Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Viral Persistence, Antibody to E1 and E2, and Hypervariable
Region 1 Sequence Stability in Hepatitis C
Virus-Inoculated Chimpanzees
SUZANNE E. BASSETT,1,2DAVID L. THOMAS,3KATHLEEN M. BRASKY,4 ANDROBERT E. LANFORD1,2*
Department of Virology and Immunology1and Department of Laboratory and Animal Medicine,4Southwest Foundation for Biomedical Research, San Antonio, Texas 78227; Department of Microbiology, University of Texas
Health Science Center at San Antonio, San Antonio, Texas 782842; and Division of Infectious Diseases, The Johns Hopkins School of Medicine, Baltimore, Maryland3
Received 22 May 1998/Accepted 26 October 1998
The relationship of viral persistence, the immune response to hepatitis C virus (HCV) envelope proteins, and envelope sequence variability was examined in chimpanzees. Antibody reactivity to the HCV envelope proteins E1 or E2 was detected by enzyme-linked immunosorbent assay (ELISA) in more than 90% of a human serum panel. Although the ELISAs appeared to be sensitive indicators of HCV infection in human serum panels, the results of a cross-sectional study revealed that a low percentage of HCV-inoculated chimpanzees had detectable antibody to E1 (22%) and E2 (15%). Viral clearance, which was recognized in 28 (61%) of the chimpanzees, was not associated with an antibody response to E1 or E2. On the contrary, antibody to E2 was observed only in viremic chimpanzees. A longitudinal study of animals that cleared the viral infection or became chronically infected confirmed the low level of antibody to E1, E2, and the HVR-1. In 10 chronically infected animals, the sequence variation in the E2 hypervariable region (HVR-1) was minimal and did not coincide with antibody to E2 or to the HVR-1. In addition, low nucleotide and amino acid sequence variation was observed in the E1 and E2 regions from two chronically infected chimpanzees. These results suggest that mechanisms in addition to the emergence of HVR-1 antibody escape variants are involved in maintaining viral persistence. The signifi-cance of antibodies to E1 and E2 in the chimpanzee animal model is discussed.
Hepatitis C virus (HCV) infections represent a serious health problem. A vaccine protective against HCV infection is not currently available, and antiviral treatments are ineffective in the majority of HCV-infected patients. Current estimates suggest that as many as 85% of HCV-infected individuals re-main persistently infected, and chronic HCV infection is asso-ciated with cirrhosis and hepatocellular carcinoma (5, 6, 37).
HCV infection appears to persist despite the presence of virus-specific cytotoxic T lymphocytes (CTL) and circulating antibodies to HCV proteins (3, 12, 16). The HCV structural proteins include the capsid and two envelope glycoproteins, E1 and E2. Several hypervariable regions (HVR) are present within the envelope glycoproteins and may facilitate the main-tenance of persistent infection (10, 15, 23, 25, 50). The most significant divergence has been observed in the first HVR (HVR-1) within E2. Since the HVR-1 may be a dominant neutralizing epitope (19), the presence within an individual of heterogeneous populations of virions, or quasispecies, may explain why HCV-specific CTL and antibodies are not suffi-cient to clear infection, since multiple variant genomes contin-uously escape neutralization (18). A greater understanding of the pathogenesis of HCV may facilitate the development of vaccines and antiviral treatments that are more-efficacious.
HCV pathogenesis is difficult to study, since small-animal models and conventional tissue culture systems have not been established. Currently, chimpanzees serve as the only animal model for HCV infection. The frequency of persistent
infec-tion in chimpanzees and humans appears to differ. Examina-tion of the virological outcome in a large cohort of HCV-inoculated chimpanzees revealed that an unexpectedly high percentage of chimpanzees cleared the virus (61%) based on reverse transcriptase (RT)-PCR negativity (7). Since an anti-body response elicited against the envelope protein has been proposed to be important for neutralization and clearance of the virus, we have examined HCV-inoculated animals for an-tibody reactivity to the envelope proteins and sequence vari-ability in the envelope domain. The results revealed that (i) a low percentage of infected chimpanzees responded to E1 and E2, (ii) viral clearance did not appear to be associated with an antibody response to E1 or E2, and (iii) persistence did not appear to be due to immune escape of variants in the E1 and E2 regions. The significance of these findings to the chimpan-zee animal model and their possible extrapolation to humans is discussed here.
MATERIALS AND METHODS
Cloning and envelope proteins into baculovirus expression vectors.An E1 fragment representing nucleotides 915 to 1421 (amino acids [aa] 192 to 360) was amplified by PCR by using a previously described plasmid containing the E1 region of the HCV-1 strain (genotype 1a) (33). The E1 domains of HCV-1 and the Hutchinson strains are 98% homologous. The downstream primer for the E1
fragment spanned nucleotides 1404 to 1421 (aa 355 to 360, 59-GAAGATCTTT
AGTGGTGGTGGTGGTGGTGCGCTATGCCCGCCAGGAC-39) and
con-tained nucleotide sequences encoding a 6-histidine tail, and aBglII site. The
upstream primer (59-TTCCCCGGGATACCAAGTGCGCAACTCC-39)
con-tained aSmaI site and spanned nucleotides 915 to 932 (aa 192 to 196). The
gel-purified E1 fragment was digested withSmaI andBglII and ligated into
gel-purifiedSmaI- andBamHI-digested pPbac (Stratagene). The vector
con-tained a translational start site in frame with a human placental alkaline
phos-phatase signal sequence (APSS) upstream of theSmaI site. The APSS directs
polypeptides to the secretory pathway of insect cells. The selected clone was designated pE1–360.
* Corresponding author. Mailing address: Department of Virology and Immunology, Southwest Foundation for Biomedical Research, 7620 N.W. Loop 410, San Antonio, TX 78228. Phone: (210) 670-3245. Fax: (210) 670-3329. E-mail: [email protected].
1118
on November 9, 2019 by guest
http://jvi.asm.org/
An E2 fragment representing aa 364 to 715 from the HCV-1 strain was amplified by PCR as described above for E1 by using plasmid DNA containing the E2 region as the template (33). The E2 domains of the HCV-1 and Hutchin-son strains are approximately 94% homologous. The downstream primer for the
E2 fragment spanned nucleotides 2469 to 2486 (aa 710 to 715, 59-GGGCTGC
AGTTAGTGGTGGTGGTGGTGGTGCTTAATGGCCCAGGACGC-39) and
contained sequences encoding a 6-histidine tail and aPstI site. The upstream
primer (59-GGGGATCCATATGGTGGGGAACTGGGCGAAG-39)
con-tained aBamHI site and an ATG initiation codon and spanned nucleotides 1431
to 1451 (aa 364 to 370). The E2 fragment was gel purified, cleaved withBamHI
andPstI, and ligated into gel-purified pBacPAK9 (BamHI/PstI; Clontech). The
selected clone was designated pE2–715.
Transfections ofSf9 cells with pE1–360 and pE2–715 were performed
accord-ing to instructions provided by Clontech. Plaque assays were performed with medium derived from transfected cultures, and several plaques were
character-ized to confirm that the desired protein was expressed inSf9 cells infected with
the recombinant virus. Three plaque purifications were performed to ensure clonal recombinant virus.
Sf9 cells and baculovirus infections.Procedures involving the cultivation of insect cells and the production and use of recombinant baculoviruses were
essentially as described (44). For the purification of E1 and E2,Sf9 cells were
grown to a cell density of approximately 23106to 33106cells/ml in a 250-ml
spinner culture as previously described (32). Cells were pelleted at 1,0003gfor
10 m in and resuspended in 25 ml of virus stock for 1 h at 27°C. After infection, 225 ml of Grace’s medium supplemented with 2% fetal bovine serum and 0.1% Pluronic F-68 (JRH Biosciences) was added to the spinner of infected cells.
Purification of HCV recombinant envelope proteins.Sf9 spinner cultures infected with E1 and E2 recombinant baculoviruses were harvested at 72 h postinfection. The cells were washed three times with cold phosphate-buffered saline (PBS) and were extracted in 15 ml of EB buffer (50 mM Tris-HCl, pH 9.0; 100 mM NaCl, 1% Nonidet P-40) for 15 min, during which time the container
was vortexed lightly every 5 min. The cell lysate was clarified at 31,0003gfor 20
min.
