0022-538X/91/031291-13$02.00/0
Copyright© 1991, American Society for Microbiology
Induction of Human Immunodeficiency Virus Type 1 Expression
in
Chronically Infected Cells Is Associated Primarily with a
Shift in RNA
Splicing Patterns
NELSONL.
MICHAEL,'*
PAUL MORROW,2 JOSEPH MOSCA,3 MARYANNE VAHEY,1 DONALD S. BURKE,4 ANDROBERTR.REDFIELD'
Departmentof RetroviralResearch1 and Division of
Retrovirology,
Walter Reed ArmyInstitute of Research, 13 Taft Court Suite 200, Rockville, Maryland 20850, and SRATechnologies2
andHenry M. JacksonFoundation,3 Rockville, Maryland 20850 Received 11 September1990/Accepted28 November 1990
We have analyzed the kinetics of human immunodeficiency virus type 1 (HIV-1) RNA induction in chronicallyinfected T cells and promonocytes. A substantial amount of splicedmRNAs and assembled virions wasfound in resting cells. Induction increased the steady-state level of total HIV-1 RNA by 4-fold but increased the level ofunspliced transcripts by 25-fold. This increase in unspliced RNA was reflected in the amount of virus seenby electronmicroscopy. These data suggest a mechanism for the induction ofHIV-1 RNA in chronically infected cells involving a shift in splicing greatly favoring the stability of unspliced viral RNA with only a modest increase in total viralRNA. Analysis of the relative abundance of transcript classes is critical to the measurement of HIV-1 viral replication kinetics.
We are interested in investigating the molecular mecha-nismsthatmediate viralgene expression in human immuno-deficiency virustype 1 (HIV-1)-infected cells. Experiments in cellsacutely infectedwith HIV-1 show an initial rise in the multiply splicedtranscripts, tat, rev, and nef, followed by a riseinthe level of both singlyspliced andunsplicedspecies (1, 10, 14,20, 27, 32). Tat increases viraltranscriptionfrom the5'long terminalrepeat (LTR) in these modelsystems(2, 23, 29, 39) and may facilitate elongation ofinitiated tran-scripts (25, 29, 37). Rev serves to increase the steady-state level of cytoplasmic unspliced viral RNA byeither directly interacting with the splicing machinery (5, 6) or increasing theefflux of thesetranscriptsoutofthenucleoplasm, where theyaresusceptibletobeing spliced (12,15,22,32). Thus, in models of acute infection, a mechanism of HIV-1 RNA metabolism can be demonstrated that functions early in infectionto express regulatorygenes and thenswitchesto a late phase ofexpression of both viral structural genes and genomicRNA.
In contrast to acute infections, less is known about the detailed molecular events of HIV-1inductionin chronically infected cells activated by cytokines, phorbol esters, and otherviruses (9, 13, 16, 17,41). Previous workhas showna five- to sixfold induction of virus by analysis of reverse transcriptase activityand coreprotein p24levels in culture supernatants, by immunoblot analysis of cellular protein, andby measurements ofreporter geneactivity from the 5' LTR. In situ hybridization studies inT cells demonstrated that less than 20% of
resting
cells werepositive
for viralRNA;
ofthese, only 2%
werestrongly positive (9). Among
these cells 10to 15%were positive byimmunofluorescence for viral proteins prior to induction, whereas 100% were positive postinduction.Thesedatawereinterpreted tomean that cells chronically infected with HIV-1 exist in a latent rather than a low-level-expressing state. RNA slot blot analysiswasconsistentwithamechanism of
transcriptional
*Correspondingauthor.
activation thatwasbasedon anincrease inthe steady-state levelofviraltranscripts originating fromthe 5' LTRregion (11).
We have investigated the kinetics of HIV-1 RNA induc-tion in chronically infected promonocytes and T cells by a combination of RNA hybridization studies, reverse tran-scriptase-polymerase chain reaction (RT-PCR) assays, and transcript end mapping. These data were correlated with measurementsof virionassembly by supernatant p24 antigen analysisand by transmission electron microscopy. Expres-sion ofviral RNA in unstimulated cells is substantial. These cellscontainthree-tofourfoldless HIV-1 RNAthan stimu-latedcells, but virtuallynoneofthese RNAs areunspliced. The molecular mechanism of HIV-1 RNA induction in chronically infected cellsisassociatedpredominantlywitha shift in splicing favoring unspliced (genomic) RNA. The amountofunsplicedviralRNA,nottheamountoftotalviral RNA, is correlated with the observed increase in virion assembly.
MATERIALS ANDMETHODS
Cell lines and plasmids. All cell lines and the plasmid pBH10were obtained from the AIDS Research and Refer-enceReagentProgram
(National
InstitutesofHealth Repos-itory). ACH-2 isanA3.01derivativeT-celllinecarrying
1or2 proviral copies per cell (9). Ul is a U937 derivative promonocyte line (17). Both can be induced to
produce
infectious viralparticles.
Cells were grown out of frozen seeds and maintained in RPMI 160 medium supplemented with 10%fetalcalf serum,glutamine, andstandard antibiot-ics in a5% CO2environment. Cellsgrowing
at adensity
of106/ml
were induced inmediumadjusted
to20nM phorbol-12-myristate-13-acetate (PMA;Sigma).
Cells were main-tained in this environment untilthey
were harvested. All experimentswereperformed
atleast twice with ACH-2 and Ul cells expanded out ofa frozen seed obtaineddirectly
from the National Institutes of Health
Repository.
These experimentswerethenconfirmedwith cells obtained froma 1291on November 10, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 10, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 10, 2019 by guest
http://jvi.asm.org/
1292 MICHAEL ET AL.
second frozen seed sent from the National Institutes of Health
Repository.
pBHGEM
wasconstructedby
subcloning
the 8.9-kbSstIfragment
ofpBH10
into the SmaI site ofpGEMSZf-(Promega
Biotec).
RNA
preparation
and blotanalysis.
Total cellular RNA wasprepared by
thechaotropic
salt methodessentially
as describedpreviously
(7).
Samples
of 5 or 10 ,ug wereelectrophoresed through
1.0%formaldehyde-agarose gels.
Any given
time course was contained on asingle gel
for accuratecomparison
between timepoints.
Gelsweretreated with 0.05 NNaOH for40 min at room temperature beforecapillary
transfer toHybond-N
nylon
filters(Amersham
Corp.)
to ensure uniform transferefficiency
of RNA mole-cules from 75 bases to over10,000
bases in chainlength.
Samples
forquantitation
were run intriplicate.
The blots were cross-linked with 121xJ
of UV irradiation and thenprehybridized
andhybridized
in a solution of 50% forma-mide-6x SSC(lx
SSCis 0.15 MNaClplus
0.015 Msodiumcitrate)-20
mM Tris-HCl(pH 7.4)-5x
Denhardt solution-250,ugof salmonsperm DNA per mlat42°C.
The[32P]RNA
probe
wasprepared
fromHindIII-digested pBHGEM
with SP6RNApolymerase.
RNAwaspurified through prepacked
Sephadex
G-50 spuncolumns(5
Prime-3Prime,
Inc.)
before use. After 12 to 16 h ofhybridization,
blots were washed twice for15minwith2x SSC-0.1% sodiumdodecyl
sulfate at room temperature and then twice for 15 min with 0.1xSSC-0.1% sodium
dodecyl
sulfate at50°C.
Filterswere
subjected
todirectautoradiography
for2to5 h at-80°C
with XAR-5 film(Kodak,
Inc.)
andLightning
Plus
intensifying
screens(Dupont).
Washed filters weredirectly
analyzed
for boundradioactivity
in aBetaScope
scanner
(BetaGen, Inc.)
witha5-log
linear range of quanti-tation. Theaverageoftriplicate signals
wasusedtocompare relativesignal
intensities.Signals
were normalized with aprobe
toP-actin
(Clontech, Inc.).
