Identification of Novel Structural Determinants in MW965 Env
That Regulate the Neutralization Phenotype and
Conformational Masking Potential of Primary HIV-1 Isolates
Zakiya M. Qualls,
aAlok Choudhary,
aWilliam Honnen,
aRaja Prattipati,
aJames E. Robinson,
bAbraham Pinter
aaPublic Health Research Institute Center, New Jersey Medical School, Rutgers University, Newark, New Jersey,
USA
bDepartment of Pediatrics, Tulane University Medical Center, New Orleans, Louisiana, USA
ABSTRACT
The subtype C HIV-1 isolate MW965.26 is a highly neutralization-sensitive
tier 1a primary isolate that is widely used in vaccine studies, but the basis for the
sensitive neutralization phenotype of this isolate is not known. Substituting the
MW965.26 V1/V2 domain into a neutralization-sensitive SF162 Env clone resulted in
high resistance to standard anti-V3 monoclonal antibodies, demonstrating that this
region possesses strong masking activity in a standard Env backbone and indicating
that determinants elsewhere in MW965.26 Env are responsible for its unusual
neu-tralization sensitivity. Key determinants for this phenotype were mapped by
generat-ing chimeric Envs between MW965.26 Env and a typical resistant Env clone, the
con-sensus C (ConC) clone, and localized to two residues, Cys384 in the C3 domain and
Asn502 in the C5 domain. Substituting the sensitizing mutations Y384C and K502N
at these positions into several resistant primary Envs resulted in conversion to
neutralization-sensitive phenotypes, demonstrating the generalizability of this effect.
In contrast to the sensitizing effects of these substitutions on normally masked
epitopes, these mutations reduced the sensitivity of VRC01-like epitopes overlapping
the CD4-binding domain, while they had no effect on several other classes of
broadly neutralizing epitopes, including members of several lineages of V2-dependent
quaternary epitopes and representatives of N332 glycan-dependent epitopes (PGT121)
and quaternary, cleavage-dependent epitopes centered at the gp41-gp120 interface
on intact HIV-1 Env trimers (PGT151). These results identify novel substitutions in
gp120 that regulate the expression of alternative conformations of Env and
differen-tially affect the exposure of different classes of epitopes, thereby influencing the
neutralization phenotype of primary HIV-1 isolates.
IMPORTANCE
A better understanding of the mechanisms that determine the wide
range of neutralization sensitivity of circulating primary HIV-1 isolates would provide
important information about the natural structural and conformational diversity of
HIV-1 Env and how this affects the neutralization phenotype. A useful way of
study-ing this is to determine the molecular basis for the unusually high neutralization
sensitivities of the limited number of available tier 1a viruses. This study localized
the neutralization sensitivity of MW965.26, an extremely sensitive subtype C-derived
primary isolate, to two rare substitutions in the C3 and C5 domains and
demon-strated that the sequences at these positions differentially affect the presentation of
epitopes recognized by different classes of standard and conformation-dependent
broadly neutralizing antibodies. These results provide novel insight into how these
regions regulate the neutralization phenotype and provide tools for controlling the
Env conformation that could have applications both for structural studies and in
vaccine design.
KEYWORDS
BNAbs, Env, HIV-1, gp120, gp160, neutralization resistance
Received16 October 2017Accepted28
November 2017
Accepted manuscript posted online13
December 2017
CitationQualls ZM, Choudhary A, Honnen W,
Prattipati R, Robinson JE, Pinter A. 2018. Identification of novel structural determinants in MW965 Env that regulate the neutralization phenotype and conformational masking potential of primary HIV-1 isolates. J Virol 92:e01779-17.https://doi.org/10.1128/JVI .01779-17.
EditorWesley I. Sundquist, University of Utah
Copyright© 2018 American Society for
Microbiology.All Rights Reserved.
Address correspondence to Abraham Pinter, [email protected].
PATHOGENESIS AND IMMUNITY
crossm
on November 6, 2019 by guest
http://jvi.asm.org/
H
IV-1 Env proteins contain highly immunogenic epitopes that readily induce
anti-body responses upon either infection or vaccination (reviewed in reference 1).
These responses are represented by large numbers of monoclonal antibodies (MAbs)
isolated from vaccinated or infected subjects that are directed against standard
non-glycosylated epitopes in the V3 region (2–9), the V1/V2 domain (10–14), the
CD4-binding domain (15–18), and CD4-induced epitopes (16, 19–22). Such so-called
con-ventional antibodies possess a narrow neutralization breadth, limited mostly to a small
class of neutralization-sensitive tier 1a and tier 1b Envs, with little, if any, neutralization
activity against typical tier 2/3 primary isolates (8, 23–25).
More recent studies have documented the existence of multiple classes of broadly
neutralizing antibodies (BNAbs) that have been isolated from chronically infected
subjects (26–31). These BNAbs possess remarkable neutralization potencies and
breadths, extending to relatively neutralization-resistant tier 2 and tier 3 clinical isolates,
and passive antibody delivery studies have demonstrated that the presence of such
antibodies can effectively control viremia in several animal models (32–35). The
epitopes targeted by these responses are highly conformational and frequently include
components of one or more N-linked glycans. The BNAbs that target these sites often
possess extensive levels of somatic mutations compared to germ line sequences (26,
29, 36–38) or have unusually long CDR3 regions that contribute to their activity (26, 27,
31, 39, 40). Such antibodies are produced by only a fraction of infected patients, require
considerable time for development, and are not induced by vaccination (41–43).
For many primary HIV-1 isolates, the inability of readily inducible conventional
antibodies to neutralize viral infection is due to occlusion of the target epitopes in the
native Env trimer by conformational masking (44, 45) and by a continuously evolving
glycan shield (46). It has been shown for a number of primary Envs that the V1/V2
domain plays a critical role in such masking. Deletion of the V2 loop in resistant Envs
resulted in increased neutralization sensitivity to patient sera and standard MAbs
against conventional neutralization epitopes (47–50), and exchanging the V1/V2
do-main between the neutralization-sensitive SF162 Env and the neutralization-resistant
JR-FL Env resulted in the switching of neutralization phenotypes (8, 51). The high glycan
content of the V1/V2 domain contributes to this effect, since deletions of glycans in this
region in a number of Env clones significantly enhanced sensitivity to neutralization by
conventional antibodies (8, 52–54). The recent elucidation of the close association
between the V1/V2 and V3 domains at the apex of the native HIV-1 Env trimer (55, 56)
provides a possible mechanism for these effects, which presumably involve a
combi-nation of hydrophobic and electrostatic interactions between conserved residues in V2
and V3 that lead to steric shielding of sites in V3 by the bulk of peptide residues and
glycans in the V1 and V2 domain.
Despite the importance of V1/V2 masking as a determinant of resistance, this does
not always correlate with the neutralization phenotype, indicating that additional
mechanisms also contribute toward this effect. A good example of this is provided by
the unusual neutralization sensitivity of isolate MW965.26 (MW965), a primary clade C
virus isolated from the peripheral blood mononuclear cells (PBMCs) of a patient in
Malawi with a late-stage (Fiebig VI) primary infection (24). This Env was the most
sensitive to neutralization by polyclonal patient sera and selected MAbs in a panel of
over 100 molecularly cloned HIV-1 Env pseudoviruses representing a broad range of
genetic and geographic diversity (24) and was also highly sensitive to neutralization by
antibodies induced by vaccination with various V1/V2, V3, and gp160 immunogens (13,
57, 58). The V1/V2 domain of the MW965 Env is longer and more highly glycosylated
than that of many highly neutralization-resistant HIV-1 isolates, suggesting that the
MW965 V1/V2 sequence does not contribute to the neutralization-sensitive phenotype
of this Env and implicating an independent mechanism. We reasoned that elucidating
the molecular determinants responsible for the unusual neutralization sensitivity of this
isolate would increase our understanding of the variety of mechanisms that regulate
conformational masking and the neutralization phenotype in primary HIV-1 isolates and
improve our understanding of the correlation between Env sequence and function.
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
Through the creation of a series of consensus C (ConC)-MW965 chimeric Envs, novel
regulators of Env conformation and neutralization sensitivity that accounted for the
unusual phenotype of the MW965 Env were identified. These studies also identified Env
constructs that possessed intermediate structures and phenotypes that may be useful
for structural studies and perhaps serve as novel vaccine immunogens.
RESULTS
Inherent masking activity of the V1/V2 loop of neutralization-sensitive
MW965.26.
The V3 domain is a potentially sensitive neutralization domain that is
known to be highly sensitive to conformational masking by the V1/V2 region (8, 51).