E1 and E2 were purified over an agaroseGalanthus nivalis(snowdrop) lectin
I column (Vector Laboratories). A 1-ml column (1.5 by 15 cm, low pressure; Bio-Rad) of lectin agarose resin was equilibrated with EB buffer. The soluble cell lysate (16 ml) was passed over the resin two times at a rate of approximately 0.5 ml/min. The resin was washed with 30 ml of EB buffer followed by 20 ml of purification buffer (20 mM Tris-HCl, pH 8.0; 100 mM NaCl). The envelope
proteins were eluted from the resin with 20 ml of 1 Mmethyl-a-D
-mannopyr-anoside (Sigma) in purification buffer at a flow rate of 0.5 ml/min. Fractions (1 ml) were collected and analyzed for the presence of E1 or E2 by Western blot analysis. Fractions containing the envelope proteins were pooled and passed over a Talon metal affinity resin (Clontech).
Talon resin (3 ml) was packed into a column and washed with 4 volumes of Talon elution buffer (100 mM imidizole, 20 mM Tris-HCl, 100 mM NaCl; pH 8.0) followed by 4 volumes of purification buffer. E1 or E2 Western blot-positive fractions from lectin affinity chromatography were pooled, and purification buffer was added to the pool to obtain a volume 10 times that of the resin (30 ml). The pool was passed over the column twice at a rate of 0.5 ml/min, and the resin was washed with 72 ml (24 volumes) of purification buffer. The envelope proteins were eluted from the column with 20 ml of 100 mM imidizole in purification buffer, and fractions (1 ml) were collected at a rate of 0.5 ml/min. The fractions with high levels of E1 or E2 reactivity by Western blotting were pooled and dialyzed in 1 liter of PBS for 16 h. The dialyzed fractions were clarified by
centrifugation at 3,0003gfor 15 min. The protein concentration was estimated
by using a Micro-BCA protein assay (Pierce). Western blot analysis was also performed on start, flowthrough, and pooled column fractions to determine the efficiency of the purification.
SDS-PAGE and Western blot analysis.Discontinuous sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed as previously described (28, 29). The gels were either silver stained according to the procedure of Wray et al. (51) or were further processed for Western blot analysis as previously described (30). For Western blot analysis, the gels were transferred to a Pall Biosupport Flourotrans polyvinylidene difluoride membrane (ICN Bio-medical). The primary antibodies were rabbit antisera produced against synthetic peptides derived from capsid/E1 (aa 185 to 210) and E2 (aa 439 to 466) (33).
E1 and E2 ELISAs.Microtiter plates (flat-bottom, high-binding ELISA plates; Corning) were coated with antigen for 16 h at 4°C in borate-buffered saline (48.5 mM boric acid, 145 mM NaCl, 6.0 mM NaOH, 50 mM KCl [pH 8.2]). The
volume of reagent added to the microtiter wells was 100ml for all steps except for
the blocking (200ml) and washing (300ml) steps. A titration assay was performed
to determine the optimal coat concentrations (;5 ng/well for E1;;100 ng/well
for E2). Wells were blocked with PBS containing 5% nonfat dry milk (NFDM) and 0.05% Tween 20 (NFDM-PBS-Tween) at 37°C for 2 h and were washed three times with PBS-Tween. The primary antibody was diluted 1:200 in NFDM-PBS-Tween and was incubated with the wells for 2 h at 37°C. The wells were washed three times with PBS-Tween. Binding of primary antibodies was detected with horseradish peroxidase-conjugated goat anti-human immunoglobulin G sec-ondary antibody (Southern Biotechnology Associates, Inc.). The conjugate was diluted in NFDM-PBS-Tween (1:1,000). Wells were incubated with the conju-gate at 37°C for 1 h, washed three times in PBS-Tween, and incubated with the
peroxidase substrate solution (0.54 mM 2,29
-azino-bis[3-ethylbenzthiazoline-6-sulfonic acid], 0.03% H2O2, 0.1 M citric acid; pH 4.0) at room temperature for
30 min. The color reaction was stopped with 10% SDS. The absorbance at 410 nm (reference wavelength, 490 nm) was read by using a Dynatech MR700 microtiter plate unit. The raw data were analyzed with a computer program in Microsoft Excel designed specifically for these assays by Mark Sharp (unpub-lished data). A human or chimpanzee pool of normal, anti-HCV negative sera was run in triplicate on every microtiter plate. The average optical density (OD) reading from the negative control wells was multiplied by 2.5 to establish a cutoff value. A negative value indicated that the average OD reading from a serum sample run in duplicate was below the cutoff value.
A peptide-based ELISA was developed by using a synthetic peptide repre-senting the E2 hypervariable region (HVR-1; aa 384 to 413) of the Hutchinson strain (genotype 1a). Wells were coated with 200 ng of peptide, and the ELISA was performed as described above. Positive controls included serum samples from an HCV-infected chimpanzee or an HCV 2.0-positive human.
Analysis of sera for anti-HCV antibodies by ELISA and recombination im-munoblot assay (RIBA).Chimpanzee and human sera were analyzed for anti-HCV antibody by using a second-generation ELISA (anti-HCV 2.0 ELISA; Ortho Diagnostic Systems, Raritan, N.J.). Chimpanzee sera were also analyzed for HCV antibodies to individual HCV proteins by using HCV BLOT 3.0 (Genelabs Diagnostics). These assays were performed according to the manufacturer’s instructions.
RT-PCR amplification and sequence analysis of E2 HVR-1, E1, and E2. RNA extraction, cDNA synthesis and first-round PCR amplification were the same as described previously, except for the primers used in the RT-PCR reaction (7).
For first-round amplification of the E2 HVR-1, the downstream primer (59-GA
GTGAAGCAATATACCGGAC-39) spanned nucleotides 1852 to 1872 (aa 503
to 510), and the upstream primer (59-ATGGTGGGGAACTGGGCGAAGGT
C-39) spanned nucleotides 1431 to 1454 (aa 363 to 371). In the second-round
amplification of the E2 HVR-1, the downstream primer (59-CACAAGCTTTA
GTGCCAGCAGTAGGGGCGC-39) spanned nucleotides 1787 to 1807 (aa 482
to 488), and the upstream primer (59-CACGAATTCCATGGTGCTGCTGCT
ATTTGCCGGCGTCGACG-39) spanned nucleotides 1461 to 1488 (aa 373 to
382). Additionally, overlapping regions spanning aa 192 to 791, which included the E1 and E2 genes, were amplified from serum representing the acute and chronic phases of disease. PCR fragments were cloned and sequenced or, in some cases, the PCR products were directly sequenced. Automated DNA se-quence analysis was performed at the University of Texas Health Science Center at San Antonio at the Center for Advanced DNA Technologies.
RESULTS
Purification of E1 and E2 and ELISA development.In order
to evaluate the immune response of chimpanzees to E1 and E2, the proteins were expressed in insect cells by using recom-binant baculovirus constructs. We have previously demon-strated that the humoral immune response to E1 and E2 was primarily to denaturation-sensitive epitopes (33). Thus, the carboxy-terminal hydrophobic domains of E1 and E2 were deleted to express these proteins in the soluble fraction of the cell to facilitate purification under nondenaturing conditions. The E1 (aa 192 to 360) and E2 (aa 364 to 715) constructs were engineered with truncations at the 39end, followed by nucle-otides encoding a 6-histidine tail. Signal sequences at the amino terminus of E1 and E2 were included to direct the protein into the secretory pathway. In addition, the E1 gene was cloned into a vector with a human placental alkaline phos-phatase signal sequence at the amino terminus in an attempt to increase secretion into the culture medium. Approximately 60% of the E1 and E2 was present in the soluble fractions of the cells, and the proteins were glycosylated (33). Although approximately 20% of E2 was secreted into the medium, E1 was not secreted. E1 and E2 proteins were purified fromSf9 cell lysates under native conditions by using an agarose G. nivalislectin I and a Talon metal affinity resin as described in Materials and Methods. Approximately 120mg of E1 and 2.5 mg of E2 were purified per 2 3 109 cells (1-liter spinner culture) based on BCA analysis. Western blot analysis revealed that the purification was not highly efficient, since a large fraction of E1 and E2 was present in the unbound fractions of both resins (data not shown). However, this purification scheme yielded proteins with very low reactivity to normal human sera as determined by ELISA. The purity of E1 and E2
on November 9, 2019 by guest
http://jvi.asm.org/
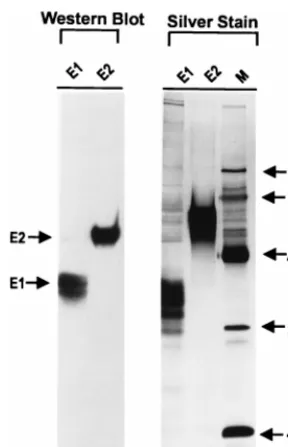
was assessed by silver staining (Fig. 1) and was estimated to be approximately 80 to 95%.
