JOURNAL OF VIROLOGY, Feb. 1980,p.697-707
0022-538X/80/02-0697/11$02.00/0 Vol. 33, No. 2
Distinct
Transfonration
Phenotypes Induced by Polyoma
Virus
and Simian Virus 40 in Rat Fibroblasts and Their
Control
by an Early
Viral Gene Function
BERNARD PERBAL* AND MINOO RASSOULZADEGAN Centre de Biochimie, Universitede Nice,06034 Nice, France
Several transformed cell lines established from Fisher rat cells (FR 3T3)
infected withwild-type polyoma virus orsimian virus 40 or early
temperature-sensitivemutants(polyomatsa and simian virus 40tsA30)werestudied for their
transformation phenotypes.The distinct patterns which were obtained for
poly-oma and simian virus 40 transformants led to the conclusion that these two
virusesexpressdifferenttransformingabilitiesin ratcells. Theresults obtained
withtemperature-sensitive mutant-derived transformantsindicate that all of the
transformation characteristicsstudied sofarmay beunder thecontrolof a viral
function in polyoma tsa-transformed cells.
Transformed cell lines have been established recently inour laboratory from normal diploid Fisher rat cells (FR 3T3) infected either with
wild-type (WT) polyoma virus or simian virus 40 (SV40) orwith theirearly
temperature-sen-sitive (ts) mutants (polyoma tsa and SV40 tsA30). Inboth cases, transformation by thets
viruses led to two distinctclasses of cell lines: "N" transformants, which reverted to the
nor-mal phenotype athigh temperatures, and "A" transformants, which still behaved as
trans-formed cells at the restrictive temperature (8,
15).Inpolyomatsaand SV40tsA N
transform-ants,theexpression ofatleastpartof the
trans-formedphenotype thusappears tobe underthe
control of a function involving the "large T" viralpolypeptidewhich isaffected by mutations of the A/a group of both polyoma virus and
SV40 (6,14, 16).
Except fortheirtemperaturesensitivity,tsA
and N cells appearedtobevery similartoWT transformantsintheirgrowthpropertiesatlow
temperatures.However,variations incellgrowth characteristics and in theratesof
2-deoxyglucose
uptake
andofplasminogen
activator(PA)
pro-ductionwereobserved within these classes. This variabilityin theexpression ofthe transformed phenotypemight
correspond
totheearliercon-cept of a stepwise progression toward the ulti-matetumoralstate (18). The
phenotypic
varia-bility was most apparent among SV40
trans-formants, as
expected
onthe basis of the pre-vious work of Risser and Pollack(12).
Since mostof thepolyoma
transformantsexhibited afully transformed
phenotype
for all these char-acters,one mightask whetherasignificant
dif-ferenceexists between the
transforming
abilities of the twoviruses in thissystem.Experimental resultspresented in this report
ledus totheconclusion that there is anactual difference between thespectrumoftransformed phenotypes induced by polyoma virus and SV40 in FR 3T3cells. They also indicate thateventhe
"maximal" transformation phenotypes maybe
under the control ofaviralfunctioninpolyoma
tsa transformants. Evidence is also presented that in suchcellsagenetic drift towarda heat-stable transformedphenotype is observed incell culture, which may be due to the selection of
spontaneous cellular mutationsepistatic to the viral transforming function(s).
MATERIALS AND METHODS
Cellculture. Unless otherwise specified,cellswere grown in Dulbecco-modified Eagle medium (GIBCO Laboratories) supplemented with 10% newbom calf serum (GIBCO Laboratories). They were routinely propagated at 33°C by seeding every 3rd day at a density of 5 x 104 cells per 6-cm petri plate, one transfer thus being equivalent to three to five cell generations.
The isolation of transformed cell lines has been described previously (8, 15). After three successive isolationsstartingfromisolatedcolonies,stockswere constituted afteraminimal number of transfers and kept inliquid nitrogen for further studies.
Theability of cellstogrow withoutattachmentto asolidsubstratewasassayedinbasalEaglemedium
(GIBCO Laboratories)supplementedwith 10%serum
and agaroseaspreviouslydescribed(9).
Assays for PAproduction. (i) Preparationof substrates. Purified human plasminogen wasfrom theCentre NationaldeTransfusionSanguine, Paris,
France.Thebatchusedthroughouttheseexperiments
containedonlytwopolypeptidespecieswithapparent
molecular weights ofbetween 94,000 and 96,000 as
revealedby sodiumdodecylsulfate-polyacrylamidegel
electrophoresis. Plasminogen waskept desiccated at 697
on November 10, 2019 by guest
http://jvi.asm.org/
698 PERBAL AND RASSOULZADEGAN -20°C and dissolved before use in 0.05 M borate buffer, pH 9.0, containing 0.1 M NaCl and 1 mM EDTA.
Casein (according to Hammarsten, Merck & Co., Inc.) was tritiated by a minor modification of the
proceduredescribedbyRiceand Means(10).Sodium [3H]borohydride (12.5 mCi;Commissariatal'Energie Atomique,Saclay, France)wasprogressivelyaddedto 2 mlofformaldehyde-activated casein (80 mg/mlin 0.2Mborate buffer, pH 9.0).The reactionwasstopped
byadjustingtopH2with1 NHCl;thesolutionwas
thenneutralized andextensively dialyzed against0.05 M boratebuffer, pH 9.0, containing0.1 M NaCl and1 mM EDTA.Specificradioactivities obtainedwere
be-tween0.6 and 2.5Ci/mmolofcasein,correspondingto thelabelingof 5 to 10% of thelysineresidues.
Hydrolysisof[3H]caseinwasfollowedby counting
theradioactivityof the supernatant afterprecipitation withtrichloroaceticacid(10%finalconcentration)and centrifugationfor 5minat2,000rpm.
(ii) Qualitativeassayfor extracellular PA.
As-says described in the next two sections deal with activator(s)excreted in the mediumorboundtothe extemal cell membrane or both. Theseassays were
performedwith intactcells,whereastotal PAactivity
wasmeasured incellextracts.
