Copyright0 1974 AmericanSociety for Microbiology Printedin U.S.A.
Newcastle Disease Virus-Specific
RNA: Polyacrylamide Gel
Analysis
of
Single-Stranded
RNA
and
Hybrid Duplexes
NICOLAI V. KAVERIN ANDNATALIA L. VARICH
Ivanovsky Institute of Virology,Academy of MedicalSciences, U.S.S.R.
Received for publication6July 1973
Newcastledisease virus-specific [3H
]uridine-labeled
18SRNAwasresolved bypolyacrylamide gel electrophoresis into several components with molecular weights from 450,000to 840,000.The analysis of35and 24S virus-specificRNA
alsorevealed several componentsineachsedimentationalclass. The conversion
of 18S RNA into double-stranded form by hybridization with an excess of
unlabeled virion RNA improved the resolution in polyacrylamide gels and revealedatleast six distinctcomponents.Thesamesixclasses ofhybrid duplexes wererevealed when 32P-labeled50Svirion RNAwashybridizedwithanexcessof
18S RNA. The applicability of polyacrylamide gel electrophoresis of hybrid duplexestotheanalysis of viral genome structure isdiscussed.
A large part ofvirus-specific RNA formedin
paramyxovirus-infected cells is represented by
molecules ofsubgenomic size (18 to 35S)
com-plementary to virion RNA (1, 2). Messenger
function in the synthesis ofvirus proteins was
tentatively ascribed to the complementary
RNA
by
Kingsbury asearlyas1966 (11). Sincethenanumber of datawerereported confirming
this suggestion. The most
abundant
class ofcomplementary
RNA,
18SRNA, possessessev-eral characteristics which make ita
particularly
good candidate for the role of paramyxoviral
mRNA: a "monocistronic" size
(-
7 x 105daltons), the presence of
polyadenylic
acid-sequences (19), and
heterogeneity.
The latterwas proposed in order toexplain theability of
18S RNAto convert alargepart of virionRNA
(over 50%, i.e., >3 x 106
daltons)
intodouble-stranded form
by hybridization
(2).Polyacryl-amide gel
electrophoresis
ofparamyxovirus-specific RNA revealed a
heterogeneity
in theregion
corresponding
to 18SRNA, butnoindi-vidual specieswereresolved (12, 13).
In this paper we present a series of
experi-ments onpolyacrylamide gel
analysis
ofvirus-specific RNA isolated from Newcastle disease
virus (NDV)-infected cells. Both
single-stranded RNAs andhybrid duplexes were
ana-lyzed. The latter approach was used with a
double purpose: first, toimprovetheresolution
ofRNAspeciesinthe
gel;
second,
as anattemptto find out whether individual species of 18S
RNA
correspond
tospecific
template regions
inthe genomic RNA. For the first purpose the
hybrids oflabeled 18S RNA with an excessof
virion RNA weresubjected to gel electrophore-sis. For the second purpose the products of
hybridizationoflabeledvirion50S RNAwithan
excess of18S RNAwere analyzed.
MATERIALS AND METHODS
Egg-grown Beaudette strain of NDV (thermostable
clone C) and chicken embryo cell (CEC) monolayer
cultures were used. Theprocedures of infection and
[3H]uridine-labeling of the cells as well as labeling
and purification of the virus, RNA extraction, and
ratezonal centrifugation have been described (8).
RNA-RNA hybridization. The procedure
de-scribed by Kingsbury (11) was used with slight
modifications (8). If the product of hybridization had
tobeanalyzed further, the annealing was performed
ina2-mlvolume. Afterannealing, RNasewasadded
(final concentration 10ug/ml); thesample was
incu-bated at37C for 30 min, treated with Pronase (100
ug/ml,60minat37C) in ordertodestroyRNase(6),
and extracted twice with phenol and precipitated
with 2 volumes of ethanol and one-tenth volume of
16%sodiumacetate.
