JOURNAL OF VIROLOGY, Dec. 1975, p. 1444-1452 Copyright(©1975 American Society forMicrobiology
Vol.16, No.6
Printed in US.A.
Complementation
Between
Temperature-Sensitive
and
Deletion Mutants of Reovirus
D. A. SPANDIDOS AND A. F. GRAHAM*
Department ofBiochemistry, McGill University MedicalSchool, Montreal, Quebec H3G1Y6Canada
Received for publication 24June1975
Avery lowlevel of complementation has been found in conventional crosses
between various classes oftemperature-sensitive (ts) mutants of reovirus. A
moredefinitivetestforcomplementationwasdevised throughaplaqueassay on
cell monolayers mixedly infected with defective reovirions lacking the
L,
seg-ment and prototype tS mutants from one or other of the known classes of
reovirusmutants. An increase in thenumber of plaquesonthe mixedly infected
platesoverthaton control plates infected with defective virionsor ts mutants
alone indicated that the ts mutant had been complemented by the defective virus. Class A, B, D, F, and G mutants were complemented at 39C by the
defective viruses, whereas class C and Emutantswerenot.Intests todetermine whethercomplementationwasreciprocalitwasfound that the defective virions werecomplementedbyaclass Gmutantbutnotby the class Cmutant.This and
previous work (D. A. Spandidos and A. F. Graham, 1975) has therefore shown that of thesevenknownclasses oftsmutantsthe class Cmutantis theonlyone
thatneither complementsnoriscomplementedby the defective virions. For this
reasontheclass C ts mutationhasbeen assignedtothe L,segmentof the viral
genome.
A number of temperature-sensitive (ts) mu-tants of reovirus first isolated by Fields and Joklik (6) have been classified into seven groups by geneticrecombination tests (4, 6). It is thought that each group ofmutants repre-sents mutations in one or another of the 10 doubled-stranded (dsRNA) segments of the viral genome and that recombination results from exchange ofwholesegmentsduring repli-cationofthetwoparental viraltypes.
Although recombination is readily demon-strable with this virus, complementation be-tweenthe several mutantgroupstestedis very low. Thus, Fields and Joklik (6) reported that complementation indices between pairs of class A mutantsand betweenAandBmutantswere between 2and 7. Such indices are on the border-lineof significance. In adifferent type of test it was found (11) that in cells co-infected with pairs ofdsRNA-mutantsundernonpermissive conditions there was a small but significant
increase in the total amount of virus-specific RNA synthesized over that of the mutants takenseparately.Thisobservationsuggesteda low level of complementation between C, D, and E class mutants. On the other hand we havefound thatdefective reovirus lackingthe
L,
segment isreadilycomplementedatthe non-permissivetemperaturebythe Eclassmutant, although there is no reciprocalcomplementa-tionofthe E functionbythe defectivereovirus
(21). This result classified the E mutation as beingin acis-function whereas the
L,
genomic segment codes for a trans-function. Conse-quently, lack of complementation between some mutant pairsmight be explicable ifsev-eral of the reoviral functionsarecis-acting. An opportunity to test this proposition was provided by our recentisolationof
L,-defective
virions (21). In the present work we have de-visedasimpletestwith thesedefectivevirions to show that they will complement the known classes ofts mutants except C and E. On the other handit isfound that allts mutantsexcept the C classwill complement thegrowth of defec-tivevirions atthenonpermissivetemperature. Theseresultssuggestthatof the knownmutant classesonlythe E mutation is in acis-function, whereasthe C mutation is inatrans-function thatresidesintheL1segmentofthegenome.MATERIALS AND METHODS
Cells and virus.Lcellsweregrowninsuspension
in Eagle minimal essential medium supplemented
with5%fetal calfserum.
Buffers, chemicals, labeled precursors, and the conditions forgrowth and purification ofstandard (infectious) and defective virions have been de-scribed (21).
Thetwowild-typestrainsof reovirus serotype 3 previously described (21) have been used.According
tothenomenclature introducedintheprevious pa-per (21) these strainswere designatedR,3 andR23
1444
on November 10, 2019 by guest
http://jvi.asm.org/
COMPLEMENTATION BETWEEN REOVIRUS MUTANTS where the superscript 3 represents the viral
sero-type. We havealso used the Lang strain of reovirus type 1 (17)
(R,1).
The following ts mutants of R23 reovirusisolated by B. Fields and hiscollaborators (6) have been used as prototypes of the mutantclasses: R23A (201), R23B (352), R23C (447), R23D
(357),R23D (585),R23E (320),R23F (556), R23G (453). Thesemutantswerekindly supplied by Fields. The
deletionmutantR13d(L,)hasalso been used: this is
a defective virion lacking the L1 genomicsegment whose isolation has beendescribed (21). Permissive temperatureforthe plaqueassay(17)was31C and
the nonpermissive temperature was 39C. For the present work all virus strainswere freshly cloned.
