CopyrightC)1993, AmericanSocietyforMicrobiology
Bovine
Leukemia
Virus RNA
Sequences
Involved in
Dimerization
and
Specific
gag
Protein
Binding:
Close Relation
to
the
Packaging
Sites of
Avian, Murine,
and
Human
Retroviruses
IYOKOKATOH,1* TERUOYASUNAGA,2t ANDYOSHIYUKIYOSHINAKA'
Microbiological Research Institute, Otsuka Pharmaceutical Co., Ltd., 463-10Kagasuno, Kawauchi, Tokushima 771-01,1and Computation Center, TheInstituteof Chemical and
Physical Research, 2-1 Hirosawa, Saitama 351_01,2Japan
Received 1September1992/Accepted 15 December1992
In vitro detection ofa specific complex of the bovine leukemia virus (BLV) MA(p15) protein and the 5'-terminalRNAdimer led to thehypothesisthat theNH2-terminaldomain ofretrovirusgagproteinprecursor
isinvolved intheselective viral RNApackagingmechanism. Herewedescribemappingof the BLV RNA for
dimer-formingandMA(plS)-bindingabilitiesbyasimplecDNAprobingmethodfollowedbymutationanalyses
with thereactiveU5-5'gagRNA. The RNAdimerization is mediated bytheregionharboringU5,theprimer binding site (PBS), and the 30 bases immediately downstream of PBS. This conclusion is supported by computer-assisted RNA secondary-structure analysis which predicted a multibranched stem-loop folding
throughoutthe dimerregiondetermined. Anotherregionfrom PBS tothe 5'-terminal 60 residues ofthegag gene, partially overlapping the dimer region, likely provides essential elements for the MA(pl5) binding reaction, although the presence of either the 3' or5' neighboring sequences increases the complex-forming efficiencysignificantly, and each of thesubstructures predicted withinthecore region has,ifany,onlyvery
weakaffinitytoMA(p15).Thesein vitro characterizations of the BLV RNAmayreflectgeneralfeatures of the specific protein-RNAinteraction in thepackagingeventsof variousretroviruses.5'-terminal folded structures ofretroviral RNA molecules and theirbiologicalactivitiesarediscussed.
Although the molecular mechanism ofretrovirus replica-tion has been characterizedtoagreatextent,several critical
processesremain tobeexplained. Amongthem isthestage
of virusparticle formation, the stage atwhich assemblyof thegagprecursor polyprotein andspecific association ofa
domain of theprecursor with viral RNAare thought to be primary biochemical events leading to productive virus morphogenesis.
Two schools of evidence have beenpresentedin thestudy of the virus protein-RNA interaction. Results of in vitro mutagenesis experiments emphasizedthatthe COOH-termi-nalnucleocapsid (NC) domain of thegagprecursorand its zincfinger-like motifsarerequiredfor viral RNApackaging (3, 9, 10, 15, 29-31). Direct RNAbinding experimentswith purifiedviral proteins suggested that the NH2-terminal do-main ofthe matrix-associated (MA) proteinorits adjacent
polypeptide plays the key role (17, 21, 39, 40). The NC proteinwasshowntobeanonspecificRNA-binding protein
(16, 17, 20) thatformstheviralnucleocapsid complex. On the otherhand,locationof thepackaging signalin the
genomeRNAhas been assessedbydeletion and recombina-tion analyses, andno significant difference amongthe rela-tivepositionsdetermined for retroviruses of variousspecies has been found(3, 18,23, 28, 33, 41, 43, 45). Recent results ofdetailed examinations ofmurine leukemiavirus(MuLV) and Roussarcomavirus, however, indicate that packaging
sequences should be in a more extended region than was
originallydetermined(1,5-7, 24). Another intriguing feature of retroviralgenome encapsidation isthat theviral RNA is incorporated into the particles as a dimer of the identical
*Correspondingauthor.
tPresentaddress: GenomeInformationResearchCenter,Osaka
University, 3-1Yamadaoka, SuitaCity,Osaka565, Japan.
molecule. It has longbeenpostulated that the dimer struc-ture is closely related to the packaging events. Although hypotheticaldimermodelswerededuced from possible base
pairingswithin the 5'-terminal RNAsequences, their
prob-abilities have not been experimentally validated (8, 13). Recently, thepresence ofafoldedstructureofa stem-loop
with multiplebranches has been reported for thecis pack-aging element of human immunodeficiency virus type 1 (HIV-1) on the basis of computer analysis in combination withenzymeand chemicalprobing experiments (12). Asyet,
however, no direct in vitro evidence points to the folded structureasthesignal recognized by the HIVgagprecursor
protein (26).
Hereweshowthatminimum elements required for bovine
leukemia virus(BLV) RNA dimerization and complex for-mation with MA(p15)arelocated inaregion corresponding to the packaging site determined for various retroviruses. Ourelectrophoretic analysisoftheprotein-RNAinteraction
may account for one of the critical steps of retrovirus packagingofdimericviralRNA.
MATERUILSANDMETHODS
Purification ofMA(pl5). BLVwasprepared from culture
fluid of Bat2Cll cells (36) as described previously (17).
MA(p15)waspurifiedfromthe BLVpreparations byacetone
treatmentfollowed by Sephacryl S-200 column chromatog-raphy and reverse-phase high-performance liquid chroma-tography (RP-HPLC) as described previously (17). The purified protein fractionwaslyophilized, solubilized in H20
with 0.025% Nonidet P-40, and quantitated by the Lowry method (25). TheMA(pl5) solutionwas adjustedto 50 ,uM andstoredat4°C.
Synthesis of RNA fragments. Viral RNA fragments and
1830
on November 9, 2019 by guest
http://jvi.asm.org/
DIMERIZATION AND gag PROTEIN BINDING IN BLV 1831 their mutants weresynthesizedby the SP6 RNApolymerase
reaction as describedpreviously (17). 32P-labeledU5-5'gag and mutant RNAs wereproduced in reaction buffer (40 mM Tris-HCl [pH 7.5], 6 mMMgCl2, 2 mM spermidine, 10 mM dithiothreitol, 1 U of human placenta RNase inhibitor (Takara) per
p1)
containing 0.5 mM each GTP, UTP, and ATP, 0.05 mM CTP, 0.025 mmol [-32P]CTP, 2 ,g of linearized template DNA, and 20 U of SP6RNApolymerase (Takara)inatotal volume of 20P,l.