The firstassayis derived from the method described
byGoldberg (2).Cell cultures (usuallyasclonal
colo-niesin6-cmdishes)wereoverlaid with 4.5 ml ofamix containing0.5to1%agarose(IndubioseA37;Industrie Biologique
Frangaise),
2.5%(wt/vol)commercialnon-fat dried milk(Francelait), and 20,ugofpurified plas-minogen. For celLs growing on plastic, the culture
mediumwasremovedand thecells werewashed twice
with prewarmed phosphate-buffered saline before pouringtheagarosemix. Forthe detection of the PA
produced bycoloniesgrowing in softagar, the total volume of the overlay was reduced to 2 ml. Plates
wereexamined forclear proteolysisareasafter
over-nightincubationateither 33or400C.For thevarious
celllinestested,theextentofproteolysiswasusually
foundtobe correlated with thecellnumber and with
therateofactivatorproduction assayedquantitatively
asdescribedbelow. Thesensitivityofthe methodwas
such thatafour-toeight-cell microcolonyandpossibly
asingleproducingcellgaverise toadetectablelysis
areaafter8hof incubation.
(iii) Quantitative measurement. For assay of
extracellular PA, thecelllayerwasincubatedin the
presenceof[3H]casein andplasminogen for increasing periodsof time. Aspreviouslypointedout byRifkin andPollack(11),thekinetics of caseinolysis measured inthiswayisacomplexonesince it isinfluenced by
parameters such as the rate of secretion of PA, its stabilityinthe reactionmedium,and the interaction ofplasminwithplasminogenandactivator.
Cells were grown in 3.5-cm-diameter petri dishes
andwashedthree timeswith 5 mlof Eagle medium. Twomillilitersof thesamemedium containing25,ug ofpurified plasminogen and100 jgof [3H]casein was
addedtotheplates, whichwerethen incubatedatthe appropriatetemperature. Samples(200,uleach)were
taken at various time intervals, and the amount of hydrolyzedcaseinwasmeasured aftertrichloroacetic
acidprecipitation.Plasminogen-independent
caseino-lysiswasestimated in control assays without addition ofplasminogen.Spontaneous hydrolysisof[3H]casein
didnot exceed 0.1% under these conditions. Results were expressed as specific activities relative to the total cellprotein (counts per minute permilligram of protein). Protein concentrations wereassayedas de-scribed byeitherLowryetal. (5) orBramhall et al.
(1).
(iv) Total cell-associated PA.Cell cultures were washed threetimeswith 5 ml of 0.1 M
Tris-hydrochlo-ride buffer (pH 7.4) containing 0.01 M EDTA and lysed in1ml of thesamebuffercontaining0.5% Triton X-100(Serva)andnoEDTA.Samples (4001l each) of this extract were incubated in Tris-hydrochloride
buffer(pH7.4) in thepresenceof[3H]casein (200
jig)
andplasminogen (1 ug)for 45 minat370C. Unlabeled casein (1ml, 10 mg/ml) was addedtoeach reaction mixture, and the nonhydrolyzed casein wasprecipi-tated with trichloroaceticacid. Theextentof caseino-lysis wasestimated from the 3Hradioactivity meas-ured in 0.5 ml of the supernatant after low-speed
centrifugation.
Uptakeof2-deoxyglucose.Ratesofuptakeof 2-deoxyglucoseweremeasuredasdescribedpreviously
(8). Valueswerenormalizedasratiosof the cell-asso-ciated3H radioactivityatagivenincubationtimeto thetotalamountofproteinonthesame coverslips.
Theamountofproteinwasdetermined byremoving
the cover slips from the scintillation mixture after counting, washing with methanol, and staining with Coomassie brilliant blue forproteindeterminationby
themethod ofBramhalletal.(1).
RESULTS
Transformed phenotypes among
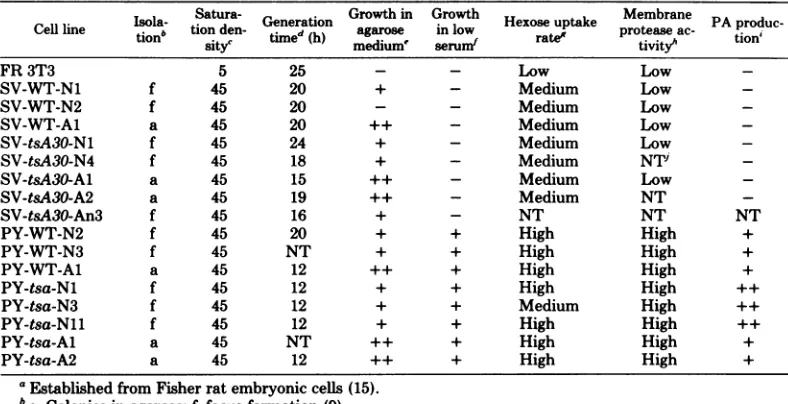
poly-omavirus and SV40derivatives. Table 1 is
a summary of the growth characteristics with andwithout anchorage and the rates of hexose uptake and PA production for 16 independently
isolatedcell lines. Eight lines were transformed
by polyoma virus (either WT or tsa) and by SV40 (either WT or tsA30). In all cases, the
transformed cells and the normal control cells (FR 3T3) were grown at330Candtested after a
limitednumber ofpassages in culture (three to eight).
All of the transformants thus far tested ex-hibitedhigh saturation densities whengrown in
Dulbecco-modified Eagle medium supple-mented with 10% calf serum. With only one exception (SV-WT-N2 cells), all ofthem grew
well in agar medium. Slightly higher plating
efficiencieshave beenrecorded for clones origi-nally selected as agar colonies, as compared with thosederived from foci (8, 15).
Incontrast to this relatively static picture, all oftheother growth and biochemical
character-isticslisted in Table 1 were found to discriminate
betweenatleasttwoclasses of transformed phe-notypes.