Oligodeoxynucleotide
synthesis
andsequences.Primersandprobes
weresynthesized by using phosphorimadate
chemis-tryonanApplied
Biosystems
DNAsynthesizer.
The nucle-otide sequences of theprimers
andprobes
used in theseexperiments
are as follows(BRU
sequence numbers aregiven
withinparentheses).
Primers:FPL-1,
5' GGCTAACT AGGGAACCCACTG3'(44 through 64); FPL-2,
5'GTCCC TGTTCGGGCGCCACTG3'(201
through 181);
NEF-2,5'CGAGAGCTGCATCCGGAGTACTT
3'(8975 through 8997);
TPL-2,
GCACTCAAGGCAAGCTTTATTGAGGCTTA 3'(9225 through
9197);
FPS-3,
5'GACGCTCTCGCACCCATC TC3'(354 through
335); E17,
5'GCTTTGATCCCATAAAC TGATTA 3'(6147 through 6125);
FPR-1,AAGCCTCAATAAAGCTTGCCTTGAGTGC
3'(66
through
93); TRN-2, 5'CTCTCTCTCCACCTTCTTCTTC
3' (8038 through 8017);RK-1,
5'CAGTGGCGCCCGAACAGGGAC
3'(179through199).
Probes:LTRP-3,
5'GCCTCAATAAAGCTTGCCTTG AGTGC 3'(68 through
110); NEFP, 5' TGCTTTTTGCCTGTACTGGGTCTCTCTGGTTAGACCAG
3' (9115 through9152); TPS-1,
5'GAAGAAGCGGAGACAGCGACGAAG
3'
(5558 through
5581); FPS-1,
5'GGCGACTGGTGAGTACGCCAAAAATTTTGAC
3' (282 through 312).RT-PCR assays.
Semiquantitative
RT-PCR assays wereperformed
on several incremental amounts of total cellular RNAtemplate.
All assays were done intriplicate. Samples of1.0,
2.5, 5.0,
and 10 ng of total cellular RNA were converted to cDNAby primer
extension with 10 pmol ofspecific
3'primer
with 200 U of cloned RNase H-Moloney murine leukemiavirusRT(BethesdaResearchLaboratories) for 1 h at450C
in a20-,d
reactionvolume. One halfofthereaction was amplified in a 100-pd reaction volume for 30 cycleswithaPCR scheme of 95°C for 30 s, 55°C for 30 s, and 72°C for 3 min. The amplified products were
separated
through horizontal 1% agarose gels, transferred to nylon filters, and probed with 32P-end-labeled oligodeoxyribonu-cleotides. Theprobesequencesrepresentedregionsinternal to butnotoverlappingwith the primersused for
amplifica-tion. Hybridization signals were quantified directly from filters in a Betascope scanner. To confirm that
amplified
signals were both RNA dependent and HIV-1 specific, control amplifications were performed without RT, with template pretreatment with DNase and RNase, and with uninfected H9 cell RNA. Quantitations were performed in the linear part of the amplification signal versus template inputcurve. RNAamounts were normalized by hybridiza-tion to a 3-actinprobe.
Nuclease Si and primer extension analyses. RNA end mapping was performed with nuclease S1 and radiolabeled RNA probes. Synthesis ofuniformly labeled, complemen-taryprobes was accomplished by in vitro transcription with SP6 polymerase from pJM123 (a pGEM-based vector con-taining nucleotides 300 to 760 of HXB2) and purification through a Sephadex G-50 spun column. Total cellular RNA (10 pug) was hybridized in 20 mM piperazine-N,N'-bis(2-ethanesulfonic acid) (pH 6.5)-80% formamide-2 M NaCI-5 mM EDTA with 1.5 x
105
cpm ofpJM123 probe in a10-pd volume at 50°C overnight. The annealed products were digested with 10 U of nucleaseS1 in a 300-pd volume at 37°C for 30min. Samples were precipitated with ethanol, dried in vacuo, suspended in formamide loading buffer, and heat denatured. Digested material was subsequently electro-phoresed through a denaturing polyacrylamide-ureagelat30 mA for 2 h before direct autoradiography. The protected RNAs were sized against a parallel lane containing end-labeled DNAfragments from an HaeIII digest ofpBR322.Primer extension studies were performed with an end-labeled oligonucleotide (O-JM76) that is complementary to nucleotides901 to 930ofHXB2.Total cellular RNA (5 to 10 ,ug)washybridizedin 12pI of 83 mMTris-HCl(pH
7.5)-125
mM KCl with 3 x 105 cpm ofprobe at 65°C for 1 h. The mixture was heated to 85°C for 2min, followed by 65°C for 2 min, and cooled to room temperature for 30 min. The primerwas then extended by the addition of 8 pl1 of RT buffer containing 9 mM MgCl2, 30 mM dithiothreitol, 150 ,ug of dactinomycin per ml, 1.5 mM deoxynucleoside triphos-phates,and 1 ,ul (200 U) ofMoloney murine leukemia virus RT. Thereactionwas incubatedat 37°C for 30min and then incubatedat42°Cforanother 30 min. The reaction products wereprecipitatedwith ethanol, dried in vacuo, suspended in 80% formamideloading buffer, heat denatured, and cooled on ice. Reaction products were electrophoresed through denaturing polyacrylamide-ureagels and analyzed by direct autoradiography.
p24 antigen analyses. Culture supernatants were diluted appropriately before p24 quantitation. Samples were proc-essedfor reactivity by using a commercially available anti-gen capture kit(AbbottLaboratories).
Electron microscopy. Cells to be analyzed by electron microscopy were grown to a density of 106 cells per ml beforeinduction withPMA. Samples were taken for analysis just before stimulationand after 10 and 21 h of stimulation. Cells were washed twice with cold phosphate-buffered sa-line,followed by fixation in 2.5% glutaraldehyde-phosphate-buffered saline on ice for 1 h. Samples were postfixed in 1% osmium tetroxide for 2 h, stained en bloc with 2% uranyl J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
A
A(C I-I1 ' T h 11
B
44-+k[
Ikh
Ul
2hr 4hr 22hr 28hr 53hr
H9 If IF
{
If'T
9.4kb
4.4
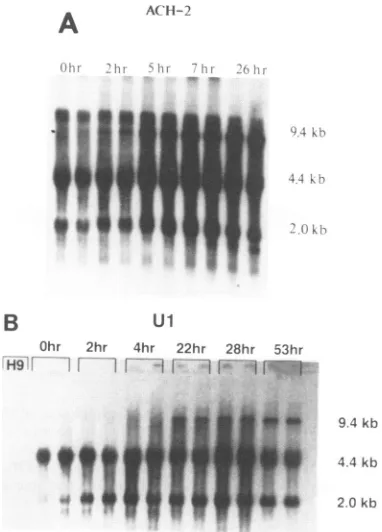
FIG. 1. Kinetics of HIV-1 RNA induction favors stability of
unspliced RNA. (A) Northern blot analysis of total cellular RNA
from ACH-2cells. Cellswereharvestedatthe indicated timesafter
exposure to PMA, and RNA was prepared by the chaotropic salt
method. RNA (10 p.g)was electrophoresed in each oftwoduplicate
lanes through a 1% formaldehyde-agarose gelfor each time point
shown, transferred toanylonmembrane,and subsequently probed with anRNAprobe commontothe 3' end of all HIV-1transcripts
(seeMaterialsand Methods). All signals shownwerefromthesame
2.5-h exposure ofa single nylon membrane. Hybridization signals
were quantified in a radioanalytic imager (BetaScope; BetaGen,
Inc.)and normalizedtoa
P-actin
probe. Theapparentmolecularsizeof each transcript class (shown to the right ofthe autoradiograph)
was determined from the comigration ofa known array of DNA
fragments and by the positions ofrRNAs and tRNAs in the total
RNA preparation. (B) Northern blot analysis of total cellular RNA
from Ul cells. This blot was performed essentially as in panel A.
