Sequence, Function, and Regulation of
the Vmw65 Gene of
Herpes Simplex Virus
Type
2
RICHARD F. GREAVESt ANDPETER O'HARE*
Marie CurieResearchInstitute, The Chart, Oxted, Surrey RH8 OTL, United Kingdom
Received 10 June 1991/Accepted 21 August 1991
We determined thesequenceofthegeneforthe virion transactivator protein Vmw65 of herpes simplex virus
type 2 (HSV-2), strain 333. An analysis of the coding sequence revealed an overall high degreeofprimary
sequence conservation (86%) relative to the HSV-1 protein, although the carboxy-terminal region which
encompassesthepowerfulacidic transactivation domain of the HSV-1 proteinwasslightly less well conserved (70%). One importantchange in this regionwas thepresence ofa proline residue ina regionof the HSV-2 protein which is thoughttoformanamphipathic alpha-helix intheHSV-1homolog.Despitetheoccurrenceof this helix-disruptingresidue, theHSV-2 protein exhibited powerful transactivation properties for
immediate-earlytarget promoters.We also demonstrated that the HSV-2 protein formsatranscriptional complex (TRF.C) with the cellular Oct-i protein and target TAATGARAT elements from immediate-early promoters. A
comparison of upstream sequences from the two Vmw65 genes revealed good conservation of proximal
promoterelementsbut considerable divergenceelsewhere. Specifically, theHSV-2promoteralone carries9.5
copiesofa9-bpdirect repeat(GGGGCGGGA) ending 85 bpupstreamofthe conserved TTAAAT element. An
analysisoftranscription factor bindingsites invitrorevealed thatcellular factorSplboundtothe directrepeat
sequence of the HSV-2 promoter and thatcellular factor USF boundto aproximal element presentinboth
HSV-1 andHSV-2promoters. Mutational analysis of the HSV-2promoterdemonstrated that theintegrity of bothofthesebindingsiteswas importantforthefull activityof thepromoter.
The virion protein Vmw65 (VP16, a-TIF, ICP25) of herpes simplex virus (HSV) performstwo known functions during the virus lifecycle. It isamajor structuralcomponentof the virion (25, 52) and as such has an essential function for
normal virus assembly (1, 41). Vmw65 also specifically
transactivates immediate-early (IE) gene expression after virus infection (9, 47). This function is dispensible for infection in tissue cultures at a high multiplicity but is
essential fornormal virusreplicationatlow multiplicities of infection and for virulence after intracranial or
intraperito-neal inoculation in mice (2).
Although Vmw65 does notindependentlybind DNA (35, 37), it forms a complex with cellular transcription factor
Oct-1, which bindstothe octamerorTAATGARAT motifs presentinHSV IEgenes(37,42, 43,48, 49, 53). Induction of
transcription is mediated by the acidic transactivation
do-main locatedatthecarboxy terminus of Vmw65.
Mutagen-esis of the HSV type 1 (HSV-1) protein has allowed the dissection of these constituent activities (20, 55). Residues 49to388 of HSV-1 Vmw65aresufficient for the formation of
transcriptional complex TRF.C (21), which includes
Vmw65, a TAATGARAT element, Oct-1 protein, and a
second cellular factor (27, 30, 61). The formation of this
complexcorrelates tightlywithtransactivation by wild-type Vmw65 (21, 43), providing a mechanism for selective
loca-tion of the activaloca-tion domain(whichis itselfdispensible for TRF.Cformation)ontotargetgenes. We have demonstrated thatpointmutations in theregion between residues 366 and 387 frequently affect the ability of the Vmw65 protein to
participate in TRF.C and have predictedthat this region is
directlyinvolved atan interface within the TRF.Ccomplex
*Correspondingauthor.
t Presentaddress:DepartmentofMicrobiologyandImmunology,
StanfordUniversity School ofMedicine, Stanford,CA 94305-5402.
(21). Consistent with this proposal is the abolition of
com-plex formation bytheinsertion of short two- tofour-residue peptides at (among others) position 369 or 379 (1, 59).
Furthermore, we have shown that a peptide derived from
this region can specifically interfere with transcriptional
complexformation(23).
The acidic transactivation domain is contained within residues 413 to 490 of Vmw65 and retains function when linked to the DNAbinding domains of other proteins (11, 50). This region is predicted to form amphipathic alpha-helices, consistentwith theproposalthathelix formation isa
requirement foracidic activation function(34). Surprisingly, however, the recent results of Cress and Triezenberg (12)
indicate thatalpha-helixformation maybeunimportant and thatspecific hydrophobic interactions and anoverall nega-tivechargearethe crucial featuresrequiredfor function. The
acidicregionisthoughttoactivatetranscription by
contact-ing TFIID (54), TFIIB (33), or a transcriptional adapter (4,
28).
The Vmw65 protein has two identified homologs, both among the alphaherpesviruses. The Vmw65 protein of
HSV-2 has been shown to activate IE gene transcription (45), although its sequence is presently unknown. In con-trast, the Vmw65 protein of varicella-zoster virus, UL10,
whose sequencehas been determined(15), seemsnot to act as a transcriptional regulator or to form a complex with
cellular transcription factors (21a). Knowledge of the
se-quenceandpropertiesof the HSV-2homologwillclearlybe
useful in advancingourunderstanding of structure-function
relationships withinVmw65.
MATERIALSANDMETHODS
Plasmids, mutagenesis, and sequencing. Plasmids pRG90,
pRG91, and pRG131 were used for double-stranded
se-quencing of the HSV-2 Vmw65 gene from strain 333 and 6705
CopyrightC) 1991, American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
contain, respectively, 1.6-kbp KpnI, 0.75-kbp XhoI-KpnI,
and 1.2-kbp SstI-KpnI fragments from plasmid pGR135 (45) inserted into pUC19. Expression vector pRG1 contains HSV-2 strain 333 sequences from an XhoI site in the untranslated leader of Vmw65 to anSstI site in the next open reading frame downstream, cloned via linkers between the HindIII and EcoRI sites of plasmid pCMV-IL2 (13). This procedure places the HSV-2 Vmw65 coding sequence under the control of the powerful human cytomegalovirus IE promoter in a vector which contains the simian virus 40 origin of replication and which can therefore replicate in COS cells. The analogous expression vector pRG70 ex-presses Vmw65 from HSV-1 strain MP and has already been described (21).
Plasmids pRG154, pRG155, pRG163, and pRG164 (see Fig. 7 for a summary) all contain Vmw65 promoter se-quences cloned (via linkers) in the appropriate orientations upstream of the chloramphenicol acetyltransferase (CAT) coding sequence ofplasmid pCATB' (58). The HSV-1 pro-moter sequence was numbered by designating thecentral T ofthe sequence GCTGT +1 (seehighlighting in Fig. 2). This site has been mapped as the mRNA cap site (+2bases) by
Dalrymple et al. (14) and Pellet et al. (46). The HSV-2 promoter sequence was numbered by naming the homolo-gous T residue in the HSV-2 promoter +1 also (see Fig. 2).
Plasmid pRG154, the parent constructfor promoter studies,
contains a 418-bpXhoIfragment from the Vmw65 promoter ofHSV-2 strain 333, stretching from -288 (in the
penulti-mate codon of the HSV-2 UL49 homolog) to +131, 34 bp
upstream of the predictedinitiator codonforVmw65. Mutations were introduced into the promoter sequences by subcloning the sequences into pTZ vectors (39) and performing oligonucleotide-directed mutagenesis on
single-stranded derivatives (31) witha Bio-Rad Mutagene kit. All mutations were verified by double-stranded sequencing of plasmids. Plasmid pRG164 isidenticaltopRG154exceptfor
a single point mutation resulting in an XhoI site at -57. Plasmid pRG174 is identical to pRG154 except for two
transversions which generateanSphI siteat -109. Plasmid pRG178 is similar to pRG174 except that sequences up-streamof theMluI site at -211 have been deleted. Plasmid
pRG163issimilar to
pRG174
exceptthat the viral sequences upstream of the novelSphI site(-109) have been deleted. SeeFig. 7 for a summary oftheseplasmids.