(i) Generationtime. As previously reported
on November 10, 2019 by guest
http://jvi.asm.org/
PHENOTYPES OF POLYOMA AND SV40 TRANSFORMANTS 699
TABLE 1. Summary of the results oftransformation assays performed on various derivatives from the FR 3T3cell linea
Cellline Isola- Satura- Generation Growthin Growth Hexose uptake Membrane PA produc-Celllie tionb tionden
timed
(h) agarose inlow rate proteaseac- tion'sityc medium' serum r
tivit3th
FR 3T3 5 25 - - Low Low
-SV-WT-Nl f 45 20 + - Medium Low
-SV-WT-N2 f 45 20 - - Medium Low
-SV-WT-A1 a 45 20 ++ - Medium Low
-SV-tsA30-Nl f 45 24 + - Medium Low
-SV-tsA30-N4 f 45 18 + - Medium NTi
-SV-tsA30-Al a 45 15 ++ - Medium Low
-SV-tsA30-A2 a 45 19 ++ - Medium NT
-SV-tsA30-An3 f 45 16 + - NT NT NT
PY-WT-N2 f 45 20 + + High High +
PY-WT-N3 f 45 NT + + High High +
PY-WT-A1 a 45 12 ++ + High High +
PY-tsa-Nl f 45 12 + + High High ++
PY-tsa-N3 f 45 12 + + Medium High ++
PY-tsa-Nll f 45 12 + + High High ++
PY-tsa-Al a 45 NT ++ + High High +
PY-tsa-A2 a 45 12 ++ + High High +
aEstablished from Fisherratembryonic
cells
(15).ba,Colonies in agarose; f, focus formation (9).
cCell densitiesatthegrowth plateau.
Cells
x104 per square centimeter.d Measured inDulbecco-modified Eagle mediumsupplemented with10%
newborn
calf serum (15).'Measuredby seeding5x
104 cells
in soft agar andcountingthecolonies8dayslater(9). Growthability in agarosewasscoredasfollows: ++,morethan 75% of the cellinput gave risetocolonies; +, 25%; -, less than 0.002%.fSymbols: -, slow growth and low saturationdensity similartothose observed in the presence ofhighserum concentrations (15); +, generation time and maximal cell density similarto those observed at high serum concentrations.
'Explanation ofterms:low, values similartothat shown for FR 3T3cells (Fig. 2); medium, values in the samerangeasthat shown forSV-WT-A1;high, values similartothat shown for PY-tsa-Nl.
hHydrolysis of[3H]casein in theabsence ofplasminogen after8hof incubation at 33°C (see text). Low, Slighthydrolysis(verysimilartothat of FR3T3); high,significantlyhigherrateofhydrolysis (can reachavalue which is10to15% that obtained in the presence of plasminogen).
'Symbols:++,positive reactionby the agaroverlayassayandrateof[3H]casein hydrolysissimilartothat obtainedforPY-tsa-N1cells(Fig.5); +,rateof[3H]caseinhydrolysisin thesamerangeasthatshown for PY-tsa-Al cells(Fig.5); -,nodetectablecaseinolysis byeither assay.
iNT, Not tested.
(15), all polyoma transformants tested except
one(PY-WT-N2) exhibited generation times of near 12 h at 33°C in Dulbecco-modified Eagle
medium supplemented with 10%calfserum, as
compared with 25h forFR3T3cellsunder the same conditions. SV40 transformnantsappeared
tobesignificantly different in this respect,as all
of themgrewwithdoublingtimesrangingfrom
15 to 24 h. This property of the established
transformants is in agreement with the
obser-vation that, after virusinfection, theappearance of visible foci or agar colonies requires 5 to 6
weeksat33°C for SV40,ascomparedwith 2to
3weeks forpolyomavirus.
(ii) Growthatlowserumconcentrations.
Severaltransformed cell lines and theFR 3T3
parentalline weregrownin mediumcontaining 2% calfserumuntiltheyreached agrowth
pla-teau. As previously reported (15), the FR 3T3
line attainedacell
density
of106
cellsper 6-cmplate
and then ceased to divide. On the otherhand,
as shown inFig.
1, SV40- andpolyoma
virus-transformedcells grew
beyond
theconflu-ence state to reach
higher
saturation densities (4x106
and7x106
cellsperplate,
respectively).
(iii)
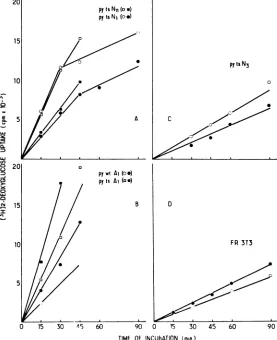
Rateof2-deoxyglucose uptake.
Cellswereseededoncover
slips
atalowdensity
and tested for therate of hexoseuptake during
ex-ponential growth
at33°C.
Inall cases, kineticsoftransportwereobserved toremain linearfor
atleastthefirst30minof incubation.
Results obtained with six
representative
celllinesareshownin
Fig.
2.Foreverycellline,
therate of
2-deoxyglucose
uptake
wasincreased ascompared
with thatofnormalFR 3T3cells. Two classes oftransformantscould, however,
bedis-tinguished:
afirst group,exemplified by
thePY-tsa-Al and
-Nil
lines,
was characterizedby
a VOL. 33,1980on November 10, 2019 by guest
http://jvi.asm.org/
700 PERBAL AND RASSOULZADEGAN
3~~eS ,~.m'".,.3
FIG. 1. Typical saturation density ofFR3T3, SV-tsA30-Al,andPY-tsa-Al cell lines.Cells(5x104/60-mm petri dish)wereseeded andgrownfor 15daysin the presenceof2%ocalfserum. Thegrowth mediumwas changedeveryotherdayatthebeginninganddailyat the endoftheexperiment. Monolayerswerestained with Giemsa(Rhone-Poulenc, Lyon,France).
°
15 vE/
C\j =5
0
v15 30 45 60
Time of incubation (min)
FIG. 2. Kineticsof 2-deoxyglucose uptakein
nor-mal andtransformedFR 3T3 cells. Cellswereseeded on coverslips, grown at33°C, and assayed for 2-deoxyglucoseaspreviously reported (8).Lines repre-sent the average ofseveral independent
measure-ments. Symbols: A, PY-tsa-Al; V, PY-tsa-Nll; *,
SV-tsA30-Nl;x,SV-tsA30-Al;0,PY-tsa-N3;O, FR
3T3 normalcells.
rate of uptake at least five times higher than that of the normal cells, whereas in cell lines
such as PY-tsa-N3, tsA30-N1, and
SV-tsA3O-Al, these valueswereonly increased bya factor of1.5 to 2.