Incubationwasfor 1 hat37°C. Unlabeled U5-5'gag was synthesized by incubating for 16 h at 37°C the followingreaction mixture(50
jlI):
the same buffer containing 0.5 mM all four ribonucleoside triphosphates, 1 ,ug of pAM5G cleaved by SailI (pAM5G/ Sall; SalI from Boehringer Mannheim) perPIu,
and30 U of the enzyme. Afterthepolymerase reaction, the templatewas removed by DNase I (8 U) digestion for 10 min. Products were deproteinized bytwo extractions with phenol-chloro-form-isoamyl alcohol (25:24:1) followed by one extraction withchloroform-isoamyl alcohol(24:1)and thenprecipitated with ethanol.Dried RNAwasdissolved inH20 and quanti-tated. Aliquots were stored at -70°C. For the MA(p15) bindingassay, RNAsolutionwasmadeonice in 2x binding buffercontaining 20 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid(HEPES;pH7.9), 12mMMgCl2,20 mM NaCl, 2 mM dithiothreitol, and 1 U of RNase inhibitor perPI.
cDNA probing. DNAs consisting of 18 or 30 bases com-plementary tothe U5-5' gag RNA sequences were synthe-sized by a DNA synthesizer (Applied Biosystems model 381A) and purified by RP-HPLC through a C18 column
(Waters)
withanacetonitrilegradient
from 5to20%in 0.1 M triethylamine acetate (pH 7.0). In one set of experiments, syntheticcDNAswere5' end labeled with[_y-32P]ATP
(New England Nuclear) and T4 polynucleotide kinase (New En-glandBioLabs), phenol extracted, and ethanolprecipitated. Each labeled cDNAwas addedtotheSP6 RNApolymerase reaction mixture (20P,)
containing pAM5G/SalI (2P,g)
and fourunlabeled ribonucleosidetriphosphates priorto incuba-tion. In another set of cDNA probing assays, unlabeled syntheticcDNAwasaddedtothe RNApolymerase reactionto produce U5-5' gag labeled with [a-
2P]CTP.
Unlabeled ATP, GTP, andUTPwereused at0.5 mM each; CTPwasadjusted to make a final concentration of0.075 mM. Incu-bationwasfor 1 hat37°C.DNasetreatment waseliminated. Construction ofplasmidsfor theexpressionofmutantRNA segments. The wild-type plasmid previously referred to as pAMGhas been renamed
pAM5G
for easieridentification in comparisonwith various DNAs used in this work. Deletion and fusion mutantswere newly constructed fromparental
plasmids
pAM5G
and pAMGPE(17).
Their identities and construction methods are described below. Nucleotide po-sitionswere numbered in relation to thetranscription
start site, +1. DNA fragments generated by restriction enzyme digestionsarealways indicated belowasA-B,representing
thesequencesfrom the5'end
(A site)
tothe 3' end(B site)
intheorientation of theBLV
coding
sequences(38).
Inthecase of noncohesive
ligation,
ends were blunted with T4 DNApolymerase priortotheligation
reaction unless spec-ified otherwise. Plasmids obtained from transformants of EscherichiacoliHB101werescreenedby
restrictionenzyme mapping.Construction of deletion mutants. (i)
pAM5GdP.
Plasmid pAM5G was cleaved into twofragments bydigestion
with BclI (at position+340)
and XmnI(at
aunique
site in the vector, pAM18[Pharmacia]).
Onefragment,
XmnI-BclI(+340),
wasdigested
with BstNI(+301)
to remove theprimer bindingsite
(PBS)
andligated
totheotherfragment,
BclI
(+341)-XmnI.
(ii)
pAM5Gda.
The BLV U5-5' gag sequences, +147 to+940,
were excised frompAM5G by
digestion
withEcoRI andSphI (at
theunique
cleavage
sites derived from themultiple
cloning
site of thevector)
andfurtherdigested
with AvaIl(+369)
to remove the 5'-terminal sequences. The deletedfragment,
AvaII(+370)-SphI,
waslinkedtotheSall(in
themultiple
cloning
site)-BclI (+340) fragment
which containedthevectorand the BLV sequencesfrom +147to +340.(iii)
pAM5GdPdK.
pAM5GdPdao
was constructedby
the cohesiveligatipns
of four DNAfragments,
Avall(+370)-SphI,
SphI-XmnI,
XmnI-BstNI(+321),
and aBstNI-AvaIIadaptor
madeby annealing
two strandsofsynthetic
oligo-nucleotides,
5'AGGAGA3' and3'CCTCTCTG5'.(iv)pAM5Gd,3.
pAM5G
wasincubatedwithAvaIItomake apartial
digestion.
Linear forms wereisolated,
further cleavedby
BssHII(+487)
to removeAvaII(+373)-BssHII
(+487),
andrecircularized.(v) pAM5Gded3. BclI
(+345)-BssHII (+487)
sequences wereremovedbydigestion
ofpAMSG
with thetwoenzymes and recircularized.(vi)
pAM5GdPdcKdI.
TheXmnI-BstNI(+301) fragment
was linkedto the BssHII
(+488)-XmnI fragment.
(vii) pAMLG.The BstNI
(+302)-SailI
(+936) fragment
was isolated frompAM5G
andinserted intothepAM18
vector at theSmaIcloning
site.(viii)
pAMgag.
TheSacl(+147)-DraI
(+535)
region
wasdeleted from
pAM5G by
digestion
of thetwoenzymes. Construction of recombinantmutants.HindIII
(in
themul-tiple cloning
site)-BamHI (+5015)
sequencesencoding
the3' end of gagtothe5' end ofenv wereremovedfrompAMGPE
to obtain
fragment
BamHI(+5016)-HindIII (AME),
whichcorresponds
tothe linear formofpAME
(17).
Recombinants between U5-5' gag and the env gene weregenerated by
inserting
segmentsfromU5-5' gag betweentheHindlIl
and BamHIendsof the AMEfragment.
(i)
pAMSLE
andpAM(5L)E.
The SacI(+ 147)-BssHII
(+488) fragment
wasisolated frompAM5G
andlinkedtothe AMEfragment.
Theresulting plasmids
with sense and antisense insertions werecalledpAM5LE
andpAM(5L)E,
respectively.
(ii)
pAMLE. The HaeII(+286)-BssHII (+487) fragment
was
ligated
toAME.(iii)
pAM5E.
Similarly,
the Sacl(+147)-HaeII
(+290)
sequenceswereinserted 5' to theenv
region.
(iv) pAMa+E.
Synthetic
double-stranded DNAfrom po-sitions +322to+386 with 5'HindIIIand 3' BamHIcohesive endswas linkedtoAME.(v)
pAMa+,3E.
TheBamHI(+5016)-ApaI
(+5206)
region
of
pAMa+E
wasreplaced by synthetic
DNAfrompositions
+410to +471with 5' BamHI and 3' ApaI
sticky
ends.(vi)
pAMPE.
pAMa+,BE
was cleavedby
HindIII
and BamHIto remove theat
sequencesandwasthen recircular-ized.Plasmids
containing
the gag sequences were cleavedby
SalI,
and those with the env sequences were cleavedby
AvaII;
theplasmids
werethenphenol
extractedandethanolprecipitated
asdescribedaboveto serveasthetemplates
for the RNAsynthesis
reaction.Gelmobility shift assay. The RNA
gel
mobility
shift assay wasperformed
by
modificationof the methodsdescribed for the HIV RevRNA-binding
protein
(14,
27)
and those for varioustranscription
factors that bind to theirtarget
DNA sequences(22). Briefly,
RNAsolution in 2xbinding
buffer VOL.67, 1993on November 9, 2019 by guest
http://jvi.asm.org/
BLV
A U5-5gag
(Sac e
+147
r
PBS leader
L--~~~~~~~~~~~
---B cDNAssynthesized
4A4 9A4 9.2 4I1
tt:
3. 3.39.3 9.1 1.2 72 1.4 3.2 34
13
C Dimer formation
D MA(p15)-binding
.. :] 3~~~.
