JOURNAL OFVIROLOGY, Feb. 1995, p. 773–778 Vol. 69, No. 2 0022-538X/95/$04.0010
Copyrightq1995, American Society for Microbiology
Sequence Evolution of the Hypervariable Region in the Putative
Envelope Region E2/NS1 of Hepatitis C Virus Is Correlated
with Specific Humoral Immune Responses
LEEN-JANVANDOORN,1* INUSHA CAPRILES,1GEERT MAERTENS,2ROBERT DELEYS,2
KENNETH MURRAY,3TON KOS,4HUUB SCHELLEKENS,5ANDWIM QUINT1
Innogenetics NV, Ghent, Belgium2; University of Edinburgh, Edinburgh, Scotland3; and
TNO Primate Centre, Rijswijk,4and Department of Molecular Biology1and
Department of Immunology and Infectious Diseases,5Diagnostic
Centre SSDZ, Delft, The Netherlands
Received 18 July 1994/Accepted 4 November 1994
Sequence evolution of the hypervariable region 1 (HVR1) in the N terminus of E2/NS1 of hepatitis C virus (HCV) was studied retrospectively in six chimpanzees inoculated with the same genotype 1b strain, containing a unique predominant HVR1 sequence. Immediately after inoculation, all animals contained the same HVR predominant sequence. Two animals developed an acute self-limiting infection. Anti-HVR1 immunoglobulin G (IgG) was produced 40 to 60 days after inoculation and rapidly disappeared after normalization of transami-nases. Another chimpanzee, previously infected with human immunodeficiency virus type 1, showed a delayed response to HVR1 epitopes after superinfection with HCV. No sequence variation of HVR1 was observed in these two animals during the transient viremia in the acute phase. Three other chimpanzees developed a chronic HCV infection. During follow up, sequence evolution occurred in two animals and their anti-HVR1 response remained at varying but detectable levels. The first mutations occurred immediately after the production of anti-HVR1 during the acute phase. However, IgM anti-HVR1 was not detectable. Remarkably, HVR1 sequences remained conserved for more than 6 years in another chronically infected animal. This correlated with the complete absence of detectable anti-HVR1 during this period. Seven years after inoculation, anti-HVR1 IgG was produced and coincided with an HVR1 alteration. These results strongly suggest the involvement of neutralizing anti-HVR antibodies in sequence evolution of HVR1 through immune selection.
Hepatitis C virus (HCV) is associated with the majority of posttransfusion and sporadic viral hepatitis cases. The virus, classified as a relative of the flavi- and pestiviruses, possesses a positive-sense, single-stranded RNA genome of approximately
9,400 nucleotides (6, 7). Besides 59and 39untranslated regions,
the viral genome encodes a capsid protein (C), two putative envelope proteins (E1 and E2/NS1), and four nonstructural proteins (NS2 to NS5). NS5 encodes the putative RNA-depen-dent RNA polymerase.
Analysis of numerous HCV isolates revealed significant se-quence heterogeneity among different viral isolates and re-sulted in the identification of at least six major genotypes and several subtypes (24). Sequence variation is not evenly
distrib-uted along the viral genome. The 59 untranslated region is
highly conserved and is therefore generally used as a target for diagnosis of viremia by PCR. The core, E1, and NS5 regions are more heterogeneous (1, 24, 25). The most variable region was identified in the N terminus of the E2/NS1 region (13, 29). This hypervariable region (HVR1) of approximately 27 amino acids (residues 384 to 410) has been shown to be highly vari-able among and within different patients (13, 17, 29, 30). HVR1 can be used to identify individual HCV isolates, which is of particular interest for epidemiological studies (32).
Sequence variation in HVR1 has been observed during chronic HCV infections in humans (9, 12, 16) and chimpanzees (22, 28) and appears to be comparable to that observed in the human immunodeficiency virus type 1 (HIV-1) gp120 V3
do-main (31). Furthermore, the HVR1 dodo-main appears to be structurally flexible and contains linear B-cell epitopes that are well exposed at the outside of the virus and probably subjected to immune selection (18, 19, 23, 26, 31). According to the immune selection hypothesis, neutralizing antibodies to HVR1 epitopes cause positive selection of escape mutants, which may play a role in the mechanism of persistent viral infection (23, 31). Moreover, sequence variation of HVR1 may have impor-tant clinical implications, such as the efficacy of interferon treatment (21).
