Vol.62,No. 12
Hepatitis B Virus Particles Contain
a
Polypeptide
Encoded
by the
Largest Open Reading Frame:
A
Putative Reverse
Transcriptase
DAVID H. MACK,' WILL BLOCH,2 NRAPENDRA NATH,3 AND JOHN J. SNINSKY1*
DepartmentofInfectiousDiseases' andDepartment ofProtein Chemistry,2 Cetus Corporation, 1400Fifty-ThirdStreet,
Emeryville, California 94608, and Blood Services Laboratories,AmericanRedCross, Bethesda, Maryland 200143
Received 28 March1988/Accepted 29 August 1988
Asegmentof thelargestopenreading frame of hepatitisBvirus (HBV)wasinserted intoanopenreading framevectordirecting the exparessioninEscherichia coli ofa fusion molecule containing 143 HBV-encoded
amino acids. The fusionprotein was used togenerate antiserum which served in immunoblots toidentify a
polypeptide with a molecular mass of 65 kilodaltons in HBV particles. Because of the small number of moleculesinvirusparticles,unambiguous detectionrequiredthedevelopmentofahighlysensitiveimmunoblot procedure.
The largest open reading frame (ORF P) of human
hepa-titis B virus (HBV) could encode a polypeptide containing
approximately 845 amino acids; its size alone led Galibertet
al. (4)tosuggestthat it is thegenefor the DNApolymerase activity typically identified in virus particles (5). The other members of the hepadnaviruses isolated from woodchucks (3), ground squirrels (10), and ducks (7) contain a similarly
sized andpositioned ORF. SummersandMason(14) initially demonstrated that duck hepatitis B virus replication
pro-ceeds through a reverse transcription step in immature
cytoplasmic nucleocapsids. Other laboratories have
ex-tended this observation to the mammalian viruses (8, 11). Accordingly, theenzyme responsible forDNA polymerase
activity inmatureextracellularparticles mayalso be ableto useRNAas atemplate. Consistentwith thisproposalwasa
report by Toh et al. (15) that a region ofthe polypeptide
predicted by ORF P shows limited but striking similarity in
amino acid sequence to the reverse transcriptases of Rous
sarcoma virus and murine leukemia virus. This report
de-scribes the cloning and bacterial expression of the itternal
segment of the putative hepadnaviral polymerase,
previ-ously shown to be conserved among elements which
repli-catethroughanRNA intermediate,as aportion ofa
tripar-titefusion protein. This proteinwasused togeneraterabbit
antiserum which permited the identification of an ORF P
geneproduct within virusparticles. Unambiguous
confirma-tion thatsuch proteins exist within virus particles,
presum-ably because of the low copy number of the proteins,
required the development of a highly sensitive detection
procedure for immunoblotted proteins.
Efforts to increase the sensitivity of solid-phase noniso-topic detection for Southern blot analysis by using horserad-ish peroxidase have benefited dramatically by the use of
tetrameth'ylbenzidine (TMB) rather than diaminobenzidine (DAB) as a substrate (12, 13; W. Bloch and D. Birch, unpublished results). We chose to determine initially the quantitative advantage thatTMBprovides in Western blots (immunoblots) since the number of copies of the viral polymerase polypeptide(s) in a virus particle could be
ex-tremely low. Twoidentical immunoblots(16)weregenerated
which contained adilution seriesofpurified aminoglycoside
phosphotransferase (3')I [APH(3')I] (Fig. 1).
*Corresponding author.