FIG. 1. Summaryof cDNAprobingassays.(A)Structure of the BLV U5-5'gagRNAfragment.RNAencompassingtheSacIsiteat +147
totheSalI siteat +936wasinvitrosynthesized. R, U5, PBS, leader,andgagregionsaremapped. The 5'splicedonorsite inthegenome RNA is 47 basesupstreamfrom the 5' end of the U5-5'gagsegment.(B)cDNAprobes. DNAssynthesizedforprobingassaysareindicated
by bars placedintheir targetsequences.cDNA 4.4wascomplementaryto+95to +124,whichwerenotcontainedinthe RNAfragment. (C) Inhibitionof RNA dimer formation. cDNAs that interferedwiththeRNAdimer formation in at leastoneofthetwo sets ofprobingassays areshownbyfilled bars. Probes9.1, 4.1, 1.1, 7.2,and1.3hadsequencescomplementaryto +245 to+274,+275 to+304,+322to+339,+340
to+369,and +350to+379, respectively. (D)Alteration ofthe RNAmobilityshift. cDNAs1.1,7.2, 3.1,and3.2,which influencedcomplex formation with MA(p15), are indicatedby dotted bars. Probes 3.1 and 3.2 were complementary to +410 to +439 and +440 to +469, respectively.
wasmixedwithMA(pl5) proteinsolutionataratio of 1:1on
ice and incubated for 10 min at 20°C. One volume of the loading buffer (30% glycerol, 0.02% xylene cyanol, 0.02% bromophenol blue)wasaddedper5 volumes of the reaction mixture,from which 3 plwasloadedperslot. Sampleswere runfor 3 hat120 Vat4°C.Theelectrophoresis gelwasmade of2.5%polyacrylamide and0.5%agaroseinTBEbuffer(90 mMTris-borate, 2 mM EDTA [pH 8.0]).
AssayresultswerenotaffectedeitherbyNaCl concentra-tion in thebindingreaction withinarangefrom10to50mM
orby pHbetween 7.5 and 8.0. Thepresenceof100mMNaCl reduced the band shift efficiency of U5-5' gag by 30% without relaxingthebinding specificityofMA(pl5).
Prediction of theBLVRNAsecondarystructure.
Minimum-energysecondarystructureswereconstructedbyoperating a SUN computer with programs FOLDRNA and SQUIG-GLES supplied by the University of Wisconsin Genetics Computer Group. BLVsequence datawere fromreference 38.
RESULTS
cDNAprobingassays.To determine thefunctionalmapof the U5-5'gagRNA for dimerization and MA(p15)-binding activities,weappliedasimplecDNAprobingmethodonthe basis of the following assumptions: that short single-stranded DNA probes complementary to the U5-5' gag
sequencescan hybridizeto the nascent RNA chains
gener-atedbythepolymerasereaction,which results inmasking of the activesequences and/or disruption of theinherent
fold-ing by imposition of an RNA/DNA double helix; and that subsequentelectrophoretic analysisincombination with the proteinbindingreactionpermits theestimation of the active regions.TwosetsofcDNAprobingassays werecarriedout asdescribed in Materials andMethods. cDNAprobes of18 or30basesweresynthesizedtoscantheRNAsequences as
shown inFig. 1. Probe 4.4wasdirectedto30basesin the3' partofthe Rregion,whichisnotcontained intheU5-5'gag
RNA andtherefore usedas aninactive cDNAprobe.
In thefirst assay, eachcDNAwas 32Pphosphorylated at
the 5' terminus and added to the SP6 RNA polymerase reaction. Recovered RNA was electrophoresed through a
nondenaturing gel to monitor dimerization and complex formation efficiency. Typical results are shown in Fig. 2A. The cold U5-5' gag RNA probed with various 32PcDNAs (odd-numbered lanes with the exception of lanes 1 and 9) appeared as dimer and monomer bands at the positions corresponding to those of the internally labeled U5-5'gag
RNA (lanes 1 and 9). Probe 4.4 failed to forma detectable
band(lane17). Unhybridized probemolecules madeabroad bandatthe bottom of eachlane.
As described previously, the U5-5'gag RNA efficiently
dimerizedtoform the dimer andmonomerbands inaratio of
4:1onthe basis of theradioactivitycountineachband(lane 1). Hybridization ofprobes 1.1, 7.2, and 1.3 (lanes 21, 23, and 13) affected the RNAdimerization to alter theratio of dimer/monomer populations to 1:4, 1:5, and 1:1,
respec-tively. Other cDNAs didnotappearto impair dimer forma-tion, althoughtheir hybridization efficiencies were not sig-nificantly differentfromthoseofthethreeinhibitory probes,
ascompared bythebandintensities.
After incubation with the MA(p15) protein, U5-5' gag
formed a shifted band presumed to contain the U5-5'gag
dimer(Fig. 2A, lanes 2 and10). Itis thought that MA(pl5) hasanRNA-annealing activity concomitant with the binding
activity (17). Bymild heattreatmentofthe RNA priortothe proteinbinding reaction, both the dimer andmonomershifts
becamedetectable in thesamelane,asdescribed previously.
On a 3.5% polyacrylamide gel, the two band shifts were
detectedby 30% reductionin theirmobilities. In aparallel
experiment on a 2.5% polyacrylamide-0.5% agarose
com-posite gel, their shifts were found as 60 to 70% slower migration than for the free dimers and monomers. From
theseobservations, shifted bands detected in thisstudywere
judged bytheir relativemobilitiestorepresenteither dimer
ormonomercomplexes.
Many of the
[32P]cDNAIRNA
hybrids normally reacted withtheprotein,asjudged by theappearanceof the shifted(SaI)
+940
I
|~ R U5
71
gagon November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.77.536.71.233.2]DIMERIZATION AND gag PROTEIN BINDING IN BLV 1833
A
SC-U5-5g;g 9 2 91 12 -3.J3; 933 :,
MA(D15) - + + + +
sd- g g
d- _
m-UUI
--'P WNA
2 3 4 5 6 7 t1
4.4 4.1 1.1 7.2
MA(p15)- + -+ + +
id-g
17 11 2 13 1. '
73 3.1 3.2
_ _
* .
._
B
4 9.1 92 93
-MA(p15 - + _ +
-bd- _S_J*
m-4
2 3 4
FIG. 2. Resultsof cDNA probingassays.(A) [32P]cDNA probing
assay. Unlabeled US-S' gag synthesized in the presence of
32P-labeledcDNAwasanalyzed bypolyacrylamide-agarose gel
electro-phoresis after incubation with (lanes +) or without (lanes -)
MA(p15). Control experiments with 32P-labeled US-S' gag are
shownin lanes 1, 2, 9, and10. Input cDNAis indicated for eachset
ofexperiments. Positions of dimers (d),monomers(m), andshifted
dimers (sd) ofU5-5'gagRNA are indicated. The position offree
[32PJcDNA is also indicated. Lanes are numbered at the bottom.