The wild-type strainsormutants asreceived from
Fields were plaque titeredat 31C andanisolated
plaquewaspicked. Eachplaquewasgrownuptoa
large stock in L cells and virus from the final lysates
was then purified through the standard procedure
foruseinthe various experiments.
Standardcomplementationtest.Inthistestthe ability ofanytwo mutants tocomplement each other
wasdeterminedby co-infectingamonolayerculture
(approximately 5 x 106 cells in ascrew-capbottle)
withthetwo mutants atamultiplicity of infection
(MOI) of 10 PFU/cell for eachmutant.Thedeletion mutant
RI3D
(Lj)was used ata MOI of200 parti-cles/cell. Similar cell cultures were infected witheachmutantseparatelyatanMOI of 10 PFU/cellor
200particles/cell ofR13d (L1) virions. In these experi-ments the MOI is given as added MOI. Since the
adsorption rate, measuredon numerousoccasions,
isapproximately 80% in 1 h the adsorbed MOI is 8. After1hatroomtemperature topermitadsorption ofvirus the cultureswerewashedtwice with5 mlof Eagle minimal essential medium, then 5mlof mini-malessential medium containing 2% fetal calf se-rumwasadded and the cultureswereplacedat39C. Viralgrowthwasterminatedat18 hbyfreezing the
culturesat-70C. Whenrequired, the cultureswere
thawed, frozen and thawed three additional times, gently sonically treated for 2 min, and plaque ti-teredat31and 39C.
The complementation index forany twots mu-tantsXand Ywascalculatedaccordingto the form-ula(6):
(XY39)31- (XY39)39/(X39)31 + (y39)31
where (XY39)31 and(XY39)3' are titers of themixed
yieldgrownat39butassayedat31and 39C, respec-tively. (X39)31 and (y3,)31 are titers ofsingle yields
grownat39C and assayedat31C.
RESULTS
Relative plating efficiency of viral strains
at 31 and 39 C. All experimentsinthis work weredone with purifiedvirus. Since each viral stock was developed from a single plaque and
the level ofcomplementation inthis systemis lowitwasessentialtoknowthe relative plating
efficiency for each stock at 31 and 39C. The results of suchplaque assaysareshownin
Ta-ble 1 along with estimates of the number of particles perPFU. Each ts mutantdiffered by
at least a factor of 3 x 10-4 in its plaque-forming ability at the two temperatures. The
very lowlevel of infectious virus contained in
thispreparationof defective virus,
R,3d (L,)
is practically all wild type; very little ofthe Eclassmutantusedto help the defectiveduring its growth(21) ispresent.
Standard complementation
test.-Comple-mentationtestsbetweenvariousmutantswere
carriedoutby the standard proceduredescribed in Materials and Methods. Table 2 shows one
exampleof the results ofthetestbetween class A andBmutants. Thecomplementation index in this experiment is 5 and in a number of
similartestsbetween thesamemutantsit
var-ied between 2 and 10. In other experiments complementation between the A mutant and defectivevirionsgaveindicesbetween1 and5. One set of such results is shown in Table 3 where thecomplementationindexwasfoundto
beapproximately2.
Atcomplementation levelsaslowasthese it iseasytoexplainthe variations in theresults. Anyoneindexdependsontheplaque titrations
TABLE 1. Efficiency ofplatingof10 strainsofpurifiedreovirus
Strainso PFU/ODU.
assayed
at Efficiencyof platingParticles/PFU
Straine
~31
C 39C 39C/31 C assayedat31Cb
Rid
(LI) 4.7 x 105 1.2 x 105 2.6 x 10-l 5 x 106R2 4.2 x 1010 3.3 x 1010 0.8 50
R2A(201) 4.2 x 1010 0.6 x 106 2.3 x 10-4 50
R2B (352) 3.0 x 1010 4.2 x 105 1.4 x 10-5 70
R2C (447) 1.4 x 1010 4.5 x 105 3.2 x 10-5 150
R2D(357) 1.6 x 1010 2.4 x 106 1.5 x 10-4 130
R2D (585) 1.4 x 1010 3.8 x 106f 2.7 x 10-4 150
R2E(320) 2.1 x 1010 7.0 x 106 3.3 x 10-5 100
R2F (556) 2.9 x 1010 1.3 x 106 4.5 x 10-5 70
R2G (456) 2.6 x 1010 1.4 x 106f 5.6 x 10-5 80
aAllstrains wereclonedand clonesweregrowntolargestocks fromwhichvirus waspurified.
bCalculated from the ratio1
OD2,0
=2.1 x 1012particles(20). For the defectivevirionsR13(Lj)the ratio1OD260= 2.35 x 1012particleswas used.