The sensitivities of MW965 Env and a control clade C Env, ConC Env, to neutralization
by standard V3-specific antibodies were initially compared for four V3-specific MAbs
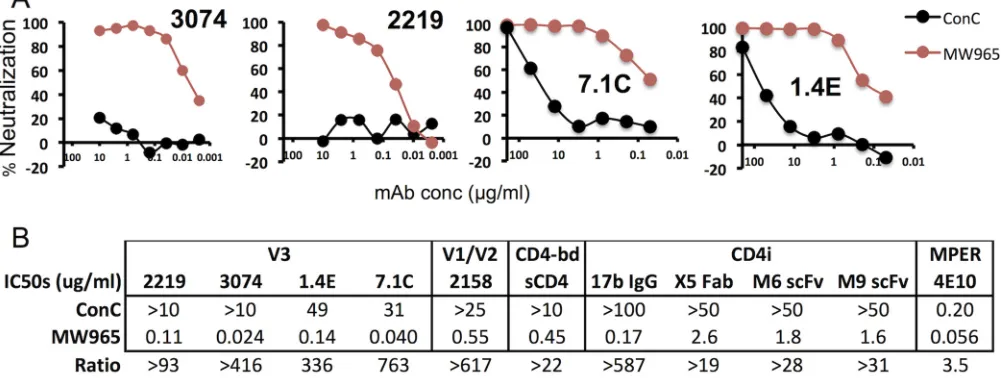
(19, 23, 59, 60), and representative neutralization curves are shown in Fig. 1A. The
neutralization sensitivity of the MW965 Env to all of these MAbs was much greater than
that of the ConC Env, with 50% inhibitory concentrations (IC
50s) ranging from 0.024 to
0.14
g/ml for MW965 Env versus
⬎
10 to 49
g/ml for ConC Env, a range of a
⬎
90- to
775-fold difference in neutralization sensitivity for the two Envs (Fig. 1B). Similar
neutralization patterns were observed for MAbs to other standard targets throughout
Env, including soluble CD4 (sCD4), V1/V2, and CD4-induced epitopes, and an anti-MPER
antibody (Fig. 1B). The V3 sequences of ConC and MW965 Envs are highly similar, with
only a single difference at position 322 (A for MW965 Env and D for ConC Env) being
detected. Introducing these substitutions into the two Envs had no effect on their
neutralization sensitivities to anti-V3 antibodies (data not shown), showing that the
difference in the neutralization sensitivities of these viruses was due to differences in
the exposures of the target epitopes.
The hypervariable V1 and V2 regions of MW965 Env are longer and more highly
glycosylated than those of the resistant ConC Env, with a total of nine and seven
N-linked glycans, respectively (Fig. 2A). In comparison, the V1/V2 sequence of SF162,
another widely used tier 1a isolate, contains only four N-linked glycans, and this
hypoglycosylation is known to correlate with the reduced masking activity of the SF162
V1/V2 domain and the high neutralization sensitivity of this isolate (51, 52, 61). In order
to directly examine the masking potential of the MW965 V1/V2 sequence, we examined
the effect of substituting this sequence into SF162-V3
ConB, a highly
neutralization-sensitive variant of SF162. This construct was used for neutralization experiments
because the introduction of the consensus B (ConB) V3 sequence into SF162 Env further
FIG 1Comparison of sensitivities of MW965.26 and ConC Envs to neutralization by anti-V3 MAbs. (A) Representative neutralization curves. (B) Summary of IC50s
(in micrograms per milliliter) of MW965 and ConC neutralization by four anti-V3 MAbs, V1/V2 MAb 2158, CD4-binding domain (CD4-bd) ligand, sCD4, and four CD4-induced (CD4i) MAbs or antibody fragments. The IC50s reported are averages from at least three independent experiments.
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:3.585.43.544.80.269.2]increased the neutralization sensitivity of this isolate, by reducing a modest effect of
V3-specific masking inherent to the native SF162 sequence (8, 62). As shown in Fig. 2B
and C, both MW965 and SF162-V3
ConBwere highly sensitive to neutralization by
four anti-V3 MAbs, with IC
50s in the low nanogram-per-milliliter range. Substituting the
MW965 V1/V2 sequence converted the SF162-V3
ConBEnv into a highly
neutralization-resistant Env with IC
50s ranging from
⬎
10
g/ml for MAbs 3074 and 2219 to 46
g/ml
for MAb 7.1C and 200
g/ml for MAb 1.4E, corresponding to
⬎
5,600- to
⬎
21,000-fold
increases in resistance to neutralization by the last two anti-V3 MAbs relative to that of
the SF162 parent (Fig. 2C). In addition, exchanging the V1/V2 sequence of MW965 into
the ConC Env backbone resulted in a neutralization-resistant phenotype similar to that
of wild-type (WT) ConC (data not show). These results showed that, consistent with its
sequence and predicted properties, the V1/V2 domain of MW965 possessed potent
masking activity for V3 epitopes in a standard Env backbone and suggested that
determinants elsewhere in the MW965 sequence overcame this masking activity and
were responsible for the high neutralization sensitivity of the MW965 isolate.
Sensitizing determinants of the MW965.26 Env were localized in the C3 and C5
domains of gp120.
To identify regions in the MW965 Env responsible for its unusual
phenotype, a series of chimeric sequences between the MW965 and ConC Env
se-quences was created (Fig. 3A). Representative neutralization curves are shown for
V3-specific MAb 7.1C in Fig. 3B, and IC
50s for 2 anti-V3 MAbs are tabulated in Fig. 3C.
The MW965 chimera containing the ConC C1-V3 region (construct 1) had a sensitivity
to neutralization by the anti-V3 MAbs similar to that of wild-type MW965, confirming
the lack of a role for either the V1/V2 or V3 sequence on sensitivity and ruling out the
possibility of contributions from other positions in the N-terminal region of the MW965
gp120 toward this neutralization phenotype. Further introduction of the ConC C3 and
V4 domains [MW965(ConC C1-V4) (construct 2)] resulted in
⬃
3- to 11-fold increased
resistance over that of the parental MW965, while introduction of the ConC C4-C5
domains into chimera 1 [MW965(ConC C1-V3/C4-C5) (construct 3)] resulted in
⬎
150- to
400-fold increased resistance. Substituting the ConC gp41 domain [MW965(ConC
C1-V3, gp41) (construct 4)] into chimera 1 had no significant effect on neutralization,
ruling out a significant effect of the transmembrane (TM) sequence on the
neutraliza-FIG 2Analysis of the V1/V2 masking activity of MW965.26 in an SF162 backbone. (A) V1/V2 amino acid alignments of ConC, MW965, and SF162, with N-linked glycosylation sites being underlined and highlighted in bold. Also highlighted are residues at positions 167 and 169 in the SF162 sequence that contribute to neutralization sensitivity and potential N-linked glycosylation sites (PNLGs). (B) Representative neutralization curves comparing the V3 sensitivity of WT MW965 and SF162 to that of chimeric SF162 Env with MW965 V1/V2. (C) Schematic of the Env chimeras used in this experiment and summary of the IC50s for four anti-V3
MAbs. The IC50s reported are averages from at least three independent experiments.
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:4.585.54.537.68.297.2]tion phenotype. These results demonstrate the presence of significant neutralization
resistance determinants in both the C3-V4 and C4-C5 regions of Env but not in other
Env domains.
In order to further localize neutralization determinants, smaller regions of gp120
were exchanged between the MW965 and ConC sequences (Fig. 4). Whereas
introduc-ing the individual V4, C4, and V5 domains of ConC into MW965 Env had minimal effects
on neutralization sensitivity to three V3-specific antibodies, substituting the ConC C3
resulted in a
⬎
2- to 5-fold increase in IC
50s, while substituting the C5 domain
resulted in
⬎
50- to 140-fold increases in resistance for these MAbs (Fig. 4A and B).
Consistent results were obtained for the reverse chimeras, with ConC(C3
MW965) and
ConC(C5
MW965) becoming
⬎
5- to 26-fold and 10- to
⬎
300-fold more sensitive to
neutralization by these anti-V3 MAbs, respectively, than the ConC parent (Fig. 4C).
While neither of the MW965 single-domain chimeras was as resistant to neutralization
as the parental ConC Env, the combined substitution of the ConC C3 and C5 domains
[MW965(ConC C3
⫹
C5)] resulted in a resistant phenotype similar to that of the ConC
parent (Fig. 4A and D).
Further localization of the C3 neutralization determinant to a novel Cys at
position 384 at the C terminus of the C3 domain.
The C3 region exchanged between
the Envs comprised 52 amino acids localized between C331 at the C terminus of the V3
domain and C385 at the N terminus of the V4 domain and contained seven
substitu-tions between the MW965 and ConC sequences (Fig. 5A). The MW965 substitusubstitu-tions
were introduced as single or double mutations into the ConC sequence, and
neutral-ization data and neutralneutral-ization curves for representative anti-V3 MAbs are shown in Fig.