Binding of 9.3,9.4, 3.3, and3.4did notinterfere with either dimer
formation or protein binding. (B) Probing assay with unlabeled
cDNA.Resultswith probes4.1, 9.1, 9.2, and 9.3aswellas results
ofacontrol experiment(-)are shown. Probes 4.1and 9.1 impaired
thedimerformationof32P-labeledUS-S'gagand also affectedband
retardation. Results with probes 9.2, 9.3, and other cDNAswere
similartothose shown in panelA.
dimer band (Fig. 2A, lanes 4, 6, 8, 12, 16, 20, and 26). In contrast, U5-5' gag hybridized with probe 1.1 or 7.2 exhib-ited unusual electrophoretic features after incubation with MA(p15). As observed in lane 22, the dimers of the RNA/1.1 hybrid shifted normally, while the monomers stayed at the freeposition. The
RNA/7.2
hybrids made noclear bandshift and formed a diffusion through lane 24. Another notable profile was observed with probes 3.1 and 3.2 (lanes 27 to 30). Although dimers were formed in the same efficiency as in the controlU5-5'
gag RNA, disruption of the shifted band was detected (lanes 28 and 30). An extended smear was formed along lane 30.In the second cDNA
probing
assay, the RNA polymerase reaction contained[ax-
2P]CTP
as a labeled nucleoside triphosphate and unlabeled probe cDNA so that effects of the input probe were monitored by the radiolabel within the body of the RNA. Resultsobtained
in this probing method were similar to those in the first experiments; however, probes 4.1 and 9.1 obviously inhibited dimerization (Fig. 2B, lanes 1 and 3). The dimerized RNA in the presence of probe 4.1 or 9.1 was observed to form complexes with MA(p15). The monomers were less reactive than the control mono-mers, and long smears were found between the shifted dimers and free monomers (lanes 2 and 4). Probes 4.1 and 9.1 had a complementary stretch within their sequences, which may have resulted in a different binding feature after a phosphorylation reaction containing spermidine.From the results of the two cDNA probing experiments, dimer formation was suspected in the sequences covered with probes 9.1, 4.1, 1.1, and 7.2. Since probes 1.3 and 7.2 had 19 bases in common, the weak interfering activity of probe 1.3 was represented by probe 7.2. The MA(p15) binding was expected to involve the dimer-forming regions and the sequences covered with 3.1 and 3.2 (Fig.
1C
and D). For convenience, the 7.2-, 3.1 + 3.2-, and 4.1 + 9.1-directed regions are referred to as cx,,B,
and -y, respectively.Deletion
analysis
of U5-5' gag. To confirm that thea, 1,y,
and PBS regions are related to dimer formation and complex formation, a deletion experiment was carried out. Structures of the mutant RNAs synthesized are illustrated in Fig. 3A. In the structure names, dP,dot,
anddo
are used to represent specific deletions as follows: dP, deletion of the3'-terminal
17 or 12 bases ofUS
and the adjacent 18 bases of PBS; da, deletion of the 25 bases of the otregion; andd,B,
deletion ofthe sequences from +373 to +487 harboring the 1 region. The wild-type and mutant RNAs were
synthesized
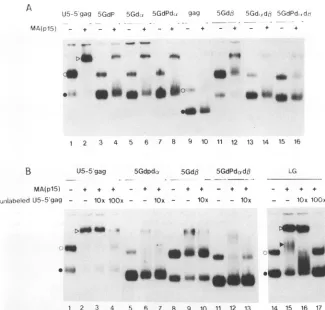
with 32p label in parallel reactions and analyzed for their dimerization and MA(p15)-binding abilities.Only 20% of the RNA molecules in the 5Gdcx, 5GdP, and 5GdPdcx preparations were dimerized (Fig. 4A), suggesting that removal of either the PBS orcxregion results in a loss of the dimerization potential ofU5-5'gag. These mutant mono-mers did not make a discrete band shift after the MA(p15) binding reaction (lanes 4, 6, and 8), while the dimers were able to make a clear shift as did the wild-type RNA (lane 2). Deletion of the 13-containing region did not alter the dimerization efficiency (lane 11). However, only 40% of the 5Gd1 dimer population participated in the complex forma-tion with MA(p15) (lane 12). Moreover, the dimer complex did not appear as a firm band. These results indicate that the 1 region is not required for RNA dimerization but may be involved in the protein binding reaction.
5Gdcxd1 and 5GdPdcxd13 also poorly dimerized (Fig. 4A, lanes 13 and 15). Their dimer shifts (in lanes 14 and16) were not solid as those of 5Gdcx, 5GdP, and 5GdPdcx.Thecontrol gag RNA made no band shift after the protein binding
VOL. 67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.72.306.109.560.2],+147 U5-5'gag I R_
HaeIBStNI B IIAva I
rU
I U5 l--I
PBS
Bs8 Dra +940
r+417
-I-
~~
gaggag-
1-i R
I
R
I
zR
I
RR
I
R+245 +304 +340 +369 +410 +46
r- -I r r
'Y Of0a
+302 r+341 U5
I...=~
+370 30 ...
U5 [...
U5
U5
69
gag
Wag
IHz
ITh:
+3M +488
+344,
.
. [image:5.612.134.469.74.422.2]+302.
...
:1
ga"
r+302ga
,+535
ga
IPr-Bam H Aja Ava I
1 +5501
L2IZ
envII
L
R r +491a,ll
It
+147,
l
gn
I
u [*{i1L
fwIIi1Ii
r+2_IP
E~Z
LZIrIIIII
Lz~cI
1111
env
1
R
+289-i .
...
env
l er r+322 +386, r+410 .471, r5203M~~~~~~~C~
a
lZ~EIIVII
W
...-
envI
It
FIG. 3. Structures of deletion and fusion mutants.Wild-typeU5-5'gagismappedforU5, PBS, leader,andgag regions. Positions of restrictionenzymecleavagesites used for theconstructionof mutanttemplatesareshown. y,PBS,a, and regionsare also indicated. (A)
RNAs with various deletionsin the leaderand/or5' terminusof thegagregion.5GdP, SGda, 5GdPda, 5GdI, 5GdadI3,5GdPdadp,LG,and gag were synthesized by SP6 RNApolymerase reactions using as templates DNAs pAM5GdP, pAM5Gda, pAM5GdPda, pAM5Gdp, pAM5Gdadp3, pAM5GdPdadI3, pAMLG, and pAMgag, linearized by SalI digestion. Deleted sequences are indicated by dotted lines. Nucleotide positionsarealso indicated.(B)Recombinant RNAsmade between the 5'regionand theenvregion.E,5LE,(5L)E,LE,Pa+ ,E, Pa+E,andPEweresynthesizedfromtemplates pAME, pAM5LE,pAM(5L)E, pAMLE, pAMPa+,3E, pAMPa+E,andpAM,3Ecleavedby
AvaI. Linkages generated bygenemanipulationareshownbydotted lines. 5'-terminalsequencesarealignedwiththewild-typeU5-5'gag
shown above.
reaction (lane 10), although the dimerandmonomer bands appearedtobeslightlysmeared.
Thus,in agreementwiththeresults obtained in the cDNA probing assays, the deletion analyses (Fig. 4A) suggested that the PBS and a regions contribute primarily to dimer
formation and that a sequences contribute to MA(p15) binding.
To compare the affinities for MA(p15) amongwild-type andmutant RNAs, we next performed competition assays
with unlabeled U5-5'gagRNAasdescribedpreviously (Fig. 4B). Inthewild-type RNA, 90and 40% of the total radioac-tivities were retained in the shifted dimer band in the
presence of 10- and 100-fold excess amounts of the cold competitor, respectively (lanes 3 and 4). In contrast, the complexes of 5GdPda, 5GdP, and 5GdPdaodI3 RNAs were
effectively dissociated by a 10-fold concentration of the wild-typeRNA. Less than 5% of theinput 32pcountswere
found in the retarded bandin lanes 7, 10, and 13.