1445 16, 1975
on November 10, 2019 by guest
http://jvi.asm.org/
1446 SPANDIDOS AND GRAHAM TABLE 2. Virusyields aftersingle and mixed
infectionswith R23A (201) and R23 (352)a
InputMOI Yedo rgn
(PFU/cell) Incu-
YieU/ld
ofsproenya
bation (PFU/ml) assayed atR23A R23B temp 31 C 39 C
(201) (352)
10 0 31 7.0 x 107 3.0 x 104
39 5.0x103 1.5x103
0 10 31 5.0x 107 2.0x 104
39 3.0x103 1.2x103
10 10 39 5.5x 104 1.5x 104
aComplementation index: 5.5 x 104 - 1.5 x 104/3.0 x 103 + 5.0 x 103 = 5.
TABLE 3. Virusyields after single and mixed infections withR23A (201) and R,3d(L )a InputMOI
Parti- Incu-
~~Yield
of progenyPalrt-
PFU/ bation (PFU/ml) assayed at cellcel RcellR2
3A temptep 31C 39 CR,Ld
(201)(L,)
200 0 31 4.0 x 102 2.0 x 102 39 1.5x102 1.4x102 0 10 31 7.0 x 107 3.0 x 104 39 5.0x 103 1.5X 103 200 10 39 2.1 x 104 1.0X 104
aComplementation index: 2.1 x 104 - 1.0 x 104/1.5 x 102+ 5.0 x 103 = 2.
ofthree lysates with their inherent errors as well as on the viral yields from the three cul-tures which are low at 39 C and could vary considerably from one experiment to another. In fact, onemight wellquestionwhether these complementation results taken by themselves have any real meaning. It was thought that complementation tests carried out with one member of the pair being the defective virion might be moreconvincing because of the very low background provided by the defective vi-rionitself. Clearly, from the results of Table 3 thisexpectation was not borne out, although in some experiments with defective virions there appearedtobe a low level ofcomplementation. Insearching for a more definitive and less tedi-ous means to study complementation in this system we have devised the test described in
the next section.
Plate test forcomplementation between ts mutants and R13d (L1) virus. This test was designedtofindwhether the R13d
(L,)-defective
virions could complement ts mutants. It is madepossible by the observation (unpublished data) thatLcells infected with these defective
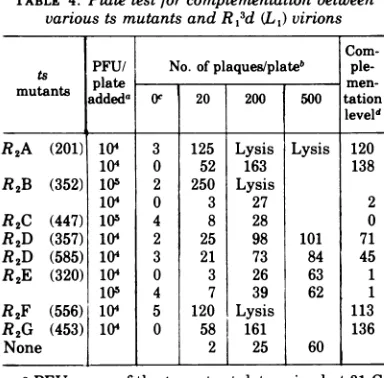
virionsare notrapidly killed. Confluent mono-layers of L cells were infected with a high MOI of defective virions along with a known number of PFU ofagivents mutant.After adsorption of the virus the plates were overlayed as in the standard plaque assay and incubated at 39 C, and the plaquesthat developedwerescored.An increase in the number of plaques over the controls would represent complementation of the ts mutant by defective virions. A representa-tive setof data is shown in Table 4.
[image:3.502.266.458.314.503.2]To determine whether complementation oc-curs between a ts mutantand a given MOI of defective virions the total number of plaques scored at that MOI must be reduced by the number of plaques resulting from the defective virions alone and reduced again by the number of plaques given by the ts mutant in the ab-sence ofdefective virions. The resulting num-ber is calculated as a ratio of the number of PFUof the ts mutant added to the plate and is thendefinedasthecomplementation level(see
TABLE 4. Plate testfor complementation between various ts mutants and
R,3d
(L1) virionsCom-ts PFU/ No. ofplaques/plateb
ple-mtant plate
men-mutants
'addeda
Oc 20 200 500 tation leveldR2A (201) 104 3 125 Lysis Lysis 120
104 0 52 163 138
R2B (352) 105 2 250 Lysis
104 0 3 27 2
R2C (447) 102 4 8 28 0
R2D (357) 104 2 25 98 101 71
R2D (585) 104 3 21 73 84 45
R2E (320) 104 0 3 26 63 1
102 4 7 39 62 1
R2F (556) 104 5 120
Lysis
113R2G (453)
104
0
58 161 136None 2 25 60
aPFUassayof thets mutantdeterminedat31C.
b Plates incubatedat39C. The resultsrepresent theaveragenumber ofplaques onduplicateplates.