We investigated the relationship between sequence evolu-tion of HVR1 of a single well-defined HCV type 1b inoculum and the humoral immune response to the corresponding epitopes during acute and chronic infections in six chimpan-zees.
MATERIALS AND METHODS
Chimpanzees.Experimental HCV infections in six chimpanzees (Phil, Peggy, Hans, Coen, Socrates, and Sophie) were performed between 1982 and 1987 and have been described in detail earlier (28). Briefly, chimpanzees Phil, Peggy, and Hans were inoculated with different quantities of an HCV inoculum, derived from another chimpanzee (27). Sequence analysis showed that this inoculum contained HCV genotype 1b. HCV was transferred from chimpanzee Phil to chimpanzees Socrates and Coen, and acute-phase plasma from Socrates was further passaged into chimpanzee Sophie. All chimpanzees were inoculated intravenously. Chimpanzees Phil, Peggy, and Hans developed a chronic infec-tion, whereas Coen, Socrates, and Sophie experienced only acute infection. Coen had been previously infected with HIV-1. These HCV infections were initially monitored by measurements of alanine aminotransferase (ALT) andg -glutamyl-transferase (g-GT) in serum.
Sequence analysis.Amplification primers and direct sequencing procedures have been described in detail elsewhere (28). Briefly, HCV RNA was isolated from serum with the guanidinium thiocyanate-phenol method (4), reverse tran-scribed into cDNA, and subsequently amplified by nested PCR. One of the * Corresponding author. Mailing address: Diagnostic Centre SSDZ,
Dept. of Molecular Biology, PO Box 5010, 2600 GA Delft, The Neth-erlands. Phone: 31-15-604556. Fax: 31-15-604550.
773
on November 9, 2019 by guest
http://jvi.asm.org/
primers in the nested reaction was biotinylated, allowing preparation of single-stranded DNA suitable for direct sequencing of the PCR product (14).
Anti-HVR1 antibody detection.Two overlapping biotinylated peptides cover-ing the hypervariable region were synthesised on a Millipore 9050 PepSynthe-sizer. Peptide A (Bio-GSTYVTGGASGRSTHGLVSLF) covers amino acid res-idues 384 to 403, and peptide B (Bio-GRSTHGLVSLFNLGAQQKVQLINT) corresponds to residues 394 to 417. Peptides were dissolved in dimethyl sulfox-ide, diluted with phosphate-buffered saline (PBS) to 10 mg/ml, and stored at 2208C. Dilutions of peptides were prepared in PBS with 5% glycerol.
An enzyme-linked immunosorbent assay (ELISA) was developed on the basis of biotinylated peptides A and B. Streptavidin (Boehringer, Mannheim, Ger-many) was coated onto 96-well Maxisorp plates (Nunc, Roskilde, Denmark) by incubating 200 ml (3 mg/ml) in each well for 1 h at 378C.
Plates were washed three times with PBS, and 100 ml of the peptide solution (1 mg/ml) was added. Dilution experiments showed that 100 ng of peptide per well was sufficient to obtain an optimal signal. For each serum sample to be tested, duplicate wells were coated with either peptide A or B or without peptide for 1 h at 378C. (Each serum sample requires six wells.) After three washes with PBS, the wells were blocked with PBSTG (PBS containing 0.05% Tween 20 and 0.5% gelatin) for 1 h at 378C.
Serum samples were diluted 1:100 in PBSTG, and 100 ml was added to the wells and incubated for 1 h at 378C. The wells were washed eight times with PBST (PBS with 0.5% Tween 20), and conjugate was added. Goat anti-human immunoglobulin G (IgG) peroxidase (produced at the Diagnostic Centre SSDZ) was appropriately diluted in PBSTG, added to the wells, and incubated at 378C. After 1 h, the wells were washed eight times with PBST, and the substrate (3,39,5,59-tetramethylbenzidine [TMB]; Aldrich, Steinheim, Germany) was added. The reaction was stopped after 30 min by adding 100 ml of 3 N H2SO4–4 M urea. Color development was measured in a plate reader at 450 nm.