Anothermutant,LG, consistingof the whole Lregionand thegagsequences,hadaninteresting phenotype.The rate of
dimerization (Fig. 4B,lane 14)was comparable tothoseof 5Gdat, 5GdP, and their relatedmutants. Despite the lackof sufficient dimerization activity, the LG RNA sustained a
moreintensereactivitywithMA(p15)thandid5Gdot,5GdP,
5Gd,B, or the related mutants. The LG RNA made an
additional band of slowermigration representing the mono-mercomplexaswellasthe band ofshifted dimers (lane15). Inaddition,one-fourth of theinputmonomerpopulationwas
incorporatedinto the dimercomplexesafterincubation with MA(p15);the dimercomplexinlanes 15 and 16 containedan
increased 32P radioactivity. Moreover, the LG
dimer-MA(p15) complex was retained in the competition assay
with 10-fold concentration of the wild-type RNA (lane 16) butnotthemonomer-MA(pl5) complex.Theyregionlikely
participates in the RNAdimerization but is lessresponsible forMA(p15) bindingandtheMA(p15)-induced RNA dimer-ization.
Experiments with recombinant RNAs. We tested whether the y, PBS, a, and ,B sequences can exert their functions when linked 5' to the env sequences that are not directly A
5GdP 5Gda
5GdPda
5Gdf
5Gdadf
5GdPdad,O
LG
gag
B E
5LE (5L)E
LE
5E
PcxOE
Poi+E
OE
u
WV
n
11
on November 9, 2019 by guest
http://jvi.asm.org/
DIMERIZATION AND gag PROTEIN BINDING IN BLV 1835
AU5-5gag 5GdP 5Gdit 5GdPd/ gag 5GdrJ 5Gd1dr: 5GdPd ,d,,
MA(p15) - + - + - + - + - + - + - + - +
--wo 4
64.
t0 _ * _a
ea so
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
U5-S gag 5Gdpdci 5Gd3 5GdPdad3
MA(p15} + i + - + + + +
-wilabeledU5-5gag - - lOx lOOx - - lOx - - lOx - - lox
LG
+ l 1 - - 10K lOOx
Fi> > > >
Z~~~~~~~~-
0 ; Zseem
00 ** sa-.-41 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
FIG. 4. Analysis of deletionmutantsfor dimer formationand complex formation with MA(p15). (A) MA(plS)bindingassaywithmutants
havingdeletions in the PBS-leader region. The wild-typeandmutantRNAswereincubated with (lanes +)orwithout (lanes -) MA(plS) (25
,uM)andanalyzedon anativegel. Positions of dimersandmonomers areindicated byopenandfilled circles,respectively, for theU5-5'gag
andgagRNAs(lanes1and9).The complex of the U5-5'gagRNAdimerand MA(p15) is indicated byatriangle in lane 2. (B) Band shift and
competitionassays.U5-5'gagandmutantRNAs with deletionsin the PBS-leaderorR-U5 regionwereexamined for MA(p15)-binding ability.
In the competition assays, concentrations of the wild-type competitor, U5-5'gag, are indicated as 1Ox (10-fold) or 10Ox (100-fold) in
comparisonwith that of the labeled RNA. -,reactions without thecompetitor.
relatedtothe packaging mechanism.Aseriesof recombinant RNAssynthesized for thispurposeis shown in Fig.3B, and the results oftheMA(pl5) bindingassay areshown in Fig.5.
Thecontrol envRNA, E,waspoorly dimerized, and it did not make a discrete band shift after the MA(plS) binding
reaction(Fig. 5A,lanes5 and6).
The 5LE RNA with a sense insertion of the 5'-terminal portion ofU5-5'gag formed a solid dimerband containing
40% of the total3ZPradioactivity(Fig. 5A,lane7) and made
aclear dimer shift(lane 8)similartothat ofU5-5'gag(lanes
2to4).Themonomershiftwasnotdetectableas afirm band.
Therecombinant RNA retained theprotein-binding activity in thecompetitionassaywitha10-foldexcessamountof the cold U5-5' gag RNA (lane 9) but not with a 100-fold
concentration of thecompetitor (lane 10).Incontrast,(5L)E with the opposite strand of the U5-leader region was
defi-cient in both self-dimerization and complex formation with MA(pl5) (lanes 11 to 14). These results indicate that the positive strand of the U5-leader segment from positions +147to+491 bears minimum structuralrequirementsfor the
twoRNA functions.
Neither the LE RNA containing the complete leader
sequencesnor5Ewith theU5 regiondimerized sufficiently (Fig. 5B, lanes 1 and9) in comparison with 5LE (Fig. SA, lane 7), ensuring the cooperation of U5 with PBS-leader
region in viral RNA dimerization. LE could make the
monomer and dimer band shifts firmly; shifts were,
how-ever, prevented by the 10-fold concentration ofU5-5'gag
(Fig. SB,lanes 1 to4). ThePa+1E RNA with PBS-a and 1B elementsinatruncated formmadeaband shift profile similar tothat of LE(lanes5to8) but less efficiently.
Dimers of5E behaved like those of5GdPdad3 (Fig. 4B, lanes 11 to 13) after the protein binding reaction (Fig. 5B, lanes 9 to 11), while monomers smeared through lane 10.
Neither PotaE nor ,BE formed a visible dimer band. The
Pa+Emonomersfailedtogiveaclear shift in lane13,while
the ,BE monomers could form a broad and diffuse band
barely separatedfrom theunboundmonomerband(lane 16)
in the absence of thewild-type competitor. The results in Fig. 5B indicate that each of the small elements in the U5-PBS-a-,B regionispoorlyreactive toMA(pl5).
It should be noted that thegag-tailed RNAs, 5GandLG,
were significantly more active in dimerization aswellas in complexformation thanwere the correspondingenv-tailed RNAs,5LEandLE, respectively.Since thegagregionfrom position +535to +940 itself hadadetectabledimer-forming naturebut didnotcause theband shiftwith MA(pl5) (Fig. 4A,lanes 9 and 10), itislikelythat thegagRNA structure influences the overallRNAfoldingandthereby supportsthe upstreamRNA functions.
B VOL. 67,1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.148.473.77.387.2]A
U5-5'gag E 5LE (5L)E
MA(p15) - + + + - + - + + + - + + +
unlabeled U5-5 gag - - lOx100x - - - - lOx100x - - lOx100x
... A
_0
0 as*, d
1 2 3 4 5 6 7 8 9 10 11 12 13 14
B
LE Pt3dE 5E PL+E 3EMA(p15) - + + + - + + + - + + - + + - + +
unlabeledU5-5'gag - - lox10Ox - - lOx 1OOx - - lox _ - lOx - - lox
0~~~~~~~
[image:7.612.157.440.349.754.2]1 23 4 56 78 910 11121314 1516 17
FIG. 5. Analysisofrecombinant RNAs for dimerization andMA(p15)-bindingactivities. (A)Results for theparentalRNAs,U5-5 gag, and Eand for recombinantRNAs,5LE,and(5L)E; (B)results for other recombinantscontainingshortsegmentsfrom the5'-terminalregion
andthebodyoftheenvregion.Themonomerbandof eachRNAis indicatedbyafilled circle. Detectable dimer bandsaremarkedwith open circles. Shifted dimers andmonomers areshownby trianglesandfilledtriangles,respectively. SampleswereincubatedwithMA(p15)(+)or
in buffer alone (-). Concentrations of thecompetitorU5-5' gagareindicated as lOx (10-fold)or 1OOx (100-fold) relativetothe testRNA concentration.