Total cellular RNA (5
,ug)
from uninfected H9 cells waselectro-phoresedinparallelwith duplicate
5-,ug
samples from Ul cells. Allsignals represent those from the same 16-h exposure of a single
nylon membrane.
acetate, and examined in a Hitachi HU-12A transmission
electronmicroscope.
RESULTS
Kinetics of HIV-1 RNA induction favors stability of
un-splicedRNA.Westudiedthekinetics ofRNAexpression and
virion production in chronically infected cells induced by
PMA to elucidate the relationship between HIV-1 RNA metabolism andvirion biogenesis. Figure1showstheresults ofatypical blot analysis ofRNA extracted from cells over
the course of 1 to 2 days after PMA exposure. Gels were
treatedwith dilute sodiumhydroxide before transfer, which
resultedin auniform transfer efficiency ofRNAsfrom 75to over 10,000 bases in length. Blots were probed with a
32P-labeled RNA that represents the 3' genomic region
commontoall HIV-1transcripts. Transcriptsize classesare
therefore represented by their hybridization signals in a
manner directly proportional to their stoichiometry in the cellular RNA pool.
Figure 1Ashows a time course of HIV-1 RNA induction from PMA-stimulated ACH-2 cells. The 0-h lanesshow the steady-state levels of HIV-1 transcripts before stimulation. Although very low levels of 9.4-kb RNA were observed, substantial amounts of 4.4- and 2.0-kb RNA, corresponding to singly and doubly spliced transcript classes, respectively, were expressed. An increase in the 2.0-kb RNA level, but no change in the 4.4-kb RNA class, was observed after 2 h of stimulation. Unspliced transcripts had begun to accumulate after 5 h of stimulation. Singly spliced RNA had also begun to increase at this time. The amount of 2.0-kb RNA had reached a plateau after 7 h of stimulation, whereas the amount of the larger species continued to rise. The rate of increase of the unspliced RNA exceeded that of the 4.4-kb RNA. Themajordifference between the 7-hpoststimulation RNA pool and the pool representing 26 h ofstimulation was that the level of 4.4-kb RNA had reached aplateau,whereas the amount of unspliced 9.4-kb RNA had continued to increase. Although the 2.0-kb RNA species hadbegun to fall at 26 h, afaint band at 1.8 kb had become discernible. Little change in the RNA splicing pattern was observed whenthe length of stimulation was extended to 48 h. No HIV-1-specific signals were observed in parallel experiments with RNA extracted from an uninfected T-cell line (H9).
Quantitation of the ACH-2 data revealed afivefold rise in the steady-state level of totalHIV-1-specific RNA after 26 h of PMA stimulation. Unspliced RNAaccumulated to a level 20-fold higher compared with the unstimulated level. The rise in singly spliced RNA was fourfold over the same time period. The rise in the level of singly spliced RNA initially coincided with the rise in the level of the 2.0-kb species. However, the 4.4-kb RNA continued toaccumulate after the 2.0-kb RNA reached a new steady-state level. The doubly spliced transcript class attained a level of expression that was threefold higher than the basal level in this experiment. Analysis of the induction kinetics of HIV-1 RNA in Ul cells is shownin Fig.1B. Whereas the steady-state levels of spliced RNAs, sized at 4.4 and 2.0 kb, were substantial in unstimulatedUl cells, the level of unspliced RNA was only twice that of the background radioactivity on the blot by radioanalytic imaging. The level of 2.0-kb RNA doubled after 2 h of PMA stimulation, but the amount of the 4.4 kb species did not change significantly from the basal level. Both of the spliced transcript class levels were increased after 4 h of stimulation. Unspliced RNA had also begun to accumulate at this point. Furtherincreases in all specieshad occurred after 22 h ofstimulation. Unspliced RNA accumu-lated more rapidly than spliced RNA. Nofurther accumula-tion of 2.0- or 4.4-kb RNA was seenafter 28 hofstimulation. There was little additional accumulation of HIV-1 RNA at time points between 28 and 53 h ofstimulation. However, cell viability began to fall below80% after 48 h. Quantitation of the RNA induction kinetics from Ul cells indicated a threefold rise in total HIV-1 RNAover thetimecourse
(Fig.
1B). The 4.4- and 2.0-kb RNA sizeclasses rosethreefold, as well. Unspliced RNA accumulated to a level 25-fold over that of unspliced RNA in unstimulated Ul cells.
These data suggest that the induction of HIV-1 RNA in chronically infected T cells andpromonocytes is associated with only a modestincrease in the steady-state level of total RNA but a dramatic increase in the stability ofunspliced viral RNA. These datasuggest three major implications for HIV-1 gene expression. First, unspliced RNA
stability,
not de novo synthesis, is the dominant molecular mechanism0
q
". 4.0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.85.276.75.341.2]1294 MICHAEL ET AL.
FPLi FPL2 FPS3
U3|4--R41U
LUiiiEI-GAG1 GAG4
-_
4-FPR1 RK1
NEF2 TPL2
-_
4-U3 R U5
E4- T4- N4-E17 TRN2 NEF3
Genomic
env
1./
nef -
_-...-FIG. 2. TranscriptionmapofHIV-1 with location ofoligonucleotide primersforRT-PCRanalysis.Aschematicdepictionof the HIV-1
genomeisgiven aligned against the majormRNAtranscriptclasses. The location and orientation ofprimersareshownbyarrows.The LTRs
areshown as subdivided boxes. Intervening sequencesaregiven as dottedlines, andcoding sequencesare givenas solid lines for each
transcript class. FPL-1 and FPL-2aswellasNEF-2 and TPL-2amplifyall HIV-1 RNAs.GAG-1andGAG-4 detectonly unspliced transcripts. FPS-3 and E17 detect singly spliced transcripts. FPS-3andNEF-3coamplifythetat/revandnef doubly splicedmRNAsbutgiveadiscrete
signal for each. This figure isnotdrawntoscale.
associated with HIV-1 RNA induction in these cells. Sec-ond, the induction of multiply spliced RNAs temporally precedestheinductionofsinglysplicedRNAs. The increase in the levelofthese species is followed byasharpincrease
in the amountofunsplicedviral RNA. The kinetics of viral RNAinductioninchronically infected cellsrecapitulate the molecular events associated with HIV-1 replication and expression in acutely infected cells (1, 14, 27, 32). Third, contrary to the belief that these cells are virologically
quiescent (9),weobserve that the basal level of viral RNA in resting T cells and promonocytes is substantial. This RNA exists primarily in thesplicedform.
Expression of splicedmRNAs is induced to a lesserextent
than thatofunspliced RNA-analysis byRT-PCR assays. A complementaryapproachtotranscriptanalysiswastakenby using semiquantitative RT-PCR assays. Oligonucleotide
primers were synthesized according to the nucleotide
se-quencesflanking major splice donor and acceptor sites on
HIV-1 RNA as shown in schematic form in Fig. 2. This
techniqueenabledustodiscriminate between thetat/revand nef mRNAs owing to the slightly smaller region 5' to the envelope genethat isfound in the nef transcriptversus the
tat/rev species. The results of triplicate experiments in the
linearpartof theRT-PCRassay curve are givenasrelative
changesintranscript levelsatmaximal inductioncompared withthebasal level (Table 1).