Number ofparticlesof
R,3d
(L,) per cell deter-mined from OD250 of purified virus (see Table 1 legend).dThecomplementation level for any coinfection between a tsmutantand defectivesisdefinedas Number ofplaques from mixed infection
- number of plaques from defectives alone - number ofplaques fromtsmutantalone x 104
Number of PFU ofts mutantplated
For mutants R2A (201) andR2F (556) the column under20R,d (L,)particles/cellwasusedtocalculate the complementation level. Forthe othermutants the column200R,d (L,)particles/cell wasused.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
COMPLEMENTATION BETWEEN REOVIRUS MUTANTS
legend to Table 4). Essentiallythe complemen-tationlevel asdefined representsthose cells, a verysmall proportion of mixedly infected cells infact, in whichcomplementation is so efficient that the progeny virus develops a plaque de-spitethenonpermissive temperature. Comple-mentation probably occurs in other mixedly in-fected cells as well but at too low a level to be perpetuated into a plaque. Complementation was measured in this test and not recombina-tion, because, as we will show in a later paper, there is no detectable recombination between
thesedefectivevirionsand anyknown class of ts mutants.
Acomplementation level as defined in Table 4can becalculated for each mutant at each of
thethree MOIs of defective particles.Whenthis isdone it is found that complementation is most
efficientat anMOIof 200 R1
(L,)
particles/cell (Table4). In two cases, R2A and R2F, lysis was almostcomplete on the plates, and the comple-mentation levels have been computed on the results obtained with an MOI of 20 defective virions/cell.It is clear that the E mutant is not comple-mentedbyany MOI of defectives used (column 7, Table 4). This is the expected result for this mutantbecause it hasalready beenshown in a
differentwaythat the E mutant is not
comple-mented by defectives andthatthe E ts muta-tion is in a cis-function (21). Thus, the result with the E mutant provides a base line for comparison(Table4).Against thisbaselinethe A, B, F, and G mutants are clearly comple-mented. The level of complementation of the two Dmutantsislowerbutwellabove the base
line. It can be calculated that approximately 1.4%ofthe cells infected with 104 PFU/cell of B mutantshow complementationinthis test (col-umn 5,Table4). Forthe two D mutants 0.5 to 0.7% of the cells infected with 104 PFU/ml
showed complementation. On theother hand, the C mutant was not complemented and this resultdefines the C mutation to be either in a
cis-functionorinthe
L,
segmentof the genome.Complementation ofdefective virions by
ts mutants. Previously weshowed that when cells wereco-infectedwithdefectivevirionsand class A, B, or E ts mutants at39C there was extensive growth ofthe defective virions (21). Allthree mutantshad complemented the miss-ing
L,
function. Indeed it would be predicted that any ts mutant wouldcomplement this de-fective virion unlessithadthe mutationintheL,
genomic segment, and the results of Table 4 suggested that the class C mutant might be in this category. The following experiment was therefore carried out to determine whether the C mutantwouldcomplement the growth ofde-fective virions using as controls the wild-type
virusand G mutant.
Three suspension cultures were co-infected with defective virions and one or another of wild-type virus, G mutant, or C mutant and placed at 39C. A second set of three cultures was infected in the same way and placed at 31C. The same amount of [3H]uridine was added, with actinomycin D, to each culture to label the progeny virus. When the cells had partially lysed, the cultures were centrifuged and a known amount of14C-labeled, purified wild-type virus (which contained no defective virions) was added to each sediment. This 14C-labeled virus was to act as a carrier, amarker to measure the efficiency of each purification step, and subsequently as a sedimentation marker in gradientanalyses. Virus from each sedimentwas purified and the recovery of 3H and14C at eachstepwasdetermined. Fromthe respective value for recovery of added '4C-la-beledvirus and theamountof material with an
optical density at260 nm
(OD260)
in each puri-fiedpreparation the yield of viral particles per cellwascalculatedfor the various cultures. The resultsareshown in Table 5.Overall recovery of added 14C-labeled virus
varied from 70 to 80%. The co-infection with
wild-type virus and
R,d (Lj)
at 31C gave anormalyieldofparticles per cell (21), whereas the yields with the two ts mutants were
consid-erablylower;these have been consistent obser-vations.Theyieldsofparticlespercellwereall
considerably lessat 39 Cthan at31 Cand, in
fact, novirusparticlesweresynthesized inthe co-infectionbetweenthe C mutantand
Rld
(Lj)
virions at 39C consistent with the view that the class C ts mutation resides in the
L,
seg-ment.Todeterminethe nature of theparticles
syn-thesized, the final purified virus from each of the sixcultures wasdigested with
chymotryp-sin to convertthe virionsto cores (12, 15) and theproductswerecentrifugedtoequilibriumin
CsClgradients. The results for the31and39C
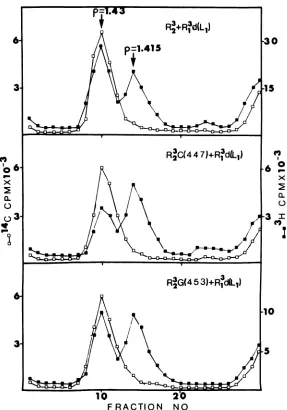
culturesareshown inFig. 1and 2,respectively.