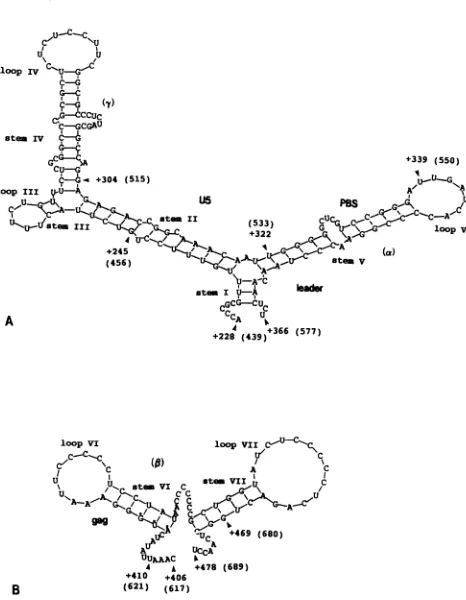
Computer-assisted analysis of BLV 5'-terminal RNA fold-ing. The dimer-forming and MA(p15)-binding sequences
determined above were examined for their formation of
secondarystructures by free-energyminimizationanalysis. As shown in Fig. 6A,abranchedstem-loopstructurewas
predictedintheregion encompassingU5tothe axregion. In
theprediction,theU5sequencesformlongstemsII andIV,
a shortstemIII,and associated bulges andloops. ThePBS andasequencesformstemV. Three basepairingsbetween the5' terminus ofU5and the3' terminus ofamakestemI.
In the region,twoseparatestem-loopstructuresVI and VII, consistingof residues +417to +437and +447to+471, respectively, were predicted (Fig. 6B). Stem-loop VI is
complementary to cDNA probe 3.1 and stem-loop VII is complementary to probe 3.2, which may account for their inhibitory effectonthe MA(pl5) bindingreaction(Fig. 2A).
DISCUSSION
Dimerization of BLVandother retrovirus RNA.We have examined thedimer-forming feature of the 5'-terminal BLV RNAbyacDNAprobing method and mutation experiments
with theU5-5'gagRNAfragments.Allof the experimental
results maybe interpreted asimplying that RNA
dimeriza-tion ismediatedby theregion encompassing U5, PBS, and the 3'flanking30 bases of PBS(a).
Computer-assisted RNA sequenceanalysis suggested for-mation ofasecondarystructureofastem-loopwithmultiple branches throughout the entire region determined. In that model,the ysequencescontributetostem-loops II, III, and IV. Elevenresidues of PBS and one base of its 5' adjacent region arefoundtopairwith thealsequencesto makestem V. Because retrovirus RNA dimerization is assumed to result fromintermolecular basepairingsduetothe presence ofpalindromic sequences (8), the model seems to support ourdata.
Stem-loop V is analogous to the secondary structure proposed for avian retrovirus RNAs, in which the 3' U5-PBS region hybridizesthe 5'-terminal leader sequences to build up a more extended stem without branching (2, 13). The involvement of thePBS-aregion in BLV RNA dimerization agreeswith the dimer modelpredicted for avian and murine viruses in earlierstudies (8).
We have alsoanalyzedHIV RNAfor dimer formationby cDNAprobing. The preliminary result shows that the 5'-terminal extended stem-loop II of the reported secondary
on November 9, 2019 by guest
http://jvi.asm.org/
DIMERIZATION AND gag PROTEIN BINDING IN BLV 1837
loop IV
stemIv
loopIII
(7)
(515)
U5
47
+245 (456)
PBS
(a) stemV kader
A
A0A +478 (689)
+410 +406
B (S121) (617)
FIG. 6. RNA secondary structures predicted for the dimer-forming andMA(p15)-binding regions by computer-assisted energy
minimization analysis. (A) Multibranched stem-loop structure
formed by thedimerization sequences +228 to +366. Nucleotide numbers originate from the RNAcapsite(+ 1). Position numbers in
thesequenced provirusgenome(38)arealso shown in parentheses.
Stems ItoIVandassociating loopsareindicated. Thestart(+245)
and end (+304) positions of the -y region and ofPBS (+322 and
+339) aremarked. Nucleotide numberings in the proviralgenome arealsoindicated in parentheses. (B) Twostem-loops found in the ,B
region. Sequences from +406to+478wereanalyzed for secondary
structure. The AUG translation start codon of thegag gene is marked by dots. The ,B region encompassed nucleotides +410 to
+469. Stem-loopsVIand VIIareindicated.
structure in the HIV leader (12) is probably involved in dimerization and thatU5 andPBS maynotberequired.
The MuLV dimerization site has been reported to be presentwithin the
*
sequences,where a double stem-loop was predicted (37). However, the folding of the recent prediction deviates from the long-standing model (8) andseemstobecomparabletothe BLV structurewithregard
tobothlocation and overall structure.
BLV RNA dimerization and primer
tRNAPr'
binding. In retrovirusparticles,tRNAprimerbindingiscompatiblewithgenome RNA dimerization (8). A primer-associated RNA dimer model, as well as a primer-free dimer model, was
proposedfor avian andmouseretroviruses(8,13). However, it is inferred fromourcDNAprobingand deletion analyses
that BLV RNA dimerization is competitive with tRNAPr, binding; bindingof cDNA 1.1 and deletion of PBSinhibited the dimerization. Also, it should be noted that the 3'-terminal primer tRNAsequences are locked inthe charac-teristic cloverleafstructurebyformation of the acceptorand TgCstems. Theprocessof tRNAannealingmaybea more
sophisticated dynamicreaction than hasbeeninterpreted by
earlier experiments (35). Unfolding of the genome RNA dimer structure, inadditiontounwinding of the tRNAstems (11), might beinvolved.
RNA structures required for MA(pl5) binding. Deletion and chimericmutationanalyses implicatethe region encom-passing PBStothe5' end ofthegag sequences, PBS-a-,B,as sequences essential for thespecificinteraction withMA(pl5) tomake acomplex that isdetected as adiscrete band in the gel retardation assay. The 1 region was predicted to form twoisolatedstem-loops VI and VII, while the PBS-a region likely forms stem-loop V, involved in the dimer conforma-tion asdescribed above.
It is possible that PBS-a and ,B cooperatively make a
three-dimensional conformationactive in protein binding. It is alsopossible that the PBS-a region in concert with thegag sequences exhibits the dimer-forming activity and thereby increases the affinity of the a structure to the protein.
BLV U5-5' gag-MA(p15) complex formation and genome packaging. Although there has been no direct information about BLV genome RNApackaging, our in vitro character-ization of U5-5'gag-MA(p15) complex formation is related to the accumulated experimental results concerning the genome encapsidation in MuLV and other retrovirus sys-temsfromtheviewpoints described below.