Polyacrylamide gel electrophoresis. The
proce-duredescribedby Schincarioland Howatson(21)was
used withsomemodifications. Thegels with
acrylam-ide concentrations of2.0% or 2.4%(wt/vol)and0.5%
agarosewereused. Thepolymerizationwasperformed
at37C in tubes10 cmlongwithaninternaldiameter
of 0.8 cm. The gelswereprerun for 1h at6 mAper
tube. Tento 20
usg
ofRNAin 50Mliterswerelayeredover the gels. Theelectrophoresis was performed at
roomtemperaturefor 3 to 5 h at 6 mApertube. The
gels were either stained with methylene blueor cut
with arazor-blade device intoslices.Thelatterwere
dissolved in 0.2mlof 30%water at70C,mixedwith
15ml ofscintillation fluid (2,5-diphenyloxazole,4g;
1,4-bis-[5-phenyloxazoly]benzene, 0.1 g; ethanol,
253
on November 10, 2019 by guest
http://jvi.asm.org/
300 ml; toluene, 700 ml) and counted in a TriCarb scintillation counter. Ribosomal RNAs of E. coli and
ofCEC wereused as markers.
Reagents. Dactinomycin (Serva, BRD), agarose
(Koch-Light,England),acrylamide (BDH,England),
sodiumdodecylsulphate (SDS) (Matheson, Coleman
andBell,England), Pronase (Calbiochem, Los
Ange-les, Calif.), pancreatic ribonuclease (Calbiochem)
[3H]uridine (Amersham, England), carrier-free
[32P]orthophosphoric acid (Isotope, U.S.S.R.) were
used.
RESULTS
Preliminary sedimentational fractionation
and polyacrylamide
gel
analysis
ofsingle-stranded
virus-specific
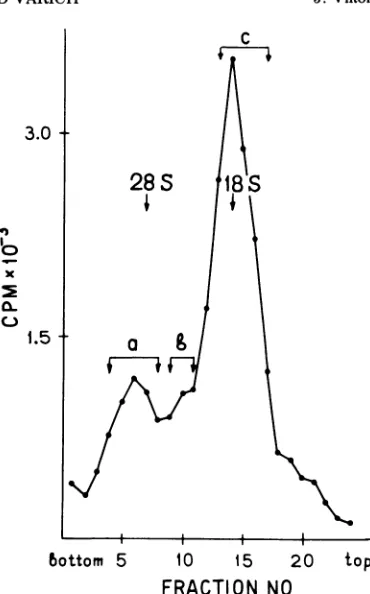
RNA. Thedistributionofvirus-specific RNA in an SDS-sucrose density gradient is shown in Fig. 1. Virus-specific RNA is distributed in three broad peaks: 35, 24 and
18SRNA. Genome-size 50S RNAsediments to
the bottom of the tube under these conditions of centrifugation. Sucha pattern is characteristic
forparamyxovirus-specific RNA extractedfrom
infected cells (1, 2). Peakfractions werepooled
as shown in Fig. 1, precipitated with ethanol,
extracted withphenol forcomplete removal of
SDS,
once moreprecipitated
withethanol,
andapplied to
polyacrylamide gel.
Thepolyacryl-amide gel electrophoresisrevealed several
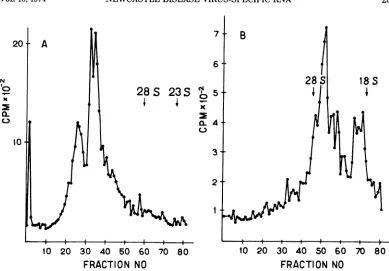
com-ponents in 35 and in 24S RNA (Fig. 2). Our
attention was confined mostly to 18SRNA for reasons discussed aboNe. The
analysis
of 18S RNA in 2% gel revealed a heterogeneity: twofaster moving components formed distinct
peaks; the rest were notresolved and formeda
slower moving zone (Fig. 3A). The analysis in
2.4% gel revealed six components (Fig.
3C).
Component VI in Fig. 3C is
represented by
ashoulder, but in several experiments it was
revealed as adistinct peak. For the calculation
ofmolecular weights ofthe individual
compo-nents ofvirus-specific 18SRNA, we used
ribo-somal RNAs with known molecularweights as
standard markers. We ran 28 and 18S RNAof
CEC as well as 23 and 16S RNA ofE. coliin
parallel tubesand stainedthem with
methylene
blue. The molecular weights for 28 and 18S
RNA were 1.65 x 106 and 0.67 x
106,
respec-tively (16), whereas for 23 and 16S the values
were 1.1 x 106 and 0.53 x 106 (16). The calculated molecular weights of individual
NDV-specific 18S RNA components are
sum-marized in Table 1.