The leading peak of 3H in each gradient of Fig. 1 and 2(p = 1.43g/ml)coincides with the 14C peak ofthe marker and represents cores derived from infectious virions, either wild-type or ts mutant.The trailing peak at p= 1.415
glml represents cores derived from defective virions(14, 15, 21). Since the relative amounts ofcores ineachgradient represents the relative amounts of standard and defective virions in thepurified virus,itisclear from Fig. 1 that at 31 C roughly equal amounts of infectious and defective virions were synthesized in each of the three cultures. Because defective virions
VOL. 16, 1975 1447
on November 10, 2019 by guest
http://jvi.asm.org/
1448 SPANDIDOS AND GRAHAM TABLE 5. Yields ofpurified virus from cultures doubly infected with defective virions andwild-type
or mutant virionsa
Recovery Purifie vi Yieldofvi-Infection
ofleC-la-
erd pripaertell-rusb (%)
tures
x 10431C
R2 xRld(L1) 78 8.7 8.8
R2C xR,d(LI) 73 3.1 1.9
R2G x
Rld
(L1) 81 4.0 2.639C
R2 x
R,d
(Li) 76 3.3 2.0R2C xRld(L1) 81 1.9 0
R2G x Rld(L1) 70 2.6 1.4
aThegeneral method of carrying out the
experi-ment is described in the text. Each culture con-tained 200 ml at 5 x 10'cells/ml coinfected with 500 particles per cell ofR,3d(L1)virionsand 10PFU/cell of one or other of the viruses, R23,R23C(447), R23G (453); 0.5
i&g
of actinomycin D/m 1; 1 juCi of[3H]uridine/ml. After 40 h at 31 C or 12 h at 39 C the
cells were centrifuged and to the sediment was
added 2.35 x 1012 particles of purified R23 virus
containing 8 x 104 counts/min of14C in its RNA.
Purificationof virus from the sediment followed the
previously describedmethod (21).
bTotal14C-labeledvirus recovered after purifica-tion as apercentage of that added before purifica-tion.
cTotal purified virus recovered from each culture
includingtheadded14C-labeledvirus. Particles per
culture calculatedfromthe relationship, 2.35 x 1012
particles/OD2,0, see footnoteTable 1.
dCalculated from the results in the preceding
columnbycorrecting thefiguresforrecovery of
14C-labeled virus and then subtracting the number of
14C-labeled particlesoriginally added.
cannotreplicate alone, nor synthesizeprogeny dsRNA(unpublished data), growthof the defec-tive virions has been "helped" at 31 C by the presenceof infectious virions. A similarhelping
effect has been shown with the A, B, and E mutants at 31C (21).
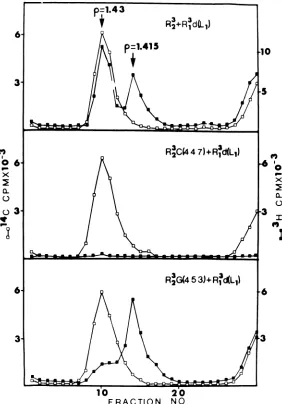
Adifferentsituationis seen at 39C (Fig. 2). Although the overall yield of particles was
con-siderably reduced (Table 5), wild-type virusat
39 C helped (complemented) the defective vi-rions asefficientlyas at31 C. Mostof thevirus formedinthe co-infection of GmutantandR
ld
(Lj)
virions at 39C wasdefective: the Gmutanthadefficientlycomplemented growth of the de-fective virion,although therewaslimited
recip-rocalcomplementation. Inacontrol culture in-fectedwithGmutantalone(resultsnotshown)
therewaspracticallynogrowth of themutant
at 39C and no defective virions were formed. Thiscontrol shows that the defectivevirionsdid not arisefromthe ts mutantitselfashasalso
beenfoundwiththe A and B mutants (21). The middle panel of Fig. 2 shows that there was no growth ofany virus at39C intheco-infection between C mutantanddefective virions.
These results are interpreted to mean that the G mutant can readily complement the growthofdefectivevirions at39C but is itself poorly complemented by the defective virion. Similar remarks applied to co-infections be-tween the defective virus and the A or B
mu-tants (21) and also to the D and F mutants (unpublished data). The low level of growth of A, B, D, F, and G mutants at 39 C in this type of experiment is consistent with therather low level of complementation of these ts mutantsby defective virions shown in Table 4. In contrast, there is nocomplementationineither direction between defective virions and the C mutantat
39C, although both virions grow reasonably welltogether at 31 C.