Mean optical densities from duplicate wells containing either peptide A or B were calculated, and the mean optical density of the control wells (no peptide) was subtracted. If the variation between two duplicate wells was higher than 10%, the samples were retested. Repeated samples gave consistent results in the same range, and large discrepancies between repeated samples were never ob-served. Serum samples from each chimpanzee were all processed on the same day, and each plate contained a positive control serum sample.
RESULTS
Sequence evolution of the hypervariable region HVR1 was studied retrospectively in six chimpanzees infected with a unique genotype 1b HCV isolate. HCV RNA was reverse transcribed into cDNA, amplified, and subjected to direct
se-quence analysis. HVR1 nucleotide and amino acid sese-quences are shown in Fig. 1 and 2, respectively. At some positions, sequence ambiguity was observed, seen as double bands on the sequencing gel. The parental HVR1 variant, present in the source plasma, was present in all chimpanzees shortly after inoculation. For chimpanzees Coen, Socrates, and Sophie, which developed acute hepatitis, several serum samples throughout the acute phase were analyzed, and no sequence alterations were observed during the transient period of vire-mia.
From the chronically infected animals, multiple sequences were determined in samples obtained up to 10 years after infection. During the acute phase in Hans and Peggy, the parental consensus sequence of the inoculum rapidly changed and variants were observed within several months. HCV RNA from chimpanzee Peggy was analyzed at multiple times during the acute phase of infection. As shown in Fig. 1 and 2, the parental sequence was detected until 62 days after infection. At 76 days after infection, a double band (C/T) appeared at po-sition 1188, but this mutation is silent. At 92 days after infec-tion, the first non-silent mutation (Leu-405 to Pro) was de-tected at position 1214, and at 125 days after infection, the second non-silent mutation (Val-411 to Ile) was detected at position 1231. Later during infection, additional mutations were detected. However, in chimpanzee Phil, no sequence alterations were detected between December 1982 and No-vember 1989. In the 1991 isolate, a single amino acid alteration (Gln-409 to Leu) was observed. In the 1989 sample, sequence ambiguity suggested the possible presence of both Ile and Val at position 415.
Antibodies against linear HVR1 epitopes, presented as overlapping peptides A and B, were determined in serial serum samples from the six chimpanzees. Peptide sequences were based on the parental sequence as detected in the original inoculum and in each chimpanzee shortly after inoculation.
FIG. 1. Alignment of nucleotide sequences from the hypervariable region (nucleotides 1140 to 1244) in the E2/NS1 region at various times during HCV infection. Nucleotide identity is indicated by dashes, Y5C or T, R5G or A, K5G or T, M5C or A, S5G or C, W5A or T. The numbers in parentheses indicate sampling time in days after inoculation.
on November 9, 2019 by guest
http://jvi.asm.org/
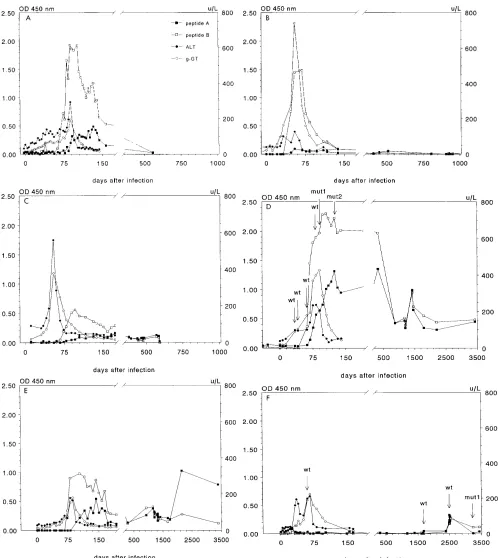
Results are shown in Fig. 3. Anti-peptide A antibody levels were lower than anti-peptide B levels in the majority of sam-ples.