Developmentof immunoblots with DAB (Sigma Chemical
Co.)wascarriedoutaspreviouslydescribed(2). For
TMB-based detection, washed filters were agitated in fumarate
buffer (100 mM sodiumfumarate, 10 mM EDTA [pH 5.4]) (fumarate buffer) for 5 min. Blotswere then incubated with
fumarate buffer containing 1% dextran sulfate for 15 min
with agitation, followed by a 5-min wash with fumarate
bufferalonetoremovebulk dextransulfate. Next,thebuffer was removed and replaced with freshly made substrate
solution(2.5 ml ofa0.1-mg/mlTMB [ICNBiologicals] stock
in100% ethanolper50 ml of fumaratebuffer) for15min with agitation. Protein bands were visualized by the addition of
hydrogen peroxidetoafinal concentrationof0.0015%; after
development for 5 miin, blots were gently agitatedwith five
changes of distilled H20. DAB and TMB provide for the detection of 5 ng and 125 pg of electrophoresed protein, respectively. The 40-fold increased sensitivity provided by TMB over DAB allows greater dilution of primary and secondary antibodies, thereby decreasing the background caused by weak cross-reactivity to unrelated polypeptides.
1
2
345
6
7
8
A
B
_::_
:.:
....
....*: .: :::
...
_...
tF it<tWwe
FIG. 1. Comparative immunoblot sensitivity withalternate
per-oxidasesubstrates. Amounts ofpurified APH(3')l lanes 1 through 8,
30ng,15ng,5ng,625pg,125pg,25pg,5pg,and 0pg,respectively)
wereloadedontwosodium dodecylsulfate-12.5% polyacrylamide gels. Following fractionation, the gels were electroblotted onto nitrocellulose membranes (16). The blots were treated identically with a 1:3,000 dilution of anti-APH(3')I serum and a 1:10,000
dilutionof goat anti-rabbitimmunoglobulin horseradish peroxidase conjugate (Bio-Rad Laboratories). Immunoblots were developed
witheither DAB(panelA)orTMB(panel B) (see text).
4786 JOURNALOFVIROLOGY,Dec. 1988,p.4786-4790
0022-538X/88/124786-05$02.00/0
Copyright © 1988,American SocietyforMicrobiology
on November 10, 2019 by guest
http://jvi.asm.org/
[image:1.612.338.534.494.631.2]Immunoblots used to monitor patient exposure to various pathogens and to confirmthe enzyme-linked immunosorbent assays which measure antibodies to the recently identified exogenous humanretroviruses may benefit from
peroxidase-linkeddetection in which the chromogenic substrate is TMB. The open reading frame (ORF) expression vector chosen
for this study (pTACNEO 2.1) is one of a series of nine vectors specifically designed to allow expression of DNA fragments of any translational reading frame containing single-stranded termini with the sequence GATC (S. Kwok,
unpublished results). Briefly, transcription is regulated by the hybrid TAC promoter and efficient translation is pro-vided by the ribosome-binding site of the ,-galactosidase
(,B-Gal)gene andeight N-terminal codons. The ,-Gal codons are followed by a polylinker region containing a unique
BamHIendonuclease site and the entire gene for a modified 262-amino-acid APH(3')I (F. C. Lawyer, F. C. Stoffel, and D. H. Gelfand, Abstr. Mol. Biol. Yeast 1983, p. 190). In summary, these coding segments allow the production of in-frame tripartite (tribrid) proteins containing 8 amino acids
of ,-Galatthe Nterminus, the amino acids encoded by an inserted DNA fragment, and the 262 amino acids encoded by theantibiotic resistance gene at the C terminus (Fig. 2).
TheHBV(adw2) Sau3A DNAfragmentchosenfor
inser-tionintoanexpression vector has thepotentialto encode 143
amino acids corresponding to codons 502 to 644 of ORF P
(17). This fragmentcontains the region shown to be similar in amino acid sequence to the largestpredicted polypeptides of the other hepadnaviruses and reverse transcriptases of ret-roviruses (15; D. H. Mack and J. J. Sninsky, Proc. Natl. Acad. Sci. USA, in press). An additional codon is contrib-uted because the linker region of the vector encodes the sameresidue as the viral sequence at position 645(proline). Since the HBVfragment chosen for expression lacks other
uninterrupted reading frames, the production of a fusion protein of the expected size unambiguously defines the
amino acids directed by the inserted DNA as encoded by ORF P.