PrimersFPL-1 and FPL-2amplifya161-bp fragment in the
5' LTR region that is common to all HIV-1 transcripts. NEF-2and TPL-2 amplifya 251-bpfragment inthe 3' LTR
region, which is also represented inall HIV-1 RNAs. Results with thesetwoprimer pairs showed a threefold increase in
total HIV-1 RNAafter induction with PMA in both ACH-2 andUlcells, consistent with the data obtained from
North-ernRNAblottingexperiments (Table1). GAG-1and GAG-4
amplifya216-bp fragment ofthegaggene,whichconstrains
thespecificity of this primer pairtounsplicedRNA. Bythis analysis, ACH-2 cells showeda29-fold increase in unspliced
RNA after 26 h of induction. Ul cells showed a 20-fold
increase in theamountofunsplicedRNAby thistechnique. This compares with 20- and 25-fold induction ofunspliced viral RNA inACH-2andUlcells,respectively, byNorthern blotexperiments (Fig. 1). The small differences obtainedin quantitating unspliced RNA by RT-PCR versus Northern blotting couldbe
explained
bythe presenceof smallamounts ofRNA from thegag-pol region
thathad beenspliced
out from the precursor molecule but not yet degraded. This material would notappearasunsplicedRNAin aNorthern blot butwouldincrease the apparent level ofunspliced
RNA in resting cells byRT-PCR analysis.The cognate sequences for primer pair FPS-3 and E17, although separated by more than 5,800 nucleotides in the proviral genome, are brought into close
approximation
on singlyspliced
HIV-1 RNA molecules. An825-bp
fragment
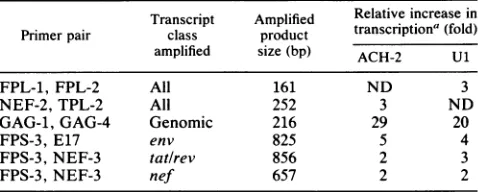
TABLE 1. Induction ofunsplicedviral RNA exceeds that of
splicedRNAby RT-PCR analysis
Transcript Amplified Relative increase in
Primerpair class product transcriptiona(fold)
amplified size(bp) ACH-2 U1
FPL-1, FPL-2 All 161 ND 3
NEF-2,TPL-2 All 252 3 ND
GAG-1, GAG-4 Genomic 216 29 20
FPS-3,E17 env 825 5 4
FPS-3,NEF-3
tat/rev
856 2 3FPS-3,NEF-3 nef 657 2 2
aTheradioactivityin thesignalsfromRT-PCR analyseswasquantitated by
theuseofaradioanalyticalimagingsystem. Thearithmeticaverage of three
separate reactions was determined and thendiminished bythebackground
activity on the blot. All samples represented equal amounts of RNA as
determinedspectrophotometricallyand then confirmedby hybridizationto a
,3-actin probe. Induction of any giventranscript classwas thencalculatedby
dividingthe amountofradioactivityin RNAsamples fromcells after
stimu-lation bythe amount ofradioactivity in RNA samples fromresting cells.
Several different RNAtemplateamounts wereused to ensure that the assay wasin a lineartemplate-to-signal region.ND,Notdetermined.
Ta/rev
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.138.491.77.273.2] [image:4.612.324.563.554.650.2]corresponding
to the env transcript was amplified with these primers. A fivefold change in the steady-state level of this transcript class was observed in ACH-2 cells after PMA induction. These data agree with the results obtained previ-ously from Northern blotting experiments. A fourfold in-crease in the level of singly spliced RNA was observed inUl cells after induction. These data also correlated well with the Northern blotting experiments. Primer pair FPS-3 and NEF-3 coamplify an 856-bp fragment and a 657-bp fragment corresponding to the tatlrev and nef mRNAs, respectively. ACH-2 cells were enriched twofold in both tatlrev and nef mRNAs after induction. Ulcells showed a threefold rise in the level oftatlrevmRNA and a twofold rise in the level of nef transcripts. A similar rise in these doubly spliced species was demonstrated previously by Northern blotting (Fig. 1). The RT-PCR data support the Northern blot data by confirming that accumulation of unspliced HIV-1 RNA, not an increase in total viral RNA, is the primary molecular event associated with viral RNA induction in chronically infected T cells and promonocytes. We further observe that the substantial amount of spliced viral RNA seen in resting T cells and promonocytes by Northern blotting is also con-firmed by RT-PCR analysis. We were able to specifically extend this observation to the nef mRNA by the use of RT-PCR.Induction does not involve a shift in the RNA start site. We performed transcript mapping studies of HIV-1 RNA from ACH-2 and Ul cells with both Si nuclease and primer extension analyses to learn whether induction was associ-ated with a shift in the RNA start site. A 420-nucleotide uniformly labeled RNA probe, corresponding to the nucleo-tide sequence position 300 to 720 of HXB2, was generated with SP6 RNA polymerase and radiolabeled nucleotide triphosphates. Due to the repetitive genomic organization of HIV-1 LTRs, the probe used in these experiments for the detection of RNA transcripts initiating from the 5' LTR cap site also detected RNA transcripts terminating in the 3' LTR. This resulted in both a 265-bp fragment and a 232-bp protected fragment in the Si nuclease protection assays (Fig. 3A). The 265-bp protected fragment represents the distance from the RNA start site at position + 1 within the R region of the 5' LTR to position 720 within the gag gene. The 232-bp protected fragment represents the distance between position 9384 within the 3' LTR and the RNA polyadenylation site. These two fragments were the only ones protected by both Ul and ACH-2 RNA extracted from unstimulated cells. RNA from stimulated Ul and ACH-2 cells predominately protected these same two fragments, although a number of minor species were also seen. These data show that the HIV-1 RNA start site in unstimulated Ul and ACH-2 cells does not shift after induction. These molecules also termi-nated at the same site in the R region of the 3' LTR.
The same RNAs were analyzed by primer extension studies. An end-labeled gag region primer complementary to HXB2 nucleotide sequence position 901 to 930 was hybrid-ized toUland ACH-2 RNA before extension with RT. The expected extension product would be a 475-nucleotide spe-cies representing the distance between position 930 within the gag gene to the RNA start site within the 5' LTR (Fig. 3B). Very little extended material was seen with unstimu-lated Ul RNA, but an intense signal migrating at 475 nucleotides was seen with stimulatedUl RNA. A faint signal migrating at 475 nucleotides could be seen withunstimulated ACH-2 RNA. This signal was much more intense when stimulated ACH-2 was used as template for primer exten-sion. ACH-2 RNA also produced a number ofsmallerprimer
extension products. The strongest signal migrated at 150 nucleotides. This would map a strongprimer extension stop approximately 40 nucleotides downstream ofthe major 5' splicedonor site atnucleotideposition743. Itispossiblethat this strong stop is simply an artifact of the RT reaction. However, the data could alsobe interpretedas
showing
that afraction of the largeintron commontoall identifiedspliced
HIV-1 RNA species is astable species in the ACH-2 RNA pool. Theseprimerextensiondata support the
findings
from S1nuclease protection studies that thepredominant5' RNA start site in stimulatedUl and ACH-2 cells does not repre-sent a shiftfrom the start site in unstimulated cells. These data furthersubstantiate the datafromNorthernblotting
and RT-PCRexperimentsindicatingthatmostofHIV-1 RNA in unstimulated Ul and ACH-2 cells exists in thespliced
form. Induction of unspliced viral RNA parallels viral particle assembly. We were interested indefining
therelationship
between the inductionofHIV-1 RNAclasses and measure-ments of virion assembly. Concomitant viral
RNA,
p24
antigen, and viralparticleanalyseswere
performed
tostudy
the relationship of individual viral RNAspecies
with the induction of HIV-1 virions. The viral RNAdatapresented
previously werecompared withquantitativemeasurementof supernatant viral p24 antigen (Fig. 4). ACH-2 cells showa
20-fold rise in p24 level after 26 h of stimulation. This correlates roughly with the induction of
unspliced
RNA in these cells but isfourtimeshigherthan the rise in total viral RNA (Fig. 1A,Table 1). Theinductionofp24in ACH-2 cells similarlyeclipsed the increase inspliced
RNAs. Incontrast, the level of supernatant p24antigen
in Ul cells roseonly
2.5-fold in 26 h, whereas the induction of
unspliced
viral RNA was 25-fold.Total Ulviral RNAincreasedproportion-ally with p24 antigen at 26 h, but this
relationship
wastransient (Fig. 1B, Table 1). At53 h of
stimulation,
thep24
data suggest a 10-fold induction of viral
replication
when RNA data indicate a 25-fold rise inunspliced
RNA and a3-fold increase intotal RNA.