In chimpanzees Socrates and Sophie (Fig. 3A and B), show-ing acute self-limitshow-ing hepatitis, high levels of anti-HVR1 IgG antibodies were produced 40 to 60 days after inoculation, co-inciding with the rise of both liver-specific enzymes ALT and g-GT. Anti-HVR1 IgG levels rapidly diminished after normal-ization of the serum ALT levels and dropped below the detec-tion limit approximately 150 to 200 days after infecdetec-tion. In chimpanzee Coen (Fig. 3C), low anti-HVR1 antibody levels became detectable 60 to 80 days after infection. Compared with those of the other animals, the antibody response seemed delayed, since it occurred after normalization rather than dur-ing elevation of liver enzymes. Eventually, the modest antibody levels decreased to just above the detection limit.
In chronically infected chimpanzees Peggy and Hans (Fig. 3D and E), high levels of anti-HVR1 antibodies also became rapidly detectable after inoculation, again coinciding with liver enzyme elevations. However, after normalization of these en-zyme levels, fluctuating levels of anti-HVR1 remained present for prolonged periods.
Chimpanzee Phil did not develop antibodies against pep-tides A and B throughout the acute phase of the disease (Fig. 3F). Low levels of anti-HVR1 antibodies were first detected in 1989, more than 6 years after inoculation.
DISCUSSION
The sequence evolution of the hypervariable region in the 59
terminus of E2/NS1 encoding one of the putative envelope proteins was studied in a well-characterized group of six chim-panzees infected with HCV.
RNA genomes have a relatively high mutation rate, because of errors of the RNA-dependent RNA polymerase during copying of the template RNA. An RNA virus population exists as a distribution of mutant genomes termed viral quasispecies, comprising coexisting mutant genomes with a consensus or master sequence (8, 12, 20). If a specific mutant has a higher competitive fitness than the parental sequence, or if selective forces such as an immune response evolve, this can influence the distribution between different viral variants and will con-sequently cause a change in the consensus sequence. This two-step process of mutant generation followed by selection results in antigenic diversification of RNA viruses. The im-mune response is assumed to select variants directly, although alternative mechanisms may also contribute, e.g., selection of variants that interact better with cell receptors (8).
Recently, Weiner and coworkers suggested that HVR1 do-mains of HCV are subjected to immune selection (31). Anti-bodies against this presumably well-exposed epitope of the putative viral envelope protein (23, 26, 31) possibly neutralize a specific virus variant, resulting in the selection of escape mutants whose evolution is favored by a transient failure of the host immune system to neutralize the newly emerging variant. This escape mutant rapidly becomes predominant while the replication of the parental variant is efficiently inhibited. Dif-ferent HVR1 sequences were detected during difDif-ferent epi-sodes of chronic hepatitis, and alteration of the sequence was accompanied by the presence of specific antibodies to corre-sponding HVR1 peptides (31).
We studied the relationship between sequence evolution of the hypervariable region and the humoral immune response to the corresponding epitopes in six chimpanzees that had been infected with an identical HCV inoculum (28). The availability of many serial serum samples from these animals allowed the precise monitoring of sequence evolution and antibody forma-tion. Sequences were determined by direct sequence analysis of PCR-amplified cDNA fragments, avoiding the possible sam-pling errors encountered by analysis of individual clones. Also, errors by reverse transcriptase and Taq DNA polymerase are unlikely to be detected. Direct sequence analysis allows detec-tion of the master sequence and is particularly useful for mea-suring changes in this consensus sequence of a population of genomes. On the other hand, direct sequencing does not pro-vide detailed information about minor variants in the popula-tion.