As shown in Table 1, all of the SV40-trans-formed lines tested exhibited a relatively low
rateofuptake, whereassevenof eight polyoma transformants behaved like the PY-tsa-A1 and
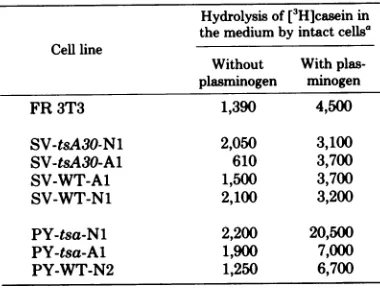
TABLE 2. External proteases and PA in FR 3T3 cells and variousrepresentative transformants
Hydrolysis of[3H]caseinin the mediumby intactcellsa Cellline
Without With plas-plasminogen minogen
FR 3T3 1,390 4,500
SV-tsA30-N1 2,050 3,100
SV-tsA30-A1 610 3,700
SV-WT-A1 1,500 3,700
SV-WT-N1 2,100 3,200
PY-tsa-Nl 2,200 20,500
PY-tsa-A1 1,900 7,000
PY-WT-N2 1,250 6,700
Therateofhydrolysisof
[3H]casein
wasmeasured asindicated in thetextin cells grownat33°C;values represent the ratios ofacid-soluble 3H radioactivityafter6hof incubationat330Ctothe total cellprotein.
-Nil
lines(PY-tsa-N3
being theonly exceptionobservedamong this set of polyoma
virus-trans-formedlines).
(iv)
Surface proteases and external PA.Hydrolysisof
[3H]casein
(activityreleased in themediumorbound to the
external cell
membrane orboth[see above]) was measured either in the presence of plasminogen (assaying both PA and nonspecific protease activities) or in its absence(protease activity). The plasminogen-independ-entactivity was slightly increased in most
trans-formantsascompared with that of FR 3T3 cells. Theplasminogen-dependent activity was, how-ever, again clearly different in the two sets of transformants; whereas
all
SV40transformants
exhibitedlow levels of activity, as did the origi-nal normal
cells,
significantly higher values wereobserved for polyoma
virus-transformed
lines(Tables1and 2). In addition, whencolonies were tested by the qualitative casein overlay assay, none of the SV40 transformants induced a de-tectable caseinolysis, whereas
all
of the polyomaon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.514.120.410.83.191.2] [image:4.514.88.233.247.495.2] [image:4.514.269.459.264.408.2]PHENOTYPES OF POLYOMA AND SV40 TRANSFORMANTS 701
transformants were clearly positive (data not
shown).
Withonly fewexceptions, the 15independent
transformants tested so far may therefore be
distributedamong twoclasses of transformation phenotypes.Afirstgroupof lines ("low
transfor-mation levels") wascharacterized by relatively long generationtimes,poor growth atlowserum
concentrations, amoderate increase in 2-deoxy-glucose uptake, and no detectable increase in PA production. A second group ("maximal transformation") wasclearly different from the normal phenotype for all of these characters. The fact that the spectrum oftransfornation phenotypes inducedbypolyoma virus in FR 3T3
rat fibroblasts differed
significantly
from the widely distributedrangeof transformation levels inducedby
SV40 (12) will be further docu-mentedbelow.Resultsreportedsofar also indicate that the
E
4-
I-0
C)
0X
r
transformation levelsobservedin various trans-formantsare notrelated in any simple way to
the selection procedureor to the A and Ntypes
among tsa- andtsA30-transformedcells. It was
of interesttodeterminewhether in thetsNlines
which expressed the maximal transformation levels (polyoma tsa-Nl and -Nil), all of the
characters assayedare temperaturedependent. Temperaturedependence of the saturation
den-sity of these lines,aswellastheir growthrates
and ability to grow in low serum and in agar
medium, has beenreported previously (15). Temperature sensitivity of hexose
up-take
andof PAproduction
inpolyoma tsa-N transformants. (i) 2-Deoxyglucose up-take. Cellsweregrownateither33or410C,
and kinetics of[3H]2-deoxyglucose
uptake weremeasured at both temperatures. As shown in Fig. 3, the initialrates of hexose uptake were
decreasedby about 50% in tsa-Npolyoma
trans-pytsN3
D
FR3T3
15 30 45 60 90
TIME OF INCUbATION (min)
FIG. 3. Thermosensitivityof 2-deoxyglucoseuptakeintypeN
transformed
celllines. Therateof
deoxyglu-coseuptakewasmeasuredaspreviouslydescribed(8)ateither33°C (open symbols)or41°C(closedsymbols)
forall cell lines tested. VOL. 33,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.514.121.398.299.639.2]702 PERBAL AND RASSOULZADEGAN
formants grown at
41°C
relative to a parallel culture grown at33°C,
whereasa 1.5- to2-fold increase was observed for FR 3T3cells,
tsa-Atransformants,
and WTpolyoma
transformants.A similar observation was
reported
previously forSV40tsA30-transformed
cells(8).Asshown inFig. 4, whenPY-tsa-N1cells were
shifted from 33 to
410C,
their rate of uptake dropped quickly and within 4 h after thetem-perature shiftwasdown tothe level measured under thesameconditions in culturesgrown at 410C (Fig. 3A,30min).
(ii) PA. As shown inFig. 5,
production
of PAasmeasured
by
theplasminogen-dependent
hy-drolysis of[3H]casein
wasfoundtobetempera-turesensitiveinPY-tsa-N1cells.Thesame
con-clusion was reached
independently (data
notshown) from
qualitative
overlay
assays on iso-lated colonies. Since these assaysonly
reflect the presence of PA on the outside of the cells (membraneandmedium), similarmeasurements20
pyLs Ai
n 15_
AiJ
_.