Plasmid pBB5 encodes the IEllOK gene and contains a
4,365-bp BstXIfragment of HSV-1strain MP cloned into the
SmaI site of pUC19. This
fragment
contains IEllOK pro-moter and coding sequences but lacks LAT promoterse-quences (3). Plasmid pBB37 (2a) encodes the IE175K gene under the control of the human
cytomegalovirus
IE pro-moterandcontains an approximately 5-kbp SalI-DraIfrag-mentof HSV-1strain MPclonedintovector
pCMV-IL2
(13).
Reporter plasmid pAB5 (42) contains the HSV-1 IEllOK
promotercloned upstreamof thecoding sequencefor CAT. Plasmid pPOH3 (44)containsthe
thymidine
kinase promoterof HSV-1clonedupstreamofthe
coding
sequencefor CAT.Plasmid sequences were determinedbychain termination sequencing with a Sequenase 2 kit
(USB).
Plasmids wereprepared for analysis by alkaline denaturation. For the HSV-2 Vmw65 gene sequence,
plasmids
pRG1,
pRG90,
pRG91, and pRG131wereusedas
templates.
For compara-tive purposes,wealsosequencedthe promoterregion
of the Vmw65 gene from HSV-1 strain MP. This sequence wasidentical to thepublished sequence(46) for HSV-1 strain
F,
with the singleexception that it lackeda G residueat -68. There are only two additional changes between the
se-quences of these two strains and the sequence of HSV-1
strain
17,
which containsaTresidue instead ofaGresidueat
position
-17 and lacksa Tresidue atposition
+78(14).
Universalprimers
were usedtocommenceHSV-2sequenc-ing;
thereafter, 15-bp
primers
derived from the determinedsequencewere
synthesized
and usedtoextend the sequence until thecomplete
gene wassequenced
on bothstrands.All
oligonucleotides
weresynthesized
on anApplied
Bio-systems 381A
synthesizer.
Complementary
oligonucleotide
pairs
forgel
retardation studies were annealedby
being
heated
together
to70°C
and thenslowly
cooled to30°C.
Oligonucleotide
pairs
containing
binding
sites fortranscrip-tion factors
PHO4,
USF,
AP-1,
andSpl
werekindly
pro-videdby Colin
Goding.
Gel retardation
analysis.
Cell extracts were incubated withprobes
in 20 ,ul ofbinding
buffer,
which contained 25 mMN-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid(HEPES) (pH 7.9),
50 mMKCl,
5 mMdithiothreitol,
1 mMsodium
EDTA,
and0.05% NonidetP-40. A5-min
incubationat
20°C
withnonspecific
DNA(0.5
,ug of sonicated salmon spermDNA per,ul
of nuclearextract; 5,ug/l,
of whole-cellextract)
was followedby
a20-min incubation at20°C
withspecific
unlabelledcompetitor
oligonucleotides
(when
appro-priate)
and thenby
a30-min incubationat20°C
withapprox-imately
0.5 ng of end-labelledoligonucleotide probe.
Gelretardation
analysis (18)
wasperformed
on 4%nondenatur-ing
polyacrylamide gels
with a 19:1acrylamide/bisacryl-amide ratio as described
previously
(21).
Quantitative
esti-mates of labelled
probe
bound were madeby
liquid
scintillation
counting
of retarded bands excised fromdriedpolyacrylamide gels.
Transient
expression
assays.COScellsand Verocellswereroutinely
cultured in Dulbecco modifiedEagle
minimales-sential medium
containing
10% newborncalfserum. Trans-fections and CAT assays werecarried out in COS orVerocells as
previously
described(21).
Whenappropriate,
cells were infected at 18 hpostinfection
with 5 PFU of HSV-1 strain MP per cell and harvested 22 h later. Aquantitative
estimate of
chloramphenicol
acetylation
was obtainedby
excision ofthe substrate and
products
fromthin-layer
chro-matography
plates
andsubsequent
measurementby
liquid
scintillation
counting
in an LKB 216 scintillation counter.Activity
isexpressed
ascounts perminuteappearing
in theacetylated
products.
Cell extract
preparation
andprotein analysis.
For large-scale nuclearextractpreparation,
HeLacells(1
x1010
to 2x 10l°
cells)
were grown insuspension
in Joklik modifiedEagle
minimal essential mediumcontaining
10% newborncalfserum.Nuclearextractswere
prepared
by
themethodofDignam
et al.(16).
Forpreparation
of transfected-cellex-tracts, 2.5 x
107
COScells weretransfected with 20 ,ug ofexpression
vectorby
themethod of Chen andOkayama
(10)
with
N,N-bis(2-hydroxyethyl)-2-aminoethanesulfonic
acid-buffered saline. Whole-cellextractswere
prepared
after40 hasdescribed
by
Wu(60).
For thecomparison
oftransactiva-tion and
complex
formationincellstransfected with Vmw65 from HSV-1 orHSV-2,
equal aliquots
of both total cellular material and solubilized extract wereanalyzed by
sodiumdodecyl
sulfate-polyacrylamide
gel
electrophoresis
(32)
fol-lowed
by
Westernimmunoblotting
(7),
using
monoclonalantibody
LP1(38)
aspreviously
described(20,
21).
Nucleotide sequence accession number. The
2,228-bp
se-quence determined in this work was submitted to the Gen-Bank data base andcanberetrieved underaccessionnumber M75098.
on November 10, 2019 by guest
http://jvi.asm.org/
1- MDLLVDELFADMNADGASPPPPRPAGGPKNTPAAPPLYATGRLSQAQLMP HSV-1
111111 MIii III
1- MDLLVDDLFAD--ADGVSPPPPRPAGGPKNTPAAPPLYATGRLSQAQLMP HSV-2
51- SPPMPVPPAALFNRLLDDLGFSAGPALCTMLDTWNEDLFSALPTNADLYR
49SPPMPVPPAlAl lLDlll GlMDTNFSAGPALC EDl P::lTADl
49- SPPMPVPPAALFNRLLDDLGFSAGPALCTMLDTWNEDLFSGFPTNADLYR
101- ECKFLSTLPSDVVEWGDAYVPERTQIDIRAHGDVAFPTLPATRDGLGLYY
99- ECKFLSTLPSDVIDWGDAHVPERSPIDIRAHGDVAFPTLPATRDELPSYY 151- EALSRFFHAELRAREESYRTVLANFCSALYRYLRASVRQLHRQAHMRGRD
11:: ll::Iliii111111 11111 111111111111111111111111 149- EAMAQFFRGELRAREESYRTVLANFCSALYRYLRASVRQLHRQAHMRGRD
201- RDLGEMLRATIADRYYRETARLARVLFLHLYLFLTREILWAAYAEQMMRP
111 1111 1111111111111111111111111:11111111 1111111 199- RDLREMLRTTIADRYYRETARLARVLFLHLYLFLSREILWAAYAEQMMRP
251- DLFDCLCCDLESWRQLAGLFQPFMFVNGALTVRGVPIEARRLRELNHIRE
1111 111111111111l 1111:11:11:1111111:1111111111111
249- DLFDGLCCDLESWRQLACLFQPLMFINGSLTVRGVPVEARRLRELNHIRE
301- HLNLPLVRSAATEEPGAPLTTPPTLHGNQARASGYFMVLIRAKLDSYSSF 299- HLNLPLVRSAAAEEPGAPLTTPPVLQGNQARSSGYFMLLIRAKLDSYSSV
351-
349-region required for TRF.C formation
TTSPSEAVMREHAYS T1fl~tIZb~?D1~D+APEEAGLAAPRL
ATSEGESVMREHAYSRGRTRNNYGSTIEGLLDLPDDDDAPAEAGLVAPRM
I>> acidic actvation domain
400- SFLPAGH-TRRLSTAPP-TDVSLGDELHLDGEDVAMAHADALDDFDLDML
111 11 11111111111111111111:1
399- SFLSAGQRPRRLSTTAPITDVSLGDELRLDGEEVDIMMWALDDFDLEML
448- GDGDSPGPGFTPHDSAPYGALDMADFEFEQMFTDALGIDEYGG -490 HSV-1 11 :11 11:1 11 111 111111:1:1
449- GDVESPSPGMT-HDPVSYGALDVDDFEFEQMFTDAMGIDDFGG -490 HSV-2 FIG. 1. Deduced primary sequence of the HSV-2 strain 333
Vmw65protein, aligned with theprimary sequenceofthe Vmw65
protein from HSV-1 strain 17 (14). Identical residue pairs are
indicated by a vertical line, and semiconserved residue pairs are
indicated by a colon. Residues in the HSV-1 proteinrequired for TRF.C formationareshaded,asis the prolineresidue atposition436
in the HSV-2 activation domain.