Although the level of viral p24
antigen
correlated with unspliced viral RNAinduction inACH-2cells,
suggesting
afunctionalrelationshipbetween thisclass ofHIV-1RNA and virion assembly, we were unable to define such a relation-ship forUlcells because of the
unreliability
of thep24
assay as a measure of viralreplication.
Weperformed
electron microscopic analysis onthese cells to obtain amore defini-tive measure of virionreplication.
Cells and supernatant fluids were analyzed forcell-associated virions andcell-free particle counts,respectively,
either before or at different timesafterstimulation.Analysis of cell-associated virions
by
transmission elec-tron microscopy is shown inFig.
5.Figure
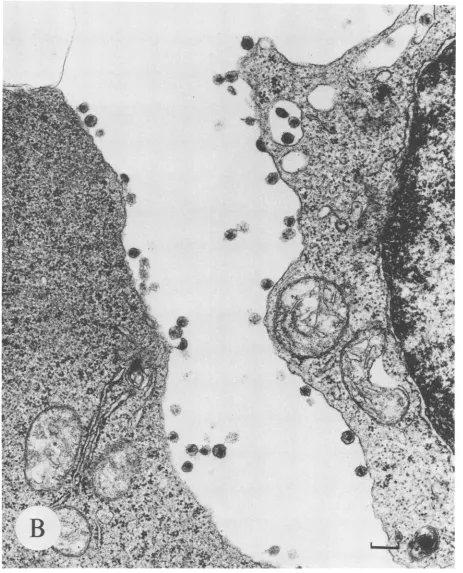
SA is a repre-sentative electron micrograph ofresting
ACH-2 cells. De-spite reportsof the latent natureof theseTcells,
analysis
of over 100 cells revealed active virionproduction
associated with 20%ofthese cells at 0 h. Theseparticles
aremorpho-logically congruent with lentivirus virions with
respect
to size (110 nm) and the presence ofa condensedcylindrical
core (19). Few virions were noted in intercellular
regions.
Most particles were associated with the cell membrane either by spatial
approximation
orby
atetherlike structure. After 21 h ofinduction,
virtually
every cell in 100 cells analyzed wasproducing
virions(Fig. SB).
More of these virions could beseenboth in intercellularregions
and within cytoplasmic vacuoles than inresting
cells,
but themajority
of particles were still associated with thecytoplasmic
mem-brane. The average load of virions per cell was also
in-creasedoverthatin
resting
ACH-2 cells.Figure
5C
showsaon November 10, 2019 by guest
http://jvi.asm.org/
1296 MICHAEL ET AL.
A. S1
NLIclease
Protection
...U.
...A
...-
.....2
PMIA - +
pr-]tFZ'~s
n TA_
5LIP-; -WiA
<:.tE 5 L .~~~~~~..P")I", 489
94
404 364
g
220
190:
,r
B. Primer
Extension)
Ul
PMA-.M
.Ae
..'.'
4-- 3
4+- 1
4+- 2
ACH-2
504
434
*26" U '34
184
T 64 9r61
---A----_T I i 3 tT'R
PcdyA
r4,PNA
L2LLE0~A
~
64'.,
r I.j_.
, ~~...s..
v 232Tp
...
.. ... ... ... ... ... ..<..
FIG. 3. Si nuclease andprimerextensionanalysesofUl and ACH-2 RNA.(A) S1nucleaseanalysis. Total cellular RNA(10p.g)from unstimulated(PMA-) and stimulated (PMA+)Ul and ACH-2cellswashybridized with auniformlylabeled RNAprobe, digestedwithSi
nuclease, electrophoresed through a6% polyacrylamide-7M ureagel,andsubjectedtodirectautoradiography.The arrow labeled"probe" identifiestheposition oftheundigested probe.The arrowslabeled1and2identify theposition oftheSi nuclease-protectedfragmentfrom the5'and 3' LTRs,respectively. Thearrowlabeled3identifies the position of theRNA-RNAhybridnotdigested bytheSi nuclease. The constructionofthe probeand thefragmentgeneratedafterRNA-RNAhybridization andsubsequentSinucleasetreatment areshown inthe diagram below thefigure. Thenumbersabovetheboxes indicate the nucleotidesequenceposition ofHXB2. Thewavylinerepresentsthe 15-nucleotide portion of the probe that is notcomplementary toHXB2. The markers are an end-labeled MspI digest ofpGEM7ZfDNA (Promega). Theexposuretimes of theautoradiographs from theUlandACH-2RNAexperimentswere2.5and24h,respectively. (B) Primer extensionanalysis. Total cellularRNA(10,ug)from unstimulated(PMA-)andstimulated (PMA+)UlandACH-2 cellswashybridizedwith a5'-end-labeledgagregion primer, O-JM76.Theannealedprimerwasextended withRTandnucleotidetriphosphates.Theresulting material
waselectrophoresedthrough a6% polyacrylamide-7M ureagel and subjected todirect autoradiography.Thepositionof O-JM76withrespect totheHXB2sequence andtheexpected lengthofthe extensionproduct are showninthediagrambelow thefigure. Markersare anend-labeled HaeIII digest of pBR322 DNA. The exposure times ofthe autoradiographs from the Ul and ACH-2 experiments were 24 and 48 h, respectively.
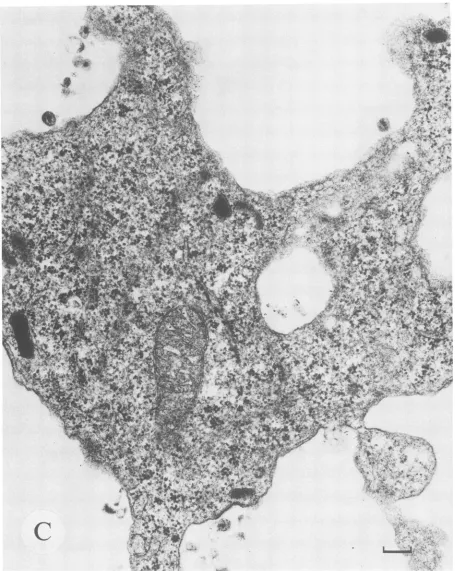
Ul cell before stimulation.Afew virions are associated with thecytoplasmicmembrane. This cell was the only one found to be producing virions after analysis of over 200 unstimu-lated Ul cells.Incontrast,Figure5Dshows a representative Ul cell after21 hof stimulation which is producinggreater numbers of both intracellular and cell surface-associated virions. Tenpercent of 100 Ul cells analyzed were produc-ing virions after induction.
Weconclude that the induction of chronically infected T cells andpromonocytes is associated with an increase in the number of cell-associated virions and that this correlates wellwith the observed increase in the steady-state level of unsplicedHIV-1RNA. No suchrelationship could be drawn between the induction of viral particles and the change in eithertotal viral RNA, spliced viral RNAs, or supernatant p24 antigen. Cell-freeparticle counts showed no consistent relationship with either p24 antigen, viral transcript, or cell-associated virus analysis. This supports the hypothesis
thatcell-associated measures of HIV-1 replication aremore reflective of true viral load than arecell-freemeasures.