Protein synthesized by the parent and recombinant plas-mids with the HBV insert in the correct orientation for
expression, before and after induction of transcription with
isopropyl ,B-D-galactopyranoside, was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (6) (Fig. 3, panel A) and immunoblot analysis (16) by using rabbit antiserum to theportion of the fusion protein encoded by the
bacterial APH(3')I gene (Fig. 3, panel B). The rabbit
anti-body recognized a major polypeptide of the predicted mo-lecular mass of ca. 45 kilodaltons (kDa) before induction
(Fig. 3, panel B, lane 3) and with increased intensity after induction(Fig. 3, panel B, lane 4). The detected protein of
lower molecular mass after induction was attributed to
proteolyticdegradation. Abandmigratingatca. 29 kDa was
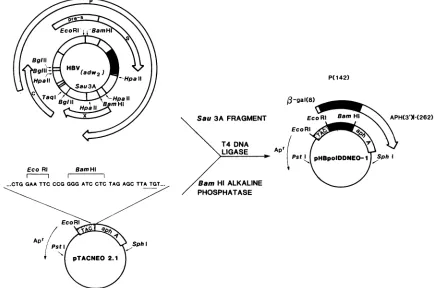
P(142)
B-gal(8)
EcoRI BamHI
m-- I
Sau3AFRAGMENT
T4 DNA LIGASE
an Hi ALKALINE PHOSPHATASE
APH(3')-(262)
FIG. 2. Vectorconstruction.Theexpressionof determinants encodedbytheORF Pregionwasdirectedby
pHBpoIDDNEO-1,
whichwasconstructed bytheinsertionofa427-base-pairSau3Afragmentcontaining putative polymerasesequences (_)intoanORFexpression
vector,pTACNEO2.1. Forthe sakeofsimplicity, onlythe HBV(adw2)genomeand thelocationof ORFs(0)identified fromsequencedata (17)aredepicted.ArrowslabeledS and Crefertothesurfaceandcoreantigengenes,respectively.Thesegmentcontiguouswith the Sgene
represents thepre-S portionoftheenvelopegeneORF.Thearrowslabeled Xand Prepresent the smallest andlargestORFs,respectively. Thebroadarrow associatedwithpHBpolDDNEO-1 represents thetribrid
P-Gal:POL:APH(3')I
protein.The thinarrowlabeled Apr denoted forpTACNEO2.1andpHBpolDDNEO-1 depicts P-lactamase. The sequencepresentedabove theparentplasmidrepresents the polylinker region betweenthetripletfortheeighthaminoacidofn-Gal(CTG)and theinitiationtriplet(ATG)forAPH(3')I.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.95.529.363.651.2]4788 NOTES
1
2
3
4
5
6
7
[image:3.612.70.549.75.264.2]PANEL
A
FIG. 3. Expression andpurificationof fusionproteins.Proteinextractsandpreparationswereseparated bysodiumdodecylsulfate-12.5% polyacrylamide gel electrophoresis. (Panel A) Protein bandswerevisualized with Coomassie blue stain.Lanes 1and3,Total bacteriallysates
of uninduced strain DG101 transformed with pTACNEO2.1(parentplasmid) andpHBpolDDNEO-1 (recombinantplasmid), respectively. Lanes 2 and 4, Lysates ofisopropyl
,3-D-thiogalactopyranoside-induced
DG101 (9) harboring pTACNEO 2.1 and pHBpolDDNEO-1, respectively. Lanes5and 6, Soluble and insolublepreparations, respectively,aftersonication of induced bacteriallysates(9) transformed with the recombinant plasmidpHBpolDDNEO-1.Lane7,Escherichiacoli-synthesized tribrid fusionprotein[0-Gal:POL:APH(3')I]
followingpreparative gelpurification (19). (Panel B) Immunoblotanalysis of gel shown inpanelA.Followingincubation withanti-APH(3')Iserum,the blotwasprobed withagoatanti-rabbitimmunoglobulin horseradishperoxidaseconjugateanddevelopedwithTMBsubstrate(see text).
also identifiedby thisanalysis from induced bacteria
harbor-ingthe parentexpressionvector(Fig.3,panel B,lane2)and
bacteriacarryingthe recombinant plasmid
(Fig.