/
_
10D
0
o pyLsNi
w
CsJ
5-0 2 4 6 8
TIME (hours)
FIG. 4. Temperature-shift experiments with PY-tsa-Nl andPY-tsa-Al cell lines. Cells were seeded onplastic cover slips(Thermanox; Lux Corp) ata density of 5x 104cells percover slip (9 by 35 mm). They were grown inDulbecco-modified Eagle me-dium (GIBCO Laboratories) buffered with 50 mM HEPES
(N-2-hydroxyethylpiperazine-N'-2-ethane-sulfonic acid; Merck & Co., Inc.)for 70 h at33°C. Cover slips were then shifted to the restrictive tem-perature (41IC) forincreasing periods of time. The rate of2-deoxyglucose uptake was measured by in-cubating the cover slips for 30 min under the condi-tions described previously (8).
were
performed
on cell extracts to determine whether a transportmechanism,
rather thansynthesis by itself, might
be thetemperature-sensitive
rate-limiting step.
Triton X-100 extracts were prepared as
de-scribed abovefrom cellsgrownatboth
temper-atures andshifted from 41 to330C
for variousperiods
oftime. PA activities in such extracts werefoundtobeabout 10-fold higher thantheactivitiesmeasured
previously
in theintactcells(Table 3),
whichmayreflecteitherthepresenceof
large
amountsofinternalPA or anactivation process in the presence of Triton X-100. Thespecific activity
of extracts from tsa-N1 cellsgrownat
410C
was20-to30-foldlowerthan that ofsimilar extractspreparedfrom cellsgrownatlowtemperatures.Upon temperatureshiftsfrom
41 to
330C,
theactivity
wasfully
restored withina short period oftime (less than 2.5 h). It is
therefore
likely
that the temperature-sensitive viralfunctionacts at acontrollevelinthe intra-cellularsynthesis
of PA rather than on atrans-port mechanism.
Preliminary
resultsfrom simi-lar experiments performed in the presence ofcycloheximide suggest that the appearance of PA activity upon a temperature shift-down is
dependentonactiveproteinsynthesis andmight therefore representaninduction of denovo
syn-thesis of PA.
(iii) Uncoupling
oftemperaturedepend-ence oftransformation
characteristics
in tsa-Ntransformants after high numbersofpassages in culture. A fully thermosensitive phenotype wasalso obtainedwith PY-tsa-N11
cells when these cells were tested soon after
their isolation. However, when cells at
higher
passage numbers were examined, they were
foundtostillbetemperaturedependent for their
rate of2-deoxyglucose uptake, but to produce
high
levelsofPAirrespective of the temperature (datanotshown). Itthereforeseems likelythat during successive generations in culture, var-iants expressing this transformation character undera different control may be selected. To furtherdocumentthisescape fromthetemper-atureviral control, wegrew early passage PY-tsa-Nl cells at330C forlarge numbersof gen-erationsandtested thematvarious
timnes
fortheamount of PAreleasedin themediumateither
33 or 410C. As shownin Table 4, the
41/330C
ratios increased progressively to a point (130 generations)atwhichthetemperatureeffectwas barelysignificant. Further studies arerequired
to determine whether clones which have lost
theirtemperatusensitive behavior contain
re-vertant viralgenomes.This does notseem
likely,
however, in view of the observation reported above that hexose uptakeandcloning efficiency in agar remain thesame as in theoriginaltsa-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.514.93.235.309.548.2]PHENOTYPES OF POLYOMA AND SV40 TRANSFORMANTS
pytsa Ni pytsa Ai
C3
Cl.. o 7v E5 A
C 0
2 4 60 2 4 6
Tm.c of incubation (houlrs)
FIG. 5. PAsynthesis by normal andtransformed FR 3T3 cells incubated at either 33 or41°C.Normal cells, polyoma transformants, andSV40transformants were grown at33°Cuntil theyreached a density of about 2
x 104 cells per 35-mm plastic petri dish. Some plates were left at33°C, whereas others were shifted to the restrictivetemperature (40 to410°C) for 24 to 48 h. Cellswere washed three times with 5 ml of prewarmed Dulbecco-modified Eagle medium without calf serum; 2 ml of medium containing 25 piofplasminogen and 100 pgof[3H]casein was added to the plates. At the indicated times, the amount of [ H]casein hydrolyzed was measured in a 2(X-ylsample. The proteinconcentration was measured after incubation of the cells in the presence of 0.2 N NaOH (1 ml). It was checked each time that the plasmin generated during the incubation
did not affect the cell morphology. Symbols: open,33°C;closed,41°C.
TABLE 3. Temperaturedependence of PA activity inextractsfrom PY-tsa-NI cells
~~con~tio~ Hydrolysisof Overlay Culture conditions
[3H]caseina
assayb33°C,2days 420,000 ++
41C,2days 15,000
41and33°C,0 h 17,000
41and 33°C,2.5h 370,000 +
41and33°C,4.5h 496,000 ++
41and33°C,7h 300,000 ++
aTriton X-100extracts werepreparedasdescribed
in the text. Theactivity is expressed as [3H]casein
hydrolyzed (counts per minute) permilligramof pro-tein in theextract.
bCellsweregrownat
330C
and incubatedfor48h at41°C. Plateswerethen shiftedto33°Cforincreasing periodsof time andincubated backtothe restrictive temperature to perform the qualitative assayas de-scribed inthelegendto Fig. 1.See Table 4, footnote b, forexplanationofsymbols.Ni andtsa-Nllcelllines
(data
notshown). Frequencyofmaximally
transformedde-rivatives after mass infection of FR 3T3 cells with either
polyoma
virus or SV40. Among the set of 16independent
transformed lines testedatthispoint
(Table 1),
mostpolyoma
transformants exhibited maximal
transforma-TABLE 4. Loss of temperature sensitivity of PA production inPY-tsa-NIcellsupon continuous
growth in cell culture
[3H]caseinhydrol-
[3H]2-ysis PA
deoxyglu-No. of Temp over- cose
up-gener- (OC) Activity 41°C/ lay as- take
ations inex- 330C Sayb (41Cra tractsa ratio
33iCora
5 33 300,000 0.03 ++ 0.5-0.6
5 41 10,000
10 33 260,000 0.10 ++ 0.5-0.6
10 41 28,000
80 33 223,000 0.45 ++ NT
80 41 102,000 +
130 33 212,000 0.66 ++ 0.5-0.6
130 41 140,000 +
aTotal PA measuredin Triton X-100 extracts
ex-pressedascountsperminutepermilligram ofprotein.
bPerformedas in Fig. 1. Symbols: ++, clear large
halos after 8 h ofincubation; +, need to wait 24 to 48 h to seeclearhalos; -, no halos within 36 h of incu-bation.