Hybridization of the labeled virus-specific
18S RNA with an excess ofvirion RNA and
the analysis of hybrids in polyacrylamide
gel. The conversion of single-stranded RNA
into double-stranded form by hybridization
3.0
C~)
xa-1.5
bottom 5
1015
20
top
FRACTION
NO
FIG. 1. Fractionation ofvirus-specific NDV RNA
by SDS-sucrose density gradient centrifugation.
NDV-infected CEC were labeled with [3H]uridine
from 7 to10hpostinfection after 2 h of pretreatment
with dactinomycin (2
Ag/ml).
SDS-phenol-extractedRNA was layered on 50-ml 15 to 30% SDS-sucrose
density gradient and centrifuged inanS W-25.2 rotorof
aSpinco L2ultracentrifuge at 20,000 rpm for 16 h at
25 C. Fractionswerecollected and radioactivity was
determinedinsamples (one-twentiethof the fraction
volume). Chosen fractions werepooled as shown in
the figure (a, b, c) for further analysis. Arrows
indicate theposition of CEC 28 and 18S ribosomal
RNA(28.8 and17.5Svedberg units as determined by
analytical centrifugation in a Spinco E
ultracentri-fuge).
withcomplementarystrands has been shown to
facilitatethe resolution of RNA in
polyacrylam-ide gels (7). Annealing of [3H
]uridine-labeled
NDV-specific 18SRNA with an excess of
unla-beled virion RNA rendered 95 to 100% of the
labelribonuclease resistant. When the product
of hybridization after ribonuclease-Pronase
treatment (see Materials and Methods) was
analyzed in2% gel, six peaks were consistently
resolved (Fig. 4). The components may be
tentatively identified with the peaks of
single-stranded RNA(Fig.3C). If the doubled
molecu-lar weight of the respective single-stranded
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.263.448.63.360.2]VIRUS-SPECIFIC
N
0
C.L
7t
B
6
C 5
x
t0 4
28
3
2
18S
0
20
30 40 50
60 70 80
10 20 30 40 50 60 70 80 [image:3.493.49.438.62.333.2]FRACTION NO
FRACTION
NO
FIG. 2. Electrophoresis of 35 and 24SNDV-specific RNA in polyacrylamide gel. RNA from pooled sucrose
gradientfractions (a and bregions, Fig. 1) was precipitated with ethanol and re-extracted with phenol, and a
partofit was analyzed in2%oacrylamide-0.5% agarose gel. A, 35S RNA (region a) electrophoresis during 5h;
B, 24S RNA(region b)electrophoresisduring4h. Arrowsindicate the positions of marker RNA run in parallel
gels.Migration here and in all electrophoregrams is toward the anode on the right.
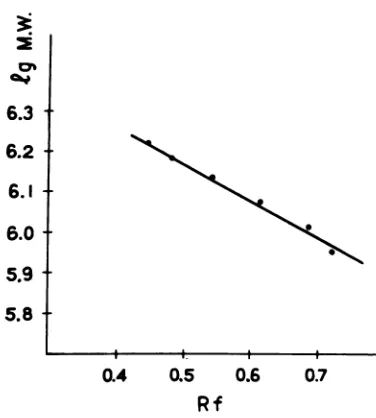
component is ascribed to each class of the
hybrids, the dependenceoftheir relative
mobil-ity on the log of molecular weightseems to be
fairly linear (Fig. 5). This confirms toacertain
extentthe identification ofhybrid classes with
individual single-stranded components. This
argument, however, is based on anassumption
that the dependenceofrelativemobility onlog molecular weight described forsingle-stranded RNA (14) holds also forhybrid duplexes.
Hybridization of labeled 50S virion RNA
withanexcessofvirus-specific 18SRNAand
the analysis of hybrids in polyacrylamide
gel. Virion RNA ofegg-grown Beaudette strain
of NDV is represented by molecules with a single polarityand does notproduceany
signifi-cant amount of double-stranded structures
when self-annealed (9, 18), differing in this
respect from Sendai virus (18, 20). For this
reason labeled virion NDV RNA may be used
forannealingwithanexcessofcomplementary virus-specific RNAforasubsequent analysisof hybridization product.