RESULTS
HSV-2 Vmw65 protein sequence. We obtained the
se-quence(Fig. 1andFig.2) of 2,228bpoftheHSV-2 strain 333 genomeknowntocontainthe genefor Vmw65(UL48in the nomenclature of McGeoch et al. [36] for HSV-1). Our determined sequence begins with the last 10 codons ofthe
presumptive HSV-2UL49homolog. The stopcodon forthe UL49readingframe is followed after 29 bp bya consensus
polyadenylation signal (AATAAA) (Fig. 2). On the basis of the overall alignment of the HSV-2 sequence with the
correspondingsequenceof HSV-1 (14, 46)and the
assump-tion that the primary sequence homologies, including, for
example, the TATA box sequence, are indicative of
tran-script homology, the cap site and initiator codon of the HSV-2 Vmw65 genecanbepredictedwith virtualcertainty (Fig. 2),particularly since the resultant mRNA would have the capacity to encode a 490-residue protein with 86%
primary homology to the HSV-1 Vmw65 protein (Fig. 1).
The end of the HSV-2 openreadingframe isfollowed after 91bp byaconsensuspolyadenylation signal (AATAAA)and
subsequently by aTATAA signal, which may be the TATA box site of the HSV-2 UL47homolog. The completeDNA sequence oftheregion canberetrieved from the GenBank database underaccession number M75098.
UL49 UL49
TaqI stop poly A
-233 GTCGA CTTC---CGTACC---CAGAC CACCAA HSV-1 I-28ICTCAG1C1 C1C1T1A 1A1G1 1A1 11 H1S1
-288 CTCGAG aGACCCCGGCCGCGTTCAGACGACAGAC~ CATCAC HSV-2 poly A
-193 CAGGGGTTCA---TTCGGTGTTGGC----GTT----GCGTG---CCTT
I11 1iii III 11 III III -238 GGTCGATTTAACCACTTCGCTGTCAGCACGCGTTTGTGGCGAGGGGCGGG
MluI
-159 TGTTTCCCAATCCGACGG-GGACCGGGACTGG---GTGGCGGG--GGG
11 1 11111 11 1 111111 III
-188 AGGGGCGGGAGGGGCGGGAGGGGCGGGAGGGGCGGGAGGGGCGGGAGGGG
9.5 perfect direct 9bp repeats - SP1 sites
-117 TGGGTTGGAC---AGCCGCCCTCGGTTCG---CCTT---CAC
III 11I 11 11 11 11 III
-138 CGGGAGGGGCGGGAGGGGCGGGAG=GGCGACACGCCTCCCTTCCGAGCGC
USF? CTF?
-84 GTGACAGGAGCCAAT -88
-38
USF SPi?
-GTG-GGGGAAGTCACGAGGTACGGGGCGGCC 1111 1I1111111111111111 11111111
GGGGGACGGGCCGCCCGGAGCGTGGGGAAGTCACGAGGTTTGGGGCGGCA
USF SPi?
'TATA'box mRNA cap
CGTGCGGGTTGCTTAAATGCGGGGTGGCGACCACGGGCT?TCATTCCTCGl -38 CTGGGGTGATACTTAAATGCGGGGTGGTGGACGCGAGATGTCAGTCCTCG
'TATA'box
+13 GGAACGGACGGGGTTCCCGCTGCCCACTTCCCCCCATAAGGTCCGTCCGG
111111111 1111 11111 I111 III
+13 GGGACGCACGGCACCCCCGGCGA----TTCCCTTCGCGAGGGCC--CCGG
111111II11 11I111
+57 -CCTTTTTC---TGTCGCGGGTCC-CGGATCCCTCCCCCCC---TCTC
EcoRV
+113 ACACTCTCTGGGCGGGCGGGGACGATCGCATCAAAA
111111111 Ill III 11III
+97 -CGC-CGCCGGGCGCTCGGGCACG-TCTCATTCGCC
Met
+163 TTCCCGTATCAACCCCACCCASM +185 HSV-1
11111 11 I111 111
+145 TTCCCGGACCCAACCGCCCCC2I +168 HSV-2 +63
AGCCCGATATCGTCT 1IIIH11111
CTCTCGAGATCGTTA XhoI
Met
FIG. 2. Alignment of nucleotide sequences upstream of the reading frames for Vmw65 inHSV-1 strainMPand HSV-2 strain
333. The stop codons and polyadenylation signals for UL49 are
shaded,as aretheinitiator codonsfor Vmw65.Restrictionsites used
for reporter constructs are marked, and the positions of point
mutationsused to createSphI sitesareindicatedby asterisks. Also
shadedaretheputativeTATAboxes,theHSV-1transcriptionstart
site(14, 46), thebinding site for USF demonstrated inthis paper,
andtheSpl binding directrepeatsof the HSV-2 promoter.
There is very good alignment between the HSV-1 and HSV-2 proteins
(86%
homology), and the greatest diver-gence in the primary sequence occurs in the carboxyl-terminal acidic domain(70%
homology). Theoverall strong homology isconsistentwith conservation of the dual roles of Vmw65 as an essential structural component and as the viriontransactivatorofIE genes. As formostregionsof the genome, theregion
encoding
Vmw65 can be containedwithin viable intertypic recombinants(40).
The primarysequenceof the HSV-2
protein
in theregion
fromresidues 366to388(which
wehavedemonstratedtobe crucial for Vmw65-Oct-1 complexformation)
showed two fairly conservativechanges(Ala-367
toGly; Lys-370
toArg)
(Fig. 1). Although we have determined that
lysine
370 is critical forcomplex formationin the HSV-1protein,
wehave also shown that its mutation toarginine
istolerated,
consis-tent with the presence of
arginine
at thisposition
in the HSV-2protein (21b).Ananalysis oftheprimarysequenceof the acidicdomain
of the HSV-2
protein
revealed a moreunexpected
feature: the presenceofaproline
residueatposition
436(Fig.
1).
TheXhoI stop
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.322.552.76.415.2]a)
A\k (A\Mi
( AMP
b)
ISV'
I(SV-V
0-I|.i11:~~~~~~~~~~~~~JK(.\ r_ _
IWA
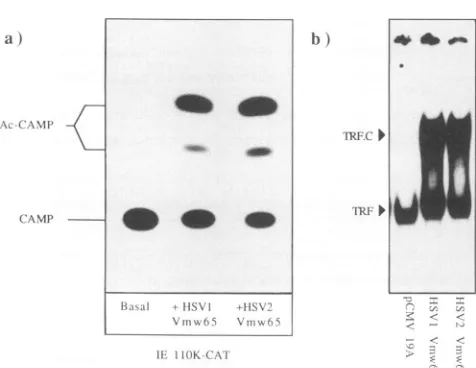
FIG. 3. (a) Transactivation of the IE 110K-CAT construct by
cotransfection with Vmw65 expression vectors. COS cells were
transfected with 20 ng oftarget plasmid and 10 ng ofpRG50 for
HSV-1Vmw65 expression, pRG1 for HSV-2 Vmw65 expression, or
controlvectorpCMV19Aforbasal activity.CAT activity in soluble
extracts was measured at40hposttransfection. Ac-CAMP, acety-lated chloramphenicol products; CAMP, chloramphenicol
sub-strate. (b) Formation of the TRF.C transcriptional complex on a
TAATGARAT element by HSV-1 Vmw65 and HSV-2 Vmw65.