5A and B, respectively. ConC mutants E335T/D336I, R344G/S346E, and K360E/E362K
were all similarly resistant to neutralization by anti-V3 MAbs as the parental ConC.
However, introducing the single Y384C mutation at the final position in this sequence
into ConC Env resulted in increased sensitivities to neutralization of
⬎
59-, 34-, and
9-fold by anti-V3 MAbs 2219, 7.1C, and 1.4E, respectively.
In order to test whether this effect was specific to the introduction of cysteine rather
than to the loss of the highly conserved tyrosine normally present at this position, two
additional mutant ConC Envs were constructed in which Y384 was replaced with either
a serine or an alanine, which are similar in size to cysteine. If the effect was due to the
FIG 3Initial mapping of determinants influencing the masking potential of MW965.26 by comparing the neutralization sensitivity of Env chimeras. (A) Schematic representation of chimeric MW965.26/ConC Envs, numbered 1 to 4, used in this experiment; (B) representative neutralization curves for the 7.1C MAb; (C) summary of IC50neutralization endpoints for two representative V3-specific MAbs. The IC50s reported are averages from at
least three independent experiments.
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:5.585.41.501.71.282.2]loss of tyrosine, these would also be expected to result in increased neutralization
sensitivity. Both ConC Y384S and ConC Y384A remained resistant to neutralization (Fig.
5A and B, right), indicating that the effect was specific to the presence of cysteine at
position 384.
Localizing the neutralization determinants in C5 to position 502 near the end
of the C5 domain.
The fragment swaps described in Fig. 3 and 4 demonstrated that the
major MW965 sensitizing determinant is located in the C5 domain. This region of gp120
is relatively conserved between these two sequences, with only three differences
between the ConC and MW965 sequences, R476K, K500G, and K502N, being detected
(Fig. 6A). The R476K substitution is a conservative mutation and was present in
⬃
7.5%
of the sequences in the database (4,183 sequences were analyzed;
http://www.hiv.lanl
.gov/
), suggesting that this is not a key factor. The other substitutions involved changes
in charge; the K500G substitution is rare in primary isolates, occurring in only 4% of Env
sequences, while the asparagine at position 502 is unique to MW965. These two
mutations were therefore introduced into the MW965 sequence, and their effect on
neutralization sensitivity was determined. The G500K mutation had a minimal effect on
neutralization sensitivity, while the N502K mutation resulted in a significant (
⬎
90- to
⬎
250-fold) increase in neutralization resistance to anti-V3 MAbs 2219, 1.4E, and 7.1C
(Fig. 6A and B).
To further explore the unmasking potential of 502N, the reciprocal K502N mutation
was introduced into the ConC Env (Fig. 6C). The mutant with this Env was
⬎
33- to
128-fold more sensitive to neutralization than parental ConC by the anti-V3 MAbs
FIG 4Residues influencing the masking potential of the MW965.26 V1/V2 map to the C3 and C5 domains of gp120. (A) Schematic representation of chimeric Envs, and summary of IC50s obtained for three anti-V3 MAbs (the IC50s reported are averages from three independent experiments); (B to D) representative
curves of MAb 1.4E neutralization of reciprocal single-domain MW965-ConC chimeras (B, C) and the C3 and C5 double-domain chimera (D).
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:6.585.40.540.67.422.2]tested. Significant increases in neutralization sensitivity were also observed when the
K502N mutation was introduced into two additional tier 2/3 primary isolates; a
⬎
33 to
⬎
250-fold increase in neutralization sensitivity was observed in clade C isolate 53M
(Fig. 6D), and a
⬎
7- to 50-fold increase in sensitivity was observed in clade A isolate
BG505 (Fig. 6E). Substituting the 502 mutation into clade B isolates AC10.0 and RHPA
yielded Envs that were not infectious (data not shown).
Lysine at position 502 is present in 82% of the sequences in the database (Fig. 7A),
and the N502K substitution had the strongest masking effect on MW965 Env (350-fold
for MAb 1.4E). Two other basic residues are the most common natural substitutions at
this position (Arg, 15%; Gln, 1.2%), and these had smaller masking effects (
⬃
25-fold for
Arg and
⬃
8.5-fold for Gln), while rarer neutral or acidic substitutions, such as alanine
(0.3% of isolates), threonine (0.2% of isolates), and glutamic acid (0.05% of isolates),
resulted in smaller decreases or increases in neutralization sensitivities (as much as a
7.7-fold increase in neutralization sensitivity for Env 502E) compared to the
neutraliza-tion sensitivity of parental MW965 Env (Fig. 7B and C). These results indicate that a
strongly basic amino acid at position 502 is needed to maintain the
neutralization-resistant phenotype, while substitution of neutral or acidic residues increases
neutral-ization sensitivity, even in the presence of a masking V1/V2 domain.
Combination of sequences at positions 384 and 502 replicates the parental
phenotypes.
While the effect of individual mutations at positions 384 and 502 of
gp120 affected the neutralization phenotypes of the ConC and MW965 Envs, the
combination of the two mutations exerted a stronger effect than the individual
mutations, often achieving the full level of sensitivity or resistance exhibited by the
parental Env sequences (Fig. 8). In addition to affecting the sensitivity of epitopes in the
V3 domain, similar reciprocal effects were also seen for sCD4 and for MAbs against
standard V1/V2 (e.g., MAb 2158) and CD4-induced (e.g., MAb 17b) epitopes and, to a
lesser extent, for an MPER-specific MAb (4E10) as well. This suggests that the
combi-nation of residues at these two positions strongly influences the expression of alternate
conformations of the native Env trimer that differ in the level of exposure of multiple
regions bearing potential neutralization epitopes normally occluded in common Env
structures.
FIG 5Y384C accounts for the contribution of the C3 domain to the increased neutralization sensitivity of MW965. (A) C3 sequences of the WT and mutant Envs constructed, with the table comparing the IC50s for three anti-V3 antibodies. The IC50s reported are averages from at least three independent experiments. (B)
Representative neutralization curves with V3-specific MAb 7.1C for ConC-MW965 C3 chimeric Envs (left) and for Env with a serine or alanine substitution at position 384 (right).
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.42.540.77.311.2]Effect of C3 and C5 mutations on sensitivity to broadly neutralizing MAbs.
In
contrast to the standard MAbs used in the studies described above, several families of
broadly neutralizing antibodies (BNAbs) that possess strong neutralizing activities
against many tier 2 and tier 3 isolates have been described. The effect of mutations at
positions 384 and 502 on sensitivity to a number of these BNAbs was tested. We first
studied the family of MAbs directed to glycan-dependent, quaternary epitopes
local-ized to the V1/V2 domain. These included the prototypical PG9 and PG16 MAbs (27),
PGT145 (39), and VRC26.08 and VRC26.03, two more recently described MAbs directed
against related epitopes isolated from an HIV-1 clade C-infected donor that possessed
potent neutralizing activities against many subtype C viruses (30, 31). The tier 2 ConC
isolate was strongly neutralized by all of these MAbs, while the parental MW965 was
less sensitive (Fig. 9A, top row, and B). In contrast to their effect on sensitivity to
standard MAbs, introduction of the MW965 C3 and C5 residues into the ConC sequence
did not appreciably affect the neutralization sensitivity to these BNAbs, while
introduc-tion of the ConC sequences at these posiintroduc-tions into the MW965 backbone had variable
effects for different members of this family. Moderate increases in sensitivity were seen
for PG9 and PG16 (IC
50increases of 19- and 12-fold, respectively), while a
⬍
2-fold effect
was seen for PGT145 (39), and no effect was seen for VRC26.08 and VRC26.03.
FIG 6Effect of substitutions at position 502 on neutralization sensitivity. (A) Amino acid sequences of the C5 point mutants constructed, with the table comparing the IC50s for three anti-V3 antibodies (the IC50s reported are averages from three independent experiments); (B)
neutralization curves for parental and mutant MW965 constructs with anti-V3 MAb 1.4E; (C) representative neutralization curves for parental and ConC K502 mutant Envs with anti-V3 MAb 1.4E; (D, E) neutralization curves for parental and K502N mutant Envs ZM53 (D) and BG505 (E) with anti-V3 MAb 1.4E.