Titration assays were performed to determine the optimal coat concentrations for E1 and E2 and the optimal dilution of primary antibody. An ELISA was performed with a human serum that reacted with both E1 and E2, and a coat con-centration that yielded an optimal signal-to-noise ratio was chosen. Titration assays with several human sera were also performed to choose an antibody dilution with an optimal signal-to-background ratio. Additionally, serum samples from two individuals yielding high and medium OD readings were included in every ELISA plate as positive controls to examine the consistency of the ELISA. Serum samples collected from 36 normal human blood donors and from 36 chimpanzees that were not exposed to HCV were examined for E1 and E2 reactivity by ELISA to ensure that false-positive reactions were insignificant. No false-positive reactivity was observed in these panels, so normal human and chimpanzee sera were indepen-dently pooled and used as negative controls.
Humoral immune response to envelope proteins in
HCV-infected human cohorts.Prior to analysis of the chimpanzee
cohort, human serum panels were examined to assess the per-formance of the assay and the frequency of E1 and E2 anti-bodies (anti-E1 and anti-E2) according to a widely used, com-mercially available test. Ortho-HCV 2.0-positive human serum samples from three different human cohorts were examined for anti-E1 and anti-E2 by ELISA (Table 1). Panel 1 was com-prised of sera from intravenous drug users (IVDUs) from a cohort that has been previously described and in which more than 95% of the Ortho-HCV 2.0 positive sera are also positive as determined by RIBA and/or for HCV RNA (45). Anti-E1 and anti-E2 reactivity were observed in 58% (149 of 257) and 93% (239 of 257) of the serum samples, respectively. There-fore, these ELISAs appeared to be sensitive indicators of HCV infection. Panel 2 consisted of plasma rejected from the blood
bank based on elevated alanine aminotransferase (ALT) val-ues prior to 1992, when blood donations were not screened for anti-HCV. Only HCV 2.0-positive samples were included in this panel. Anti-E1 and anti-E2 were observed in 44 and 62% of the samples, respectively. The higher percentage of antibody reactivity in the IVDU panel compared to the blood donor panel may be due to a longer duration of infection, as well as to potential continuous inoculation with HCV in the IVDU cohort. Panel 3 represented individuals with recently acquired symptomatic non-A, non-B hepatitis (NANBH), indicating an acute infection, which was retrospectively screened to include only HCV 2.0-positive sera. Anti-E1 and anti-E2 reactivity were observed in 22% (4 of 18) and 33% (6 of 18) of the individuals, respectively. The low frequency of reactivity to E1 and E2 in this panel may be explained by the short duration of infection in the acute panel. Alternatively, the low frequency of reactivity may be an artifact due to the small sampling size.
Humoral immune response to envelope proteins in a cohort
of HCV-inoculated chimpanzees. Serum samples from 46
HCV-inoculated chimpanzees were analyzed for the presence of anti-HCV antibody by HCV 2.0 ELISA and for the presence of HCV RNA by RT-PCR as previously described (7). The chimpanzees were exposed to HCV inocula during the past 2 to 19 years, with an average duration of 10.6 years since inoc-ulation. Of the 46 chimpanzees, at least 14 had received the Hutchinson strain inoculum, at least 3 had received the HCV-1 inoculum, and 28 had received unknown inocula. Antibody to HCV proteins was detected in 48% (22 of 46) of the chimpan-zees by the HCV 2.0 ELISA (Table 2). The majority of HCV-inoculated chimpanzees did not have detectable antibody as
FIG. 1. SDS-PAGE analysis of purified E1 and E2 proteins. Western blot
analysis of E1 (100 ng) and E2 (5.2mg) was performed as described in Materials
and Methods. Purified E1 (500 ng) and E2 (1.3mg) were analyzed by SDS-PAGE
and silver staining. Recombinant E1 and E2 proteins were purified fromSf9 cell
lysates by using an agaroseG. nivalis(snowdrop) lectin 1 and a Talon metal
[image:3.612.103.247.67.291.2]affinity resin as described in Materials and Methods.
TABLE 1. E1 and E2 antibody status of anti-HCV-positive human and chimpanzee serum panels
Panel
no. Panela No. tested
% Positive (no. positive/total
no.)bfor:
Anti-E1 Anti-E2
1 IVDU 257 58 (149/257) 93 (239/257)
2 ALT 34 44 (15/34) 62 (21/34)
3 NANBH (acute) 18 22 (4/18) 33 (6/18) 4 Chimpanzeec 22 18 (4/22) 32 (7/22)
aAll of the serum samples in the panels were found to be anti-HCV positive
by HCV 2.0
bSerum samples were analyzed for antibodies to E1 and E2 as described in
Materials and Methods.
cThe E2-positive animals included a single chimpanzee that was (i) considered
[image:3.612.312.551.602.691.2]“borderline” by the standard E2 ELISA in 1995 but found to be positive in 1997 and (ii) positive in 1995 for an E2 ELISA when five times more antigen was used to coat the wells.
TABLE 2. Immune response to E1 and E2 in HCV-inoculated chimpanzees
Status No. of chimpanzees (%) No. positive (%)
afor:
Anti-E1 Anti-E2
Anti-HCV1 22 (48) 4 (18) 7 (32)
Anti-HCV2 24 (52) 6 (25) 0 (0)
RNA1b 18 (39) 3 (17) 7 (39)
RNA2 28 (61) 7 (25) 0 (0)
Total 46 10 (22) 7 (15)
aSerum samples were analyzed by HCV 2.0, E1, and E2 ELISAs as described
in Materials and Methods
bSerum samples were analyzed for HCV RNA by RT-PCR analysis as
de-scribed in Materials and Methods.
on November 9, 2019 by guest
http://jvi.asm.org/
determined by ELISA against the envelope proteins. Anti-E1 and anti-E2 reactivity were observed in 22% (10 of 46) and 15% (7 of 46) of the HCV-inoculated chimpanzees, respec-tively. Of the HCV 2.0-positive animals, only 18% (4 of 22) and 32% (7 of 22) were anti-E1 and anti-E2 positive, respectively (Tables 1 and 2). None of the HCV 2.0-negative animals had antibody to E2, although 25% were anti-E1 positive (Table 2). Therefore, only HCV 2.0-positive animals had antibody to E2, while the anti-E1 response was similar for both HCV 2.0-positive and -negative animals.
Humoral immune responses to the envelope proteins in
viremic chimpanzees.Although HCV infections after
inocula-tion had been confirmed in all 46 of the animals in this study (7), HCV RNA was detected in recently collected serum sam-ples from only 18 (39%) of the chimpanzees (Table 2). Of the viremic chimpanzees, anti-E1 antibody was detected in 17% (3 of 18) and anti-E2 antibody was detected in 39% (7 of 18) (Table 2). All of the anti-E2 positive chimpanzees were viremic (7 of 7) (Table 2). In contrast, anti-E1 was detected in both viremic (17%) and nonviremic animals (25%). Antibody to E1 and E2 were simultaneously detected (E11E21) in 17% of the
viremic chimpanzees (Table 3). Antibody reactivity to E1 but not to E2 (E11E22) was not observed in the viremic animals,
and an antibody response elicited against E2 in the absence of anti-E1 antibody (E12E21was observed in 22% of the viremic
animals (Table 3). The majority of viremic chimpanzees lacked a detectable humoral immune response to both E1 and E2 (61% E12E22) (Table 3).