DISCUSSION
Molecular mechanism ofHIV-1 RNA induction. We have demonstrated that the stimulation of virion production in chronically infectedTcells andpromonocytesis associated with a 3-to 5-fold increase in the steady-state level of total HIV-1 RNA and a 20- to 25-fold increase in the level of unspliced or genomic RNA. The relative amounts of tran-script classes, determined by radioanalytic imaging in the resting RNA pool of Ul cells, reveal that 3.5% of HIV-1 transcriptsexist in theunsplicedform, 63% of the transcripts are singly spliced, and 33% are doubly spliced. If we normalizetothe minimal copynumber,thenfor every copy ofunsplicedHIV-1 RNAthereare18copies ofsinglyspliced transcripts and 9 copies ofdoubly spliced transcripts. The J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.114.518.78.388.2]60000 1 E 0. CL 40000 20000
0 10 20 30 40 50 60
Time post-induction (hr)
FIG. 4. Supernatant p24 antigen analysis correlates poorly with
viralRNA induction. The level ofsupernatant viral p24 antigen is
shown asafunction of time after initiation of induction with PMA
for ACH-2 and Ul cells. Cell suspensions were cleared by
low-speed centrifugation, and aliquots were frozen for subsequent p24
antigendetermination by antigencapture.ACH-2supernatantswere
diluted1:100 and Ul supernatants werediluted 1:300 toquantitate
the resultant p24 values in the linearrangeoftheassay.
number of unspliced transcripts increases to 25 copies, whereas the copy number of singly and doubly spliced
species following induction increases to 36 copies and 24
copies, respectively. If little or no turnover ofpreexisting transcripts is assumed,then thefraction ofRNAmolecules that remain unspliced rises from4%in the restingRNApool
to approximately 40% in the stimulated RNA pool. This 10-fold difference in stability of theprimaryHIV-1 transcript
would explain why the 3-fold rise in total HIV-1 RNA in stimulated Ul cells is accompanied by a nearly 30-fold
increase in unspliced RNA. Similar analysis of ACH-2
HIV-1 RNA pools is consistent with a shift in the steady-state level of unspliced RNA from 6.6% to 30%. A 5-fold increase in the fraction of unspliced RNA in ACH-2 cells
could account for the 20-fold increase in unspliced
tran-scripts coupled with onlya5-fold increase in total viral RNA
if increasedturnoverof preexisting transcriptsdid notoccur
in these cells.
The data presented do notexclude the possibility that the
rateof denovoRNAsynthesis has also risen inconcertwith the overall increase in unspliced RNA. This would be
possible, however, onlyif therate of degradation ofspliced
RNAs,butnotgenomic RNA, alsoincreasedconcomitantly.
Although it is theoretically possible, we consider this
sce-nario less likely. The mechanism of increased stability of
unspliced viral RNA could be either directly dependenton or
independent of the nuclear splicing machinery. The data
presentedareconsistentwithamechanismin whichgenomic
RNA moleculesareshielded from spliceosomes inanactive sense and the rate of nuclear-to-cytoplasmic efflux remains unchangedfrom that of resting cells. Conversely, the data
arealso consistent with amechanismthat is independentof
a direct effect on the splicing machinery where induction
could be associated with an increased efflux of unspliced
viral RNAout of the nucleus intothe cytoplasm. Thereare
data to support both mechanisms from previous work in
transiently infected cells. These reports strongly implicate
the rev gene product as the viral factor that mediates the
differential expression of unspliced viral RNA in the late
infection period (5, 6, 12, 15, 32). Althoughourdataindicate
only a threefold rise in the transcript encoding rev, it is
possible that the level or activity of the Rev protein exerts its effect on RNA stability out of proportion to its transcript level. We are studying the induction of cells chronically infected with HIV-1 at the translational level to address this question.
The Si nuclease and primer extension analyses of viral RNAs confirm that transcriptionalupregulation of HIV-1 is not associated with changes in the majority ofthe 5' or 3' ends of viral mRNAs. There is no evidence, therefore, for the existence of stable upstream viral RNA sequences that might influence the subsequent processingofnewly synthe-sized transcripts. These data further suggest that the same transcriptional start sites that support basal-level viral expression in chronically infected cells also support the elevated level of viral RNA expression asthese cells become activated. Although the specific molecular mechanism that mediates the induction of HIV-1 RNA inchronically infected T cells and promonocytes remains unclear, this mechanism is associated primarily with a shift in the pattern of HIV-1 RNA splicing, favoring the stability ofgenomic RNA. This mechanism significantly enriches the steady-statefraction of unspliced HIV-1 RNA in the cellularRNA pool ofstimulated cells. If this mechanism does directly involve the splicing machinery of the host cell and notstrictly the rate ofnuclear RNA efflux, then it would represent avariation of alternative splicing. Alternative splicing as amechanism ofdifferential gene expression has been demonstrated in biologic systems as diverse as adenovirus, simian virus 40, Drosophila mela-nogaster, and rats (4, 30). Coding information is extended beyond the strictly linear information contained in a tran-scription unit by allowing fordifferent coding sequences of that unit to be spliced together to formfunctionally distinct populations of mRNA molecules. The Drosophila suppres-sor-of-white-apricot and transformer genes, among
others,
are especially relevant to the observations in this report. Alternative splicing of the precursor RNA
molecuie
at these loci results in an mRNA that is either fully enabled orfully
disabled for subsequenttranslation of thefunctional protein encoded by the locus (3, 8). InHIV-1-infected cells,
splicing
of a precursor viral RNAmolecule would preventthespliced product from packaging into a functional virion. Unlike other models ofalternative splicing, both the precursor and the product molecule are functional in HIV-1 gene expres-sion. The commitment of agenomic RNAmolecule to
being
spliced, however, removes that molecule from the
pool
of viral RNA available for virion assembly. Thus, the cellular commitment to splice HIV-1 RNA impingesdirectly
on virion biogenesis. Since only unspliced HIV-1 RNA can support the translation of the gag-pol loci, the decision to splice these molecules may also indirectly block virion assembly by reducing the pool of availablegag-pol
gene products that are integral components ofcompleted virions. The molecularcommitment to splice, as wellastheselection of specific splicedonor-acceptorpairs,serves as aregulatory
mechanism of viral geneexpression forHIV-1.
Comparison
of the relative abundance of multiply spliced
transcripts
found in HIV-1-infected T cells and monocytes
speaks
to this mechanism ofregulation directly (36). Recent work onthe pre-mRNA splicing factor SF2 from HeLa cells has identified it asnecessary for thestabilization or
assembly
of the earliest specific prespliceosome complex(28).
This fac-tor is particularly attractive as a candidate host-encoded protein that may beimportant in thedifferentialcommitment of HIV-1 RNAs tobecoming splicedbyinteractingwith viral regulatory elements such as Rev.Experiments in acutely infected cells have elucidated a
ACH-2
-D- Ul
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.56.294.76.224.2]1298 MICHAEL ET AL.
t5
X,
6>K.
s* < J >r' Al1 Jlb
{zX~~4
<"-.
iAl~ ~ ~ 4
.4vV"~ ~
* A,
$%d
A
SIt *k3¢}
.;4,, c§;
Z"1C
_,,4wz&. ,t /
$4*
X
4''eN
;
;'
''S
,
',*..
"
"-45
A
r*>w~4
wS,*
\s*¢
9*.t$b
# * i ?*or4
-,.
FIG. 5. Induction of virionproduction bytransmission electronmicroscopycorrelateswith
unspliced
HIV-1RNA.Photomicrographs
areshown of thin sections fromglutaraldehyde-fixedand osmiumtetroxide-stained cellpellets examined
by
transmission electronmicroscopy.