Complementation of R,3d
(L,)
virions by reovirus type 1.Whereas types 1 and 3 reovi-ruses areserologically distinct, the sizes of the three L segments of the two serotypes are simi-lar. It was therefore of interest to determine whether thetype 1viruswouldcomplementthe type3-defectivevirions. Aculture ofLcells was co-infected withR,1virus andR,3d
(L,)
virions at31 C and the viralprogeny was labeled with [3H]uridine during its growth. After purifica-tion '4C-labeled R23 virus was added as a marker and theresultingmixturewasdigested with chymotrypsin to convert it to cores and subjected to isopycnic centrifugation in CsCl (Fig. 3a).In acontrol experimentaculturewas infectedwith type 1virusalone andsubjectedto similar manipulations. The results of the con-trol experiment (Fig. 3b) show that infection with type 1 virus alone did not give rise to defective virus. However, the gradientof Fig. 3a shows a considerable peak at the buoyant densityofdefective cores (p = 1.415 gIml) that resulted from the mixedinfection. Thustype 1 viruscanreadilyhelpthegrowth of thetype 3-defective virions. We have recently suggested that the L1 segment of the viral genome codes for the X2 polypeptide of the viral core (13). If thisis sothedefectivecores,resulting fromthe co-infection ofR,1
virus andR,3d
(Ll) virions, should be aphenotypicmixturecontainingthe X2polypeptideoftype1virus with the remain-ingcore peptides,X1,1Ai,
and0-2(20), being type 3 inorigin. Experiments are in progress to test thisprediction.DISCUSSION
Theuseof defectivevirionsinthisstudyhas servedtwomainpurposes:first,todemonstrate
unequivocately that complementation in the
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.502.59.256.75.237.2]COMPLEMENTATION BETWEEN REOVIRUS MUTANTS
p=1.43
R3+R3d(Ll)
p=1.415 3
R3C(4
47)+R3d(L1)
0~
0
/~~~~~~~~l
3FRACTION NO
FIG. 1. Cesium chloridegradient analysis ofthe viralcoresobtainedfromthe 31Cprogenyof co-infection
betweendefective virions and wild-typeortsvirus.Thepurifiedviralyields fromthe 31 C cultures described inTable 5,containing "4C-labeled marker R23 virus,weredialyzed against0.05 MTris-hydrochloride buffer,
pH8.0,anddigested for60 minwith100 jgofchymotrypsinperml. Theresultingmixturewaslayeredovera
preformed gradient ofCsCl (p=135 -1.46g/ml) andcentrifuged for6hat37,5(X) rpmintheBeckmanSW40
rotor.Fractionswerecollected from the bottom of the punctured tubeandanalyzed for densityand radioactiv-ity. The particularcross usedineachcoinfection isdescribed in theappropriate panel. Symbols: (5)14C(
labeledmarkervirus; (U)3H-labeledprogenyoftheco-infection.
reovirus system can be recognized by genetic
techniquesand, second, toallocatethe class C
mutationtothe L1segmentof theviralgenome.
Levels ofcomplementation in the standard
testweretoolowandvariabletobesureof their
significance. However,wehavedevisedamuch more sensitive test for complementation in
which cell monolayers were firstspread with
defective virions, then superinfected with ats mutantandoverlayed. Insomeof thesecrosses
asignificant fractionof thedoublyinfected cells gave rise to plaques (Table 4). Although no
recombination has been detected between
R,d
(L1) virions andanyclass oftsmutants (unpub-lished data), there is still a possibility that some co-infected cells might give rise to ts+ recombinants. Even so, a preliminary
comple-mentation would have to occur between the
parental virions to provide the genomes to
re-combine witheach other. Whatever the
mecha-VOL. 16, 1975 1449
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.502.105.392.66.476.2]1450 SPANDIDOS AND GRAHAM
0
U
x 0
x0 U I
I,
FIG. 2. Cesiumchloridegradient analysisof the viralcoresobtained from the39C progenyof co-infection betweendefectivevirions andwild-typeor tsvirus.Thepurifiedviralyieldsfromthe39C culturesdescribed inTable 5 were treatedasdescribedinthelegendtoFig.1.Symbols: (O)14C-labeled marker virus; (M) 3H-labeled progenyoftheco-infection.
nism of plaque development in this test the primary event must be complementation. Thus, all known classes of ts mutants except C andE arecomplemented byR d(L1)virionsat
the nonpermissive temperature.
Conversely, growth ofdefective virusatthe nonpermissive temperature is clearly comple-mentedbywild-typevirusandthe class G
mu-tant (Fig. 2), bytheA, B, and E mutants (21), by D and F mutants (unpublished data), and evenbyserotype1reovirus(Fig. 3).The class C mutant is the only one that neither comple-mentsdefectivevirionsnoriscomplementedby
them. Thus, with some confidence we can as-sign the class C ts mutation to the L1 segment of theviral genome.