3, panel B,lanes 3 and 4). This polypeptide is thought to be the
inefficiently expressed, truncated APH(3')I translatedfrom
the internal ATG. Neither the ca. 45-kDa or 29-kDa bands were identified bypreimmune serum (for example, seeFig.
4, panel A). After sonication andcentrifugation of bacteria carryingtherecombinantplasmid
(pHBPOLDDNEO-1),
the same twoproteinswerepresent in thepellet(Fig. 3, panel B,lane 6) butnot in the supernatant (Fig. 3, panel B, lane 5). Afterpreparative sodium dodecylsulfate-polyacrylamide gel electrophoresis purification
(19),
the tribrid fusion protein represented theonlyreadily detectable band by Coomassieblue staining (Fig. 3,panel A, lane 7).
The purified fusion protein was used to generate rabbit antiserum.With the TMB-baseddetectionsystem, the rabbit
anti-tribrid
serum clearly identified a protein from virus particle preparations migratingatca. 65 kDa(Fig.4,panel C,lane 1). The polypeptide recognized by the generated anti-serum is similar in molecular mass to one of two DNA
polymerase-reverse transcriptase activities (-70 kDa and -90 kDa) in virus particles from a differentiated human hepatoma cell linetransfected withgenomic HBV DNA (1). Bacteria harboring a recombinant plasmid that directs the
synthesis of ahybrid fusion protein containing eight amino
acids of ,-Gal at the N terminus and 262 amino acids of
APH(3')I at the C terminus (directed by pTACNEO 1.1) served as asourcefor control hybrid protein (Fig. 4, panels Athrough E,lanes 3). Neither DAB visualization of horse-radishperoxidase-linked secondaryantibody nor
autoradio-graphic
visualization
ofiodinated proteinA had thesensitiv-ityneededtodetect the ORF P-encodedpolypeptide invirus
particleswith this antiserum. This band was not recognized
by preimmune serumfrom thesame rabbit (Fig. 4, panel A, lane1) or byantiserumto
APH(3')I
(Fig. 4,panelB,lane1).Uninfected human sera prepared and probed in a manner
similar to that for sera containing HBV particles failed to generatethe ca. 65-kDa band (data not shown).
Preadsorp-tion of antiserum with various antigens in a manner previ-ously described (9)confirms that the recognition of theca. 65-kDa protein in virus particle preparations by the gener-ated antiserum is specifically relgener-ated to theORFP determi-nants within the molecule. Anti-tribrid serumpreincubated
withthetribrid proteinnolongerrecognized theviral poly-peptide(Fig. 4, panel D, lane 1), tribrid, or hybridproteins
(Fig. 4, panel D, lanes 2 and 3, respectively). Preincubation
of the anti-tribrid serum with an equivalent concentration of the hybrid protein did not block recognition of the viral polypeptide or tribrid fusion protein (Fig. 4, panel E, lanes 1 and 2, respectively) but eliminated recognition of thehybrid
protein(Fig. 4, panel E, lane 3).
Ifthe HBVORF Pregionwas transcribed and translated in its entirety, a 92-kDa polypeptide would result. The identification of a ca. 65-kDa protein suggests that the primary translation product isproteolyticallycleaved orthat the detectedpolypeptideresultsfromtranslation initiation at one of the multiple internal ATGs or is translated from a spliced mRNA. The precedent ofproteolytic maturation of the reverse transcriptase of the related retroviruses, and
preliminaryresultsby Will et al. (18)suggestingthe presence ofcore-polymerase fusion proteins in extracts of hepatocel-lularcarcinoma-containing HBV sequences, would perhaps favorproteolytic processing as the likely maturationevent. Thecentral location within the ORF of the ORF P segment expressed in the tribridproteinprevents adetermination of whether the protein identified from virus particle prepara-tions is truncated at either orbothpredictedtermini.