Labeled virion50SRNA wasobtained either
from [32P]labeled NDV purified by centrifuga-tion through potassiumtartrate (8, 10) orfrom
the virus partially purified by differential
cen-trifugation. In the latter case thefractionation
of RNA in SDS-sucrose gradient provides a
sufficiently pure preparation of50SRNA (Fig.
6), whereas the lossofvirusduring purification
isnegligible.
The
hybridization
withanexcess ofunlabeled18S RNA should convert 50 to 60% of virion
RNA into
double-stranded
form (2).To besurethat 18S RNA was really present in excess, a
series ofdilutionsof18S RNApreparation was
used in every experiment. The amount of 18S
RNAinthesample usedforfurtheranalysis, as
shown in Table 2, may be considered
saturat-ing. Theproduct ofhybridization after
ribonu-clease treatment had a sedimentation coeffi-cient closetotheoneexpectedforthe duplexof
18SRNAmolecule (Fig. 7). The analysisofthe
product of annealing inpolyacrylamidegel(Fig.
8) revealedthe samecomponentsasthe analysis
ofhybrids oflabeled 18SRNA (Fig. 4).
Itshould bekeptinmindthatRNAduplexes
inFig.8arerepresentedbyfragments oflabeled
viralgenome RNAhybridizedwith correspond-ing unlabeled virus-specific RNA. Such frag-mentsmay beexpectedtobepresent in
equimo-255
on November 10, 2019 by guest
http://jvi.asm.org/
9
A
8
7
N O6
5
C-,
4
3
2
10 20 30 40 50 60 70 80
FRACTION NO
B
28
S
18S
10 20 30 40 50 60 70 80
FRACTION NO
C
28S 23S
20 i
N~ ~ ~ FATO NO
C-)
10
10 20 30 40 50 60 70
[image:4.493.61.455.60.544.2]FRACTION NO
FIG. 3. Electrophoresis of NDV-specific 18S RNA inpolyacrylamidegel. A, Total RNA extracted from
un-infected CEC labeled for 3 h with [3H]uridinewassubjectedtoelectrophoresis in 2%o gel for3.25h.B, Virus-specific 18S RNAwasisolatedfromsucrosedensitygradient (Fig.1, region c) andapartof itwassubjectedto
electrophoresis in 2%gel for 3.25 h. C, 18S RNAwasanalyzed in 2.4% gel for4h.
lar amounts. In an attempt to calculate the relations obtained in three experiments are
molarrelations,weascribedthedoubled molec- shown inTable 3. The componentsIII, IV, and
ularweight of the correspondingsingle-stranded V were consistently present in approximately
component to each class ofthe duplexes and equimolar amounts, whereas the molar
rela-dividedthesumofradioactivityineach peak by tions for components I, II, and VI were lower
the respective molecular weight. The molar and morevariable. 12
cN .
0-C.,)
6-I
1*
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Molecularweights of the componentsof
NDV-specific 18SRNAa
Component Mol wtb No. of
no. (x 10-') determinations
I 446 1
II 521 9.9 4
III 608 7.8 6
IV 696 4.9 6
V 760 7.2 6
VI 840 9.7 6
aThree different preparations of
virus-specific
3H-labeled 18S RNA were used. Peak I either migrated
out ofthegel (if an attempt was made to resolve the
heavier components) or was poorly resolved. One of
the experiments is shown in Fig. 3C. 'Mean ± standard error.
4.
N
O 3
x
a.
C-,
2
10 20 30 40 50 60 70 80
FRACTION NO
FIG. 4. Polyacrylamide gel electrophoresis of
la-beled 18S RNA converted into double-strandedform
by hybridization with an excess of virion RNA.
3H-labeled virus-specific RNA was isolated as
de-scribed in the legend to Fig. 1 and annealed with
unlabeled virion RNA.A 300-agportionoftotal 18S
RNA containing200,000counts/minoflabeled
virus-specificRNAwasmixed with290ugofunlabeled 50S
RNA,annealed,andtreatedasdescribed in Materials
and Methods. The annealingrendered
ribonuclease-resistant91%of3H-18S RNA. Theresulting
prepara-tionwasdissolved in 0.2 mlof electrophoresis buffer.
A0.05-mlportionwasappliedto2%gelandrunfor4
h.