(A)AnACH-2 cellbefore stimulation. Magnification,ca. x30,240. Bar,0.5 p.m.(B)TwoACH-2cells after 21 hof stimulation withPMA. Magnification,ca. x42,840.Bar,0.2
pum.
(C)A Ul cell before stimulation.Magnification,
ca.x50,400.
Bar,0.2pum.
(D)TwoUl cells after21 hofPMAstimulation. Magnification,ca. x50,400. Bar,0.2 p.m.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.91.543.105.668.2]1
X, N
I.
af,I/
FIG. S-Continued.
temporal pattern of HIV-1 RNAexpression that is divided intoearlyand latephases.Theearly phaseis associated with
an accumulation of small, doubly spliced transcripts. The
late phase is associated with an increase in the fraction of
singly spliced
species
andunspliced
viral RNA. This pattern isconsistentwithaswitch from theproduction
ofregulatory
geneproductstostructural gene
products
andgenomic
RNA as initial infectionultimately
subverts the host to theon November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.82.545.77.650.2]J.VIROL. 1300 MICHAEL ET AL.
C 4k'<go ";',.": '\
: , .I-, . ' .
AgF$
FIG. 5-Continued.
ductionofprogenyviralparticles. We show that induction is
associated with a sharp increase in the production of viral
particles in chronically infected cells. We have demonstrated thatinduction of virions in these cells isaccompanied bya
shift in viral RNAexpression thatisqualitatively similarto that seen incellsacutelyinfected with HIV-1. In thisregard, these data are consistent with those recently published by others(35).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.83.538.76.647.2]VOL.65, 1991 INDUCTION OF HIV-1 IN CHRONICALLY INFECTED CELLS 1301
'*.!;-6% /' _ -;~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~0.Xtt , v ;.
X,
/'
.'.
**S
,
i
* !XX
t'#. v
'A
Bs
_
;'
ip ; > tZ jt X; ,r ;;, ...
WS*t;e, , [, ^ . . . t ,^ t >*
.1,
J,'<
e!
~~~~~~A
'.,,'jA''t>,
4t
t
Ri
+
ai S
X
N~~~~~~~~~~~~Fs
: D . . XAPF . t ;. ;,, !'j,,.,Yv
44t t ;<--N,M,Li <-';..i
viralRN.W~ ~ ~eesrc yteosrainta oh
teersigTclsadpoooye
sabotoefutretn
C-
n
1clscnansbtata
mut
f
o
hti ul
tmlae
el.Gvnta hs
xeiet
HIV-1 RNA.
Although
only
approximately
5% of these wereperformed
onlogarithmically growing
cells, itislikely
on November 10, 2019 by guest
http://jvi.asm.org/
1302 MICHAEL ET AL.
thatbasal-level
transcription
isrelatively
brisk but that the half-life ofunspliced
viral RNA in the nucleus is brief. In contrast, stimulated cellsproduced only
three tofive times theamountofHIV-1 RNAcompared
withresting cells,
butthey produced
far more viralparticles.
Nuclearregulatory
factors suchasNF-KB(11, 21, 26, 31,
33, 34, 38, 40)
andSP1(18, 24)
have beenimplicated
asimportant
inmediating
the effect ofinducersonincreasedviraltranscription
from the 5' LTR.Binding
sites for thisprotein
have been shownin the upstream controlregions
of the HIV-1 genome(33).
A mechanismthatonly
increased thesteady-state
level oftotal HIV-1RNA,
however,
would notexplain
the observations made in theexperiments
described in this report. A con-certedmechanismthatboth increasestranscription
fromthe 5'LTR andgreatly
decreasesthe percentageofnascentviral RNA molecules that arespliced
would berequired
to pro-duce themajor
shift in RNAsplicing
pattern observed in stimulated ACH-2 and Ul cells. Thevirally
encoded Revprotein
isprobably
afactor thatis involved in this upregu-lationofunspliced
RNAand virionassembly.
Previous reports have made reference to
resting
ACH-2 and Ul cells asvirologically quiescent (9, 17).
The datapresented
in this paper show that substantial amounts of HIV-1RNAaremadeby
these cellsin theresting
state.We further show that unstimulated ACH-2 cellsproduce large
amounts of
morphologically
intact viralparticles. Although
we havenot demonstratedthat these virions arealso
func-tionally intact,
it is a reasonableassumption
thatthey
arecapable
ofinfecting appropriate
targetcellstobegin
another round of the viral lifecycle.
Resting
Ul cellsproduce
apaucity
ofvirions,
incomparison
tothe situation in ACH-2 cells. Theamountof totalHIV-1-specific
RNAinresting
Ul cellsby radioanalytic imaging
of Northern blots isapproxi-mately
two-thirdsthat ofresting
ACH-2cells. The percent-ageofunspliced
RNAinthe cellularRNApool
ofresting
Ul cellsisroughly
half thatofresting
ACH-2 cells.Itisunlikely,
therefore,
that the differentialproduction
of virionsby
resting
Ul and ACH-2 cells canbeexplained solely by
the relativestoichiometry
ofbasal RNAtranscript
levels. These twocelllinesseemtorepresenttwovery distinctfunctional states of viralexpression.
The block in afully productive
infective stateinresting
Ul cells seems toberegulated
notonly
at thelevel ofgenomic
RNAstability
but also at the level ofvirionassembly. Resting
ACH-2 cellsmay bemoreleaky
thanUl cellsatthis level of viralexpression.
Basedon the observed levels of viral RNA and virionproduction
in thesecellsbeforeinduction,
wefeelthat thevirologic
status ofthese cells is mostaccurately
described as alow-level-expressing
state.HIV-1 RNA transcriptclasses define viral burden. These results have shown thatthe change in virionproduction in
chronically
infected T cells and promonocytes associated with inductionisnotreflectedbyeither the measurement of supernatant viralp24antigen
orthelevelof total viral RNA. The level ofspliced
HIV-1 RNAs was similarly crude as a benchmark of viral replication. Measurement ofunspliced viral RNAprovideda muchmorefaithful reflection of viral burden in these cells as they passed from a relatively low-levelstateof viralexpressionto afully productive state. Theimplications
of these results extend the rationale foremploying
RNAexpression
as avirologicmeasure in HIV-1. Unlike DNAanalysis,
viral RNA analysis provides a mea-sureoftheactively
expressingcompartment of HIV-1 in the total viral load. The resultspresentedin this paper refine the measurementofviral loadby focusing
effortson theanalysis offunctional classes of HIV-1 viral RNA in infectedpa-tients. The possible relationship of specific HIV-1 mRNA classes, notsimply total viral RNAlevel, may have
impli-cations for clinical diseaseprogressionandprognostication. Moreover, the virologic response to clinical interventions such aschemotherapy, receptorblockade, and
passive
and active immunotherapy may be reflectedby changingHIV-1 RNA transcript patterns. These transcript patterns may augment established measures of virologic responses to therapy. Wearecurrentlyinvestigatingthesepossibilities inpatient populations
from multicenter intervention trials. ACKNOWLEDGMENTSWe thank Victoria Polonis and Maria Wood for assistance with cell cultureand Phil Ray, FrancineMcCutcheon, and Eric
Sanders-Buell for thesynthesis of the oligonucleotides used in thisproject.
David Ritchey performed the Si nuclease and primer extension studies. Arnold Fowler helped with the p24 antigen analyses, and Jan Endlich assisted with electron microscopic analysis. Victoria Hunterprovided graphic designassistance.