Humoral immune responses to the envelope proteins in
nonviremic chimpanzees. Viral clearance had apparently
oc-curred in 28 chimpanzees (61%), since HCV RNA was not detected in the serum samples collected in 1995 (Table 2). Convalescence was supported by the observation that most RT-PCR-negative chimpanzees were also anti-HCV-negative by ELISA (Ab2PCR2; 86%, 24 of 28) and that nested PCR
followed by Southern hybridization failed to reveal HCV RNA in serum from antibody-positive but RT-PCR negative (Ab1
PCR2) chimpanzees. Anti-E1 was detected in 25% (7 of 28) of
the HCV RNA-negative chimpanzees, while none of these animals had a detectable anti-E2 response (Table 2). The fre-quency of anti-E1 in HCV 2.0- and HCV RNA-negative chim-panzees (Ab2PCR2) was also 25% (6 of 24). Of the
conva-lescent chimpanzees, 25% were E11E22, none were E11E21
or E12E21, and the majority were E12E22(75%) (Table 3).
In summary, convalescent animals lacked anti-E2 but had a percentage of anti-E1 reactivity similar to the viremic animals. The E1 response in viremic and nonviremic animals was not significantly different, while differences in the E2 response were significant (P50.009 by chi-square analysis). Although anti-E1 did not appear to correlate with chronicity or conva-lescence, antibody to E1 in the absence of antibody to E2 may be a potential marker for convalescence in some individuals (Table 3).
Longitudinal evaluation of humoral immune responses to
the envelope proteins.An anti-E2 antibody response appeared
[image:4.612.53.295.80.152.2]to correlate with persistent HCV infection rather than conva-lescence in the cross-sectional study (Table 2). To determine whether anti-E2 was elicited prior to convalescence but de-clined after clearance of viremia, a longitudinal study of the humoral immune response in six HCV-infected chimpanzees was performed. The Hutchinson strain of HCV was serially passaged into six chimpanzees as described previously. Viral clearance was observed in four animals within 5 months posti-noculation, with either gradual or rapid loss of anti-HCV an-tibody based on the HCV 2.0 ELISA. A chronic carrier profile characterized by persistent HCV RNA was observed in two animals. One of these chimpanzees was RT-PCR positive and antibody negative for 5 years. This animal would not have been detected with the standard assay employed by blood banks and thus represents a “silent carrier”. Serial serum samples from these six chimpanzees were examined for anti-E1 and anti-E2 by ELISA. Anti-E1 was not detected in the serial serum sam-ples from any of the chimpanzees. An anti-E2 response was not observed in the serial bleeds of the convalescent animals and
[image:4.612.148.457.544.704.2]FIG. 2. Antibody response to E1, E2, and the E2 HVR-1. Serial bleeds obtained up to 9 years postinoculation with HCV from chimpanzee x174 were examined by ELISA for reactivity to native, purified E1 and E2, and a synthetic peptide to the HVR-1 as described in Materials and Methods.
TABLE 3. Correlation of antibodies to E1 and E2
E1 and E2 statusa No. of
chimpanzees
No. of animals (%)b
RNA1(n518) RNA2(n528)
E11E21 3 3 (17) 0 (0)
E11E22 7 0 (0) 7 (25)
E12E21 4 4 (22) 0 (0)
E12E22 32 11 (61) 21 (75)
aSerum samples were analyzed for E1 and E2 antibody by ELISA as described
in Materials and Methods.
bSerum samples were analyzed for HCV RNA by RT-PCR analysis as
de-scribed in Materials and Methods.
on November 9, 2019 by guest
http://jvi.asm.org/
was detected in only one viremic animal, x174 (Fig. 2). Al-though this animal was HCV 2.0 positive by 3 months postin-oculation, the anti-E2 response was not detected until 7 years postinoculation, though it remained positive thereafter. Anti-bodies to the HVR-1 and to E1 were not detected in serum samples from x174. Although the number of animals examined was small, the results of the longitudinal study agreed with those of the cross-sectional study in that convalescent animals did not have detectable levels of antibody to E2 and suggested that the low reactivity to E1 and E2 observed in the cross-sectional study was not due to the sampling of a single time point.
Antibody responses to E2 and sequence changes in the E2
HVR-1.Several studies have suggested that viral persistence
may be associated with the emergence of viruses that vary in the HVR-1 that are able to escape neutralizing antibody. To examine the role of an antibody response to the HVR-1 in chimpanzees, serial serum samples from the six chimpanzees inoculated with the Hutchinson strain of HCV were examined for anti-HVR-1 antibody. An ELISA was developed by using a synthetic peptide homologous to the HVR-1 of the Hutchinson strain of HCV. An anti-HVR-1 antibody response was not observed in these chimpanzees, irrespective of whether they convalesced or progressed to chronic infection. Although none of the chimpanzees were positive in the HVR-1 ELISA, all were positive for anti-HCV at some time point during the analysis and four demonstrated antibodies in a different pep-tide-based ELISA to the NS4A region (data not shown). The lack of anti-HVR-1 antibody was confirmed by an immunopre-cipitation assay with a GST–HVR-1 fusion protein (data not shown).
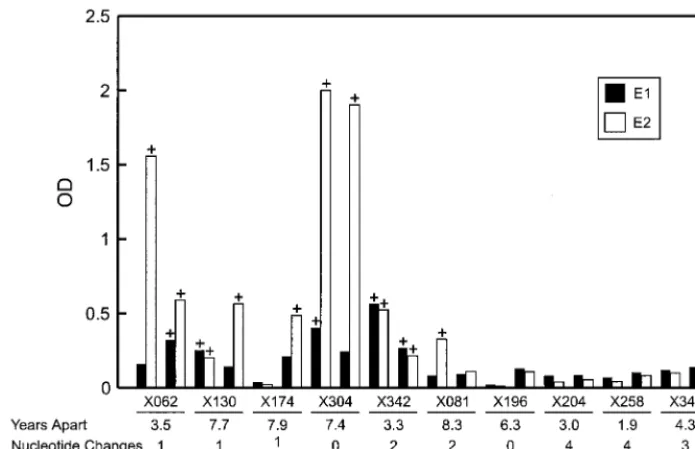
The relationship between an antibody response to E2 or to the HVR-1 and nucleotide changes in the HVR-1 was exam-ined in 10 chronically infected animals. The animals were se-lected to contain five anti-E2-positive and five anti-E2-negative chimpanzees; four were infected with the Hutchinson strain of HCV (x62, x81, x174, and x196), and six were infected with undesignated strains of HCV (Table 4 and Fig. 3). A detect-able anti-HVR-1 antibody response was observed in only one animal (x62). Reactivity to the HVR-1 peptide was confirmed in this animal by analysis of separate serum samples collected 3 years apart.
[image:5.612.51.552.81.233.2]To evaluate the relationship of anti-E2 and sequence changes in the HVR-1, a sequence spanning the HVR-1 was amplified from temporally spaced chimpanzee sera represent-ing, as closely as possible, the acute infection and a recently
FIG. 3. Alignment of amino acid sequences of the E2 HVR-1. The amino acid sequences of the HVR-1s from the acute and chronic phases from 10 chimpanzees are aligned with the amino acid sequence of the HVR from the Hutchinson strain of HCV. Residues identical to the Hutchinson sequence are indicated by a period. Residues differing between the acute and chronic samples are shown in boldface. The clinical records indicate that animals x81, x174, x196, and x62 received the Hutchinson inoculum; animals x204 and x258 received a common inoculum; animals x341 and x342 received a common inoculum, and it is suspected that animal x304 received this inoculum as well. The inoculum for animal x130 was not indicated.