Prelim-inary experiments with polyclonal sera generated to the
region encompassed by codons 34 to 314 of ORF P ex-pressed as a tripartite protein suggest that the ca. 65-kDa
protein lacks the extreme N-terminal residuespredicted by
theORF(unpublished results).
Antiviral strategies designed to intercede in persistent
1
2
3
4
5
6
7
92.5-66.2
45.0
-31.0
-
21.5-14.4x
PANEL B
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
1
2
3
92.5-66.2
-
45.0-
31.0-
21.5-1
2
3
1
2
3
-_~~~~~~~~~~~~~~~~~~~~~~~~~~~~
I_
1
2
3
1
2
3
V
T
H
V T
H
V
T
H
V
T
H
V
T
H
A
B
C
D
E
FIG. 4. Immunoblotidentification ofa HBVORF P-encoded polypeptide. Lane 1 of each panel contains 20
p.l
ofpartially purified, 650-fold concentratedHBVpreparation(19). Lanes 2 and 3 of each panel contain 0.1jig
ofpurified tribrid[P-Gal:POL:APH(3')I]
andhybrid[P-Gal: APH(3')I] protein, respectively. Tripletlanes containing virus (V), tribrid (T), and hybrid (H) proteins were probed with preimmune rabbit serum(panelA), anti-APH(3')I serum (panel B), andanti-,B-Gal:POL:APH(3')I serum (panel C). Triplets (panels D and E) were treated as for panel Cexceptthat antiserum waspreadsorbed with the tribrid and hybrid proteins, respectively (16). Following incubation with rabbit antiserum, all laneswereprobed withgoatanti-rabbit immunoglobulin horseradish peroxidase conjugate and developed with TMB substrate (see text).viral infection will benefit from the characterization of pro-teinsrequired for viral replication. Sinceprecursor polypep-tides often are functionally inactive, our demonstration of the size ofthe HBV particulate polymerase (and perhaps reversetranscriptase) should assist in the eventual produc-tionofenzymaticallyactive polypeptidesinheterologous or homologous expression systems. In addition, the sequence similarity of the ORF P region used in the tribrid fusion protein to the other hepadnaviruses and the reverse tran-scriptases of retroviruses suggest that the anti-tribrid serum mayrecognize thereverse transcriptase of other viruses.
We kindly thank E. Schaeffer for critical discussions, D. Birch andR.Goodson forhelpful discussions concerning the optimization of TMB-based detection of Southern blots priortopublication, S. Kwok for providing pTACNEO ORF vectors, S. Stoffel and D. Gelfand forproviding purified APH(3')I protein and rabbit polyclo-nalanti-APH(3')I sera, S. Nilson andE. Ladnerforpreparation of the figures, andR. Bengelsdorf for excellent manuscript prepara-tion.
The research described in this report was supported by Cetus
Corporation.
LITERATURECITED
1. Bavand, M. R., andL.Orgad.1988. Twoproteinswithreverse
transcriptase activities associated with hepatitis B virus-like particles.J. Virol.62:626-628.
2. DeBlas,A.L.,and H.Cherwinski. 1983. Detection ofantigens onnitrocellulose paperimmunoblots with monoclonal antibod-ies. Anal. Biochem. 133:214-219.
3. Galibert, F., T. N. Chen, and E. Mandart. 1982. Nucleotide sequenceofaclonedwoodchuckhepatitisvirus genome:
com-parisonwith thehepatitisBvirussequence. J.Virol.61:51-65. 4. Galibert, F., E. Mandart,F. Fitoussi,P. Tiollais,and P. Char-nay. 1979.NucleotidesequenceofthehepatitisBvirus genome (subtype ayw) cloned inE.coli.Nature(London)281:646-650. 5. Kaplan,P.M.,R. L.Greenman,J.L.Gerin,R. H.Purcell,and W.S. Robinson. 1973. DNApolymeraseassociatedwith human hepatitis B antigen.J. Virol. 12:995-1005.