Onepractical upshotof this work may be in the use of
R,d
(L,) virions to select new ts mutantsthat are notcomplemented bythe de-fectivevirions. Sevendifferentclasses of tsmu-tants areknown and, since each class is pres-entlythoughttorepresent a different segment of the genome, there should be three classes remaining to be discovered. Moreover,onlyone
mutantineach of classes C and Eisknown(4) anditwould be usefulto havemore. Theplate
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.502.123.405.71.475.2]COMPLEMENTATION BETWEEN REOVIRUS MUTANTS
0
0 I-C)
04
6
0-x
iqt
[image:8.502.49.444.59.341.2]FRACTION NO
FIG. 3. Cesium chloride gradient analysis of viral cores obtained from the co-infection between defective type 3 virions and type I reovirus. A culture containing 200 ml at 5 x 105 Lcellslmlwas co-infectedwith 2,000
RI3d
(L1) virions and 10 PFU ofR11 viruslcell, and 05 pg ofantinomycin D per ml and 10juCi
of[3H]uridine/mlwereadded. A second culture was treated inthe same way after infection with 10PFUlcellof
R1'virus.Theprogenyvirus was purified, mixed withpurified I4C-labeled R23virus as amarkerand treated asdescribed in the legend to Fig. 1. (a) Co-infection withR23d (L1)andR11virions; (b) infection with
RI'
virus. (0) "IC-labeledmarker virus;(U)3H-labeled progeny of the infection.
method described in Table 4 could be used to
testnewly isolatedts mutantsfor
complementa-tionby R1d (L1) virions. Thosets mutants not
complementedinthetestwould be classCorE or a newclass.Infact,onemight
predict
thatatleast one new class ofmutants would be
iso-latedinsuchatest,namely thosewithts
muta-tionsin the viriontranscriptase (1, 19), which
should beacis-acting function. Althoughithas beensuggested that theviriontranscriptase of the class Gmutantisslightlyts(2) itisshown thatthis mutant iscomplemented by
R,d
(L1)virus (Table 4) and that it complements the
defectivevirion(Fig. 2). Consequentlythe
ma-jor mutation inthe G class mustbe elsewhere than inthetranscriptase. There is no evidence that either the C or E mutation affects the viriontranscriptasealthoughbothmutants are dsRNA- (2, 11). In fact, the evidenceis to the contrary inthat bothgenomeshave been shown tobetranscribedininfected cells under nonper-missive conditions (2, 11) and such transcrip-tion mustbe carriedoutby the parental virion
polymerase.As will bedescribedindetail
else-where the genome of
RId
(L1) virions is also transcribed in vivo, although no dsRNA is formed.Althoughtheplatetestwith
R,d
(L1) virions shows that they will complement various ts mutants,itdoesnotprovideamethodforclassi-fying such mutants into complementing
groups. The plate test, taken in conjunction with the results ofthe reciprocaltest(e.g., Fig. 1and2),has nevertheless specifiedthat theA,
B, C, D, F, and Gts mutations are in
trans-functions, whereas the E mutation isin a cis-function.Complementation should therefore oc-curbetweents mutants of reovirusindifferent trans-actingfunctions. This encourages one to proceed with a systematic application of the standardcomplementation test tothe reovirus mutants. Even though complementation indi-ces may be ofborderline significance in some crosses, it is important to find whether the classification ofmutantsaccordingtotheir func-tional differences correspondstothatbased on recombination frequencies(4, 6).
Pure populations of different deletion
mu-VOL. 16, 1975 1451
on November 10, 2019 by guest
http://jvi.asm.org/
1452 SPANDIDOS AND GRAHAM
tants will be needed to allocate other tsmutant classes to their specific segments by the comple-mentation plate test described here. Since six of the seven known viral functions are trans-act-ing any one of these could theoretically be de-leted. In fact, on continued passage of practi-cally all strains of reovirus, wild type or ts, a spectrum of defectives is obtained (unpublished data) some with double deletions of whole seg-ments and at least one with a partial deletion. Some ts mutants, forexample the class C mu-tant, generate defectives with high frequency even atlowmultiplicities of infection (18). Isola-tionof specific defective virions from such mix-tures, in which virions with
L,
deletions are usuallyinlarge excess, is aformidable techni-calproblemfor which there is no apparent gen-eral solution. On the other handproduction of defective viruses maywellmodify the course of an infection with the parental strain as has been discussed by Huang and Baltimore (10).Evidenceisaccumulatingthat thedevelopment
of persistent viral infection bothincell cultures and in animals frequently involves the selec-tion of ts mutants during the infection (16). In fact subacute neurotropic infections in mice have beeninduced through exposure of the ani-mals to certain tsmutantsof reovirus(5, 7) and ofmeasles virus(8). Developmentand mainte-nance of these chronic infectionscould be sim-plyexplained by the long-term limited growth of thetsmutantsunder temperature conditions veryclose to nonpermissive. It is alsopossible
that some subacute viral infections may be the result of a complex interplay between a ts mu-tantand defective virionsgeneratedfrom it or from the parentalwild-type strains,ashas been shown for persistent infections with vesicular stomatitis virus forexample (3, 9). Continued
analyses of the sort presented in this paper should givesome insight intohow such mixed viral populations can maintain themselves throughmutualcomplementation.