[image:3.612.58.299.89.370.2]HVR1 sequence alterations were not detected during the transient viremia in acutely infected chimpanzees Socrates and Sophie, although humoral immune responses against HVR1 were rapidly detectable after inoculation, coinciding with ele-vated levels of ALT and g-GT. The period of viremia was probably too brief to allow the selection of escape mutants. By the time the anti-HVR1 antibodies reach a significant level, the viral RNA has already been cleared from the circulation (28). Chimpanzee Coen experienced an HCV infection with tran-sient viremia. After normalization of ALT levels, HCV RNA was undetectable for at least 2 years (28). However, a single sample obtained 10 years after infection did contain detectable HCV RNA. Therefore, we conclude that chimpanzee Coen was also chronically infected. Unfortunately, additional sam-ples were not available for HCV analysis. Because this animal had been previously infected with HIV-1, viral interference
FIG. 2. Alignment of deduced amino acid sequences of hypervariable region (underlined) (amino acids 381 to 414) in E2/NS1 region. Amino acid identity is indicated by dashes. Italics indicate the possibility of the presence of both the original and the italicized amino acid, because of nucleotide sequence ambiguity. Asterisks indicate the possibility of multiple amino acids. The numbers in pa-rentheses indicate sampling time in days after inoculation.
VOL. 69, 1995 SEQUENCE EVOLUTION OF HCV HYPERVARIABLE REGION 775
on November 9, 2019 by guest
http://jvi.asm.org/
between HIV-1 and HCV (11) might be important. Remark-ably, the anti-HVR1 antibody response in Coen clearly oc-curred after normalization of ALT and g-GT, whereas in other animals, it coincided with elevated levels of these enzymes, suggesting immunomodulatory effects of HIV-1.
[image:4.612.57.555.67.625.2]Chimpanzee Peggy and Hans showed sequential alterations in the HVR1 region during chronic HCV infections. Most of the nucleotide alterations are replacement mutations. The pu-tative conserved amino acids Thr-385, Gly-406, Gln-409, and Leu-413 (19) also remained conserved in these chimpanzees.
FIG. 3. Production of antibodies to peptides A and B, corresponding to epitopes of hypervariable region in original inoculum, during HCV infection. ALT and g-GT levels measured during the acute phase of infection are also shown. (D and F) The presence of the parental (wild type [wt]) as well as a mutant (mut) HVR1 consensus sequence is indicated by labeled arrows. (A) Socrates; (B) Sophie; (C) Coen; (D) Peggy; (E) Hans; (F) Phil. Symbols are defined in panel A. OD450, optical density at 450 nm.
on November 9, 2019 by guest
http://jvi.asm.org/
The production of HVR1 specific antibodies and correspond-ing sequence alterations are in accordance with earlier reports (18, 19, 23). Anti-HVR1 antibodies remained present at vari-able levels, which may be partly explained by cross-reactive anti-HVR1 antibodies, and suggest the continuing stimulation of the immune system with HVR1 epitopes. The rapid de-crease in anti-HVR1 IgG in the acutely infected animals So-phie and Socrates was remarkable, since IgG responses to other viruses usually remain detectable during much longer periods. During the first vaccination experiments in chimpan-zees (5), the levels of antibodies to E1 and E2 also rapidly declined after immunization with recombinant protein. This apparent absence of long-term humoral immune response against HCV envelope epitopes may be very important for the development of an effective vaccine.
In chimpanzee Phil, chronic HCV infection without any de-tectable humoral immune response is associated with the com-plete absence of hypervariability of HVR1 epitopes for more than 6 years. Because of the absence of a specific immune response against HVR1 epitopes, there are no mutants that have a higher competitive fitness. The reason underlying this unresponsiveness of Phil remains unclear but could be associ-ated with the histocompatibility (HLA) genotype. An alterna-tive explanation might be that immunological escape is in-duced only by an effective initial antiviral immune response upon primary infection, as has been suggested for HIV-1 (33). Chimpanzee Phil was inoculated with a very low dose of virus (28). Viremia was first detected in Phil 32 days after infection, whereas in chimpanzees Socrates and Sophie HCV RNA was detectable within a week. Also, the overall anti-HCV response in Phil seemed inefficient during the acute phase of infection. Between 1989 and 1991, a single amino acid change (Gln to Leu at position 409) was observed. This can possibly be related to low levels of anti-HVR1 antibodies detected around 2,500 days after inoculation. At 2,527 days after infection, the pa-rental sequence was still detected. Unfortunately, the number of available serum samples from this period is very limited. Although the coincidence between the two observations is re-markable, we have failed to identify the cause that triggered the production of anti-HVR1 and the subsequent selection of an escape mutant 7 years after inoculation.