TABLE 4. Divergence within the E2 HVR
Chimpanzee Yra No. of changes
b Antibodyc
Rate of variation (1023)d
nt aa E2 HVR-1
x62 3.5 1 1 1 1 3.17
x130 7.7 1 1 1 2 1.44
x174 7.9 1 1 1 2 1.41
x304 7.4 0 0 1 2 0
x342 3.3 2 1 1 2 6.74
x81 8.3 2 0 2 2 2.68
x196 6.3 0 0 2 2 0
x204 3 4 2 2 2 14.8
x258 1.9 4 3 2 2 23.39
x341 4.3 3 1 2 2 7.75
Total 53.6 18 10 5 1 NA
Average 0.34e 0.19e NA NA 3.73
aThe duration of time between the collection of temporally spaced serum samples.
bNucleotide (nt) and amino acid (aa) changes in the HVR-1 sequences from chimpanzee sera representing as closely as possible the acute and chronic phases of
infection were analyzed as described in Materials and Methods.
cSerum samples were analyzed for the presence (1) or absence (2) E2 and HVR-1 antibody as described in Materials and Methods. NA, not applicable. dThe rate of variation was calculated as the number of nucleotide changes/site/years between temporally spaced bleeds.
eAverage changes per year.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.327.521.382.641.2]acquired serum sample. Sera collected during the acute phase of infection were not available from x62, x304, x130, and x81; therefore, the sera analyzed were collected at the earliest time points available after infection. The duration of time between the temporally spaced chimpanzee sera was from 1.9 to 8.3 years (average55.4 years). Since the primary objective was to detect changes in the predominant sequence, rather than to analyze the total quasispecies, the initial analysis involved the sequencing of a single clone from each time point. This ap-proach should detect the emergence of variants due to a strong selective pressure. The validity of the results was confirmed by analysis of additional clones and direct sequencing of the PCR products.
Alignment of the deduced amino acid sequences with the HVR-1 sequence of the Hutchinson HCV strain emphasized several points. The sequences from animals inoculated with serial passages of Hutchinson were identical or nearly identical to the original sequence, whereas animals receiving other in-ocula were divergent, emphasizing the variability of this region. In contrast, the sequences from the acute and chronic bleeds for an individual animal were nearly identical, emphasizing the lack of divergence in this sequence in these chimpanzees. An average of 0.34 nucleotide changes/year was observed, with a range of 0 to 4 nucleotide changes per animal. Additionally, an average of 0.19 amino acid changes/year was observed, with a range of 0 to 3 amino acid changes per animal. The number of sequence variations in the HVR-1 did not coincide with the antibody response elicited against E2 or the HVR-1 (x62) or the duration of time between samples (Table 4, Fig. 3 and 4). Anti-E2 was detected in both serum samples from x304 spaced 7.4 years apart, yet no nucleotide or amino acid changes in the HVR-1 were observed. Conversely, the greatest number of nucleotide and amino acid changes was observed in an animal (x258) that lacked detectable antibodies to E2 and the HVR-1. However, since the peptide used in the ELISA was not com-pletely homologous to the HVR-1 sequenced from x258 (21 of
30 positions were homologous), anti-HVR-1 antibody may have not been detected by this ELISA. The average rate of variation, or nucleotide changes per site per year, was 3.733 1023, with a range of 0 to 23.331023nucleotide changes/site/ year. However, it should be noted that 44% of the nucleotide changes were silent and did not result in amino acid changes. The lack of a bias for nonsynonymous nucleotide changes argues against a strong immunoselection in this domain in these chimpanzees. The four animals inoculated with the Hutchinson strain exhibited only 2 aa changes compared to the inoculum over a cumulative of 22.5 years.
Since very few nucleotide and amino acid changes were observed in all 10 animals, multiple clones of the HVR-1 from each animal were not sequenced. However, we determined the dominant variant by direct sequencing of the RT-PCR frag-ments representing the HVR-1 from the most recent serum sample from five chimpanzees. Although 0 to 1 aa differences were observed, the deduced amino acid sequences of the PCR fragments closely matched the sequences of the paired clones. This suggested that the dominant sequences had been cloned and that major changes in the HVR-1 had not occurred. How-ever, limited quasispecies were present, since minor variations in the HVR-1 were observed. The sequence of the PCR frag-ment and one clone derived from a serum sample collected in 1996 from x174 were identical to the sequence of the clone representing the acute infection. However, two additional clones from 1996 and a clone from 1995 had one to four nucleotide changes and 1 to 2 aa changes.
Evaluation of E1 and E2 sequences for variability outside of
E2 HVR-1.Immune escape of variants in the HVR-1 has been
[image:6.612.129.477.70.294.2]suggested to contribute to persistent HCV infection in hu-mans. However, persistent infection was observed in three chimpanzees even in the absence of amino acid changes in the HVR-1, and changes were minimal in the other seven animals (Table 4). To determine whether variable regions exist in chim-panzees, other than the HVR-1 observed in humans, the E1
FIG. 4. Relationship of immune responses to envelope proteins and nucleotide changes in the HVR-1. Temporally spaced bleeds from 10 HCV-infected chimpanzees were examined, 5 of which had an antibody response to E2. The HVR-1 sequence was amplified from each serum sample and was cloned and sequenced as described in Materials and Methods. The sera were examined by ELISA for antibody to E1 and E2. Positive responses are indicated by a “1” at the top of the bars. The bars represent the OD values from the E1 and E2 ELISAs of paired sera, with the earliest and latest bleeds represented by the bars to the left and right, respectively. The chimpanzee numbers, the duration of time between temporally spaced sera, and the number of nucleotide and amino acid changes are noted at the bottom.
on November 9, 2019 by guest
http://jvi.asm.org/
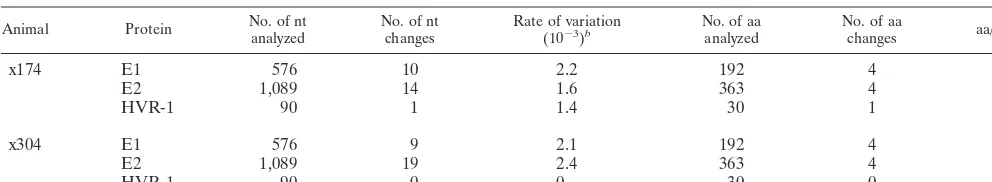
and E2 genes were sequenced from paired sera collected from x174 and x304 (Table 5). Serum samples were chosen to rep-resent the greatest possible duration of time between samples, which was 7.9 years for x174 and 7.4 years for x304. Anti-E2 was detected in the most recent serum sample from x174 and from both samples collected from x304. Although anti-E2 only recently emerged in x174 (7 years postinoculation), it was present for approximately 1 year prior to the collection of the serum sample used for the sequencing of E1 and E2. Although both animals had anti-E2, no major sequence changes in the E1 and E2 regions were observed. The rates of variation in the HVR-1, E1, and E2 regions sequenced from x174 were 1.43 1023, 2.2 3 1023, and 1.6 3 1023, respectively (Table 5). Similarly, the rates of variation in the HVR-1, E1, and E2 regions sequenced from x304 were 0, 2.13 1023, and 2.43 1023, respectively. Similar rates of variation were observed in the E1 and E2 regions, despite the presence of antibody ex-clusively to E2. Although the position of amino acid changes varied between x174 and x304, in both animals, 4 aa changes were observed within E1 and 4 aa changes were observed within E2. Interestingly, the percentage of nucleotide changes that resulted in amino acid changes was higher in E1 (40 to 44%) than in E2 (21 to 29%) (Table 5). Since the chimpanzees had detectable anti-E2 but not anti-E1, immunoselection did not appear to be driving the nonsilent nucleotide changes observed in E1. However, the E1 ELISA may not have been adequate for the detection of neutralizing antibody that may have driven immunoselection. Perhaps neutralizing antibody only recognizes E1 in the form of E1-E2 heterodimers.
DISCUSSION
We have examined HCV-inoculated chimpanzees for anti-body reactivity to the envelope proteins and sequence variabil-ity in the envelope domain. The results revealed that (i) a low percentage of HCV-inoculated chimpanzees had antibody to E1 and E2, (ii) convalescence did not appear to be associated with an antibody response to E1 or E2, and (iii) viral persis-tence did not appear to be due to the antibody escape of variants in the E1 and E2 regions.