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:8.585.43.506.70.457.2]Two BNAbs directed against different glycan-dependent epitopes (PGT121 and
PGT151) gave similar patterns as PG9/PG16; i.e., the C3 and C5 mutations had no effect
on sensitivity to neutralization in the ConC backbone, and modest increases in
sensi-tivity (
⬃
3 to 6-fold) were detected upon introduction of these mutations into the
MW965 backbone (Fig. 9A, bottom row, and B). PGT121 is a representative of a class of
broadly neutralizing antibodies that are dependent in part on the presence of a
high-mannose glycan at position 332 at the C-terminal base of the V3 loop (26), while
PGT151 recognizes an epitope comprised of amino acid residues and glycans at the
FIG 7Position 502 is an important regulator of V1/V2, masking the potential of the V3 domain in primary isolates. (A, B) Masking/unmasking effect of other common and rarer amino acids (AA) found at position 502 in circulating isolates. (C) Summary of IC50s for Envs with different substitutions at
position 502. The IC50s reported are averages from at least three independent experiments.
FIG 8Effects of combined substitutions at positions 384 and 502 on Env structure and monomeric epitope exposure. (A) Representative curves comparing the neutralization of Env mutants with double mutations at positions 384 and 502 with that of the WT Env by V3, sCD4, CD4i, and V1/V2 antibodies; (B) summary of IC50s, reported as averages from at least three independent experiments.
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:9.585.41.525.83.269.2] [image:9.585.40.545.446.707.2]gp120/gp41 interface that is preferentially expressed on properly formed, cleaved
trimers (63).
A different pattern was obtained for members of another class of broadly
neutral-izing MAbs, represented by VRC01, VRC03, and AC135, directed against sites
overlap-ping the CD4-binding domain (Fig. 9A, bottom row, and B). Similar to the other BNAbs,
the parental ConC Env was more sensitive to neutralization by these MAbs than
MW965, and introducing the ConC C3 and C5 mutations into the MW965 backbone
strongly increased the sensitivity to neutralization by these MAbs, to the extent that the
mutant MW965 Env was as sensitive as the WT ConC Env. However, in contrast to the
effect seen for the other MAbs, introducing the MW965 mutations into the ConC
backbone significantly decreased the neutralization sensitivity to these antibodies. The
effect of the individual mutations on the neutralization phenotype and epitope
expo-sure varied for each BNAb, but in the majority of the cases the 502 position had the
greatest effect on the observed neutralization phenotype (Fig. 9B).
These varied neutralization patterns suggested that mutations at these positions
stabilized alternative conformations of Env that differentially affected the formation
and exposure of these different classes of epitopes on the virion surface. To directly
measure the effect of these mutations on epitope exposure, WT or double mutant
trimeric Envs were expressed on the surface of 293T cells, fixed after being bound by
the MAb of interest, and analyzed by flow cytometry (Fig. 10A and B). The binding
patterns correlated well with the neutralization data for all four MAbs tested. For
example, anti-V3 MAb 7.1C bound better to MW965 than to ConC, while the opposite
pattern was seen for VRC03, and the double mutation reversed these patterns.
Contribution of the C3 and C5 regions of MW965 to viral infectivity.
The studies
described above defined novel mutations in the C3 and C5 domains that modify the
neutralization phenotype of primary Envs. The MW965 phenotype is rare in primary
FIG 9Effects of positions 384 and 502 on exposure of quaternary epitopes. (A) Representative curves comparing the neutralization of Env mutants with double mutations at positions 384 and 502 with that of the WT Env by QNE glycan-dependent BNAbs; (B) summary of IC50s, reported as averages from at least three
independent experiments.
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:10.585.46.541.71.372.2]isolates, and the C384 and N502 mutations responsible for this phenotype were not
found in any other isolates of an extensive Env sequence database. One possible reason
for this is that the infectivity of such viruses would be inhibited by the presence of
readily induced antibodies to the exposed targets in the V3 and CD4-binding domains,
limiting viral replication within hosts and spread to other hosts. Another possibility is
that the neutralization-sensitive phenotype correlated with a reduction in replicative
activity, which would inhibit the spread of these sequences, independent of an
acquired immune response. This possibility was suggested by the observation that the
MW965 wild-type Env was reproducibly inefficient in viral entry assays, compared to the
efficient entry of neutralization-resistant Envs. This effect was further explored by
quantitating the relative efficiency of the various phenotype-switched mutant forms in
both the MW965 and ConC backbones.
Averaged entry activities are quantitated in Fig. 11A for normalized viral inputs
(based on the p24 content) for the parental and mutant Envs. Multiple preparations of
viral pseudotypes prepared with the wild-type ConC possessed robust activity in entry
assays (
⬎
3
⫻
10
6relative light units [RLUs]), whereas the wild-type MW965
pseu-dotypes routinely possessed poor entry activity (on the order of 50,000 to 100,000
RLUs). Introducing the ConC C3 and C5 double mutations (C384Y/N502K) into the
MW965 Env resulted in large increases in entry efficiency that were close to WT ConC
levels, while introducing the key MW965 point mutations (Y384C/K502N) into ConC Env
resulted in significant decreases in entry efficiency. The single mutations had
interme-diate activities, with a greater effect being seen for the C5 (position 502)
substitu-FIG 10Effects of positions 384 and 502 on antibody binding to cell surface Env complexes. (A) Representative curves comparing the binding affinity of neutralizing antibodies to Env mutants with double mutations at positions 384 and 502 with that of the WT Env; (B) summary of half-maximal binding concentrations in comparison to the neutralizing IC50s, reported as averages from at least two and three independent experiments, respectively.
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:11.585.47.549.73.418.2]tions than for the C3 (position 384) substitutions. These results indicate that an
additional reason for the rarity of the tier 1 phenotype typified by MW965 is that the
mutations responsible for the increased neutralization sensitivity resulted in a
substan-tial reduction in replicative fitness, and the correlation between the effect of specific
mutations on both entry efficiency and neutralization resistance suggested a
mecha-nistic relationship between these factors.
To further explore the mechanism underlying entry efficiency differences, gp160
processing was studied by pulse-chase experiments conducted in transfected 293T cells
labeled with
35S (Fig. 11B). Analysis of gp160 decay indicated that WT MW965 Env was
defective in proteolytic processing compared to ConC Env and that these effects were
reversed by introduction of the C3/C5 mutations. These results suggest that the MW965
C3 and C5 mutations contribute to inefficient processing of MW965 Pr160, which in
turn may contribute to a reduction in the number of surface spikes on virions and may
be a factor in the reduced entry efficiency of this virus.
DISCUSSION
The goal of this study was to determine the structural and mechanistic basis for the
unusually high neutralization sensitivity of the tier 1a MW965.26 clade C HIV-1 isolate
to polyclonal sera from HIV-infected subjects and conventional, easily induced MAbs
directed to sites in V3 and other highly immunogenic domains (14, 24). These
exper-FIG 11Effects of positions 384 and 502 on viral infectivity and Env processing. (A) Comparison of viral infectivity, measured by RLU values. The p24 concentration was determined through ELISA, and a standardized amount of viral particles was used to infect U87-T4-CCR5 cells. Data represent averages from three independent infectivity assays. (B) Pulse-chase analysis comparing the Env (gp160) processing of the WT and mutant Envs in transfected 293T cells. Band intensity was quantified using ImageJ software.
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:12.585.43.504.71.435.2]iments revealed that the neutralization-sensitive phenotype of MW965 was expressed
despite the presence of a large and heavily glycosylated V1/V2 domain, which induced
a high level of neutralization resistance when introduced into a standard Env backbone.
This unusual phenotype was determined by a combination of two rare substitutions at
conserved positions in the C3 and C5 domains, Y384C and K502N, which altered the
conformation of the native Env complex, accounting for the lack of masking by the
V1/V2 domain and the consequent exposure of neutralization epitopes in domains that
are occluded in typical isolates. A similar effect on neutralization sensitivity was seen
when these two mutations were inserted into several other primary Envs, arguing for
the generality of these effects. These mutations had different effects on sensitivity to
several classes of broadly neutralizing antibodies that targeted potent neutralization
epitopes in the V1/V2 and CD4-binding domains. Thus, in addition to defining the basis
for the unusual neutralization phenotype of MW965, this study provides insight into
the role of different conformational forms on the exposure of classes of epitopes
targeted by rarer classes of broadly neutralizing antibodies that are formed in
response to chronic infection (64) and provides potential tools for manipulating these
conformations.
Little is known about the roles of the C3 and C5 positions identified in this study in
regard to Env function and the regulation of susceptibility to neutralization.