Analysis of the acute-phase samples of chimpanzee Peggy showed that the first HVR1 mutant emerged immediately after the production of anti-HVR1 IgG. This suggests that sequence alterations in HVR1 are not the result of rapid viral adaptation to a new host but occur only after the development of a specific antibody response. The results for chimpanzee Phil also show that sequence variation and escape mutants in the HVR1 are not essential for the development of a chronic HCV infection. Also, the efficacy of the anti-HVR1 response seems unrelated to complete clearance of the virus. The exact mechanisms of acute resolving versus persistent HCV infection remain ob-scure.
The biological function of the HVR1 region remains un-clear. It is uncertain whether sequence alterations in HVR1 are associated with long-term adaptation of the virus to a specific host with changes in cell tropism and viral entry, as has been described for the HIV-1 V3 region (2, 3, 10, 15). It might be speculated that chronic HCV infection leads to a slow adaptation to the host, with increasing clinical symptoms, such as cirrhosis and eventually hepatocellular carcinoma. Ran-domly generated HVR1 mutants may not only be subjected to a rapidly acting immune selection mechanism, but antigenic variation could also be driven by adaptive fitness of the virus to the host.
In conclusion, our results provide strong evidence for an
immune selection mechanism of HVR1 variants in HCV. How-ever, antigenic variation of HVR1 is not obligatory for the development of a chronic HCV infection. Since anti-HVR1 antibodies are considered to be neutralizing, modulation of the immune responses against these epitopes may have important implications for vaccination strategies.
ACKNOWLEDGMENTS
We acknowledge J. G. Beljaars for assistance with chimpanzee se-rum stocks and data records and P. Herbrink and H. v.d. Munckhof for valuable suggestions. A. P. Plaisier is acknowledged for assistance in preparation of the figures.
REFERENCES
1. Bukh, J., R. H. Purcell, and R. H. Miller. 1993. At least 12 genotypes of hepatitis C virus predicted by sequence analysis of the putative E1 gene of isolates collected world-wide. Proc. Natl. Acad. Sci. USA 90:8234–8238. 2. Chesebro, B., J. Nishio, S. Perryman, A. Cann, W. Obrien, I. S. Chen, and
K. Wehrly.1991. Identification of human immunodeficiency virus envelope gene sequences influencing viral entry into CD4-positive HeLa cells, T-leukemia cells, and macrophages. J. Virol. 65:5782–5789.
3. Chesebro, B., K. Wehrly, J. Nishio, and S. Perryman. 1992. Macrophage-tropic human immunodeficiency virus isolates from different patients exhibit unusual V3 envelope sequence homogeneity in comparison with T-cell-tropic isolates: definition of critical amino acids involved in cell tropism. J. Virol. 66:6547–6554.
4. Chomczynski, P., and N. Sacchi. 1987. Single step method of RNA isolation by guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162:152–159.
5. Choo, Q.-L., G. Kuo, R. Ralston, A. J. Weiner, D. Chien, G. Van Nest, J. Han, K. Berger, K. Thudium, C. Kuo, J. Kansopon, J. McFarland, A. Tabrizi, K. Ching, B. Moss, L. B. Cummins, M. Houghton, and E. Muchmore.1994. Vaccination of chimpanzees against infection by the hepatitis C virus. Proc. Natl. Acad. Sci. USA 91:1294–1298.
6. Choo, Q.-L., G. Kuo, A. J. Weiner, L. R. Overby, D. W. Bradley, and M. Houghton.1989. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 244:359–362.
7. Choo, Q.-L., K. H. Richman, J. H. Han, K. Berger, C. Lee, C. Dong, C. Gallegos, D. Coit, A. Medina-Selby, P. J. Barr, A. J. Weiner, D. W. Bradley, G. Kuo, and M. Houghton.1991. Genetic organization and diversity of the hepatitis C virus. Proc. Natl. Acad. Sci. USA 88:2451–2455.
8. Domingo, E., J. Diez, M. A. Martinez, J. Hernandez, A. Hoguin, B. Borrego, and M. G. Mateu.1993. New observations on antigenic diversification of RNA viruses: antigenic variation is not dependent on immune selection. J. Gen. Virol. 74:2039–2045.