DISCUSSION
The heterogeneity of
paramyxovirus-specific
18S RNA had been shown with the use of
polyacrylamide gel
electrophoresis
for Sendaivirus (12) and for NDV (13),
although
thenumber of size classes hadnotbeendetermined.
Rhabdovirus 13S RNA, which is similar to
paramyxoviral 18S RNA in many respects,had
been shown to consist of several (up to eight) components (21, 22). In the experiments pre-sented in this paper, the resolution power of the
gel wasincreased (7) by conversion of 18S RNA
into hybrid duplexes, and the population of18S
RNA was resolved into sixdistinct components (Fig. 4). The range of the sizes of differenit
classes of
RNA,
asdetermined by the analysis of single-stranded 18S RNA preparation (Table 1) roughly corresponds to the expected size of mRNAs for viral proteins (14). The sum of molecular weights of allthe components is -3.4 x 106,i.e.,
-50 to 60% ofthemolecular
weightof virion RNA (4, 5). This value is in good agreement with the size of the part of viral genome RNA which may be converted into
double-stranded formby hybridization withan
excess of18SRNA (2, 8; see alsoTable 2) and
probably serves as a template for 18S RNA
synthesis.
Itshould be takeninmind,however, that the
analysisofsizedistribution of18S RNAcannot
give an answer to the question whether a
particular RNA component has an individual
base sequence,i.e., whether itis atranscript of
3t
6.3
6.2
6.1
6.0
+
5.9
-5.8
S 0
0.4
0.5 0.6 0.7Rf
FIG. 5. A dependence between relative mobility
and assumed molecular mass of hybrid duplexes.
Relative mobility ofhybrid duplexes was calculated
from theexperimentrepresentedinFig.4.Sixpoints
correspond to the sixpeaksresolved in thegel. The
individual value ofmolecular mass foreach hybrid
component is taken to be equal to the doubled
molecularweightof thecorrespondingsingle-stranded
component(Table 1).
257
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.43.236.258.468.2] [image:5.493.246.434.371.580.2]
2C-0
C-)
iS
l+
Bottom 5 10 15 20 Top
FRACTION NO
FIG. 6. Isolation of 50S RNAfrom partially
puri-fied NDV. RNA was extracted from 32P-NDV
par-tiallypurified by differential centrifugation, layered
on a 39-ml 15 to 30% SDS-sucrose gradient,
cen-trifugedfor15hat 18,000 rpm and25C inanSW27
rotorofaSpinco L 65ultracentrifuge. Radioactivity
wasdetermined insamples,andfractionswerepooled
asshown in thefigure.
TABLE 2. Demonstrationofsaturatingamountof
NDV-specific 18S RNAatannealingwith32P-labeled
virion50S RNAfor furtheranalysisof hybridization
producta
18S RNAfromNo.RNase RNase
Sample NDV-infected (counts treat- Annealed
no. cellsa g
mmin)
mentsample)'
1 70 640C 410 64
2 53 636 424 66.5
3 35 610 381 62.5
4 1.3 716 37 5.2
5 0 690 23 3.3
a32P-labeled 505virion RNA (0.1Ag, 1,300counts
per min per sample) was annealed with different
amountsof 185 RNA.Inthereactionmixture usedfor
furtheranalysis (Fig. 7, 8), theexcess of18 over505
RNAwasthesame asinsample 1 inthetable.
ITotalamountofRNAinthesample, represented
mostly by cellular RNAfrom the188 zone.
cMean oftwoparallelsamples.
an individual gene, a fragment of a larger
transcript, or a transcript of two
neighboring
individual genes. The latter possibility cannot
be
disregarded
as it had beenreported
thatnucleotidesequences identical to 18S RNAare
present in35S RNA (2).
To study the regions of 50S virion RNA
serving astemplates forvirus-specific 18S RNA,
we attempted to cutthe labeled 50SRNA into
fragments complementary to individual size
classes of 18S RNA. The possibility of this
approachwas notevidentapriori: it wasbased
on a presumption of the existence of "spacer"
sequences among the template regions in 50S RNA. Such spacers would remain
single-stranded and ribonuclease-sensitive after
an-nealing with an excess of 18S RNA. If such
spacers were absent, the annealing would
pro-duce a population of hybrids containing a 50S
RNA molecule as one strand and several 18S
RNA molecules stuck end-to-end as the other
strand. In this caseribonuclease might be not
quite efficient in "cutting" such structures
between twoadjacent 18S RNA molecules.