Replace-ment of Y384 with cysteine resulted in significant increases in the sensitivity of ConC to
standard anti-V3 MAbs (Fig. 4). Previous studies have shown that there is a strong
subtype-specific diversifying selection on sites in the
␣
2-helix present in the N-terminal
region of the C3 domain of clade C isolates (65), and unique mutations within this
region of C3 have been linked with resistance to neutralization by donor plasma,
suggesting the presence of type-specific neutralization targets in this region (66). C384
is located at the C terminus of the C3 region and distant from this region, suggesting
that a direct effect on the
␣
2-helix is not likely. In the BG505 SOSIP crystal structure,
position 384 is buried deep within the hydrophobic core of C3 that helps stabilize parts
of the CD4-binding site (55, 56, 67). Disruption of this core with a cysteine may alter the
stability of this domain. This residue is also adjacent to the highly conserved cysteine
at residue 385, which forms a disulfide linkage to C418 that delineates the C4 and V4
domains and has been shown to be essential for efficient folding and secretion of Env
and formation of functional molecules (68). This disulfide bond links two antiparallel
beta-strands in a six-stranded beta-barrel that is a prominent feature of the outer
domain, and the motion of this region is coupled with that of both the
␣
2-helix and the
bridging sheet during the conformational rearrangement induced by CD4 binding (69).
It is possible that the introduction of a second cysteine at position 384 may interfere
with cysteine pairing of the proximal C385, thereby altering the conformation of the
outer domain and changing how this region interacts with other Env regions during
infection.
Residue N502 played the major role in the neutralizing sensitivity of MW965 and
conformational masking of ConC. Most viral isolates bear a basic residue at this position
(82% have K, 15% have R, 1.2% have Q), with N502 being unique to the MW965
sequence (
http://www.hiv.lanl.gov/
). Substituting basic residues at this position in
MW965 reduced the sensitivity to neutralization, with the greatest effect being seen for
K, followed by R and then Q. This position is part of a basic KAKR motif common at
positions 500 to 503 that has been reported to influence proteolytic processing at the
adjacent furin cleavage signal (KEKR) at positions 508 to 511 (70, 71).
Independently of its role in proteolytic processing, the C5 region is known to form
essential interactions with gp41, which lead to the assembly of the native Env trimer
and contribute to membrane fusion and viral entry (71, 72). Formation of the functional
gp120-gp41 complex involves interactions between the highly conserved C1 and C5
domains of gp120 and the disulfide-bonded region (DSR) of gp41 (residues 598 to 604
in Env). Substitutions in both the C1 (residues 36 to 45) and C5 (residues 491 to 501)
domains resulted in dissociation of the envelope glycoprotein subunits (73), as did
mutations within and surrounding the DSR in gp41 (74, 75), and position 502 has been
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
reported to be present in a helical structure that directly interacts with gp41 (76).
Consistent with an association between this C5 region and the gp41 DSR, Cys residues
introduced at position 501 in the C5 domain and position 605 immediately C terminal
to the DSR in gp41 resulted in the formation of a novel disulfide bond that stabilized
the trimeric gp120-gp41 complex (77). Additional modifications of this mutant resulted
in the BG505 SOSIP complex (78) that was used to model the native Env trimer and to
obtain crystallographic and cryo-electron microscopy (cryo-EM) structures of the
cleaved HIV-1 Env trimer (55, 56, 67). Although the natural interactions and plasticity of
position 502 cannot be determined from the SOSIP structure because of the close
proximity of this position to the engineered stabilizing disulfide bond introduced at
position 501, its location does, however, suggest that the K502N substitution in MW965
weakens the interaction with gp41, resulting in the conformational changes that
contribute to the neutralization phenotype.
Recent data suggest that the native Env trimer exists in multiple conformational
forms, ranging from an unliganded highly compact closed trimer that is the dominant
form in neutralization-resistant strains to an open form which is dominant after
receptor binding (reviewed in reference 64). In the closed form, the V3 domain is
complexed with the base of the V2 domain (79, 80), and quaternary glycan-dependent
epitopes recognized by broadly neutralizing antibodies are efficiently expressed in this
structure, while highly immunogenic targets present in the V3, CD4-induced, and
CD4-binding domains are effectively occluded by the bulk of the heavily glycosylated
V1/V2 region. The most straightforward hypothesis for the results described in this
study is that the rare mutations present in the C3 and C5 domains of MW965 interfere
with the stability of the closed conformation and shift the equilibrium toward the more
open conformation that exposes the highly immunogenic targets, accounting for the
neutralization-sensitive phenotype.
The mechanism responsible for the neutralization sensitivity of MW965 is distinct
from that utilized by SF162, another widely used tier 1 isolate. SF162 is extremely
sensitive to neutralization by a family of MAbs that recognize an SF162 type-specific
quaternary epitope that bridges the V2 and V3 domains (81–84), demonstrating that
this Env exists predominantly in the closed conformation. The neutralization-sensitive
phenotype of SF162 is, however, mediated by structural features of its V1/V2 domain,
including hypoglycosylation and the presence of substitutions at several conserved
positions in V2 that result in a reduced ability to mask the adjacent V3 domain and
other highly immunogenic epitopes (82, 84). Consistent with this model, replacing the
V1/V2 domain of SF162 with that of JR-FL, a neutralization-resistant isolate with a more
highly glycosylated V1/V2 (51) or introducing additional glycosylation sites into the
SF162 V1/V2 domain results in the neutralization-resistant phenotype (52, 61), while
introduction of the SF162 V1/V2 domains into JR-FL Env (51, 84) or the removal of
glycosylation sites from the V1/V2 domain of YU2, another typical tier 2 isolate (8),
results in neutralization-sensitive phenotypes. The V1/V2 domain of MW965 is heavily
glycosylated and otherwise similar to that of typical neutralization-resistant isolates,
and transplantation of this domain into the SF162 backbone induced a highly resistant
neutralization phenotype (Fig. 2), supporting the conclusion that the MW965 Env exists
predominantly in an open conformation that precludes masking by the V1/V2 domain.
Particularly interesting was the varied effects of these mutations on the sensitivity
of different classes of epitopes to neutralization (Fig. 8 and 9). Inserting the ConC
sequences into MW965 Env resulted in three distinct patterns for different classes of
antibodies: increased resistance to conventional antibodies, no effect on members of
the VRC26 lineage of V2-dependent BNAbs, and increased sensitivity to V2-dependent
BNAbs represented by PG9 and PG16 and to the VRC01-like class of antibodies.
Inserting the MW965-specific mutations into the ConC Env also resulted in three
different patterns: the exposure of conventional epitopes in V3 and other normally
masked domains in ConC Env was greatly increased, and the exposure of the VRC01-like
epitopes overlapping the CD4-binding domain was reduced, while the mutant retained
full sensitivity to BNAbs directed against different classes of quaternary epitopes,
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
including members of several lineages of quaternary V2-dependent epitopes,
repre-sentatives of the N332 glycan-dependent epitopes (PGT121), and quaternary
cleavage-dependent epitopes centered at the gp41-gp120 interface on intact HIV-1 Env trimers
(PGT151).
A key question is how to explain the concurrent sensitivity of the mutant ConC Env
both to standard anti-V3 MAbs which target the open conformation and to
quaternary-dependent antibodies which target the closed conformation of Env. One possibility is
that these mutations introduce an intermediate conformation that retains the V2-V3
complex at the apex of the trimer required for expression of quaternary epitopes while
allowing the exposure of other regions of the V3 domain that are recognized by the
standard MAbs. A recent cryo-EM study identified an intermediate structure of the
sCD4-liganded BG505 SOSIP trimer that was stabilized by binding of Fabs derived from
MAb 17b, which recognizes a CD4-induced epitope, and MAb 8ANC195, which spans a
site on the gp120-gp41 interface (67). While this structure was intermediate between
the previously described open and closed forms, it required an
⬃
40-Å displacement of
the V1/V2 domain from its position in the unliganded structure (56, 80), and it is not
obvious that this structure could express the quaternary epitopes.
An alternate hypothesis is that an equilibrium exists between the different Env
conformations that normally favors the closed structure and that the MW965-specific
residues change that equilibrium toward the open structure. A rapid equilibration
between these forms in the mutant ConC Env would allow both conformations to be
readily accessed by antibodies, accounting for sensitivity to both the standard and the
quaternary-dependent BNAbs. The equal sensitivity of the wild-type and mutant forms
of MW965 Env to the VRC26 pair of V2-dependent antibodies, in contrast to the greater
potency of PG9 and PG16 for the mutant Env, may reflect a difference in dependence
on sites within the V3 domain for recognition by these different sets of antibodies,
consistent with previous epitope mapping data for these classes of antibodies (30, 31,
85). The different effects of these mutations on epitopes in the V2/V3 apex versus the
VRC01 class of epitopes in the CD4-binding domain could indicate different
equilibra-tion rates for these domains and reflect the conformaequilibra-tional complexity of Env.