Prior to examining serum samples from HCV-inoculated chimpanzees for antibody to the envelope proteins, the validity of the E1 and E2 ELISAs was assessed by testing three panels of HCV 2.0-positive human sera (Table 1). The three panels were representative of individuals with acute NANBH, blood donors with elevated ALT levels, and IVDUs. The low per-centages of anti-E1 (22%) and anti-E2 (33%) reactivity ob-served in the acute serum panel may be due to insufficient time to mount an immune response to the envelope proteins. Nishi-hara and co-workers observed a similar percentage of anti-E2
(29%) in an acute panel (38). Panel 2 consisted of serum samples from HCV 2.0-positive blood donors with elevated ALT levels. Antibody to E1 and E2 was observed in 44 and 62% of the serum samples, respectively. Although the presence of HCV RNA was not assessed in our HCV 2.0-positive panel of blood donors, a higher percentage of anti-E2 positive sam-ples may have been observed in this panel if only viremic individuals were tested. We observed that 58 and 93% of HCV 2.0-positive sera from the IVDU panel were anti-E1 and an-ti-E2 positive, respectively (Table 1). The higher percentage of antibody reactivity in the IVDU panel compared to the other panels may be due to a greater proportion of long-term infec-tions. Alternatively, a greater breadth of immunoreactivity may be noted in IVDUs with ongoing and repeated exposure to HCV.
In our human serum panels, lower reactivity to E1 (22 to 58%) than to E2 (33 to 93%) was observed. A study of trun-cated HCV glycoproteins revealed that E2 appeared to be required for the correct folding of E1 (35). Therefore, the lower antibody reactivity to E1 compared to E2 may be ex-plained if conformational differences due to the misfolding of E1 hinder antibody recognition. The percentage of responders to E2 in the IVDU panel in our ELISA was similar to the percentages of E2 responders in chronically infected individu-als reported in several earlier studies (range5 53 to 100%) (11, 13, 21, 24, 34, 38, 47, 54). Therefore, our assay appeared to be a sensitive indicator of HCV infection.
[image:7.612.55.551.81.172.2]A low percentage of the 46 HCV-inoculated chimpanzees responded to E1 (22%) and E2 (15%) but, in contrast to the human panels, E1 reactivity was higher than E2 reactivity. The higher prevalence of anti-E1 versus anti-E2 in our chimpanzee cohort may be explained by the high percentage of convales-cent animals in the cohort. The lack of detectable anti-E1 and anti-E2 in the majority of HCV-inoculated chimpanzees did not appear to be due to a deficient humoral immune response to HCV, since even within the chimpanzees that were positive by the HCV 2.0 ELISA, only 18 and 32% of these animals were anti-E1 and anti-E2 antibody positive, respectively (Table 2). Although it is possible that the negative serological responses in some chimpanzees may have been due to differences be-tween strains or genotypes used for inoculation and the anti-gens in the serologic assays, antibody to E2 was detected in 93% of a human serum panel. This suggests that the E2 ELISA was probably sensitive to a wide variety of strains, since hu-mans are exposed to highly heterogeneous inocula. Previous studies have suggested that humoral immune responses to HCV structural proteins are observed less frequently in chim-panzees than in humans for reasons not understood (2, 14, 22, 31, 36, 48, 49).
TABLE 5. Divergence within E1 and E2a
Animal Protein No. of ntanalyzed No. of ntchanges Rate of variation(1023)b No. of aaanalyzed No. of aachanges aa/nt (%)c
x174 E1 576 10 2.2 192 4 40
E2 1,089 14 1.6 363 4 29
HVR-1 90 1 1.4 30 1 100
x304 E1 576 9 2.1 192 4 44
E2 1,089 19 2.4 363 4 21
HVR-1 90 0 0 30 0 0
aThe E1 and E2 regions analyzed spanned aa 192 to 383 (576 nucleotides [nt]) and an 384 to 746 (1,089 nucleotides), respectively. The HVR-1 region spanned aa
384 to 413 (90 nucleotides). The duration of time between the collection of serum samples was 7.9 years for x174 and 7.4 years for x304.
bThe rate of variation was calculated as the number of nucleotide changes/site/years between temporally spaced bleeds.
cThe percentage of nucleotide changes resulting in amino acid changes.
on November 9, 2019 by guest
http://jvi.asm.org/
Viral clearance was not associated with an antibody re-sponse to E1 or E2 in the cross-sectional study. Chimpanzees that cleared HCV lacked an E2 antibody response but had approximately the same reactivity to E1 as the persistently infected animals (Table 2). E2 antibody is consistently detected in chronically infected humans (11, 13, 21, 24, 34, 38, 47, 54), and several studies suggests that E2 antibody is a marker for active HCV replication (11, 20, 54). Consistent with data from human studies, E2 antibody was detected exclusively in viremic chimpanzees (Table 3). In the cross-sectional study, it could not be determined if nonviremic chimpanzees elicited an E2 antibody response that declined after viral clearance or if an-tibody to E2 was never produced.
In the longitudinal study, antibodies to E1, E2, and the HVR-1 were not detected in serial serum samples from five chimpanzees (four nonviremic, one viremic). Antibody to E2, but not to E1 or to the HVR-1, was detected in one persistently infected animal (x174), but only after 7 years postinoculation (Fig. 2). If the data from the longitudinal study can be extrap-olated to the 46 chimpanzees, it is possible that antibodies to E1, E2, and the HVR-1 were never elicited in the majority of chimpanzees and that the antibodies did not simply decline after viral clearance. Several studies have suggested that neu-tralizing antibodies target the envelope proteins (14, 19, 43) and that an early antibody response to the HVR-1 is associated with convalescence (4, 56). However, additional immune fac-tors (e.g., cytotoxic T lymphocytes and cytokines) may be in-volved in viral clearance, since antibody to E2 and to the HVR-1 was not detected in serial serum samples from the four chimpanzees that cleared the virus in the longitudinal study.
Several studies have suggested that viral persistence may be associated with the continuous emergence of viruses variant in the HVR-1 that are able to escape neutralizing antibody. How-ever, the prevalence of immune escape variants is often diffi-cult to assess, since minor variation in quasispecies due to random genetic drift may be interpreted as immunoselection. Parameters such as viral load and the presence of complex quasispecies in the inoculum may affect the rate of variation. Increased viral replication, as reflected in higher viral loads, should lead to higher rates of variation due to increased errors by RNA polymerase. A minor variant in the quasispecies may emerge as a predominant strain if it replicates more efficiently or due to T-cell immune escape variants at any position in the polyprotein. Since these changes will be in linkage with varia-tions in the HVR-1, it would appear as if a new HVR-1 was selected by immune escape mechanisms.
Mechanisms in addition to the emergence of escape variants in the HVR-1 must play a role in maintaining persistent infec-tion. Strong selective pressure did not appear to be driving the emergence of variants in our study. We observed very few amino acid changes in the HVR-1 during the time between collection of temporally spaced bleeds in 10 chimpanzees, and the percentages of silent and nonsilent nucleotide changes were similar (Table 4, Fig. 3). Additionally, antibody to E2 or the HVR-1 did not coincide with the number of nucleotide and amino acid variations in the HVR-1 (Table 4, Fig. 3 and 4). In some studies of persistently infected humans and chimpanzees, lower variation in the HVR-1 has also been observed and is sometimes associated with the absence of an antibody response to the HVR-1, near-normal ALT levels, and an improved re-sponse to interferon treatment (1, 8, 17, 26, 27, 39, 41, 46, 52, 53, 55). Since the majority of chimpanzees also have near-normal ALT levels and lack antibodies to E2, the lower vari-ation in the HVR-1 was not surprising (7, 31). Possibly, the chimpanzees may represent the healthier, more asymptomatic population of HCV-infected humans with normal ALT levels.
The entire E1 and E2 genes were sequenced from tempo-rally spaced sera collected from two animals to determine whether additional variable regions within the envelope re-gions, other than the E2 HVR-1 region, could contribute to persistence in chimpanzees. Major sequence changes in the E1 and E2 genes were not observed, although both animals had antibody to E2 in the chronic phase. The rates of variation in the E1 region from x174 and x304 were similar to the rates observed by Okamoto and coworkers and by Ogata and co-workers (9, 40, 42). In contrast to these studies, lower rates of variation were observed here in the E2 regions. The emergence of variants due to a humoral immune response was not clearly apparent. Although antibody to E2 was detectable for 7.4 years in x304 and for only 1 year in x174, the rates of variation and the percentage of nucleotide changes that resulted in amino acid changes were similar.