CMeanratio established from severalindependent measures(30-niinuptake). NT,Not tested.
tionlevels, whereas allof the
SV40-transformed
lines,although showinghighsaturationdensities and the ability to grow in agar medium,
ex-hibited relatively low levels of hexose uptake
VOL. 33,1980 703
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.514.132.372.78.286.2] [image:7.514.260.451.409.552.2]704 PERBAL AND RASSOULZADEGAN andnodetectable PAproduction. To determine
whether this observation reflectedanactual
dif-ferencebetween thetransforming abilitiesofthe twoviruses, rather thanarandomphenomenon
due to the relatively small number of clones analyzed, weused the qualitativeoverlayassay for PA production (see above) as a means for
establishing the proportionsofmaximally
trans-formed derivatives in a statistically significant way.
Parallelcultures of FR 3T3cells wereinfected
witheither WT polyoma virus orWT SV40 at
identical multiplicities (250 PFU/cell), and transformants were selected at 33°C either as
fociovergrowing attached monolayersoras
col-onies in agarose medium, under the conditions
previously used forestablishing the transformed cell linesdescribed above(8, 15). After the plates
were scored for the number of transformants,
both the attached cultures and the agarplates
were overlaid with the casein-plasminogen-aga-rosemixture, and the number of PAproducers wasdetermined after either 12-hor2-day
incu-bationsat33°C. The numberof haloswas never
observedtochange with the incubation time. As shown in Table 5, the frequencies of PA-producing clones obtainedamongthe
transform-ants were quite different after SV40 and poly-oma virus infections. The lowest values were
observed among transformants selected as foci
afterSV40infection; in thiscase, thefrequency
wasalso dependentontheselectionprocedure, ashigher proportions of PA producerswere
ob-served among the colonies grown in agar
me-dium.Highfrequencieswereobservedwith pol-yomavirus among both foci and agarcolonies.
These frequencies were not found to be de-pendent on themultiplicity of infection (0.1 to
250 PFU/cell), an observation reinforcing the conclusion that there is an actual difference
between the transforming abilities of these two viruses which cannotbe accounted for by acci-dental variations in the titers of the virus stocks. Table 5 also shows the results of an
experi-mentinwhichanonproducingestablished SV40
transformant (SV-tsA30-A2) was infected with either polyoma virus or SV40. "Supertrans-formed" PA producers could be observed, again
atagreaterfrequency with polyoma virus than
with SV40. These clones also grew faster than SV-tsA30-A2 cells, since they appeared as dis-tinct foci on the layers of transformed cells grown onplastic platesat33°C.
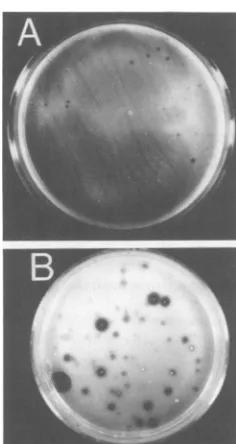
Withpolyoma virus, thediameter of the
cas-einolysis area wasnotrelatedtothesize ofthe colony, whereas the halo sizeswerealwaysmore
uniformly distributed and correlated with the diameter of the colony for SV40 transformants (Fig. 6). This observation suggests thatPA pro-duction insomecases,butnotinothers,maybe
correlated with the growth rate of the cells in suspension.
DISCUSSION
A number of independently isolated rat cell
lines transformed by SV40 and polyoma virus
werestudied for their transformation phenotype
incell culture.These lineswereestablished un-deralimited number ofdefinedconditions (cell
growth,selection procedure,useof either WTor early tsmutants,etc.) andtherefore offered an
approach for the identification of possible cor-relations between transformation characteris-tics. Their phenotype wassystematically com-pared with theparental diploid established FR 3T3 lineatearlypassagesincell culture.
Theresults obtained ledustothe operational distinction between a maximally transformed
[image:8.514.69.465.516.646.2]state and several intermediate phenotypes as
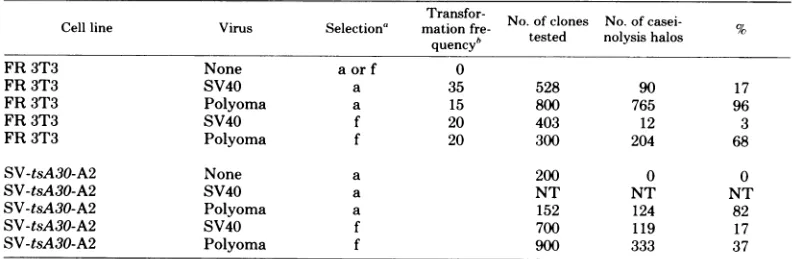
TABLE 5. Frequency of PA-producingtransformants after polyoma virus and SV40 infections of normal rat cells(FR3T3) and nonproducing SV40-transformed cells
Transfor- No of clones No. of
casei-Cell line Virus Selection' mation fre- tse oyi ao
quencyb tested nolysis halos
FR 3T3 None aorf 0
FR 3T3 SV40 a 35 528 90 17
FR3T3 Polyoma a 15 800 765 96
FR 3T3 SV40 f 20 403 12 3
FR 3T3 Polyoma f 20 300 204 68
SV-tsA30-A2 None a 200 0 0
SV-tsA30-A2 SV40 a NT NT NT
SV-tsA30-A2 Polyoma a 152 124 82
SV-tsA30-A2 SV40 f 700 119 17
SV-tsA30-A2 Polyoma f 900 333 37
aa,
Colonies in
agarosemedium;
f,focus formation (15).bNumber oftransformants per104infected
cells.
on November 10, 2019 by guest
http://jvi.asm.org/
PHENOTYPES OF POLYOMA AND
SV40
TRANSFORMANTS705
I
FIG. 6. PA-induced caseinolysis by WT SV40 transformants(A)andby WTpolyoma transformants (B)growninagarmedium.
previously reported for polyoma virus-infected
hamstercells(18) and for SV40-infectedmouse
cells (12). Thus, all polyoma virus-transformed
ratcellson oneside exhibiteda
well-character-ized transformed phenotype for any property which was assayed (growth characteristics on
solid substrate withhighor lowserum
concen-trations, abilitytogrowwithoutanchorage,rate
of hexose uptake, and production of PA), whereasall of the SV40 transformants studied expressed intermediatelevels of transformation.