This model would also be consistent with previous literature documenting the
reversal of the phenotype of neutralization-resistant tier 2/3 viruses by the introduction
of various point mutations throughout Env. In the older literature this involved either
changes in the masking potency of the V1/V2 domain through glycan mutation or
conformational rearrangement or included mutations in the TM subunit (86–89). The
mutations described in this study are unique in that they induce similar phenotypes in
multiple Envs with varied origins and sequences, do not involve modification of
glycosylation sites or sites in gp41, and have a novel pattern of effects on various
epitopes compared to those described in other previously published studies (86–89).
In terms of viral infectivity, the mutants/chimeras that resulted in a more open
conformation of Env were defective in infecting target cells compared to the infectivity
of their closed Env counterparts. A possible explanation of this observation is that
intracellular transport and precursor processing are less efficient for the unmasked
open Envs than for the closed Env forms (Fig. 11C), and the lower viral infectivity could
simply be the result of fewer functional Env trimers on the surface of unmasked/open
virions. This effect is probably independent of the effect of these mutations on the
neutralization phenotype, since introducing the G500K mutation into MW965 greatly
increased the infectivity of this Env, while having no effect on the neutralization
phenotype (data not shown).
The rarity of the two mutations responsible for the MW965 phenotype suggests that
whereas this virus may be useful for characterizing the qualitative and quantitative
nature of the humoral responses to specific immunogens, the neutralization pattern
observed does not reflect that expected for standard viral isolates. The insights gleaned
from this work may be of use in designing Env proteins for structural studies and may
be particularly helpful in the design of immunogens for induction of specific antibody
responses. For example, the increased conformational mobility of the ConC mutant
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
described in this study and its ability to efficiently coexpress both standard V3 epitopes
and quaternary epitopes may positively impact the immunogenicity of vaccines that
incorporate these structural features. Finally, insights gained from this work may also
aid immunotherapeutic clinical trials, since understanding the effect of different Env
conformational states on the exposure of different classes of epitopes will help
deter-mine the best antibody combinations needed to prevent neutralization escape.
MATERIALS AND METHODS
Generation of chimeric and mutant Envs.The MW965.26 and codon-optimized consensus C HIV-1 Envs were obtained from the NIH AIDS Reagent Program and expressed in the pcDNA3.1 mammalian expression vector (Thermo Fisher Scientific); for the MW965.26 Env,revwas coexpressed in the plasmid. Point mutations were introduced into the Envs by site-directed mutagenesis with a QuikChange site-directed mutagenesis kit (Stratagene, Inc.), and chimeric Envs were generated by blunt end ligation or Gibson assembly (New England BioLabs, Inc. [NEB]), which utilizes overlapping regions to ligate various DNA fragments into singular constructs. In some experiments, DNA fragments were generated by Phusion Hot Start High-Fidelity DNA polymerase (NEB) or synthesized as gBlock gene fragments (Integrated DNA Technologies, Inc. [IDT]) and exchanged into Env at specific restriction enzyme sites. Following transformation, colonies were screened by PCR for the desired sequence, and positive clones were confirmed by sequencing by Genewiz. All oligonucleotides were purchased from IDT.
Antibodies.Anti-V3 MAbs 2219 (23) and 3074 (25, 59) were provided by Susan Zolla-Pazner (NYUMC) or obtained from the NIH AIDS Reagent Program. Two additional broadly reactive V3 MAbs generated in the J. E. Robinson lab were included in this study (62). MAb 1.4E was isolated from a patient in the Gambia, where several CRF strains along with subtypes C and A are common (60), and Mab 7.1C was isolated from a subject in South Africa, where subtype C is dominant (19). The 1.4E antibody was isolated by antibody secretion of Epstein-Barr virus (EBV)-transformed B cell lines from patients, while 7.1C was isolated directly from cytokine-stimulated B cells in the absence of EBV transformation. The variable heavy (VH) and variable light (VL) chain regions were cloned into standard immunoglobulin expression
vectors, and the recombinant antibodies were expressed in the Expi293 expression system (Thermo Fisher Scientific) and purified by affinity chromatography on protein A columns (Thermo Fisher Scien-tific). Purified antibodies and reagents obtained from the NIH AIDS Reagent Program included sCD4 (contributed by Progenics Pharmaceuticals, Inc.) and anti-CD4-bs MAb 17b (contributed by James E. Robinson [90, 91]). The X5 Fab-, M6 scFv-, and M9 scFv-targeting CD4-induced epitopes were provided by Dimiter S. Dimitrov [17, 92]). Broadly neutralizing antibodies analyzed included PGT121, targeting the V3 loop glycan N332 (79, 93); PG9/PG16 (40, 94), PGT145 (39), and VRC26.03 and VRC26.08 (31), targeting quaternary epitopes centered in the V1/V2 domain; and VRC01, VRC03 (29), and AC135, recognizing epitopes overlapping the CD4-binding domain. These were all expressed as IgG1 isotypes from molecular clones in the Expi293 system and purified by protein A affinity chromatography.
The AC135 MAb was isolated from anin vitromemory B cell culture, using procedures described by Huang et al. (95). Briefly, non-B cells from the PBMCs were eliminated by negative selection with magnetic beads to which antibodies against the cell surface markers CD2, CD3, CD14, CD16, CD36, CD43, CD56, CD66b, and glycophorin A were attached and memory B cells were positively selected with magnetic beads conjugated with anti-CD27, a cell surface marker for memory B cells that is also expressed in low levels on plasma cells. The purified memory B cells were cocultured in 96-well plates with the MS40 L-low cell line (96) in 10% Alpha minimal essential medium supplemented with Toll-like receptor 9 agonist type B CpG oligonucleotide ODN 2006 (1M), interleukin-2 (IL-2; 10 ng/ml), IL-4 (2 ng/ml), IL-10 (100 ng/ml), and IL-21 (100 ng/ml), and cell supernatants were screened for binding activity against YU2 gp120, YU2 gp140, and a YU2 gp120 core fragment that lacked the V1, V2, and V3 variable regions and that was truncated at the N- and C-terminal regions, which was previously shown to bind anti-CD4-binding site (CD4bs) antibodies (97, 98). The B cells from the triply positive wells were lysed by adding 50l of cDNA lysis buffer (10⫻reverse transcription [RT] buffer (SuperScript III CellsDirect cDNA Synthesis System; Invitrogen), 400 ng random hexamer, 10 mM deoxynucleoside triphosphates, 0.1 M dithiothreitol [DTT], RT Sup III, RNAout, and water) per well, and the genes encoding Ig VHand VLchains
were amplified by RT-PCR and nested PCR as described previously (99).
The specificity of the AC135 MAb for a VRC01-related epitope in the CD4-binding domain was confirmed by the homology of the VHand VLchain sequences with those of VRC01-like MAbs and by the
binding of this MAb to the RSC3 protein, a resurfaced version of the stabilized core of gp120 known to express VRC01-related epitopes but not to the RSC371ΔI variant, previously shown to have reduced recognition by most VRC01-related antibodies (29).
Infectivity and neutralization assays. Infectivity and neutralization activity was determined as previously described (100) with a single-cycle infectivity assay using virions generated from the Env-defective luciferase-expressing pNL4-3.Luc.R⫺E⫺genome (101) pseudotyped with the molecularly
cloned HIV Env of interest. Briefly, 293T cells were transfected with plasmids harboring the pNL4-3.Luc.R⫺E⫺core and Env genomes, and 48 to 72 h later pseudotyped virions were harvested from the
culture supernatants. To determine specific viral infectivity, the p24 concentrations of viral lysates were measured by enzyme-linked immunosorbent assay (ELISA), a standardized amount of p24 (30 ng) from each sample was added to U87-T4-CCR5 or TZM-bl target cells, and the cells were plated out in 96-well plates in the presence of DEAE-dextran (10g/ml). For neutralization assays, virions were incubated with serial dilutions of MAbs for 1 h at 37°C prior to incubation with target cells, and the luciferase activity in the cell lysates was measured at 72 to 96 h postinfection with a microtiter plate luminometer (Harta, Inc.),
Qualls et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
using assay reagents from Promega. The IC50s reported were determined by interpolation from
neutral-ization curves or through the Excel software growth function, which calculates a predicted exponential growth, and are averages from at least three independent assays. The IC50s for neutralization reported
were compared with the IC50/IC80values reported for various MAbs in the CATNAP database, which
contains a compilation of neutralization data from the published literature (http://hiv.lanl.gov/catnap) (102), and were found to correlate with the reported values.