End-labelledoligonucleotideTAAT24 (1 ng)wasincubated together
withaHeLa cell nuclearextract (1,ul)and extracts of transfected
COScells (1,ul). Bound oligonucleotide wasresolved by
nondena-turing electrophoresis; unbound probe isnot shown in this figure.
Vmw65 was expressed in COS cells by transfection with vector pRG50 (HSV-1)orpRG1 (HSV-2). Thebindingprofile ofanextract
madefrom cells transfected with the control vector is shown in
parallel(pCMV19A).Approximatelyequalamountsof Vmw65were
detected in pRG1- and PRG50-transfected cells by Western blot
analysis of the extracts with type-common monoclonal antibody
LP1(datanot shown).
acidic activation domain has been predicted to form two
amphipathic alpha-helices, but the presence of a proline
residueinthe HSV-2 proteinmay preclude theformation of
atleastoneof these structures. Alternatively, ifahelix does
forminthis region, it wouldbe restricted toaround residues
438 to 448. This region of the HSV-2 protein is clearly
functional intranscriptional activation, as we show below;
we and others have also demonstrated that an Ala-to-Pro
switchatanadjacent residuein the HSV-1 activation domain
has no detectable effects on its function (12, 41a). Despite
the slightly lower levelofsequence homology between the
acidic domains, key features are clearly well conserved.
Negative charge and its distribution are very similar in the
two proteins (all acidic residues present in the HSV-1
activation domain are present in its HSV-2 equivalent),
althoughtherearethreeAsp-to-Glu changes. The carboxyl-terminalregionof theHSV-2protein also hastwoadditional
acidic residuesas theresult of Ala-to-Asp changes. HSV-2 Vmw65 protein function. An expression vector (pRG1) which expresses Vmw65 of HSV-2 strain 333 was
constructed. The vectorused has asimian virus40origin of replication andthe IEpromoterfrom human
[image:4.612.58.296.69.253.2]cytomegalovi-rus and can be used toproduce large quantities offoreign protein intransfected COS cells. We previously described similarvectors whichexpressthe Vmw65proteinof HSV-1 strainMP (20). Assays fortransactivationofanIE promoter-CATconstruct byHSV-1 and HSV-2 proteinsareshown in
Fig. 3a. Cotransfection ofvectors expressing Vmw65 from
HSV-1 or HSV-2 stimulated CAT expression to similar
extents, indicating that the proline residueat position436 in
the HSV-2 acidic domain has no substantial effect on the
activation function. Figure 3b shows that HSV-2 Vmw65 overexpressed in COS cells is also fully functional for the
formation of transcriptional complex TRF.C with cellular
proteins and target promoter sequences. This result was as we would predict, since TRF.C formation seems essential fortransactivation (1, 21, 43, 59).
The HSV-2 Vmw65 promoter contains a repeat structure. The intergenic region between the UL49and Vmw65(UL48) reading frames from HSV-1 strain MP andHSV-2strain333 is shown in Fig. 2. The homologywithinthis region isclearly
not as great as that within the protein coding region. How-ever, there is very clear homology just upstream of the mapped cap site of the HSV-1 transcript. This homology
includes a conserved CTTAAATGCG sequence at -27, presumably the functional TATA box (14, 46), aGGGGCG
GC sequence at -47, and a GGGGAAGTCACGAGGT
sequence at -65. On the basis of the conservation of the
nucleotide sequence and spacing, it would seem likely that
the latter two elements may representcis-acting sites shared
between the two promoters and may be binding sites for transcription factors. Upstream of the -65 element and downstream of the UL49 polyadenylation signal there is considerably less homology between the two promoters. The HSV-1 promoter carries only unique sequence in this region which contains potential binding sites for transcription fac-tors USF (CACGTG) at -87 and CTF (CCAAT) at -74. In contrast, the HSV-2 promoter has 9.5copiesof a 9-bp direct repeat (GGGGCGGGA) flanked by a unique sequence. These repeated elements closely resemble the binding site consensus sequence for cellular transcription factor
Spl
(6). On the basis of these observations, it seems that the two promoters share similar structures proximal to the transcrip-tion start site but may have different distal elements.Expression and transactivation of the HSV-2 Vmw65
pro-moter. We constructed plasmids which linked the promoter
region to the coding sequence for CAT. The fragment used spanned from the
XhoI
site at position -288 in the penulti-mate codon of UL49 to theXhoI
site at position +131 in the5'-untranslated leader region of the Vmw65 gene. The
activ-ities of the promoter and mutant versions were assessed by
determining CAT activity in transfected Vero cells. A measure of basal activity in a dose-response experiment is shown in Fig. 4a. The HSV-2 promoter construct (pRG154) showed good basal activity, approximately twice that of the delayed-early thymidine kinase promoter con-struct (pPOH3). The results of virus superinfection of trans-fected cells are shown in Fig. 4b. The HSV-2 promoter was clearly induced by virus superinfection; the induction ratio
relative to mock-infected controls was in the range of 4- to 12-fold. Superinfection by HSV-1 or HSV-2 induced
pro-moter activity to similar levels (data not shown). HSV-1 IE
transactivator proteins were introduced separately in cotransfection experiments, and typical results from these experiments are shown in Fig. 4b. Cotransfection with a vector expressing the
IEllOK
protein induced expression from the HSV-2 promoter by 25-fold, and cotransfection with a vector expressing theIE175K (ICP4) protein induced expression by approximately 15-fold.A conserved binding site for cellular transcription factor USF is required for full activity of the HSV-2 Vmw65 pro-moter in transient expression assays. The 16-bp sequence GGGGAAGTCACGAGGT is located approximately 65 bp
ob
do
on November 10, 2019 by guest
http://jvi.asm.org/
a) b)
a) CDef
x
I5
b)
basal nf basal -Ell OK +1E175K
pRG1S4 (HSV-2 promoter)
FIG. 4. (a) Dose-responseassays of CAT expression from HSV Vmw65 promoter-CAT constructs. Vero cells(106cells) were
trans-fected with various amounts of the indicated plasmids, and CAT
activity insoluble cell extracts was measured at 40 h
posttransfec-tion. (b) Induction of expression from Vmw65 promoter-CAT con-structsbysuperinfection with HSV-1 strain MP or by cotransfection with genesfor HSVIEproteins. Vero cells were transfected with
pRG154(1 ,ug) andinfectedat 18 haftertransfection with 5 PFU of
HSV-1percell. CAT activity insolubleextracts was measured at 20
h postinfection. For the cotransfection experiments, target con-structpRG154(1 ,ug) wastransfected intoVero cellstogether with
IEllOK expression vector pBB5 (50 ng) or IE175K expression vectorpBB37 (20 ng). CAT activity in soluble cell extracts was measured at 40 hposttransfection. inf, infected.
