Extracellular matrix remodelling in response to venous hypertension: Proteomics of human varicose veins
Full text
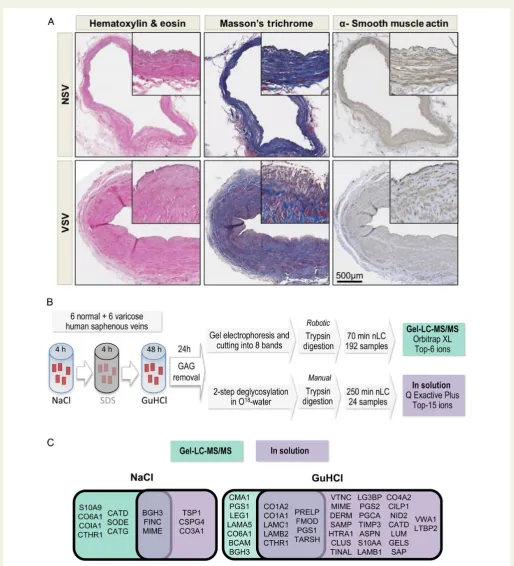
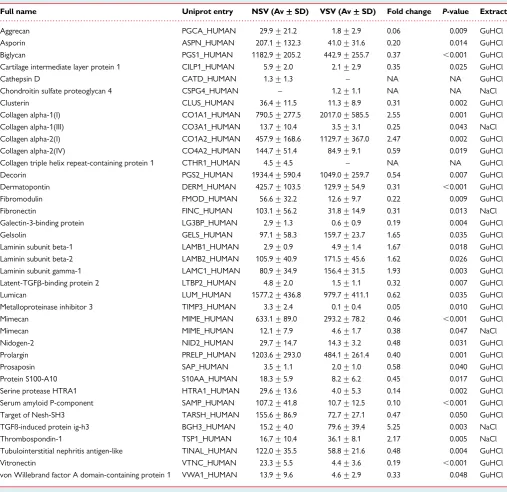
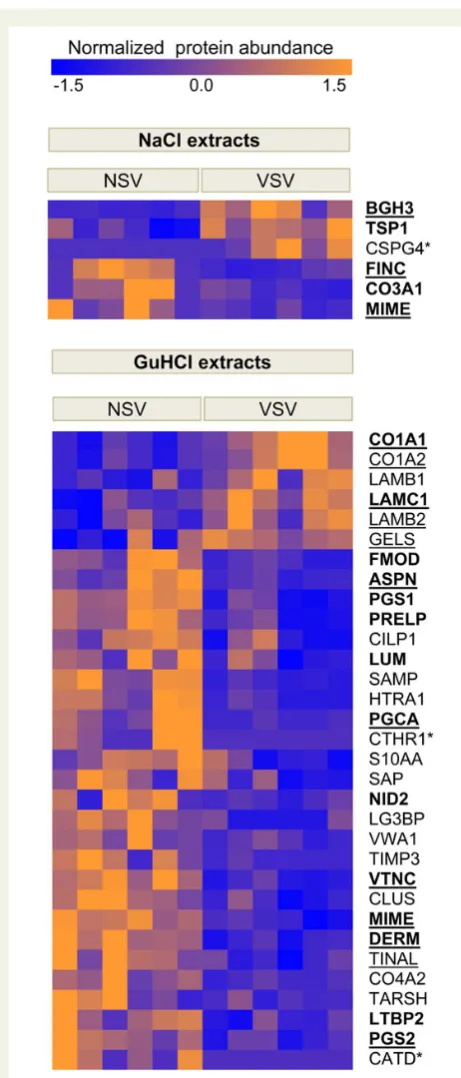
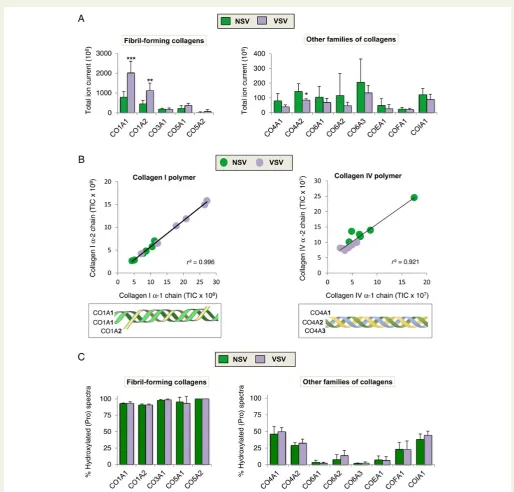
Figure
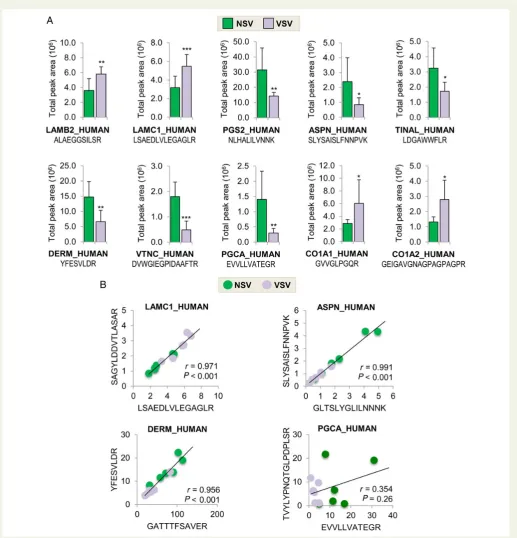
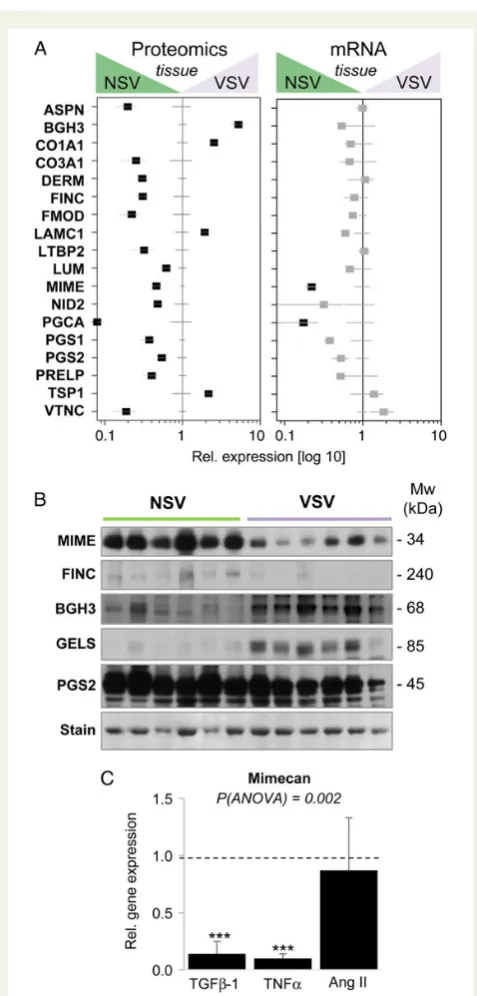
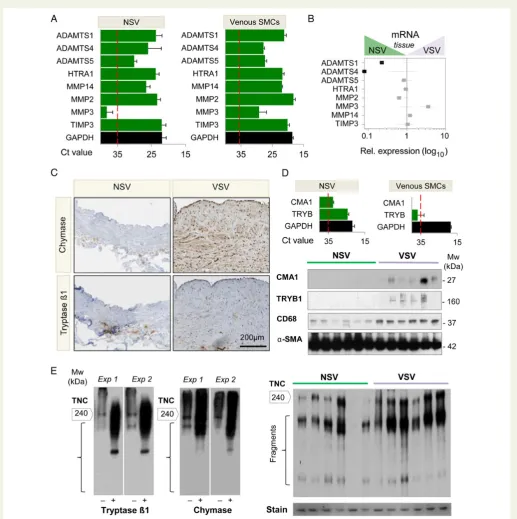
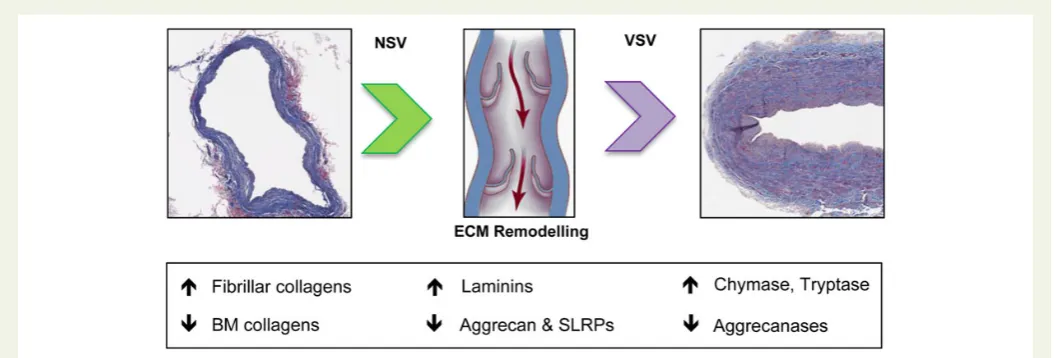
Related documents
all of these studies, one can conclude the effect of bisphosphonate therapy on changes in BMD in post-menopausal women with diabetes is variable and difficult to compare, and that
Rauth et al. 7) suggest that design thinking education (i.e. the process of teaching design thinking) is able to develop a certain mindset as creative competence that 'culminates in
Special Low Level Commands are subject to use in a external Software running on a PC. These Commands are locked until a special command
squat exercise only from the Lower Body - Primary Exercises list and after warm up sets, start with 40% of your estimated 1RM and do 5 sets of 2 repetitions off of a box. You will
It is common, for example, to distinguish between negative (freedom from) and positive (freedom to) conceptions of freedom, and their different implications for thinking about the
Examining our case a posteriori, the initial anamnestic data of milky urine and renal calculus and the emission of whitish material immediately after the stent positioning could
Approximately 40% of deaf children have mental health problems versus 20% of hearing children Deaf children more likely to be identified with ‘externalizing problems’
Proof that the technology works is in the consistently high scores that distance learners get on examinations says Weiss 1994 in a report on a variety of video applications for