Flow cytometric analysis of MAb binding affinity to trimeric Env.The ability of MAbs to bind to the trimeric form of Env on the surface of transfected 293T cells was examined by flow cytometry. WT MW965, WT ConC, or the 384/502 mutant Env was expressed in 293T cells using the TransIT-LT1 transfection reagent and protocol (Mirus Bio LLC, Madison, WI). Cells were refed after 24 h with fresh medium containing 10% serum and incubated for an additional 2 days. Cells were removed from the plates for analysis by rinsing with 2 mM EDTA–phosphate-buffered saline and then incubated with antibodies for 1 h at 4°C, washed, incubated with anti-human IgG conjugated to phycoerythrin (PE) for an additional 1 h at 4°C, and then washed and fixed in 2% paraformaldehyde. The mean fluorescence intensities (MFI) were then determined on an Accuri C6 flow cytometer using C Flow software (Accuri Cytometers Inc., Ann Arbor, MI).
Radiolabeling and pulse-chase experiments.293T cells were transfected (as described above) with a 1:1 mixture of Env and the pNL4-3.Luc.R⫺E⫺core, and after 24 h the cultures were subjected to
pulse-chase analysis as previously detailed (103). Briefly, cells were starved in cysteine- and methionine-free medium for 30 min and then pulse-labeled for 20 min with 50Ci/well ofL-[35S]cysteine (MP
Biomedicals) in a 6-well plate and chased for 0, 1, 3, 6, or 12 h by washing with medium containing cold cysteine-methionine. After the chase period, the cells were lysed and immunoprecipitated with selected, high-titer HIV-positive plasma, and labeled viral proteins were analyzed after reduction with 25 mM DTT by SDS-PAGE on 7.5% polyacrylamide gels. The gels were dried and exposed to a phosphor screen in a phosphorimager cassette and developed in a Typhoon FLA 7000 phosphorimager (GE Healthcare Life Sciences). Env and p24 band intensities were quantified using ImageJ software (104); the data shown are representative of those from three independent repeats of the pulse-chase analysis.
Accession number(s).GenBank accession numbers for the included antibodies areMG517525and MG517526for 7.1C,MG517523andMG517524for 1.4E, andMG733355andMG733356for AC135.
ACKNOWLEDGMENTS
We thank Dennis Burton, Susan Zolla-Pazner, Dimiter S. Dimitrov, and John Mascola,
who provided monoclonal antibodies used in this study.
These studies were supported by PHS grants P01-AI088610 and R01 AI102718 to
A.P., and Z.M.Q. was supported in part by a Sigma XI grant-in-aid for research.
We have no competing interests to declare.
REFERENCES
1. Mayr LM, Zolla-Pazner S. 2015. Antibodies targeting the envelope of HIV-1. Microbiol Spectr 3(1):AID-0025-2014. https://doi.org/10.1128/ microbiolspec.AID-0025-2014.
2. Jiang X, Burke V, Totrov M, Williams C, Cardozo T, Gorny MK, Zolla-Pazner S, Kong XP. 2010. Conserved structural elements in the V3 crown of HIV-1 gp120. Nat Struct Mol Biol 17:955–961.https://doi.org/ 10.1038/nsmb.1861.
3. Baillou A, Brand D, Denis F, M’Boup S, Chout R, Goudeau A, Barin F. 1993. High antigenic cross-reactivity of the V3 consensus sequences of HIV-1 gp120. AIDS Res Hum Retroviruses 9:1209 –1215.https://doi.org/ 10.1089/aid.1993.9.1209.
4. Davis KL, Gray ES, Moore PL, Decker JM, Salomon A, Montefiori DC, Graham BS, Keefer MC, Pinter A, Morris L, Hahn BH, Shaw GM. 2009. High titer HIV-1 V3-specific antibodies with broad reactivity but low neutralizing potency in acute infection and following vaccination. Vi-rology 387:414 – 426.https://doi.org/10.1016/j.virol.2009.02.022. 5. Krachmarov C, Pinter A, Honnen WJ, Gorny MK, Nyambi PN,
Zolla-Pazner S, Kayman SC. 2005. Antibodies that are cross-reactive for human immunodeficiency virus type 1 clade A and clade B V3 domains are common in patient sera from Cameroon, but their neutralization activity is usually restricted by epitope masking. J Virol 79:780 –790. https://doi.org/10.1128/JVI.79.2.780-790.2005.
6. LaRosa GJ, Davide JP, Weinhold K, Waterbury JA, Profy AT, Lewis JA, Langlois AJ, Dreesman GR, Boswell RN, Shadduck P, Bolognesi DP, Matthews TJ, Emini EA, Putney SD. 1990. Conserved sequence and structural elements in the HIV-1 principal neutralizing determinant. Science 249:932–935.https://doi.org/10.1126/science.2392685. 7. Spear GT, Takefman DM, Sharpe S, Ghassemi M, Zolla-Pazner S. 1994.
Antibodies to the HIV-1 V3 loop in serum from infected persons contribute a major proportion of immune effector functions including
complement activation, antibody binding, and neutralization. Virology 204:609 – 615.https://doi.org/10.1006/viro.1994.1575.
8. Krachmarov CP, Honnen WJ, Kayman SC, Gorny MK, Zolla-Pazner S, Pinter A. 2006. Factors determining the breadth and potency of neu-tralization by V3-specific human monoclonal antibodies derived from subjects infected with clade A or clade B strains of human immuno-deficiency virus type 1. J Virol 80:7127–7135.https://doi.org/10.1128/ JVI.02619-05.
9. Gorny MK, Revesz K, Williams C, Volsky B, Louder MK, Anyangwe CA, Krachmarov C, Kayman SC, Pinter A, Nadas A, Nyambi PN, Mascola JR, Zolla-Pazner S. 2004. The V3 loop is accessible on the surface of most human immunodeficiency virus type 1 primary isolates and serves as a neutralization epitope. J Virol 78:2394 –2404.https://doi.org/10.1128/ JVI.78.5.2394-2404.2004.
10. Moore JP, Sattentau QJ, Yoshiyama H, Thali M, Charles M, Sullivan N, Poon SW, Fung MS, Traincard F, Pinkus M, Robey G, Robinson JE, Ho DD, Sodroski J. 1993. Probing the structure of the V2 domain of human immunodeficiency virus type 1 surface glycoprotein gp120 with a panel of eight monoclonal antibodies: human immune response to the V1 and V2 domains. J Virol 67:6136 – 6151.
11. Shotton C, Arnold C, Sattentau Q, Sodroski J, McKeating JA. 1995. Identification and characterization of monoclonal antibodies specific for polymorphic antigenic determinants within the V2 region of the human immunodeficiency virus type 1 envelope glycoprotein. J Virol 69:222–230.
12. Wu Z, Kayman SC, Honnen W, Revesz K, Chen H, Vijh-Warrier S, Tilley SA, McKeating J, Shotton C, Pinter A. 1995. Characterization of neutral-ization epitopes in the V2 region of human immunodeficiency virus type 1 gp120: role of glycosylation in the correct folding of the V1/V2 domain. J Virol 69:2271–2278.
Novel Conformational Masking Determinants in HIV-1 Env Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
13. Gorny MK, Pan R, Williams C, Wang XH, Volsky B, O’Neal T, Spurrier B, Sampson JM, Li L, Seaman MS, Kong XP, Zolla-Pazner S. 2012. Func-tional and immunochemical cross-reactivity of V2-specific monoclonal antibodies from HIV-1-infected individuals. Virology 427:198 –207. https://doi.org/10.1016/j.virol.2012.02.003.
14. Li L, Wang XH, Williams C, Volsky B, Steczko O, Seaman MS, Luthra K, Nyambi P, Nadas A, Giudicelli V, Lefranc MP, Zolla-Pazner S, Gorny MK. 2015. A broad range of mutations in HIV-1 neutralizing human mono-clonal antibodies specific for V2, V3, and the CD4 binding site. Mol Immunol 66:364 –374.https://doi.org/10.1016/j.molimm.2015.04.011. 15. Pinter A, Honnen WJ, Racho ME, Tilley SA. 1993. A potent, neutralizing
human monoclonal antibody against a unique epitope overlapping the CD4-binding site of HIV-1 gp120 that is broadly conserved across North American and African virus isolates. AIDS Res Hum Retroviruses 9:985–996.https://doi.org/10.1089/aid.1993.9.985.
16. Sullivan N, Sun Y, Sattentau Q, Thali M, Wu D, Denisova G, Gershoni J, Robinson J, Moore J, Sodroski J. 1998. CD4-induced conformational changes in the human immunodeficiency virus type 1 gp120 glycoprotein: consequences for virus entry and neutralization. J Virol 72:4694 – 4703.
17. Zhang MY, Shu Y, Rudolph D, Prabakaran P, Labrijn AF, Zwick MB, Lal RB, Dimitrov DS. 2004. Improved breadth and potency of an HIV-1-neutralizing human single-chain antibody by random mutagenesis and sequential antigen panning. J Mol Biol 335:209 –219.https://doi.org/10 .1016/j.jmb.2003.09.055.