The chimpanzee animal model will most likely be used in assessing experimental therapies and vaccines. Recombinant envelope proteins have already been used in one vaccine study (14) and may be used in future studies. Therefore, a better understanding of the humoral and cellular immune response to the envelope proteins and the genetic drift of HCV in this animal model may aid in the interpretation of such important studies.
ACKNOWLEDGMENTS
This work was supported by grant AI40035 from the National Insti-tutes of Health.
We thank Mark Sharp for many helpful discussions of the data and for help with the statistical analysis.
REFERENCES
1.Abe, K., G. Inchauspe, and K. Fujisawa.1992. Genomic characterization and mutation rate of hepatitis C virus isolated from a patient who contracted hepatitis during an epidemic of non-A, non-B hepatitis in Japan. J. Gen.
Virol.73:2725–2729.
2.Abe, K., G. Inchauspe, T. Shikata, and A. M. Prince.1992. Three different
patterns of hepatitis C virus infection in chimpanzees. Hepatology15:690–
695.
3.Abrignani, S.1997. Immune responses throughout hepatitis C virus (HCV) infection: HCV from the immune system point of view. Springer Semin.
Immunopathol.19:47–55.
4.Allander, T., A. Beyene, S. H. Jacobson, L. Grillner, and M. A. A. Persson.
1997. Patients infected with the same hepatitis C virus strain display different
kinetics of the isolate-specific antibody response. J. Infect. Dis.175:26–31.
5.Alter, H. J.1995. To C or not to C: these are the questions. J. Am. Soc.
Hematol.85:1681–1695.
6.Alter, M. J., H. S. Margolis, K. Krawczynski, F. N. Judson, A. Mares, W. J. Alexander, P. Y. Hu, J. K. Miller, M. A. Gerber, R. E. Sampliner, E. L. Meeks, and M. J. Beach.1992. The natural history of community-acquired
hepatitis C in the United States. N. Engl. J. Med.327:1899–1905.
7.Bassett, S. E., K. M. Brasky, and R. E. Lanford.1998. Analysis of hepatitis C virus (HCV)-inoculated chimpanzees reveals unexpected clinical profiles.
J. Virol.72:2589–2599.
8.Booth, J. C., U. Kumar, D. Webster, J. Monjardino, and H. C. Thomas.1998. Comparison of the rate of sequence variation in the hypervariable region of E2/NS1 region of hepatitis C virus in normal and hypogammaglobulinemic
patients. Hepatology27:223–227.
9.Bukh, J., R. H. Miller, and R. H. Purcell.1995. Genetic heterogeneity of
hepatitis C virus: quasispecies and genotypes. Semin. Liver Dis.15:41–63.
10. Bukh, J., R. H. Purcell, and R. H. Miller.1993. At least 12 genotypes of hepatitis C virus predicted by sequence analysis of the putative E1 gene of
isolates collected worldwide. Proc. Natl. Acad. Sci. USA90:8234–8238.
11. Cerino, A., M. Bissolati, A. Cividini, A. Nicosia, M. Esumi, N. Hayashi, K. Mizuno, R. Slobbe, P. Oudshoorn, E. Silini, M. Asti, and M. U. Mondelli.
1997. Antibody responses to the hepatitis C virus E2 protein: relationship, to viremia and prevalence in anti-HCV seronegative subjects. J. Med. Virol.
51:1–5.
12. Chang, K. M., B. Rehermann, and F. V. Chisari.1997. Immunopathology of
hepatitis C. Springer Semin. Immunopathol.19:57–68.
13. Chien, D. Y., Q.-L. Choo, R. Ralston, R. Spaete, M. Tong, M. Houghton, and G. Kuo.1993. Persistence of HCV despite antibodies to both putative
en-velope glycoproteins. Lancet342:933. (Letter.)
14. Choo, Q.-L., G. Kuo, R. Ralston, A. Weiner, D. Chien, G. Van Nest, J. Han,
on November 9, 2019 by guest
http://jvi.asm.org/
K. Berger, K. Thudium, C. Kuo, J. Kansopon, J. McFarland, A. Tabrizi, K. Ching, B. Moss, L. B. Cummins, M. Houghton, and E. Muchmore.1994. Vaccination of chimpanzees against infection by the hepatitis C virus. Proc.
Natl. Acad. Sci. USA91:1294–1298.
15.Choo, Q.-L., K. H. Richman, J. H. Han, K. Berger, C. Lee, C. Dong, C. Gallegos, D. Corr, A. Medina-Selby, P. J. Barr, A. J. Weiner, D. W. Bradley, G. Kuo, and M. Houghton.1991. Genetic organization and diversity of the
hepatitis C virus. Pro. Natl. Acad. Sci. USA88:2451–2455.
16. Eckels, D., P. Flomenberg, and J. C. Gill.1996. Hepatitis C virus: models of
immunopathogenesis and prophylaxis. Tranfusion36:836–844.
17. Enomoto, N., C. Sato, M. Kurosaki, and F. Marumo.1994. Hepatitis C virus after interferon treatment has the variation in the hypervariable region of
envelope 2 gene. J. Hepatol.20:252–261.
18. Farci, P., J. Bukh, and R. H. Purcell.1997. The quasispecies of hepatitis C
virus and the host immune response. Springer Semin. Immunopathol.19:5–
26.
19. Farci, P., A. Shimoda, D. Wong, T. Cabezon, D. De Gioannis, A. Strazzera, Y. Shimizu, M. Shapiro, H. J. Alter, and R. H. Purcell.1996. Prevention of hepatitis C virus infection in chimpanzees by hyperimmune serum against the hypervariable region 1 of the envelope 2 protein. Proc. Natl. Acad. Sci.
USA93:15394–15399.
20. Fong, T. L., S. R. Lee, W. K. Briggs, B. Valinluck, S. Govindarajan, A. Hoffman, B. Jaczko, and A. G. Redeker.1996. Clinical significance of hep-atitis C viral RNA status and its correlation to antibodies to structural HCV antigens in anti-HCV reactive patients with normal liver tests. J. Med. Virol.
49:253–258.
21. Harada, S., R. Suzuki, A. Ando, Y. Watanabe, S. Yagi, T. Miyamura, and I. Saito.1995. Establishment of a cell line constitutively expressing E2 glyco-protein of hepatitis C virus and humoral response of hepatitis C patients to
the expressed protein. J. Gen. Virol.76:1223–1231.
22. Hilfenhaus, J., U. Krupka, T. Nowak, L. B. Cummins, K. Fuchs, and M. Roggendorf.1992. Follow-up of hepatitis C virus infection in chimpanzees: determination of viraemia and specific humoral immune response. J. Gen.
Virol.73:1015–1019.
23. Houghton, M., A. Weiner, J. Han, G. Kuo, and Q.-L. Choo.1991. Molecular biology of the hepatitis C viruses: implications for diagnosis, development
and control of disease. Hepatology14:381–388.
24. Hu¨ssy, P., H. Faust, J.-C. Wagner, G. Schmid, J. Mous, and H. Jacobsen.
1997. Evaluation of hepatitis C virus envelope proteins expressed inE. coli
and insect cells for use as tools for antibody screening. J. Hepatol.26:1179–
1186.
25. Kato, N., Y. Ootsuyama, T. Tanaka, M. Nakagawa, T. Nakazawa, K. Mu-raiso, S. Ohkoshi, M. Hijikata, and K. Shimotohno.1992. Marked sequence diversity in the putative envelop proteins of hepatitis C viruses. Virus Res.
22:107–123.
26. Kumar, U., J. Monjardino, and H. C. Thomas.1994. Hypervariable region of hepatitis C virus envelope glycoprotein (E2/NS1) in an agammaglobulinemic
patient. Gastroenterology106:1072–1075.
27. Kurosaki, M., N. Enomoto, F. Marumo, and C. Sato.1993. Rapid sequence variation of the hypervariable region of hepatitis C virus during the course of
chronic infection. Hepatology18:1293–1299.