The result shown inFig. 8indicates that the
"cutting" is performed at appropriate sites,
because thepopulation oftheduplexes (Fig. 8)
issimilarinthe number and size of the
compo-nents totheoneobtained afterhybridization of
6
28 S
18S
4S
C-,x
X
bottom 10 20 30 40 50 top
FRACTION NO
FIG. 7. Sucrose density centrifugation of hybrid
duplexes obtained by annealing of 32P-virion RNA
with an excess of18S RNA. 32P_virion50S RNA (3
,og,27,000counts/min)isolatedasshown inFig.6was
annealed with 2 mg of unlabeled 18S RNA from
infected cells. These conditions were shown to be saturating (Table 1). The product of hybridization
wastreated asdescribed in the text and divided into
two equal parts. One part was analyzed in sucrose
density gradient. The speed, 23,000 rpm, and the
otherconditions of centrifugation were as described in
thelegend toFig. 1. Theotherpart was analyzed in
polyacrylamide gel (Fig. 8).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.62.248.59.325.2] [image:6.493.263.444.346.550.2] [image:6.493.54.250.427.544.2]20
X 16
C.,
10 20 30 40 50 60 70 8o
FRACTION NO
FIG. 8. Polyacrylamide gel analysis of hybrid
du-plexes obtained by annealing of 32P-virion RNA with
an excess of 18S RNA. The product of the
hybridiza-tion experiment described in Fig. 7was analyzed in
polyacrylamidegel.Electrophoresis in2%ogel for4.25
h.
labeled 18S RNA withanexcessof virionRNA (Fig. 4).
Itshould be noted that if eachcomponent of
18S RNA isa transcript ofan individual tem-plate region, the amount of label in hybrid peaks (Fig. 8) should be distributed in equimo-lar ratios. As one can see from Table 3, this is
notgenerally thecase: only thecomponentsIII,
IV, and V are present in equimolar amounts, whereas there is a deficit of the label in the otherpeaks. The values of molar ratios for the components I, II, and VI were variable, but always less than 1.0, especially for the
compo-nentVI. Thesumofradioactivity in peaks I, II,
and VIslightly exceeds the amount needed for
the component VI to be present in equimolar
relation with components III, IV, and V. One
cannot exclude the following possibility: the
component VI is a composite of components I
and II, i.e., an mRNA-transcript oftwo neigh-boringgenes.Suchasituationcould introducea
biasinto the equimolardistribution of the label
in the population of hybrid duplexes. These considerations indicatethat, although the
popu-lation of 18S RNA isrepresented byatleast six
components (Fig. 3, 4), one cannot conclude
with certainty whether theyarethetranscripts ofsix or five template regions of the genomic
RNA.
Thedatapresentedin thispaperindicate the
applicability ofpolyacrylamide gel electropho-resis ofhybrid duplexes for the analysis of the
viralgenomeswhicharenotfragmentedinsitu.
Further elaboration of the method and its
[image:7.493.45.228.59.235.2]application to DNA-containing viruses might facilitate the study of viral genome structure and function.
TABLE 3. Molar relations of 32P-hybrid duplexesa
Componentno.
Exptno.
I II III IV V VI
1 0.85 0.85 1.07 0.95 1.0 0.22
2 0.52 0.67 1.09 1.03 1.0 0.74
3 0.69 0.79 0.93 1.03 1.0 0.43
aThe sum ofradioactivityineachpeak was divided
bythe assumedmolecularweightof thecorresponding
component. The value for component V is taken as
1.0.
b The conditions of annealing and the reaction
mixture weresimilar to those described in the legend
toFig. 7. The fractionationprocedure in the gelwas as
inFig. 8.
After this paper had been submitted for
publication, we became acquainted with a
re-cent work on this subject (3) where
virus-specific 18S RNA is resolved into several species
inpolyacrylamide gel. Our results(Fig.3C) are
in good agreement with the data published by
Collins andBratt.
ACKNOWLEDGMENTS
We wishtothankG. Petrovskyforthe analytical determi-nation ofsedimentation coefficient of ribosomal RNA and T. Logutenkovaforexcellent technical assistance.