6. Laemmli,U. K.1970.Cleavage ofstructuralproteins during the assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
7. Mandart, E., A. Kay, and F. Galibert. 1984. Nucleotide se-quenceofaclonedduck hepatitisBvirus genome: comparison with woodchuck and human hepatitis B virus sequences. J. Virol. 49:782-792.
8. Miller, R. H., P. L. Marion, and W. S. Robinson. 1984. Hepatitis B viral DNA-RNAhybridmolecules inparticles from infected liver areconverted toviral DNA molecules duringan endoge-nous DNApolymerase reaction. Virology 139:64-72.
9. Schaeffer,E.,R.L.Snyder,andJ. J.Sninsky. 1986. Identifica-tion andlocalization of pre-S-encoded polypeptides from wood-chuck and ground squirrel hepatitis viruses. J. Virol. 57:173-182.
10. Seeger, C., D. Ganem, and H. E. Varmus. 1984. Nucleotide sequence of an infectiousmolecularly clonedgenomeof ground squirrelhepatitis virus.J. Virol. 51:367-375.
11. Seeger, C., D. Ganem, and H. E. Varmus. 1986. Biochemical andgenetic evidence for thehepatitis B virus replication strat-egy. Science232:477-484.
12. Sheldon,E., D. E.Kellogg, C. Levenson, W. Bloch, L. Aldwin, D.Birch,R.Goodson,P.Sheridan,G.Horn,R.Watson,and H. Erlich. 1987. Nonisotopic M13 probes for detecting the beta-globingene:applicationtodiagnosisof sickle cellanemia. Clin. Chem. 33:1368-1371.
13. Sheldon,E.L.,D. E.KeUogg,R.Watson,C. H.Levenson, and H. A. Erlich. 1986. Use ofnonisotopic M13 probesforgenetic analysis:applicationtoHLAclassIIloci. Proc. Natl. Acad. Sci. USA 83:9085-9098.
14. Summers, J.,and W.S. Mason. 1982.Replicationof thegenome ofa hepatitis B-like virusby reverse transcription ofanRNA intermediate. Cell29:403-415.
15. Toh, H.,H.Hayashida,and T. Miyata.1983. Sequence homol-ogybetween retroviral reverse transcriptase and putative po-lymerases of hepatitis B virus and cauliflower mosaic virus. Nature(London)305:827-829.
16. Towbin,H.,T.Staehelin,andJ.Gordon. 1979.Electrophoretic
transfer ofproteins frompolyacrylamide gelsto nitrocellulose sheets:procedureandsomeapplications.Proc.Natl. Acad. Sci. USA 76:4350-4354.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.114.525.69.253.2]4790 NOTES
17. Valenzuela, P., M. Quiroga, J. Zaldivar, P. Gray, and W. J. Rutter. 1980. The nucleotide sequenceofthehepatitis B viral genomeand theidentification ofthemajorviralgenes, p.57-70. In B. Fields, R. Jaenisch, and C. F. Fox (ed.), Animal virus genetics. AcademicPress, Inc., NewYork.
18. Will, H., J.Salfeld, E. Pfaff, C. Amnso, L. Theilmann, and H.
Schaler. 1986. Putative reverse transcriptase intermediates of humanhepatitis B virus inprimary liver carcinomas. Science 231:594-596.
19. Wong, D. T., N. Nath, and J. J. Sninsky.1985. Identification of hepatitisBviruspolypeptides encoded by the entire pre-Sopen
reading frame.J.Virol. 55:223-231.
J. VIROL.


![FIG. 4.forantiserum,(seeconcentratedAPH(3')I]serum Immunoblot identification of a HBV ORF P-encoded polypeptide](https://thumb-us.123doks.com/thumbv2/123dok_us/1328639.86773/4.612.114.525.69.253/fig-forantiserum-seeconcentratedaph-serum-immunoblot-identification-encoded-polypeptide.webp)