Although all transformantsshowedanincreased rate ofdeoxyglucose uptake, noneof theSV40
transformants produceddetectable amounts of
PA. A similar dissociationof PA excretion from
othertransformation characteristicshasalready been reported in some Rous sarcoma
virus-transformed cells (19). Thelatter cells showed
levels of PA similar to that of the uninfected
cells, although theyexhibitedseveralproperties of transformed cells, including increased sugar transport, altered cellularmorphology, high
ef-ficiencyofgrowthinagar,andhighincidenceof
wing webtumors (20). Thesecretion of PA
in-hibitors bytransformed cellshas also been re-ported in severalcases(4, 13). Ourpreliminary results obtained with SV40-transformed cells
havenotrevealed the detectionofany PA
inhib-itor. The lack of PA secretion by SV40
trans-formants, togetherwith the intermediate
trans-formationphenotypeobserved forgrowthin2%
serum,growthinagar,and rate ofdeoxyglucose
uptake, may reflectdifferences in the transform-ing abilities of SV40 and polyoma virus. Al-though conflicting results have been reported regarding the correlation between tumorigenic-ityandPAproduction (3, 4, 7, 13),the
compar-isonbetween the different phenotypes observed
in culture and the ability to induce tumors in animals has been undertaken. Preliminary
re-sults indicate that the polyoma transformants
induce tumor formation in various animals (young Fisher rats,nudemice, and chicken
cho-rioallantoid membrane), whereas the SV40 transformantsaremuchless tumorigenic under
the same conditions (M. Rassoulzadegan, F. Birg, G. Meyer,andF. Cuzin,unpublisheddata).
The frequency ofPA-producing transformants
was measured after polyoma virus and SV40 infectionof FR 3T3 cells. Theresults obtained clearly showthat these two virusesinduce
dif-ferent patterns of transfornation in rat cells.
After polyomavirusinfection,96% of the trans-formantsselectedinagarosewere PAproducers, whereas only17% of the coloniesobtained under
thesameconditionswithSV40wereproducers.
Itseemsthat agarose selection leads to ahigher proportion ofPA producersamong both SV40
and polyoma transformants as compared with the selectionoffoci growingonplastic (Table 5).
It isthen possiblethat growth in agarsuspension requiresa"moretransfornedstate" than is re-quired forgrowth as foci on plastic. The super-infection of a nonproducer SV40 tsA30
trans-fornant with WTSV40 and WT polyoma virus shows that the existence of a viral genome
(SV40) in thetransformed cellsdoes not prevent
the expression ofanother since supertransfor-mationby WTpolyoma virus resultsin a much
higherfrequency of PA-positive colonies (38%)
than is observed when the cells are
retrans-formed with WT SV40 under the same
condi-tions(17%).
It therefore appears that different levels of
transfornation areinduced by SV40and poly-omavirus.Whether thesedifferencesarerelated
inmolecularterms, to the organizationofviral sequencesincells,or totheexpression of partic-ularviralproteins remainstobeestablished. It has been reported previously that ratcellsare
nonpermissive for SV40 replication, whereas
they aresemipermissive forpolyoma virus
rep-lication (reviewedinreference 17). Itwould be
of interest to determine whether the transfor-mation levelswhich are reported in this study
arerelatedtodifferentlevelsofcellular permis-sivity.Inany case, the distinct nature of thetwo
viruses appears in ourhands to be one of the criticaldeterminants of the
variability
observed among thetransformedphenotypes.Twoclasses oftransformants have been
ob-VOL. 33,1980
1W.. I
00
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.514.97.215.81.303.2]PERBAL
TABLE 6. Summaryofthethermosensitivetransformationcharacteristics in PY-tsa-NI cells
Saturation Rate of[3H]casein
hy-Growth densityin Genera- Platingeffi- Rateof 2- drolysis' Production
CellsCt(eOmcptm containingmeim timbetion agaroseciencyme-
mn
deoxyglu-ofP
Without (overlay)10% calf se- (h) diumC minogen plasmino-
assay)f
rumi gen
PY-tsa-Nl 33 >10 12 14 14,000 20,500 2,200 +
41 1.5 22 <0.01 5,500 3,400 2,700
FR 3T3 33 5 25 <0.002 2,000 4,500 1,800
41 5.1 24 <0.002 2,500 4,800 1,800
a
Cells
per 6-cmplatex 10'.bFromSeif and Cuzin(15).
cExpressedasthe percentage of the
cell
input (5x104
cells)growinginagarose medium. d Measuredaspreviously
reported (8).'Expressedas countsperminute per6hof incubation per
milligram
ofprotein.fSymbols: +,clear halos after8h ofincubation;-,nodetectablehydrolysisafter 36 h of incubation.
tained with either SV40orpolyomavirus:
trans-formed celllines oftypeN were found toregain the normal phenotype at high temperatures,
whereastypeAcell linesstillexpresstheir
trans-formation characteristics at the nonpermissive
temperature (8, 15). The different phenotypes described with the present set ofcell lines did
not permit a differentiation between A and N
transformants (polyoma tsa and SV40
tsA30).
However, recent studies led to the conclusion
that theproductionof PAmaybesusceptibleto
the growth conditions applied toinfected cells
during the selection of transformed cells (B. Perbal,manuscriptinpreparation).