9. Enomoto, N., N. Sakamoto, M. Kurosaki, F. Marumo, and C. Sato. 1993. The hypervariable region of the HCV genome changes sequentially during the progression of acute HCV infection to chronic hepatitis. J. Hepatol. 17:415–416.
10. Fouchier, R. A. M., M. Groenink, A. Kootstra, M. Tersmette, H. G. Huis-man, F. Miedema, and H. Schuitemaker.1992. Phenotype-associated se-quence variation in the third variable domain of the human immunodefi-ciency virus type 1 gp120 molecule. J. Virol. 66:3178–3187.
11. Giovannini, M., A. Tagger, M. L. Ribero, G. Zuccoti, L. Pogliani, A. Grossi, P. Ferroni, and A. Fiocchi.1990. Maternal-infant transmission of hepatitis C virus and HIV infections: a possible interaction. Lancet 336:1166. 12. Higashi, Y., S. Kakumu, K. Yoshioka, T. Wakita, M. Mizokami, K. Ohba, Y.
Ito, T. Ishikawa, M. Takayanagi, and Y. Nagai.1993. Dynamics of genome change in the E2/NS1 region of hepatitis C virus in vivo. Virology 197:659– 668.
13. Hijikata, M., N. Kato, Y. Ootsuyama, M. Nakagawa, S. Ohkoshi, and K. Shimotohno.1991. Hypervariable regions in the putative glycoprotein of hepatitis C virus. Biochem. Biophys. Res. Comm. 175:220–228.
14. Hultman, T., S. Ståhl, E. Hornes, and M. Uhle´n. Direct solid phase sequenc-ing of genomic and plasmid DNA ussequenc-ing magnetic beads as solid support. Nucleic Acids Res. 17:4937–4946.
15. Hwang, S. S., T. J. Boyle, H. K. Lyerly, and B. R. Cullen. 1991. Identification of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science 253:71–74.
16. Kato, N., Y. Ootsuyama, S. Ohkoshi, T. Nakazawa, H. Sekiya, M. Hijikata, and K. Shimotohno.1992. Characterization of hypervariable regions in the putative envelope proteins of hepatitis C virus. Biochem. Biophys. Res. Comm. 189:119–127.
17. Kato, N., Y. Ootsuyama, T. Tanaka, M. Nakagawa, T. Nakazawa, K. Mu-raiso, S. Ohkoshi, M. Hijikata, and K. Shimotohno.1992. Marked sequence diversity in the putative envelope proteins of hepatitis C viruses. Virus Res. 22:107–123.
18. Kato, N., H. Sekiya, Y. Ootsuyama Y, T. Nakazawa, M. Hijikata, S. Ohkoshi, and K. Shimotohno.1993. Humoral immune response to hypervariable
re-VOL. 69, 1995 SEQUENCE EVOLUTION OF HCV HYPERVARIABLE REGION 777
on November 9, 2019 by guest
http://jvi.asm.org/
gion 1 of the putative envelope glycoprotein (gp70) of hepatitis C virus. J. Virol. 67:3923–3930.
19. Lesniewski, R. R., K. M. Boardway, J. M. Casey, S. M. Desai, S. G. Devare, T. K. Leung, and I. K. Mushahwar. 1993. Hypervariable 59 terminus of hepatitis C virus E2/NS1 encodes antigenically distinct variants. J. Med. Virol. 40:150–156.
20. Martell, M., J. I. Esteban, J. Quer, J. Genesca`, A. J. Weiner, R. Esteban, J. Guarida, and J. Gomez. 1992. Hepatitis C virus (HCV) circulates as a population of different but closely related genomes: quasispecies nature of HCV genome distribution. J. Virol. 66:3225–3229.
21. Okada, S. I., Y. Akahane, H. Suzuki, H. Okamoto, and S. Mishiro. 1992. The degree of variability in the aminoterminal region of the E2/NS1 protein of hepatitis C virus correlates with responsiveness to interferon therapy in viremic patients. Hepatology 16:619–624.