upstream ofthe transcription start site for the HSV-1
pro-moterand isperfectly conserved in both HSV-1 and HSV-2 promoters. To test whether this sequence was a
transcrip-tion factor binding site, we synthesized 29-bp
oligonucleo-tides spanning the conserved region and examined the
protein binding profile in gel retardation assays. Using a
nuclear extractofHeLacells (16)as asourceoftranscription
factors andoligonucleotides from either the HSV-1 orthe HSV-2 promoter, weobserved identicalcomplexes
consist-ing ofa single major species and a minor, faster-migrating species (Fig. Sa).Wenoted similaritieswithin theprobesto
the binding sites for cellular factors AP-1 and USF and
binding experiments were therefore conducted with unla-belled competing oligonucleotide pairs containing known
bindingsitesforAP-1,USF,Spl,and yeastfactor PHO4. Of these, the USF site competed strongly for the specific
complex,thePH04 sitecompetedweakly,andtheAP-1and
Spl sites did not compete significantly at any of the doses tested (datanot shown). We therefore tentatively assigned
the binding activity from HeLa cell nuclei as USF. Weak
competition bythe PH04site is consistent with this
conclu-AtNII.1' iS%62 VZV HSIVI
F
- c h |- c h |- c h - c h
c)
=60
N l.SFl 0
4 n.s. >
AdMILP HS V2 ('tIA%(t
hi
i
12 3 1 2 3s1 1 x
pRGil64
hl;sE: site poitTl IttulEtitit
ha,al -110K --?'K
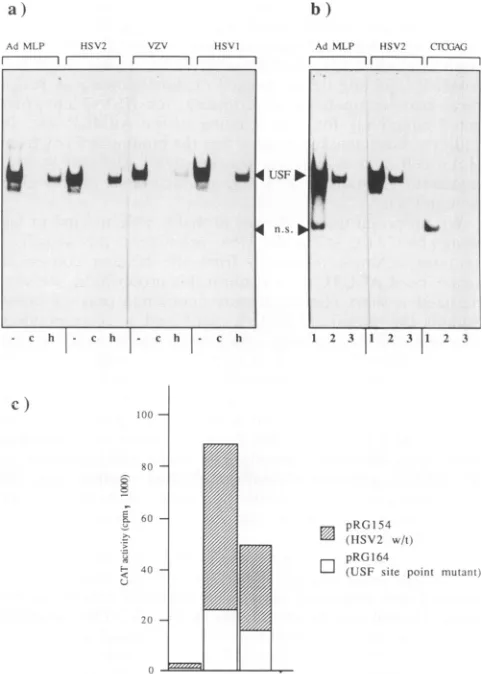
FIG. 5. (a) Binding ofHeLa cellnuclear factorUSF to similar elements in AdMLP (Ad MLP), the HSV-2 Vmw65 promoter
(HSV2), the varicella-zoster virusUL10 promoter(VZV),and the HSV-1Vmw65 promoter(HSV1). Bound end-labelled oligonucleo-tideprobeswereresolvedby nondenaturing gel electrophoresis.For purposes of increased resolution, unbound probe was
electro-phoresed offthe gel but was shown to be in excess in parallel experiments. Lanes: -, competition by nonspecific competitor
DNAonly;c,specific competition by AdMLP USF site oligonucle-otides (100-fold molar excess); h, formation ofthe complex by a
heat-treated HeLa cell nuclearextract. (b) Binding ofaHeLacell nuclear factor to USF sites in AdMLP (Ad MLP), the HSV-2
Vmw65 promoter(HSV2),and a mutantHSV-2 Vmw65 promoter
(CTCGAG). Bound end-labelled oligonucleotide probes were
re-solvedbynondenaturing gel electrophoresis;unboundprobe isnot
shown. Lanes: 1, competition by nonspecific competitor DNA at
100ng;2,competitionbynonspecific competitorDNAat500 ng;3, competition bynonspecific competitorDNAat500 ng and AdMLP
oligonucleotides (100-foldmolarexcess). USF,USFcomplex;n.s.,
nonspecific complex. (c) Inductionofexpressionfromwild-typeand
mutant Vmw65 promoter-CAT constructs by viral IE proteins
supplied by cotransfection. Target promoters (1 ,ug) were
trans-fectedinto 106Vero cellstogether with IEllOK expressionvector
pBB5 (50 ng) or IE175K expression vector pBB37 (20 ng). CAT
activity in solublecellextracts wasmeasuredat40h
posttransfec-tion. w/t, wild type.
sion, as the PH04 protein is, like USF, a helix-loop-helix protein and binds to a related motif(18a). To confirmour
conclusion,weend labelledanoligonucleotide pair
contain-ingthe USFsite from the adenovirus 2
major
late promoter (AdMLP) and used it inparallel gelretardation assays. TheW
to
W
i
4a
db
"b
40
4D 4D
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.63.312.80.371.2] [image:5.612.324.565.82.419.2]result (Fig. Sa) shows that the mobilities of complexes formed on the AdMLP USF probe and the HSV element probes were identical (lanes marked -) and that all of the complexeswere specificallycompetedforbythe unlabelled AdMLP USFsite (lanes marked c). Furthermore, in recip-rocal cross-competition experiments, the HSV-2 site
com-peted effectively for USF binding to the AdMLP site. In
addition,lanes marked hshowthat thebinding activityfrom HeLa cell nuclear extracts was relatively resistant to heat treatment (10 min at 80°C), a characteristic of the USF protein (51).
We proposed thatUSF was
probably
ableto bindto the sequence CACGAG in the HSV promoters; this sequencecontains a single mismatch from the binding consensus
sequence(CACGTG). To confirmthisproposition, we syn-thesized a short oligonucleotide containing only 16 bases around the proposed HSV-2 motif and a corresponding oligonucleotide containingasinglebase-pairmutation within the site (CACGAG to CTCGAG). The results (Fig. Sb)
demonstrated that the shortwild-typeoligonucleotide bound
the factor as efficiently as the longer probe and with an
efficiency comparabletothatoftheAdMLPprobe (compare
lanes 1 and 2from the AdMLP panel with thecorresponding
lanesfrom the HSV-2 panel). The single point mutation in the HSV-2 sequence virtually abolished binding (Fig. Sb) and,whenassayed bycompetitionassays(datanotshown),
theaffinity of USF for this site was apparently reducedbyat
least 100-fold.
To determine the significance of the USF site, we
intro-duced this point mutation into the HSV-2 promoter-CAT plasmid and measured activity in transient expression
as-says. Typical results are shown in Fig. Sc. The construct
containing the point mutation exhibited basal activity
ap-proximately three- tofourfold lower than that of the wild-type promoter but remained inducible by the IEllOK and IE175K proteins in cotransfection experiments (Fig. Sc).
Over the course ofseveral experiments, the ratio of induc-tion by either the IEllOK or the IE175K protein was
approximately the samefor the wild-type and mutant
pro-moters. These results suggest thatUSFbinding contributes
to the high basal and induced activities of the HSV-2 promoterintransient assays but that it isnotrequiredfor the
mediation of induction by viral IE products. Weanticipate
that the homologous site is involved in the previously
demonstrated constitutive and induced expression of the HSV-1 promoter (5).
The9-bp repeat elements can bind cellularfactorSpl and are required for full activity of the HSV-2 promoter in transient assays. The major difference between the HSV-2 and HSV-1 promoters is the presence of the 9-bp repeat
element,andwethereforeinvestigated whether the element
boundacellular transcription factorand whetherthis bind-ing was involved in the HSV-2 promoter activity. A 26-bp end-labelled probe containing two tandem copies of the repeat wasgenerated and was used togetherwith a HeLa cell nuclear extract in gel retardation assays. The probe bound
specifically to a factor in the nuclear extract (Fig. 6a).
Competition experiments with unlabelled oligonucleotide pairs containing AP-1, USF, yeast PHO4, and Spl sites established that the binding activity seen probably repre-sented cellular transcription factor Spl. An end-labelled
probe containing the Spl site of the adenovirus EIla late promoter(19)was runin a parallel gel retardation assay and bound a complex virtually identical in appearance and
mobilitytothatfound with the HSV-2 repeat probe (Fig. 6a).
Complexes
formed with both probes were also effectively.-\J1i-.)l.ile'SPI }IISV-SPil a)to probe repeats prtlhe
r II--- 1
I
S11 .,.
PIli D
Ul
b)
Si-,I 0
P.S.D
I
- c.
