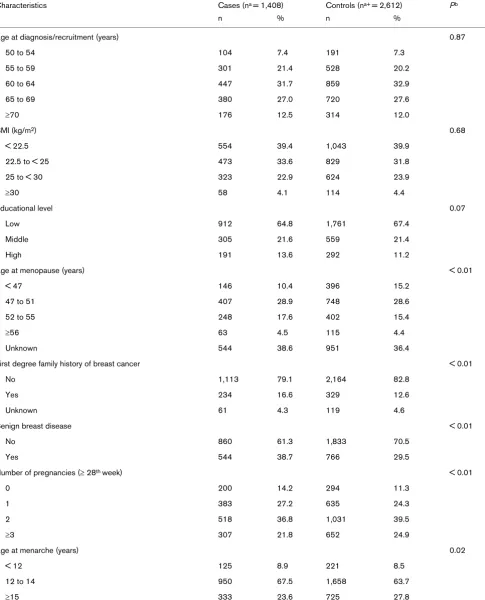
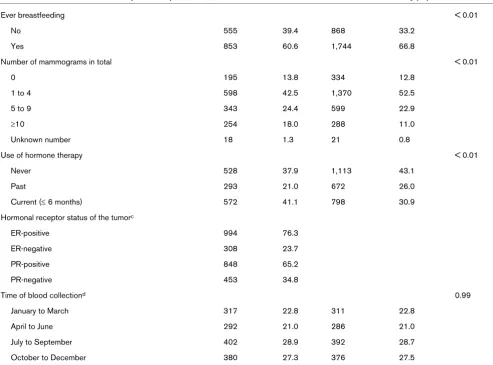
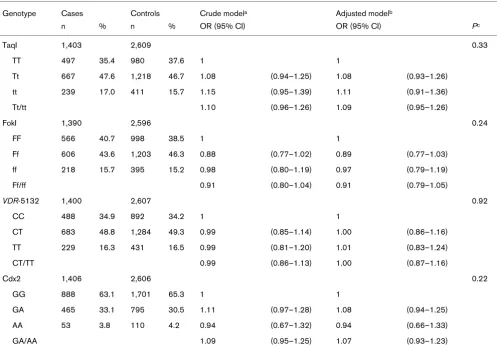
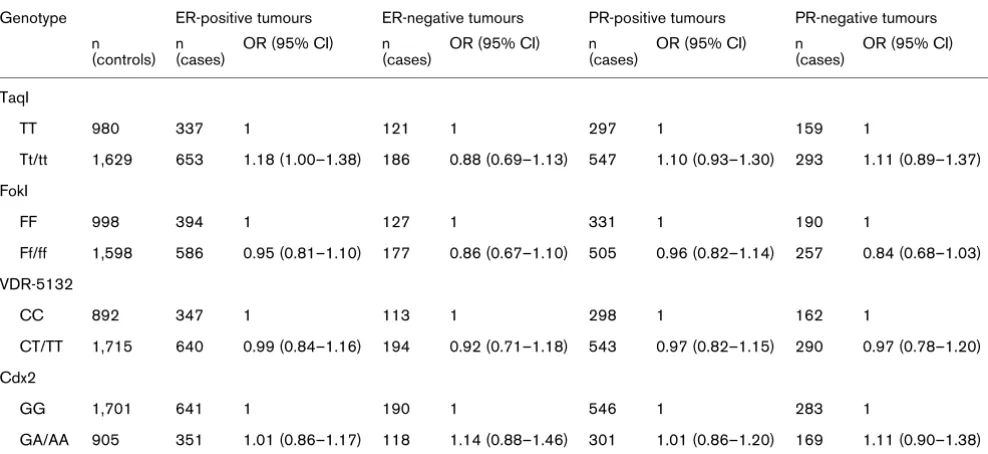
Vitamin D receptor gene polymorphisms and haplotypes and postmenopausal breast cancer risk
Full text
Figure
Related documents
Despite the limitations of the study by Mussa, the study does serve to illustrate that a methodology based on simulation, in which the necessary probability functions (e.g.
Two factors that may explain the high attendance rates at this time were the special days for lead screening that were taking place at Maari Ma and the role of an Aboriginal
While all this investment has been made for the preparation of new teachers, and meanwhile in all education undergraduate system is undertaken a very powerful reform to
Towards the end of A.L. Kennedy’s most recent work, The Blue Book
To address this gap in research, the objectives of this study are to examine (1) the knowledge of technology, (2) perceived usefulness of technology, (3) feature preferences
Even otherwise, we believe that our work substan- tially differs from all of the above attempts because we focus primarily on the performance of the cipher in the link layer
Higher improvement level was observed in distal sensory and motor latency, volitional activity and pinch power in case group compared to control group.. At the same time
In vitro anti-lithiatic activity of the extracts was demonstrated in terms of inhibition of calcium oxalate formation by the extracts in the presence and absence