LITERATURE CITED
1. Blair, C. D., and W. S. Robinson. 1968. Replication of Sendai virus. I. Comparison ofviral RNA and virus-specific RNA synthesis to Newcastle disease virus. Virology 35:537-549.
2. Bratt, M. A., and W.S. Robinson. 1967. Ribonucleic acid synthesis in cells infected with Newcastle disease virus. J.Mol. Biol. 23:1-21.
3. Collins. and M. A. Bratt. 1973. Proc. Nat. Acad. Sci. U.S.A. 70:2544-2548.
4. Compans,R.W., and P. W. Choppin.1968.Thenucleic acid of the parainfluenza virus SV5. Virology 35:289-296.
5. Duesberg, P. H. 1968. Physical propertiesofRous sar-coma virus RNA. Proc. Nat. Acad. Sci. U.S.A. 60:1511-1518.
6. Gillespie, D., and S. Spiegelman. 1965. A quantitative assayforDNA-RNAhybrids with DNA immobilizedon amembrane. J. Mol. Biol.12:829-842.
7. Ito, Y., and W. K. Joklik. 1972. Temperature-sensitive mutantsofreovirus. I. Patterns of gene expressionby mutantsof groupsC, Dand E.Virology 50:189-201. 8. Kaverin, N. V., and N. L. Varich. 1971. Virus-specific
RNA formed inNewcastle disease virus-infected cells after suppression of protein synthesis by cyclohexi-mide. Arch. Gesamte Virusforsch.35:378-384. 9. Kaverin, N.V., andN. L. Varich. 1972. Sedimentation
andhybridizationanalysisofvirus-specific RNAinthe course of Newcastle disease virus replication cycle.
Arch.Gesamte Virusforsch. 38:183-191.
10. Kingsbury, D. W. 1966.Newcastle diseasevirusRNA. I. Isolation and preliminary characterization of RNA fromvirusparticles. J. Mol. Biol. 18:195-203. 11. Kingsbury,D. W. 1966.Newcastledisease virus RNA.II.
Preferential synthesisofRNA complementary to pa-rentalviral RNAby chick embryocells. J. Mol. Biol. 18:203-214.
VOL.13,1974
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.493.243.439.82.213.2]12. Kingsbury, D. W., A. Portner, and R. W. Darlington. 1970. Properties of incomplete Sendai virions and subgenomic viral RNAs. Virology 42:857-871. 13. Lomniczi, B., A.Meager, and D. C. Burke. 1971.Virus
RNA and protein synthesis in cells infected with different strains of Newcastle disease virus. J. Gen. Virol. 13:111-120.
14. Mountcastle, W. E., R. W. Compans, and P. W. Choppin. 1971.Proteins and glycoproteins of paramyxoviruses:a
comparison of simianvirus 5,Newcastle diseasevirus, and Sendai virus. J. Virol.7:47-52.
15. Peacock, A. C., and C. W. Dingman. 1968. Molecular weightestimationandseparation ofribonucleicacid by electrophoresis in agarose-acrylamyde composite gels. Biochemistry 7:668-674.
16. Petermann, M. L. 1964. The physical and chemical properties ofribosomes. Elsevier, New York. 17. Petermann,M. L., and A. Pavlovec.1966.The subunits
and structural ribonucleic acids of Jensen sarcoma
ribosomes. Biochim. Biophys. Acta114:264-276. 18. Portner, A.,and D. W. Kingsbury.1970.Complementary
RNA'sinparamyxoviriens andparamyxovirus-infected cells. Nature (London) 228:1196-1197.
19. PridgenC., and D. W. Kingsbury.1972. Adenylate-rich
sequences in Sendai virus transcripts from infected cells. J. Virol. 10:314-317.
20. Robinson, W.S. 1970.Self-annealing ofsubgroup2
myx-ovirus RNAs.Nature(London) 225:944-945. 21. Schincariol,A.L., and A. F. Howatson.1972.Replication
ofvesicular stomatitis virus. II.Separation and charac-terization of virus-specific RNA species. Virology 49:766-783.
22. Wild, T. F. 1971. Replication of vesicular stomatitis virus: characterization of the virus-induced RNA. J. Gen.Virol. 13:295-310.