PBS-ox-1 covers the BLV RNA region analogous to the packaging sites determined for MuLV (28, 41), Rous sar-coma virus (33), avian sarcoma-leukosis virus (18, 43), spleen necrosis virus(45), and HIV (3, 23). Recent observa-tion that gag sequences are also included in the signal sufficient forheterologous RNA packaging into MuLV and Rous sarcoma virusvirions (1, 5-7) supports our results; the BLV 13 region contained the gag start codon and its 3' sequences. In addition, the downstream gag sequenceswere found to performfacilitating roles in the MA(pl5) binding reaction.
Further support is provided by the fact that deletions in the U5region of the MuLV genome resulted inblockingof the RNApackaging into the virus particles(32).Inthisstudy with BLV RNA, U5 was shown to contain a part of the region essential for RNA dimerization. Deletion of U5 resulted in a more than a 10-fold decrease in complex-forming ability (compare LG with U5-5' gag in Fig. 4B), although the RNA feature to be annealed upon MA(pl5) bindingwasretained. These notionsagreewith the explana-tion that the U5 region of MuLV may not be directly involved in the selective packaging of viral RNA but is requiredtomaintainthecorrectoverall structureof the viral genome (24).
BLV RNA dimerization appeared to lead to enhanced reactivitywithMA(pl5)protein.Conversely,MA(pl5) bind-ing often resulted inannealingofthe RNAmonomers.These observations could offer anexplanation forwhy retrovirus particles contain a70S dimer asthe genome.
With all of these in vitro results, it is necessary to determine at the cell culture level the RNA and protein sequencesrequired for BLV
packaging,
using
aDNA clone capable of infectious particle production or an equivalent assay system.There is disagreement concerning the function of avian retrovirus MA(pl9); the specific
RNA-binding
activity
de-scribed in earlier studies(20, 21)wasundetectable inrecentexperiments (42).
Furthermore,
detection of the specific interaction between MuLVppl2and thehomologous
RNA (39, 40) has never been repeated with other methods or furtherinvestigated
to determine theresponsive
RNA se-quences. Since retrovirus gag precursorprocessing
occursVOL.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.66.299.63.363.2]after formation of the immature core (19, 47), it is thegag polyprotein that first associates with the viral RNA. Unlike avian virus MA(p19) or MuLV ppl2, BLV MA(pl5) is the NH2-terminal cleavage intermediate generated by the Pr44gag processing (46) or the precursor of mature proteins MA(plO) and p4 (17).
The5'-terminal RNAstructureand virus replication. In the predicted folding of the 1 region (Fig. 6B), the gag start codon isfound in stemVI. Although the model needstobe validated experimentally, it is conceivable that the 5'-termi-nal conformation of the viral RNA and its interaction with cellular and viral proteins may determine the fate of viral RNAmolecules eithertoactasmRNA or to beencapsidated intoparticles asthe genome RNA. In HIV, the large folded structure lies immediately 5' to the AUG codon (12).
Manyfunctional elements for reverse transcription prim-ing, RNAdimerization, packaging, translation initiation and activation, and splicing nest in the 5'-terminal region of retroviral RNA. Some of these elements overlap each other orarein closeproximity, suggesting the possibility that one RNAfunction can regulate other activities. The 5'-terminal region may form an intricate three-dimensional conforma-tion. In this regard, retroviral RNA may be reminiscent of poliovirus RNA,whose 5'-terminal noncoding region forms a defined cloverleaf structure that controls translation (34, 44) and genome replication (4) by interacting with various cellular and viral factors.
ACKNOWLEDGMENT Wethank YokoSakamoto forBLVpreparation.
REFERENCES
1. Adam, M.A., and A. D. Miller. 1988.Identification ofasignal in amurine retrovirus that is sufficient forpackaging of nonretro-viral RNA into virions. J. Virol. 62:3802-3806.
2. Aiyar, A., D. Cobrinik, S. Ge, H.-J. Kung, and J. Leis. 1992. InteractionbetweenretroviralU5 RNA and the
T*C
loopofthe tRNATrP primer is required for efficient initiation ofreverse transcription. J.Virol. 66:2464-2472.3. Aldovini, A., and R. A. Young. 1990. Mutations of RNA and protein sequences involved in human immunodeficiencyvirus type 1packaging resultinproduction of noninfectious virus. J. Virol. 64:1920-1926.
4. Andino, R., E.Rieckhof, and D.Baltimore. 1990. A functional ribonucleoprotein complex forms around the 5' end of poliovi-rus RNA. Cell 63:369-380.
5. Armentano, D., S.-F. Yu, P. W. Kantoff, T. von Ruden, W. F. Anderson, and E. Gioboa. 1987. Effect of internal viral se-quences in the utility ofretroviral vectors. J. Virol. 61:1639-1646.
6. Aronoff, R., and M.Linial.1991. Specificity of retroviral RNA packaging.J.Virol. 65:71-80.
7. Bender, M. A., T. D. Palmer, R. E. Glinas, and A. D. Miller. 1987. Evidence that the packaging signal of Moloney murine leukemia virus extends into the gag region. J. Virol. 61:1639-1646.
8. Coffin, J. 1982. Structure of the retroviral genome, p. 261-368. In R.Weiss, N. Teich, H. Varmus, and J. Coffin (ed.), RNA tumor viruses. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
9. Gorelick, R. J., L. E. Henderson, J. P. Hanser, and A. Rein. 1988. Pointmutants of Moloney murineleukemia virus that fail
topackageviral RNA: evidence for specific RNA recognition by
a "zinc finger-like" protein sequence. Proc. Natl. Acad. Sci. USA 85:8420-8424.
10. Gorelick, R. J., S. M. Nigida, Jr., J. W. Bess, Jr., L. 0. Arthur, L. E. Henderson, and A. Rein. 1990. Noninfectious human immunodeficiency virus type 1 mutants deficient in genomic RNA.J.Virol. 64:3207-3211.
11. Harada, F., G. G. Peters, and J. E. Dahlberg. 1979. The primer tRNA for Moloney murine leukemia virus DNA synthesis. J. Biol. Chem. 254:10979-10985.
12. Harrison, G. P., and A. M. L.Lever. 1992. The human immu-nodeficiency virus type 1 packaging signal and major splice donor region have a conserved stable secondary structure. J. Virol. 66:4144-4153.
13. Haseltine, W. A., A. M. Maxam, and W. Gilbert. 1977. Rous sarcoma virus genome is terminally redundant: the 5' sequence. Proc. Natl. Acad. Sci. USA 74:989-993.
14. Heaphy, S., C. Dingwall,I. Ernberg, M. J. Gait, S. M. Green, J. Karn, A. D. Lowe, M. Singh, and M. A. Skinner. 1990. HIV-1 regulator of virion expression (Rev) protein binds to an RNA stem-loop structure located within the Rev response element region. Cell 60:685-693.
15. Jentoft, J. E., L. M. Smith, X. Fu, M. Johnson, and J. Leis. 1988. Conserved cysteine and histidine residues of the avian myelo-blastosis virus nucleocapsid protein are essential for viral rep-lication but are not "zinc-binding fingers." Proc. Natl. Acad. Sci. USA 85:7094-7098.
16. Karpel, R. L., L. E. Henderson, and S. Oroszlan. 1987. Interac-tions of retroviral structural proteins with single-stranded nu-cleicacids. J.Biol. Chem. 262:4961-4967.