18. Chen L, Kwon YD, Zhou T, Wu X, O’Dell S, Cavacini L, Hessell AJ, Pancera M, Tang M, Xu L, Yang ZY, Zhang MY, Arthos J, Burton DR, Dimitrov DS, Nabel GJ, Posner MR, Sodroski J, Wyatt R, Mascola JR, Kwong PD. 2009. Structural basis of immune evasion at the site of CD4 attachment on HIV-1 gp120. Science 326:1123–1127. https://doi.org/10.1126/science .1175868.
19. Robinson JE, Elliott DH, Martin EA, Micken K, Rosenberg ES. 2005. High frequencies of antibody responses to CD4 induced epitopes in HIV infected patients started on HAART during acute infection. Hum Anti-bodies 14:115–121.
20. Xiang SH, Doka N, Choudhary RK, Sodroski J, Robinson JE. 2002. Characterization of CD4-induced epitopes on the HIV type 1 gp120 envelope glycoprotein recognized by neutralizing human monoclonal antibodies. AIDS Res Hum Retroviruses 18:1207–1217.https://doi.org/ 10.1089/08892220260387959.
21. DeVico AL. 2007. CD4-induced epitopes in the HIV envelope glyco-protein, gp120. Curr HIV Res 5:561–571. https://doi.org/10.2174/ 157016207782418560.
22. Dey AK, Burke B, Sun Y, Sirokman K, Nandi A, Hartog K, Lian Y, Geonnotti AR, Montefiori D, Franti M, Martin G, Carfi A, Kessler P, Martin L, Srivastava IK, Barnett SW. 2012. Elicitation of neutralizing antibodies directed against CD4-induced epitope(s) using a CD4 mimetic cross-linked to a HIV-1 envelope glycoprotein. PLoS One 7:e30233.https:// doi.org/10.1371/journal.pone.0030233.
23. Gorny MK, Williams C, Volsky B, Revesz K, Cohen S, Polonis VR, Honnen WJ, Kayman SC, Krachmarov C, Pinter A, Zolla-Pazner S. 2002. Human monoclonal antibodies specific for conformation-sensitive epitopes of V3 neutralize human immunodeficiency virus type 1 primary isolates from various clades. J Virol 76:9035–9045.https://doi.org/10.1128/JVI .76.18.9035-9045.2002.
24. Seaman MS, Janes H, Hawkins N, Grandpre LE, Devoy C, Giri A, Coffey RT, Harris L, Wood B, Daniels MG, Bhattacharya T, Lapedes A, Polonis VR, McCutchan FE, Gilbert PB, Self SG, Korber BT, Montefiori DC, Mascola JR. 2010. Tiered categorization of a diverse panel of HIV-1 Env pseudoviruses for assessment of neutralizing antibodies. J Virol 84: 1439 –1452.https://doi.org/10.1128/JVI.02108-09.
25. Davis KL, Gray ES, Moore PL, Decker JM, Salomon A, Montefiori DC, Graham BS, Keefer MC, Pinter A, Morris L, Hahn BH, Shaw GM. 2009. High titer HIV-1 V3-specific antibodies with broad reactivity but low neutralizing potency in acute infection and following vaccination. Vi-rology 387:414 – 426.https://doi.org/10.1016/j.virol.2009.02.022. 26. Walker LM, Huber M, Doores KJ, Falkowska E, Pejchal R, Julien JP, Wang
SK, Ramos A, Chan-Hui PY, Moyle M, Mitcham JL, Hammond PW, Olsen OA, Phung P, Fling S, Wong CH, Phogat S, Wrin T, Simek MD, Protocol G Principal Investigators, Koff WC, Wilson IA, Burton DR, Poignard P. 2011. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 477:466 – 470.https://doi.org/10.1038/nature10373. 27. Walker LM, Phogat SK, Chan-Hui PY, Wagner D, Phung P, Goss JL, Wrin T, Simek MD, Fling S, Mitcham JL, Lehrman JK, Priddy FH, Olsen OA,
Frey SM, Hammond PW, Kaminsky S, Zamb T, Moyle M, Koff WC, Poignard P, Burton DR. 2009. Broad and potent neutralizing antibodies from an African donor reveal a new HIV-1 vaccine target. Science 326:285–289.https://doi.org/10.1126/science.1178746.
28. Moore PL, Gray ES, Sheward D, Madiga M, Ranchobe N, Lai Z, Honnen WJ, Nonyane M, Tumba N, Hermanus T, Sibeko S, Mlisana K, Abdool Karim SS, Williamson C, Pinter A, Morris L. 2011. Potent and broad neutralization of HIV-1 subtype C by plasma antibodies targeting a quaternary epitope including residues in the V2 loop. J Virol 85: 3128 –3141.https://doi.org/10.1128/JVI.02658-10.
29. Wu X, Yang ZY, Li Y, Hogerkorp CM, Schief WR, Seaman MS, Zhou T, Schmidt SD, Wu L, Xu L, Longo NS, McKee K, O’Dell S, Louder MK, Wycuff DL, Feng Y, Nason M, Doria-Rose N, Connors M, Kwong PD, Roederer M, Wyatt RT, Nabel GJ, Mascola JR. 2010. Rational design of envelope iden-tifies broadly neutralizing human monoclonal antibodies to HIV-1. Science 329:856 – 861.https://doi.org/10.1126/science.1187659.
30. Doria-Rose NA, Bhiman JN, Roark RS, Schramm CA, Gorman J, Chuang GY, Pancera M, Cale EM, Ernandes MJ, Louder MK, Asokan M, Bailer RT, Druz A, Fraschilla IR, Garrett NJ, Jarosinski M, Lynch RM, McKee K, O’Dell S, Pegu A, Schmidt SD, Staupe RP, Sutton MS, Wang K, Wibmer CK, Haynes BF, Abdool-Karim S, Shapiro L, Kwong PD, Moore PL, Morris L, Mascola JR. 2015. New member of the V1V2-directed CAP256-VRC26 lineage that shows increased breadth and exceptional potency. J Virol 90:76 –91.https://doi.org/10.1128/JVI.01791-15.
31. Doria-Rose NA, Schramm CA, Gorman J, Moore PL, Bhiman JN, DeKosky BJ, Ernandes MJ, Georgiev IS, Kim HJ, Pancera M, Staupe RP, Altae-Tran HR, Bailer RT, Crooks ET, Cupo A, Druz A, Garrett NJ, Hoi KH, Kong R, Louder MK, Longo NS, McKee K, Nonyane M, O’Dell S, Roark RS, Rudicell RS, Schmidt SD, Sheward DJ, Soto C, Wibmer CK, Yang Y, Zhang Z, NISC Comparative Sequencing Program, Mullikin JC, Binley JM, Sanders RW, Wilson IA, Moore JP, Ward AB, Georgiou G, Williamson C, Abdool Karim SS, Morris L, Kwong PD, Shapiro L, Mascola JR. 2014. Developmental pathway for potent V1V2-directed HIV-neutralizing antibodies. Nature 509:55– 62.https://doi.org/10.1038/nature13036.
32. Balazs AB, Chen J, Hong CM, Rao DS, Yang L, Baltimore D. 2011. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 481:81– 84.https://doi.org/10.1038/nature10660.
33. Saunders KO, Wang L, Joyce MG, Yang ZY, Balazs AB, Cheng C, Ko SY, Kong WP, Rudicell RS, Georgiev IS, Duan L, Foulds KE, Donaldson M, Xu L, Schmidt SD, Todd JP, Baltimore D, Roederer M, Haase AT, Kwong PD, Rao SS, Mascola JR, Nabel GJ. 2015. Broadly neutralizing human immu-nodeficiency virus type 1 antibody gene transfer protects nonhuman primates from mucosal simian-human immunodeficiency virus infec-tion. J Virol 89:8334 – 8345.https://doi.org/10.1128/JVI.00908-15. 34. Liu MA. 2012. Gene-based passive antibody protection from HIV. Nat
Biotechnol 30:156 –157.https://doi.org/10.1038/nbt.2114.
35. Yang L, Wang P. 2014. Passive immunization against HIV/AIDS by antibody gene transfer. Viruses 6:428 – 447. https://doi.org/10.3390/ v6020428.
36. Zhou T, Georgiev I, Wu X, Yang ZY, Dai K, Finzi A, Kwon YD, Scheid JF, Shi W, Xu L, Yang Y, Zhu J, Nussenzweig MC, Sodroski J, Shapiro L, Nabel GJ, Mascola JR, Kwong PD. 2010. Structural basis for broad and potent neutralization of HIV-1 by antibody VRC01. Science 329: 811– 817.https://doi.org/10.1126/science.1192819.
37. Scheid JF, Mouquet H, Ueberheide B, Diskin R, Klein F, Oliveira TY, Pietzsch