ACKNOWLEDGMENTS
Thiswork wassupported bygrants from the National Cancer Institute ofCanada and the Medical Research Coun-cil ofCanada.
LITERATURE CITED
1. Borsa, J., and A. F. Graham. 1968. Reovirus: RNA polymeraseactivityinpurifiedvirions.Biochem. Bio-phys.Res.Commun. 33:895-901.
2. Cross, R. K., and B. N. Fields. 1972. Temperature-sensitivemutantsof reovirustype3:studies on the
synthesis of viral RNA. Virology 50:799-809. 3. Doyle, M., and J. J. Holland. 1973. Prophylaxis and
immunization in mice by use of virus-free defective T particlesto protect againstintracerebralinfectionby vesicular stomatitis virus. Proc. Natl. Acad. Sci. U. S. A. 70:2105-2108.
4. Fields, B. N. 1971.Temperature-sensitive mutants of reovirus type 3: features of geneticrecombination. Virology46:142-148.
5. Fields, B.N. 1972.Genetic manipulation ofreovirus-a model for modification of disease? New. Engl. J. Med. 287:1026-1033.
6. Fields, B. N., and W. K. Joklik. 1969. Isolation and preliminary genetic and biochemical characterization oftemperature-sensitive mutants of reovirus. Virol-ogy 37:335-342.
7. Fields,B.N., and C.S. Raine. 1974.Alteredneurologic disease induced by reovirus mutants, p. 161-167. In W. S. Robinsonand C. F. Fox (ed.), Mechanisms of virus disease, vol. 1, ICN-UCLA Symposia on molecu-lar and cell biology. W. A. Benjamin, Inc., Menlo Park, Calif.
8. Haspel,M. V., and F. Rapp. 1975. Measles virus-an unwanted variant causing hydrocephalus. Science 187:450-451.
9. Holland, J. J., and L. P. Villarreal. 1974. Persistent non-cytocidal vesicular stomatitis virus infections me-diated by defective T particles that suppress virion transcriptase. Proc. Natl. Acad. Sci. U. S. A. 71:2956-2960.
10. Huang, A. S., andD.Baltimore. 1970. Defective viral particles and viral diseaseprocess. Nature(London) 226:325-327.
11. Ito,Y.,andW. K.Joklik. 1972.Temperature-sensitive mutants of reovirus. I. Patterns of gene expressionby mutants of groupsC,DandE.Virology50:189-201. 12. Joklik,W. K. 1972.Studiesonthe effect of
chymotryp-sin on reovirions.Virology49:700-715.
13. Lau, R. Y., D. vanAlstyne,R.Berckmans, andA. F. Graham.1975.Synthesis ofreovirus-specific polypep-tidesincellspretreated with cycloheximide.J.Virol.
16:470478.
14. Nonoyama, M., and A.F.Graham.1970.Appearanceof defectivevirions inclones ofreovirus. J.Virol. 6:693-694.
15. Nonoyama,M., Y.Watanabe, andA. F.Graham.1970. Defectivevirionsofreovirus.J.Virol.6:226-236. 16. Preble, 0. T., and J. S. Younger. 1975.
Temperature-sensitive viruses andtheetiologyofchronicand inap-parent infection. J. Infect. Dis. 131:467-473. 17. Prevec,L.,Y. Watanabe, C.J.Gaunt, andA. F.
Gra-ham. 1968.Transcriptionofthe genomes of type 1 and type 3reoviruses.J.Virol.2:289-297.
18. Schuerch,A.R.,T.Matsuhisa,and W. K.Joklik.1974. Temperature-sensitivemutantsofreovirus. VI. Mu-tant ts 447andts 557particles that lack eitherone or twogenome segments.Intervirology3:3646. 19. Shatkin,A.J., and J.D.Sipe. 1968. RNApolymerase
activityinpurifiedreovirus.Proc.Natl. Acad.Sci. U. S. A. 61:1462-1469.
20. Smith,R.E.,H. J.Zweerink,and W. K.Joklik. 1969. Polypeptide components ofvirions, top component and cores of reovirus type3.Virology39:791-810. 21. Spandidos,D.A.,and A. F. Graham.1975.
Complemen-tationofdefective reovirusbyts mutants.J. Virol. 15:954-963.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 3.R1'asRI3dvirus.type[3H]uridine/ml described Cesium chloride gradient analysis of viral cores obtained from the co-infection between defective 3 virions and type I reovirus](https://thumb-us.123doks.com/thumbv2/123dok_us/1565814.109178/8.502.49.444.59.341/described-chloride-gradient-analysis-obtained-infection-defective-reovirus.webp)