Allof thetsNcell lineswerefoundtoexpress their transformationphenotypeat33°Cbut not
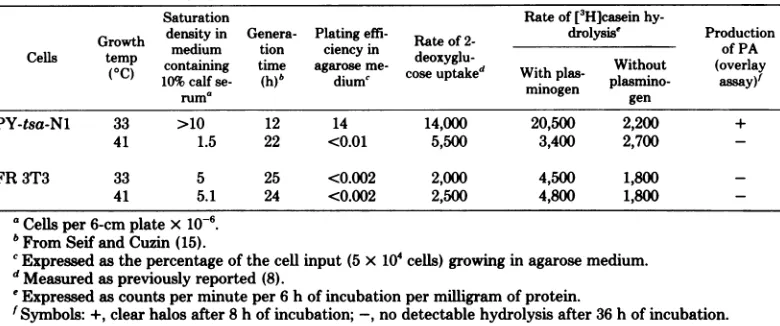
at 40to41°C. As shown in Table 6 for
PY-tsa-Ni cells,the rates of bothdeoxyglucose uptake and PA production reverted with generation time, saturation
density,
andplating
efficiencyinagarandonplastictothe valuesobtainedfor
FR 3T3 cells. Thus, the
expression
ofa wholesetofvarious transformationcharacteristics ap-pears to be dependent upon the
expression
ofthe viral functions defined
by
tsa(and tsA30)
mutations.Temperature-shift
experiments
show that the transition from one phenotype tothe othertakesplacewithin a shortperiod oftime. The drifts of PA production and cloning effi-ciencyonplastic,which werereportedabove forpolyomatsN cellsgrown for severalgenerations,
suggest that their control is unlikely to be a
simple oneandprobably involves bothcellular
and viral factors as previously reported with Rous sarcomavirus-transformed cells(20).
ACKNOWLEDGMENTS
We thankF.Cuzin for helpfulcriticism of this work and F. Birg and G. Fareed forthe critical reading of themanuscript. The technicalassistance of C. Bonifacino and L.Carbone is
gratefully acknowledged. We also thank M. Ducasse, L. Fer-aud, and R. Rahmani fortheir kind help in the study of the PY-tsa-Nl cell line and to Francelait S. A. for the generous gift of nonfat driedmilk.
This work wasmade possible by grants C. R. L. 78.1.083.1 and AT79-114from the Institut National de laSant6 et de la Recherche Medicale.
LITERATURE CITED
1. Bramhall, S., N. Noack, M. Wu, and J. R. Loewen-berg. 1969. A simple colorimetric method for determi-nation ofprotein. Ann. Biochem. 31:146-148. 2. Goldberg, A. R. 1974. Increased protease levels in
trans-formed cells:acaseinoverlay assay for the detection of plasminogenactivator production.Cell 2:95-102. 3. Jones, P. A., W. E.Laug,and W.F. Benedict. 1975.
Fibrinolyticactivity in a human fibrosarcomacell line andevidence for the induction of plasminogen activator secretionduringtumorformation.Cell6:245-252. 4. Loskutoff, D., and T. S. Edgington. 1977. Synthesis of
a fibrinolytic activator and inhibitor by endothelial cells.Proc. Natl. Acad. Sci. U.S.A. 74:3903-3907. 5. Lowry,0.H., N. J. Rosebrough, A. L. Farr, and R. J.
Randall. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265-275.
6. Paucha, E.,R.Harvey, R. Smith, and A. E. Smith.
1977.EMBO-INSERM Workshop on Early Proteins of
OncogenicDNAViruses, p. 189-198. Institut National de laSante et de la RechercheMedicale,Paris. 7. Quigley, J. 1979. Phorbol ester-induced morphological
changesintransformed chick fibroblasts: evidence for directcatalytic involvement ofplasminogen activator. Cell 7:131-141.
8. Rassoulzadegan, M., B.Perbal, and F. Cuzin. 1978. Growth control in simian virus40-transformedratcelLs: temperature-independent expression of thetransformed phenotypeintsAtransformants derived by agar selec-tion. J. Virol. 28:1-5.
9. Rassoulzadegan, M., R. Seif, and F. Cuzin. 1978. Con-ditionsleading to the establishment of the N (a gene dependent) and A (a gene independent)transformned
statesafterpolyoma virus infection of ratfibroblasts. J.
Virol.28:421-426.
10.Rice, R. H., and G. E. Means. 1971. Radioactive labeling ofproteinsinvitro. J.Biol. Chem. 246:831-832. 11. Rifkin, D. B., and R. Pollack. 1977. Production of
plas-minogen activator by established cell lines of mouse origin. J. Cell Biol. 73:47-55.
12. Risser, R., and R. Pollack. 1974. A non-selective analysis
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.514.68.458.94.257.2]PHENOTYPES OF POLYOMA AND SV40 TRANSFORMANTS 707
ofSV40 transformation ofmouse3T3cells. Virology 59:477-489.
13. Roblin, R. O., P. L Young, and T. E. Bell. 1978. Concomitant secretion bytransformed SVWI38-VA-13-2RAcells ofplasminogen activator(s) and substance(s) whichpreventtheirdetection. Biochem. Biophys. Res. Commun. 82:165-172.
14. Schaffhausen, B. S., J. E. Silver, and T.LBenjamin.
1978.Tumor antigen(s) incellsproductively infected by
wild-type polyoma virus andmutantNG-18. Proc. Natl. Acad. Sci. U.S.A. 75:79-83.
15.Seif, R., and F. Cuzin. 1977. Temperature-sensitive growthregulation inonetypeoftransformedratcells induced by thetsamutantofpolyomavirus.J. Virol. 24:721-728.
16. Smart,J.E., andY. Ito. 1978. Threespeciesofpolyoma
tumorantigens sharecommonpeptides probablynear
the amino termini of the proteins. Cell 15:1427-1439. 17.Tooze, J. 1973. The molecular biology oftumorviruses. ColdSpring Harbor Laboratory, Cold SpringHarbor, N.Y.
18.Vogt, M., and R. Dulbecco.1963.Steps in the neoplastic transformation of hamsterembryo cells by polyoma virus. Proc. Natl. Acad.Sci. U.S.A. 49:171-179. 19.Wolf, B. A., and A. R. Goldberg.1976.
Rous-sarcoma-virus-transformed fibroblasts having lowlevelsof plas-minogen activator. Proc. Natl. Acad. Sci. U.S.A. 73: 3613-3617.
20.Wolf, B. A., and A. R. Goldberg. 1978. Lack of
corre-lation between tumorigenicity and level of plasminogen activatorinfibroblasts transformed by Rous
sarcoma-virus. Proc. Natl. Acad. Sci.U.S.A. 75:4967-4971. VOL. 33, 1980