22. Okamoto, H., M. Kojima, S.-I. Okada, H. Yoshizawa, H. Iizuka, T. Tanaka, E. E. Muchmore, D. A. Peterson, Y. Ito, and S. Mishiro.1992. Genetic drift of hepatitis C virus during an 8.2 year infection in a chimpanzee: variability and stability. Virology 190:894–899.
23. Sekiya, H., N. Kato, Y. Ootsuyama, T. Nakazawa, K. Yamauchi, and K. Shimotohno.1994. Genetic alterations of the putative envelope proteins encoding region of the hepatitis C virus in the progression to relapsed phase from acute hepatitis: humoral immune response to hypervariable region 1. Int. J. Cancer 57:664–670.
24. Simmonds, P., A. Alberti, H. J. Alter, et al. 1994. A proposed system for nomenclature of hepatitis C viral genotypes. Hepatology 19:1321–1324. 25. Stuyver, L., W. Van Arnhem, A. Wyseur, F. Hernandez, and G. Maertens.
1994. Classification of hepatitis C viruses based on phylogenetic analysis of the E1 and NS5b regions and identification of 5 new subtypes. Proc. Natl. Acad. Sci. USA 91:10134–10138.
26. Taniguchi, S., H. Okamoto, M. Sakamoto, M. Kojima, F. Tsuda, T. Tanaka, E. Munekata, E. E. Muchmore, D. A. Peterson, and S. Mishiro.1993. A structurally flexible and antigenically variable N-terminal domain of the
hepatitis C virus E2/NS1 protein: implication for an escape from antibody. Virology 195:297–301.
27. Tsiquaye, K. N., R. G. Bird, G. Tovey, R. J. Wyke, R. Williams, and A. J. Zuckerman.1980. Further evidence of cellular changes associated with non-A, non-B hepatitis. J. Med. Virol. 5:63–71.
28. van Doorn, L. J., W. G. V. Quint, K. Tsiquaye, J. Voermans, D. Paelinck, T. Kos, G. Maertens, H. Schellekens, and K. Murray.1994. Longitudinal anal-ysis of Hepatitis C virus infection and genetic drift of the hypervariable region. J. Infect. Dis. 169:1226–1235.
29. Weiner, A. J., M. J. Brauer, J. Rosenblatt, K. H. Richman, J. Tung, K. Crawford, F. Bonino, G. Saracco, Q.-L. Choo, M. Houghton, and J. H. Han. 1991. Variable and hypervariable domains are found in the regions of HCV corresponding to the flavivirus envelope and NS1 proteins and the pestivirus envelope glycoproteins. Virology 180:842–848.
30. Weiner, A. J., C. Christopherson, J. E. Hall, F. Bonino, G. Saracco, M. Brunetto, K. Crawford, C. D. Marion, K. A. Crawford, S. Venkatakrishna, T. Miyamura, J. McHutchinson, T. Cuypers, and M. Houghton.1991. Sequence variation in hepatitis C viral isolates. J. Hepatol. 13(Suppl. 4):S6–S14. 31. Weiner, A. J., H. M. Geysen, C. Christopherson, J. E. Hall, T. J. Mason, G.
Saracco, F. Bonino, K. Crawford, C. D. Marion, K. A. Crawford, M. Bru-netto, P. J. Barr, T. Miyamura, J. McHutchinson, and M. Houghton.1992. Evidence for immune selection of hepatitis C virus (HCV) putative envelope glycoprotein variants: potential role in chronic HCV infections. Proc. Natl. Acad. Sci. USA 89:3468–3472.
32. Weiner, A. J., M. M. Thaler, K. Crawford, K. Ching, J. Kansopon, D. Y. Chien, J. E. Hall, F. Hu, and M. Houghton.1993. A unique, predominant hepatitis C virus variant found in an infant born to a mother with multiple variants. J. Virol. 67:4365–4368.
33. Zhang, L. Q., P. MacKenzie, A. Cleland, E. C. Holmes, A. J. Leigh Brown, and P. Simmonds.1993. Selection for specific sequences in the external envelope protein of human immunodeficiency virus type 1 upon primary infection. J. Virol. 67:3345–3356.