17. Katoh, I., H. Kyushiki, Y. Sakamoto, Y. Ikawa, and Y. Yoshi-naka. 1991. Bovine leukemia virus matrix-associated protein MA(pl5): further processing and formation of a specific com-plexwith thedimer of the 5' terminal genomic RNA fragment. J. Virol. 65:6845-6855.
18. Katz, R. A., R. W.Terry,and A. M. Skalka. 1986. A conserved cis-actingsequencein the5' leader of avian sarcoma virus RNA isrequired forpackaging.J. Virol. 59:163-167.
19. Kohl, N. E., E. A. Emini, W. A. Schleif, L. J. Davis, J. C. Heimbach, R. A. F. Dixon, E. M.Scolnick, andI. S. Sigal. 1988. Active human immunodeficiencyvirus protease is required for viralinfectivity. Proc.Natl. Acad. Sci. USA 85:4686-4690. 20. Leis, J., S. Johnson, L. S. Collins, and J. A. Traugh. 1984.
Effects of phosphorylation of avian retrovirus nucleocapsid proteinppi2onbinding ofviral RNA. J. Biol. Chem. 59:7726-7732.
21. Leis, J., J. McGinnis, and R. W. Green. 1978. Rous sarcoma virus p19 binds to specific double-stranded regions of viral RNA: effect of p19 oncleavage ofviral RNA by RNase III. Virology 84:87-98.
22. Lenardo, M.J.,C.-M. Fan, T. Maniatis, and D. Baltimore. 1989. The involvement of NF-KB in 3-interferon gene regulation reveals itsrole as widely induciblemediatorofsignal transduc-tion. Cell57:287-294.
23. Lever, A., H. Gottlinger, W. Haseltine, and J. Sodroski. 1989. Identification of a sequence required forefficientpackaging of human immunodeficiency virus type 1 RNA into virions. J. Virol.63:4085-4087.
24. Linial, M. L., and A. D. Miller. 1990. Retroviral RNA packag-ing: sequencerequirements and implications.Curr.Top. Micro-biol.Immunol. 157:125-152.
25. Lowry,0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem.193:265-275.
26. Luban, J.,and S. P. Goff. 1991. Binding of human immunode-ficiency virus type 1(HIV-1) RNA to recombinant HIV-1 gag protein. J. Virol. 65:3203-3212.
27. Malim, M. H., L. S. Tiley, D. F. McCarn, J. R. Rusche, J. Hauber, and B. R. Cullen. 1990. HIV-1 structural gene expres-sion requires binding of the Rev trans-activator to its RNA targetsequence. Cell60:675-683.
28. Mann, R., R. C.Mulligan, and D. Baltimore.1983.Construction of aretrovirus packaging mutant and its use toproduce helper-freedefective retrovirus. Cell33:153-159.
29. Meric, C.,and S. P. Goff. 1989. Characterization of Moloney murineleukemia virus mutants withsingle-amino-acid substitu-tions in the Cys-His box of thenucleocapsid protein. J.Virol. 63:1558-1568.
30. Meric, C., E. Gouilloud, and P.-F. Spahr. 1988. Mutations in Roussarcoma virus nucleocapsid proteinp12(NC): deletions of
on November 9, 2019 by guest
http://jvi.asm.org/
DIMERIZATION AND gagPROTEIN BINDING IN BLV 1839 Cys-His boxes.J.Virol. 62:3328-3333.
31. Meric, C., and P.-F. Spahr. 1986. Rous sarcomavirusnucleic acid-bindingprotein p12 isnecessaryfor viral70SRNAdimer formationandpackaging. J.Virol.60:450-459.
32. Murphy, J. E., and S. P. Goff. 1989. Construction and analysis of deletion mutations in the U5 region of Moloney murine leukemia virus: effects on RNA packaging and reverse tran-scription.J. Virol. 63:319-327.
33. Nishizawa, M., T. Koyama, and S. Kawai.1985.Unusual feature oftheleadersequenceof Rous sarcomaviruspackagingmutant TK15. J.Virol. 55:881-885.
34. Pelletier, J., and N. Sonenberg. 1988. Internal initiation of translation ofeukaryoticmRNAdirected byasequencederived frompoliovirusRNA. Nature(London) 334:320-325.
35. Peters, G., and J. E. Dahlberg. 1979. RNA-directed DNA synthesis in Moloney murine leukemia virus: interaction be-tweentheprimertRNAand thegenome RNA. J.Virol. 31:398-407.
36. Phalguni,G., and J. F. Ferrer. 1980. Detection ofa precursor-likeprotein ofbovine leukemia virusstructural polypeptidesin purified virions. J.Gen. Virol. 47:311-322.
37. Prats, A. C.,C. Roy, P. Wang, M. Erard, V. Housset, C. Gabus, C.Paoletti, and J.-L.Darlix. 1990.ciselements and trans-acting factors involved in dimer formation of murine leukemia virus RNA.J. Virol. 64:774-783.
38. Sagata, N.,T.Yasunaga, J.Tsuzuku-Kawamura,K. Ohishi, Y. Ogawa, and Y. Ikawa. 1985. The nucleotide sequenceof the genomeof bovineleukemia virus: its evolutionary relationship tootherretroviruses. Proc.Natl. Acad. Sci. USA 82:677-681. 39. Sen, A.,andG.J. Todaro. 1976.Specificityofin vitrobindingof
primatetypeCviral RNA andthe homologous viralp12core
protein.Science 193:326-328.
40. Sen, A., and G. J. Todaro. 1977. The genome-associated, specificRNAbinding proteins of avian and mammaliantypeC viruses. Cell 10:91-99.
41. Sorge, J. D., D. Wright, V. D. Erdman, and A. E.Cutting. 1984. Amphotropic retrovirus vector system for human cell gene transfer. Mol. Cell. Biol.4:1730-1737.
42. Steeg, C. M., and V. M. Vogt. 1990.RNA-binding properties of the matrix protein (p199a9) of avian sarcoma and leukemia viruses. J.Virol. 64:874-855.
43. Stoker, A. W., and M. J. Bissell. 1979. Development ofavian sarcomaandleukosis virus-basedvector-packaging cell lines.J. Virol.62:1008-1015.
44. Trono, D., R. Andino, and D. Baltimore. 1988. An RNA se-quence of hundreds ofnucleotides at the 5' endofpoliovirus RNAis involved in allowingviral protein synthesis. J. Virol. 62:2291-2299.
45. Watanabe,S., and H. M. Temin. 1983.Encapsidationsequences forspleen necrosis virus,anavianretrovirus,arebetweenthe 5' longterminalrepeatand the startof the gag gene. Proc. Natl. Acad. Sci.USA79:5986-5990.
46. Yoshinaka, Y., I. Katoh, T. D. Copeland, G. W. Smythers, and S. Oroszlan. 1986. Bovine leukemia virusprotease:purification, chemical analysis, and in vitro processing of gag precursor polyproteins. J.Virol. 57:826-832.
47. Yoshinaka, Y., and R. B.Luftig. 1977. Murine leukemia virus morphogenesis: cleavage of P70invitrocanbeaccompanied by ashift fromaconcentrically coiled internal strand("immature")
to acollapsed ("mature") form of the viruscore. Proc. Natl. Acad. Sci.USA74:3446-3450.
VOL. 67,1993