.I.n
+AdiF.2 latc SPI
i1te cr mpeltpiCtr
c) 150S
120)
A-9 (1 CS
F- 6
3
Ad E2 alae SPI
site probec
i
+AdE2 aleSil1 Site coInipCeitiIr
IrD 7
repcats copiT
S pR(3154
(IISV2 w/t)
D
pR(GI
)63rOISEV2 no repeats)
FIG. 6. (a) BindingofHeLa cell nuclear factor Spl to similar elements in the adenovirus Ella late promoter and the HSV-2
Vmw65 promoter direct repeats. Bound end-labelled
oligonucleo-tide probes were resolved by nondenaturing gel electrophoresis;
unboundprobe isnotshown. Adenovirus EIIalate promoter Spl
site oligonucleotides were used as specific competitors in the
amounts indicated. (b) Competition for Spl binding to an
end-labelled EIlalate promoter Spl site probe. Adenovirus Ella late promoter Spl site oligonucleotides and unlabelled HSV-2 repeat
oligonucleotides wereusedas specific competitorsin the amounts indicated. Bound end-labelledoligonucleotide probeswereresolved
by nondenaturinggelelectrophoresis;unboundprobeisnotshown.
Spl, Spl complex; n.s., nonspecific complex. (c) Induction of
expressionfromwild-typeandmutantVmw65promoter-CAT
con-structs by viral IE proteins supplied by cotransfection. Target
promoters (1
pLg)
were transfected into Vero cells together withIEllOK expression vector pBB5 (50 ng) or IE175K expression vector pBB37 (20 ng). CAT activity in soluble cell extracts was
measuredat40 hposttransfection. w/t,wild type.
competed
forby
a 100-fold molarexcessof unlabelled Ella late promoterSpl
site(Fig.
6a).Figure
6b showsacompe-titionassaytoexamine the relativeaffinities ofthe Ella late promoter
Spl
site and thetwotandem9-bp
repeatsforSpl.
Unlabelled
oligonucleotides
atdifferentconcentrationswereused to compete for
Spl
binding
to a labelled Ella late promoterprobe.
Quantitation
ofthisexperiment by
bandexcisionand scintillation
counting
showedthat theaffinity
ofon November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.313.546.75.451.2]HSV-2 Vmw65 6711
pRG 154 ___ iH~~~~CA
pRG164 CAT
USF
pRG174 ..pAT
pRG178 'T.hM. CATI
Sphl
pRG 163
Sphl
~~~~~S
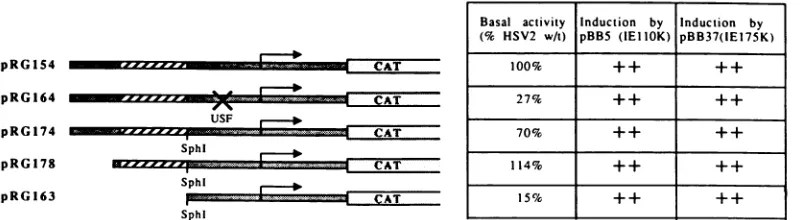
FIG. 7. Diagrammatic representation ofthe promoter-CAT constructs used, together with relative basal activities in transient assays
(averages ofatleast three independent experiments)andinducibility by viral IE transactivators(averagesofatleastthreeexperiments): ++
= 8-to25-fold. HSV-2promotersequencesareshaded,and HSV-2directrepeats arehatched.
Spl for the HSV-2 site was approximately 10- to 20-fold
lower than its affinity for the EIla latepromoter Spl site.
Toinvestigate whether the 9-bprepeatswere involvedin
the activity of the HSV-2promoter, weintroducedanSphI
siteimmediately downstream of therepeatsby mutagenesis. Nearwild-typelevels of expressionwereseenfrom plasmids intowhich this SphI site was introduced (pRG174) orfrom
which unique sequences 5' to the repeats were deleted (pRG178) (Fig. 7). A plasmid (pRG163) in which repeat sequences 5' tothe introduced SphI site were deleted was made. Typical results of the effects of this deletion on expression from the HSV-2promoter are showninFig. 6c.
Basalexpressionwasreduced by four-tofivefold.However,
expression remainedopento transactivation by the IEllOK
and IE175K proteins, suggesting that the repeats are not
required to mediate either of theseactivities (Fig. 6c). DISCUSSION
Wehavereported thesequence of the coding and regula-toryregions oftheVmw65geneof HSV-2.Acomparison of
theencoded protein sequence with theHSV-1 gene reveals
products ofidentical lengths: 490 residues. Vmw65 peptide
sequence conservation between the two viruses (86%) is high and comparabletothatseen withotheressential HSV
proteins, DNA polymerase (90%), alkaline exonuclease (80%), glycoprotein D(82%), andglycoprotein B (86%) (8, 17, 56, 57). The strong sequence conservation seenis
con-sistent with the dual role of Vmw65 in both viruses as a
majorand essential structural componentand as thevirion
transactivatorofIEgene transcription.
Our aim atthe outset of this work was a comparison of
peptide sequences in regions of the proteins required for transactivation. The region between residues 366 and 388, which we have proposed to form an interactive interface
during transcriptional complex formation (21, 23), as
ex-pected from the overall conservation, showed no major
difference between thetwo viruses. A change in the acidic activation domain wasof greater interest. The HSV-2 pro-teinhas aproline residueatposition436. This helixbreaker ispositioned centrally in a regionpredicted to form a long
amphipathicalpha-helix inthe HSV-1 protein. Ifthe struc-tureof thisregionisconserved between thetwoviruses,the
predicted helix may not be formed in either protein. Our observation(41a)thatanalanine-to-prolineswitchatresidue 436 does not affect the function of the HSV-1 activation domain would seem to confirm this idea. Our data are
consistent with a recent report which demonstrated that
prolinesubstitutionsatposition432or436 hadnosignificant
effect onthetransactivation function of Vmw65 (12). Over-all,theresultssuggesteitherthatalpha-helix formation isnot necessaryfor theactivity of this regionorthat ifahelix does
form, it is a short helix located within the region from positions 438 to 448 (at residue 448, the helix probability significantly declines and the protein sequence enters a
proline- and glycine-rich segment).
The conservation of the sequence between the two
Vmw65promoters isnot extensive and centersaround two
elementsupstreamofaconservedTTAAATelement. One of
the two conserved elements in the HSV-1 and HSV-2
promoters can bind in vitro to cellular protein USF. USF
was characterizedasthe cellulartranscription factor which
bindstothe hexamersequenceCACGTGinAdMLP(51). It
isamember, alongwithMyoD andc-myc,ofthe
helix-loop-helixfamily ofDNAbinding proteins (22).Thecorebinding
site in the HSV promoters(CACGAG)hasasinglebase-pair
mismatch relative to the characterized core site but still
binds USF with a high affinity-about 30% that of the
AdMLP site. Inaddition,ourresultswithaUSFbindingsite
point mutant in transient assays show that USF binding
correlates with efficientexpressionfrom the HSV-2Vmw65 promoter. We have alsodemonstratedthebindingof USFto anelement inaconservedpositionupstreamof thepredicted
TATA box of the varicella-zoster virus ORF10 gene, the
homologof the HSV Vmw65 gene. It isintriguingto specu-late that the preservation of this site may reflect some commonfeature intheregulationof thesehomologousgenes
during the virus life cycle. We have searched the HSV-1 genome for potential USF sites in the upstream regions of other genes. The searches revealed possible USF binding
sites intheregulatory regionsof theUL5, UL33, andUL55 genes. A binding site for USF has also recently been
characterized inanearly gene promoter of another herpes-virus, humancytomegalovirus (29).
Therepeated 9-bpmotifstructureisuniquetotheHSV-2
promoter. We show that this element isan SP1 bindingsite
and isalsorequiredfor efficientexpressionfrom the HSV-2 promoter. However, neither the USF site northe Spl site
accountsfor the inducibilityof thepromoterby IE
transac-tivators. Some other feature of the proximal promoter or
leader sequences must beresponsible.