28. Laemmli, U. K.1970. Cleavage of structural proteins during the assembly of
the head of bacteriophage T4. Nature227:680–685.
29. Lanford, R. E., and J. S. Butel.1979. Antigenic relationship of SV40 early
proteins to purified large T polypeptide. Virology97:295–306.
30. Lanford, R. E., K. D. Carey, L. E. Estlack, G. C. Smith, and R. V. Hay.1989. Analysis of plasma protein and lipoprotein synthesis in long-term primary cultures of baboon hepatocytes maintained in serum-free medium. In vitro
Cell. Dev. Biol.25:174–182.
31. Lanford, R. E., L. Notvall, L. H. Barbosa, and J. W. Eichberg.1991. Eval-uation of a chimpanzee colony for antibodies to hepatitis C virus. J. Med.
Virol.34:148–153.
32. Lanford, R. E., L. Notvall, and B. Beames.1995. Nucleotide priming and reverse transcriptase activity in hepatitis B virus polymerase expressed in
insect cells. J. Virol.69:4431–4439.
33. Lanford, R. E., L. Notvall, D. Chavez, R. White, G. Frenzel, C. Simonsen, and J. Kim.1993. Analysis of hepatitis C virus capsid, E1, and E2/Ns1
proteins expressed in insect cells. Virology197:225–235.
34. Lesniewski, R., G. Okasinski, R. Carrick, C. Van Sant, S. Desai, R. Johnson, J. Scheffel, B. Moore, and I. Mushahwar.1995. Antibody to hepatitis C virus second envelope (HCV-E2) glycoprotein: a new marker of HCV infection
closely associated with viremia. J. Med. Virol.45:415–422.
35. Michalak, J. P., C. Wychowski, A. Choukhi, J. C. Meunier, S. Ung, C. M. Rice, and J. Dubuisson.1997. Characterization of truncated forms of
hep-atitis C virus glycoproteins. J. Gen. Virol.78:2299–2306.
36. Mink, M. A., S. Benichou, P. Madaule, P. Tiollais, A. M. Prince, and G.
Inchauspe.1994. Characterization and mapping of a B-cell immunogenic domain in hepatitis C virus E2 glycoprotein using a yeast peptide library.
Virology200:246–255.
37. National Institutes of Health.1997. National Institutes of Health Consensus development conference panel statement: management of hepatitis C.
Hepa-tology26:2S–10S.
38. Nishihara, T., C. Nozaki, H. Nakatake, K. Hoshiko, M. Esumi, N. Hayashi, K. Hino, F. Hamada, K. Mizuno, and T. Shikata. 1993. Secretion and purification of hepatitis C virus NS1 glycoprotein produced by recombinant
baculovirus-infected insect cells. Gene129:207–214.
39. Odeberg, J., Z. B. Yun, A. So¨nnerborg, K. Bjoro, M. Uhle´n, and J. Lunde-berg.1997. Variation of hepatitis C virus hypervariable region 1 in
immu-nocompromised patients. J. Infect. Dis.175:938–943.
40. Ogata, N., H. J. Alter, R. H. Miller, and R. H. Purcell.1991. Nucleotide sequence and mutation rate of the H strain of hepatitis C virus. Proc. Natl.
Acad. Sci. USA88:3392–3396.
41. Okada, S.-I., Y. Akahane, H. Suzuki, H. Okamoto, and S. Mishiro.1992. The degree of variability in the amino terminal region of the E2/NS1 protein of hepatitis C virus correlates with responsiveness to interferon therapy in
viremic patients. Hepatology16:619–624.
42. Okamoto, H., M. Kojima, S.-I. Okada, H. Yoshizawa, H. Iizuka, T. Tanaka, E. E. Muchmore, D. A. Peterson, Y. Ito, and S. Mishiro.1992. Genetic drift of hepatitis C virus during an 8.2-year infection in a chimpanzee: variability
and stability. Virology190:894–899.
43. Rosa, D., S. Campagnoli, C. Moretto, E. Guenzi, I. Cousens, M. Chin, C. Dong, A. J. Weiner, J. Y. N. Lau, Q. L. Choo, D. Chien, P. Pileri, M. Houghton, and S. Abrignani.1996. A quantitative test to estimate neutral-izing antibodies to the hepatitis C virus: cytofluorimetric assessment of en-velope glycoprotein 2 binding to target cells. Proc. Natl. Acad. Sci. USA
93:1759–1763.
44. Summers, M. D., and G. E. Smith.1987. A manual of methods for baculo-virus vectors and insect cell culture procedures. Tex. Agric. Exp. Stn. Bull.
1555:1–48.
45. Thomas, D. L., D. Vlahov, L. Solomon, S. Cohn, E. Taylor, R. Garfein, and K. E. Nelson.1995. Correlates of hepatitis C virus infections among injection
drug users. Medicine (Baltimore)74:212–220.
46. Van Doorn, L.-J., I. Capriles, G. Maertens, R. DeLeys, K. Murray, T. Kos, H. Schellekens, and W. Quint.1995. Sequence evolution of the hypervariable region in the putative envelope region E2/NS1 of hepatitis C virus is
corre-lated with specific humoral immune responses. J. Virol.69:773–778.
47. Van Doorn, L.-J., K. van Hoek, G. de Martinoff, F. Bosman, L. Stuyver, T. Kos, I. Frantzen, P. Sillekens, G. Maertens, and W. Quint.1997. Serological and molecular analysis of hepatitis C virus envelope regions 1 and 2 during
acute and chronic infections in chimpanzees. J. Med. Virol.52:441–450.
48. Walker, C. M.1997. Comparative features of hepatitis C virus infection in
humans and chimpanzees. Springer Semin. Immunopathol.19:85–98.
49. Wang, Y. F., B. Brotman, L. Andrus, and A. M. Prince.1996. Immune response to epitopes of hepatitis C virus (HCV) structural proteins in
HCV-infected humans and chimpanzees. J. Infect. Dis.173:808–821.
50. Weiner, A. J., M. J. Brauer, J. Rosenblatt, K. H. Richman, J. Tung, K. Crawford, F. Bonino, G. Saracco, Q. L. Choo, M. Houghton, et al.1991. Variable and hypervariable domains are found in the regions of HCV cor-responding to the flavivirus envelope and NS1 proteins and the pestivirus
envelope glycoproteins. Virology180:842–848.
51. Wray, W., T. Boulikas, V. P. Wray, and R. Hancock.1981. Silver staining of
proteins in polyacrylamide gels. Anal. Biochem.118:197–203.
52. Yeh, B. I., K. H. Han, S. H. Oh, H. S. Kim, S. H. Hong, and Y. S. Kim.1996. Nucleotide sequence variation in the hypervariable region of the hepatitis C virus in the sera of chronic hepatitis C patients undergoing controlled
inter-feron-atherapy. J. Med. Virol.49:95–102.
53. Yoshioka, K., Y. Higashi, K. Tanaka, T. Aiyama, M. Takayanagi, A. Oku-mura, K. Iwata, Y. Nagai, and S. Kakumu.1996. Deficiency of antibody response to hypervariable region of hepatitis C virus in patients with chronic
hepatitis C. J. Hepatol.24:649–657.
54. Yuki, N., N. Hayashi, A. Kasahara, H. Hagiwara, E. Mita, K. Ohkawa, K. Katayama, H. Fusamoto, and T. Kamada.1996. Quantitative analysis of antibody to hepatitis C virus envelope 2 glycoprotein in patients with chronic
hepatitis C virus infection. Hepatology23:947–952.
55. Yuki, N., N. Hayashi, T. Moribe, Y. Matsushita, T. Tabata, T. Inoue, Y. Kanazawa, K. Ohkawa, A. Kasahara, H. Fusamoto, and T. Kamada.1997. Relation of disease activity during chronic hepatitis C infection to complexity
of hypervariable region 1 quasispecies. Hepatology25:439–444.
56. Zibert, A., H. Meisel, W. Kraas, A. Schulz, G. Jung, and M. Roggendorf.
1997. Early antibody response against hypervariable region 1 is associated
with acute self-limiting infections of hepatitis C virus. Hepatology25:1245–
1249.