andtranscription-inhibiting abilityin defec-tive interfering particles of vesicular stomatitis virus.
Virology50:159-170.
31. Perrault, J.,andJ. J.Holland.1972.Variabilityof vesicu-lar stomatitis virus autointerference with different host cells and virus serotypes. Virology 50:148-159. 32. Perrault, J., and B. L. Semler. 1979. Internal genome
deletions in twodistinct classes of defective interfering particles of vesicular stomatitis virus. Proc.Natl. Acad. Sci.U.S.A. 76:6191-6195.
33. Perrault, J., B. L. Semler, R. W. Leavitt, and J. J. Holland. 1978. Inverted complementary terminal se-quencesin defectiveinterfering particleRNAs of vesicu-larstomatitisvirus andtheirpossiblerole in autointerfer-ence, p. 527-538. In B. W. J.Mahyand R. D.Barry(ed.), Negative strand viruses and the host cell. Academic Press,Inc., New York.
34. Rao, D. D., and A. S. Huang. 1979. Synthesis of a small RNAincells coinfectedby standardanddefective
inter-feringparticles ofvesicular stomatitisvirus. Proc. Natl.
Acad.Sci. U.S.A. 76:3742-3745.
35. Rao, D. D., and A. S. Huang. 1980. RNA synthesis of vesicular stomatitis virus. X. Transcription and replica-tion bydefective interfering particles. J. Virol. 36:756-765.
36. Reichmann,M.E., D. H. L.Bishop, F. Brown, J. Crick, J.
J. Holland, C.-Y. Kang, R. Lazzarini, S. Moyer, J. Per. rault, L. Prevec, C. R. Pringle, R. R. Wagner, J. S. Youngner, and A. S. Huang. 1980. Proposal for auniform nomenclature for defective interferingviruses of vesicular stomatitis virus. J. Virol.34:792-794.
37. Relchmann, M. E., D. R. Pringle, and E. A. C. Follett.
1971. Defective particles in BHK cells infected with temperature-sensitive mutants of vesicularstomatitis vi-rus. J. Virol. 8:154-160.
38. Rose, J. K. 1980. Complete intergenic andflanking gene sequences from the genome of vesicularstomatitis virus. Cell 19:415-421.
39. Schubert, M., J. D. Keene, R. C. Herman, and R. A.
Lazzarnln. 1980. Site of the vesicular stomatitis virus
genome specifying polyadenylation and theend of the L gene mRNA. J. Virol. 34:550-559.
40. Schubert, M., J. D. Keene, and R. A.Lazzarinl. 1979. A specificinternalRNA polymeraserecognition siteof VSV RNA is involved in the generation of DI particles. Cell 18:749-757.
41. Schubert, M., J. D. Keene, R. A. Lazzarla, and S. U. Emerson. 1978. The completesequence of aunique RNA species synthesized bya DI particle.Cell15:103-112.
on November 10, 2019 by guest
http://jvi.asm.org/
42. Schubert, M., and R. A. Lazzarinl. 1981. In vivo transcrip-tion of the 5'-terminal extracistronic region of vesicular stomatitis virus RNA. J. Virol. 38:256-262.
43. SekeUlick,M.J.,and P. .Marcus. 1980. Viral interference
by defective interfering particles of vesicular stomatitis virus measured in individual cells. Virology 104:247-252.
44. Soria, M., S. P. Little,andA.S. Huang.1974. Character-izationofvesicular stomatitis virus nucleocapsids I. Com-plementary 40 S RNA molecules in nucleocapsids.
Virolo-gy61:270-280.
45. vonMagnus, P.1954. Incompleteforms of influenza virus.
Adv.Virus Res. 2:59-79.
on November 10, 2019 by guest
http://jvi.asm.org/