It is possible thatthe difference in promoter structure is
reflected in a difference in the temporal regulation ofthe
HSV-2 gene compared with the HSV-1 gene. Previous
results demonstrated that CAT constructs containing the
promoter for the HSV-1 gene were inducible by virus
superinfection and that the kinetics of Vmw65 expression during HSV-1 infection followed classical leaky-late (or
Basal activity Induction by Induction by
(% HSV2 wAt) pBB5 (IEIIOK) pBB37(1E175K)
100% ++ ++
27% ++ ++
70% ++ ++
114% ++ ++
15% ++ ++
VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.118.519.79.189.2]P-y)-type
kinetics (5, 62). Earlier results indicated that Vmw65 expression in HSV-1-infected cells was repressed but was still detected in the absence of virus DNAreplica-tion (24, 26).
Our current work indicates that the expression of the
HSV-2Vmw65 genemay be unaffectedbytheinhibition of DNA synthesis (21b), indicating that for HSV-2, Vmw65
expression may follow classical delayed-early (or 3)-type
kinetics. Comparative analysesof these twopromoters and of theregulationof theexpressionof Vmw65 in HSV-1- and HSV-2-infected cells are now in progress to examine this
point.
ACKNOWLEDGMENTS
Thisworkwas fundedbythe MarieCurieMemorial Foundation.
We thank Tony Minson for continued supply of monoclonal antibody LP1 and Colin Goding for oligonucleotides. The typing skillsof Jean Marraregratefully appreciated.
REFERENCES
1. Ace,C.I.,M.A.Dalrymple,F. H.Ramsay,V. G.Preston,and
C. M.Preston. 1988. Mutationalanalysisof theherpes simplex virus type 1 trans-inducing factor, Vmw65. J. Gen. Virol.
69:2595-2605.
2. Ace,C.I.,T. A.McKee,J.M.Ryan,J.M.Cameron,andC. M.
Preston. 1989. Construction and characterization ofa herpes simplexvirustype 1 mutant unable to transinduce
immediate-earlygeneexpression. J. Virol.63:2260-2269.
2a.Batchelor,A. Unpublishedobservations.
3. Batchelor,A.H.,and P.O'Hare. 1990.Regulationand cell-type-specific activityofapromoterlocatedupstreamof the latency-associated transcript ofherpes simplex virus type 1. J. Virol.
64:3269-3279.
4. Berger,S.L.,W. D. Cress, A.Cress, S.J.Treizenberg,and L. Guarente. 1990. Selective inhibitionof activated but not basal
transcription bythe acidicactivation domainofVP16:evidence
fortranscriptional adaptors. Cell61:1199-1208.
5. Blair, E. D., C. C. Blair, and E. K. Wagner. 1987. Herpes simplexvirus virionstimulatory proteinmRNAleader contains
sequenceelements which increasebothvirus-induced transcrip-tion andmRNA stability.J. Virol.61:2499-2508.
6. Briggs, M. R.,J. T. Kadonga, S. P. Bell,and R. Tjian. 1986.
Purification and biochemical characterisation of the promoter-specific transcription factor, Spl. Science 234:47-52.
7. Burnette, W. N. 1981. "Western blotting": electrophoretic transfer ofproteins from sodium dodecyl sulphate
polyacryl-amidegelstounmodifiednitrocellulose andradiographic
detec-tion with antibody and radioiodinated protein A. Anal. Bio-chem. 112:195-203.
8. Bzik,D.J.,C.Debroy,B. A.Fox,N.E.Pederson,and S. Person. 1986. The nucleotide sequence ofthegB glycoproteingeneof
HSV-2 andcomparisonwiththecorrespondinggene of HSV-1.
Virology155:322-333.
9. Campbell, M. E. M., J. W. Palfreyman, and C. M. Preston.
1984. Identification of herpes simplex virus DNA sequences
whichencodeatrans-acting polypeptide responsiblefor
stimu-lation ofimmediateearly transcription.J. Mol. Biol. 180:1-19. 10. Chen, C., and H. Okayama. 1987. High-efficiency
transforma-tion of mammalian cells by plasmid DNA. Mol. Cell. Biol.
7:2745-2752.
11. Cousens,D.J.,R.Greaves,C. R.Goding,andP.O'Hare.1989. The C-terminal 79 amino acids of the herpes simplex virus
regulatory protein, Vmw65, efficientlyactivatetranscriptionin yeastand mammalian cells inchimeric DNA-binding proteins.
EMBOJ.8:2337-2342.
12. Cress, W. D., and S. J. Triezenberg. 1991. Critical structural elementsoftheVP16transcriptionalactivation domain.Science 251:87-90.
13. Cullen, B. R. 1986. Trans-activation of human
immunodefi-ciencyvirusoccursviaabimodalmechanism. Cell46:973-982.
14. Dalrymple, M. A., D. J. McGeoch, A. J. Davison, and C. M.
Preston.1985. DNA sequence of theherpessimplexvirustype
1 gene whose product is responsible fortranscriptional activa-tion of immediateearlypromoters.Nucleic Acids Res. 13:7865-7879.
15. Davison, A. J., and J. E. Scott. 1986. The complete DNA sequenceof varicella-zoster virus. J. Gen. Virol. 67:1759-1816. 16. Dignam, J. D., R. M. Liebowitz, and R. G. Roeder. 1983. Accurate transcription initiation by RNA polymerase II in a
solubleextractfrom isolated mammalian nuclei. Nucleic Acids Res. 11:1475-1489.
17. Draper, K. G., G. Devi-Rao, R. H. Costa, E. D. Blair, R. L. Thompson, and E. K. Wagner. 1986. Characterization of the genes encoding the herpes simplex virus type 1 and type 2 alkaline exonucleases and overlapping proteins. J. Virol. 57: 1023-1036.
18. Fried,M., and D. M. Crothers. 1981.Equilibria and kinetics of lac repressor-operator interactions by polyacrylamidegel elec-trophoresis. Nucleic Acids Res. 13:7847-7863.
18a.Goding, C. Personal communication.
19. Goding, C. R., S. M. Temperley, and F. Fisher. 1987.Multiple
transcription factors interact with the adenovirus-2 ElI-late promoter: evidence for a novel CCAAT recognition factor. Nucleic Acids Res. 15:7761-7780.
20. Greaves, R., and P. O'Hare. 1989. Separation ofrequirements
forprotein-DNA complex assembly from those for functional activity in the herpes simplex virus regulatory protein Vmw65. J. Virol.63:1641-1650.
21. Greaves, R., and P. O'Hare. 1990. Structural requirements in the herpes simplex virus type 1 transactivator Vmw65 for interactionwith the cellular octamerprotein and target TAAT-GARAT sequences. J. Virol. 64:2716-2724.
21a.Greaves,R. F. Unpublished observations.
21b.Greaves,R.F., and P.O'Hare. Unpublishedobservations. 22. Gregor, P. D., M. Sawadogo, and R. G. Roeder. 1990. The
adenovirus major late transcription factor USF isamember of the helix-loop helix group ofregulatory proteins and binds to
DNA as a dimer. Genes Dev.4:1730-1740.
23. Haigh, A., R. F. Greaves, and P. O'Hare. 1990. Interference with the assembly of a virus-host transcription complex by
peptide competition. Nature(London) 344:257-259.
24. Hall,L.M., K. G. Draper, R. J. Frink, R. H. Costa, and E. K. Wagner. 1982.Herpes simplex virus mRNA species mapping in EcoRIfragment I. J. Virol. 43:594-607.
25. Heine, J. W., R. W. Honess, E. Cassai, and B. Roizman. 1974. Proteins specified by herpes simplex virus. XII. The virion
polypeptidesoftype 1 strains. J. Virol. 14:640-651.
26. Honess,R.W.,and D.H. Watson. 1977. Herpessimplexvirus resistance and sensitivity to phosphonoacetic acid. J. Virol. 21:584-600.
27. Katan,M., A. Haigh, C. P. Verrijzer, P. C. van der Vliet, and P. O'Hare. 1990.Characterisation ofacellular factor which
inter-actsfunctionally with oct-1 in the assembly of a multicomponent
transcriptioncomplex. Nucleic Acids Res. 18:6771-6780. 28. Kelleher,R. J., P. M. Flanagan, and R. D. Kornberg. 1990. A
novelmediator between activatorproteins andthe RNA
poly-meraseIItranscription apparatus. Cell 61:1209-1215.
29. Klucher,K.M.,and D. H.Spector. 1990. The human cytomeg-alovirus 2.7-kb RNA promoter contains a functional binding site for the adenovirus major late transcription factor. J. Virol. 64:4189-4198.
30. Kristie, T. M., J. H. LeBowitz, and P. A. Sharp. 1989. The
octamer-binding proteins form multi-protein-DNA complexes with theHSV alpha TIF regulatory protein. EMBO J. 8:4229-4238.
31. Kunkel, T. A. 1985.Rapid and efficient site-specific mutagenesis
without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
32. Laemmli, U. K. 1970.Cleavageof structural proteins during the
assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
33. Lin,Y.S., andM. R. Green. 1991. Mechanism of actionof an acidictranscriptionalactivator in vitro. Cell 64:971-981. 34. Ma, J.,and M. Ptashne. 1987. A new class of yeast
on November 10, 2019 by guest
http://jvi.asm.org/
tional activators. Cell 51:113-119.
35. Marsden, H. S., M. E. M. Campbell, L. Haarr, M. C. Frame, D.S. Parris, M. Murphy, R. G. Hope, M. T. Muller, and C. M. Preston. 1987. The 65,000-Mr DNA-binding and virion trans-inducing proteins of herpes simplex virus type 1. J. Virol. 61:2428-2437.
36. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan, M.C. Frame, D. McNab, L. J. Perry, J. E. Scott, and P. Taylor. 1988. The complete DNA sequence of the long unique region in the genome of herpes simplex virus type 1. J. Gen. Virol. 69:1531-1574.
37. McKnight, J. L., T. M. Kristie, and B. Roizman. 1987. Binding
of the virion protein mediating alpha gene induction in herpes
simplex virus 1-infected cells to its cis site requires cellular
proteins. Proc. Natl. Acad. Sci. USA 84:7061-7065.
38. McLean, C., A. Buckmaster, D. Hancock, A. Buchan, A. Fuller, and T. Minson. 1982. Monoclonal antibodies to three
non-glycosylated antigens of herpes simplex virus type 2. J. Gen.
Virol.63:297-305.
39. Mead, D. A., E. Szczesna-Skorupa, and B. Kemper. 1986.
Single-stranded DNA 'blue'T7 promoterplasmids: a versatile promoter system for cloning and protein engineering. Protein
Eng. 1:67-74.
40. Morse, L. S., L. Pereira, B. Roizman, and P. A. Schaffer. 1978. Anatomyofherpes simplexvirus (HSV) DNA. X. Mapping of
viralgenes by analysis of polypeptides and functions specified
by HSV-1 x HSV-2 recombinants. J.Virol. 26:389-410. 41. Moss, H. 1989. Properties of the herpes simplex virustype 2
trans-inducing factor Vmw65 inwild-type andmutantviruses.J.
Gen. Virol. 70:1579-1585.
41a.O'Hare, P. Unpublished observations.
42. O'Hare, P., and C. R. Goding. 1988. Herpes simplex virus
regulatory elementsand the immunoglobulin octamerdomain binda commonfactorandarebothtargetsforvirion transacti-vation.Cell52:435-445.
43. O'Hare,P., C. R. Goding, and A. Haigh. 1988. Direct combina-torial interaction between a herpes simplex virus regulatory
proteinand a cellularoctamer-binding factormediates specific inductionof virus immediate-early geneexpression. EMBO J. 7:4231-4238.
44. O'Hare, P.,andG. S.Hayward.1985.Evidence foradirectrole
for both the 175,000-and110,000-molecular-weight immediate-early proteins of herpessimplex virus inthetransactivation of delayed-earlypromoters. J. Virol.53:751-760.
45. O'Hare, P., and G. S. Hayward. 1985. Three trans-acting regulatory proteins of herpes simplex virus modulate immedi-ate-earlygene expressioninapathway involving positive and
negative feedback regulation.J. Virol.56:723-733.
46. Peliett,P.E.,J.L.C.McKnight,F.J. Jenkins, and B. Roizman. 1985.Nucleotide sequenceandpredicted amino acidsequence
ofaprotein encoded inasmallherpessimplexDNAfragment capable oftrans-inducingalpha genes. Proc. Natl. Acad. Sci. USA82:5870-5874.
47. Post, L. E., S. Mackem, and B. Roizman. 1981. Regulation of alpha genes of herpes simplex virus: expression ofchimeric genesproduced by fusion of thymidine kinase with alpha gene promoters. Cell 24:555-565.
48. Preston, C. M., M. G. Cordingley, and N. D. Stow. 1984. Analysis of DNA sequences which regulate the transcription of aherpes simplex virus immediate-early gene. J. Virol. 50:708-716.
49. Preston, C. M., M. C. Frame, and M. E. Campbell. 1988. A complex formed between cell components and an HSV struc-turalpolypeptide binds to aviral immediate early gene regula-toryDNA sequence. Cell 52:425-434.
50. Sadowski, I., J. Ma, S. Triezenberg, and M. Ptashne. 1988. GAL4-VP16 is an unusually potent transcriptional activator. Nature (London) 335:563-564.
51. Sawadogo, M., and R. Roeder. 1985. Interaction of a gene-specific transcription factor with the adenovirus major late promoterupstream of the TATA box region. Cell 43:165-175. 52. Spear, P. G., and B. Roizman. 1972. Proteins specified by herpes
simplex virus. V. Purification and structural proteins of the herpesvirion. J. Virol. 9:143-159.
53. Stern, S., M. Tanaka, and W. Herr. 1989. The Oct-1 home-odomaindirects formation ofamultiprotein-DNA complexwith theHSV transactivator VP16. Nature(London) 341:624-630. 54. Stringer, K. F., C. J. Ingles, and J. Greenblatt. 1990. Direct and
selectivebindingof an acidictranscriptional activationdomain to theTATA-box factor TFIID. Nature(London)345:783-786. 55. Triezenberg, S. J., R. C. Kingsbury, and S. L.McKnight. 1988. Functional dissection of VP16, the transactivator of herpes
simplex virus immediate early gene expression. Genes Dev. 2:718-729.
56. Tsurumi, T., K. Maeno, and Y. Nishiyama. 1987. Nucleotide sequenceof the DNApolymerasegeneofherpes simplexvirus type 2 and comparison with the type 1 counterpart. Gene 52:129-137.
57. Watson,R.J. 1983. The DNA sequenceof the herpes simplex
virus type 2glycoproteinDgene. Gene26:307-312.
58. Weeks, D. L., and N. C. Jones. 1985. Adenovirus E3-early
promoters: sequencesrequired for activation byElA. Nucleic Acids Res. 13:5389-5402.
59. Werstuck, G., and J. P. Capone.1989.Mutationalanalysisof the herpes simplex virus trans-inducing factor Vmw65. Gene 75: 213-224.
60. Wu,C. 1984. Twoprotein-bindingsites in chromatinimplicated
in the activation of heat-shock genes. Nature(London)
309:229-234.
61. Xiao, P., andJ. P.Capone. 1990. Acellular factor binds tothe
herpes simplex virus type 1 transactivator Vmwl6 and is
required for Vmw65-dependent protein-DNA complex
assem-blywithOct-1. Mol. Cell. Biol. 10:4974-4977.
62. Zhang, Y.F.,and E. K.Wagner. 1987. The kinetics of expres-sion of individualherpes simplexvirus type 1transcripts.Virus
Genes1:49-60.