REFERENCES
1. Arrigo, S.A., S. Weitsman,J.D.Rosenblatt,andI.S. Y. Chen.
1989.Analysis ofrevgenefunctiononhumanimmunodeficiency
virus type 1 replication inlymphoid cellsby usingaquantitative
polymerase chain reaction method. J. Virol. 63:4875-4881. 2. Arya, S. K., C. Guo, S. F. Josephs, and F.Wong-Staal. 1985.
Trans-activator gene of human T-lymphotropic virus type III (HTLV-III). Science 229:69-73.
3. Boggs, R. T., P. Gregor, S. Idriss, J. M. Belote, and M. McKeown. 1987. Regulation of sexual differentiation in D. melanogaster via alternative splicing of RNA from the trans-formergene. Cell 50:739-747.
4. Breitbart, R. E., A. Andreadis, and B. Nadal-Ginard. 1987. Alternativesplicing: a ubiquitous mechanism for thegeneration
of multiple protein isoforms from single genes. Annu. Rev. Biochem. 56:467-495.
5. Chang,D. D., and P. A.Sharp. 1989. Regulation by HIV rev
dependsuponrecognitionofsplice sites. Cell59:789-795. 6. Chang, D. D., and P. A. Sharp. 1990. Messenger RNA transport
and HIV revregulation. Science 249:614-615.
7. Chirgwin, J. M., A. E. Przybyla, R. J. McDonald, and W. J. Rutter. 1979. Isolation ofbiologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
8. Chou, T. B., Z. Zachar, and P. M. Bingham. 1987. Develop-mental expression of aregulatory gene is programmed at the level ofsplicing.EMBO J. 6:4095-4104.
9. Clouse,K.A.,D.Powell,I.Washington,G.Poli, K. Strebel, W. Farrar, P. Barstad, J. Kovacs, A. S. Fauci, and T. M. Folks. 1989. Monokineregulation of human immunodeficiencyvirus-1
expression in a chronically infected human T-cell clone. J.
Immunol. 142:431-438.
10. Cullen, B. R., and W. C. Green. 1989. Regulatory pathways
governingHIV-1replication.Cell 58:423-426.
11. Duh, E. J., W. J. Maury, T. M. Folks, A. S. Fauci, and A. B. Rabson. 1989. Tumor necrosis factor alpha activates human
immunodeficiency virus type 1 through induction of nuclear factorbindingto theNF-KB sitesin the long terminal repeat. Proc. Natl. Acad.Sci. USA86:5974-5978.
12. Emerman, M., R. Vazeux, and K. Peden. 1989. The rev gene
product ofthehuman immunodeficiencyvirus affects
envelope-specificRNAlocalization. Cell57:1155-1165.
13. Ensoli, B., P. Lusso, F. Schacter, S. F. Josephs, J. Rappaport, F. Negro, R. C. Gallo, and F. Wong-Staal. 1989. Human herpes
virus-6 increases HIV-1 expression in co-infected T cells via nuclear factors binding to the HIV-1 enhancer. EMBO J. 8:3019-3027.
14. Feinberg, M. B., R. F. Jarrett, A.Aldovini,R. C. Gallo, and F.
Wong-Staal.1986.HTLV-IIIexpression and production involve
complex regulation atthe levelsof splicing and translation of viral RNA. Cell46:807-817.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
15. Felber, B. K., M. Hadzopoulou-Cladaras, C. Cladaras, T. Cope-land, and G. N.Pavlakis. 1989. Rev protein of human
immuno-deficiency virustype 1 affectsthe stability and transport of the viral mRNA. Proc. Natl.Acad. Sci. USA 86:1495-1499. 16. Folks, T. M., K. A. Clouse, J. Justement, A. Rabson, E. Duh,
J. H.Kehrl,and A. S.Fauci. 1989. Tumor necrosis factor alpha
induces expression of human immunodeficiency virus in a chronically infected T-cell clone. Proc. Natl. Acad. Sci. USA 86:2365-2368.
17. Folks, T. M., J. Justement, A. Kinter, S.Schnittman, J. Oren-stein, G. Poli, and A. S. Fauci. 1988. Characterization of a promonocyte clone chronically infected with HIV and inducible
by13-phorbol-12-myristate acetate. J. Immunol. 140:1117-1122. 18. Garcia, J. A., F. K. Wu, R. Mitsuyasu, and R. B. Gaynor. 1987.
Interactions of cellular proteins involved in the transcriptional
regulation of the human immunodeficiency virus. EMBO J. 6:3761-3770.
19. Gonda, M. A., F. Wong-Staal, R. C. Gallo, J. E. Clements, 0. Narayan, and R. V. Gilden. 1985. Sequence homology and
morphologic similarity ofHTLV-IIL andvisna virus, a
patho-logic lentivirus. Science227:173-177.
20. Green, W. C. 1990.Regulationof HIV-1 geneexpression.Annu. Rev. Immunol. 8:453-475.
21. Griffin, G. E., K. Leung, T. M. Folks, S. Kunkel, and G. J. Nabel. 1989. Activation ofHIV geneexpression during mono-cyte differentiation by induction ofNF-KB. Nature (London) 339:70-73.
22. Hammarskjold, M.-L., J. Heimer, B. Hammarskjold, I. Sang-wan, L. Albert, and D. Rekosh. 1989. Regulation of human
immunodeficiency virusenvexpression bythe revgeneproduct.
J. Virol. 63:1959-1966.
23. Hauber, J., A. Perkins, E. P. Heimer, and B. R.Cullen. 1987.
Trans-activation ofhumanimmunodeficiency virusgene
expres-sion is mediated by nuclearevents.Proc.Natl.Acad.Sci. USA 84:6364-6368.
24. Jones, K. A., J. T. Kadonaga, P. A. Luciw, and R. Tjian. 1986.
Activation of the AIDS retrovirus promoter by the cellular transcriptionfactor, SP1. Science232:755-759.
25. Kao, S.-Y., A. Calman, P. A. Luciw, and B. M. Peterlin. 1987.
Antitermination of transcription withinthelong terminalrepeat
ofHIV-1bytatgeneproduct.Nature(London)330:489-493. 26. Kaufman, J. D., G. Valandra, G. Roderiquez, G. Busher, C.
Giri, and M. A. Norcross. 1987. Phorbolesterenhances human
immunodeficiency virus-promotedgeneexpression andacts on arepeated10-base-pair functional enhancerelement.Mol. Cell. Biol. 7:3759-3766.
27. Kim, S., R. Byrn, J. Groopman, and D. Baltimore. 1989.
Temporal aspects ofDNA andRNA synthesis during human
immunodeficiency virus infection: evidence for differentialgene
expression. J. Virol. 63:3708-3713.
28. Krainer, A. R., G. C. Conway, and D. Kozak. 1990. Purification
and characterization of pre-mRNA splicing factor SF2 from HeLacells. GenesDev. 4:1158-1171.
29. Laspia, M. F., A. P. Rice, and M. B. Mathews. 1989. HIV-1 tat
protein increasestranscriptional initiation and stabilizes
elonga-tion. Cell59:283-292.
30. Leff, S. E., M. G. Rosenfield, and R. M. Evans. 1986.Complex transcriptional units:diversity in gene expression byalternative
RNAsplicing. Annu. Rev. Biochem. 55:1091-1117.
31. Lenardo, M. J., and D. Baltimore. 1989. NF-KB: apleiotropic mediator of inducible and tissue specific gene control. Cell
58:227-229.
32. Malim, M. H., J. Hauber,Shu-Yun Le, J. V. Maizel, and B. R.
Culien. 1989. The HIV-1 rev trans-activator acts through a structured target sequence to activate nuclear export of
un-splicedviral RNA. Nature (London) 338:254-257.
33. Nabel, G., and D. Baltimore. 1987. An inducibletranscription factor activates expression ofhumanimmunodeficiencyvirus in Tcells. Nature (London) 325:711-713.
34. Pomerantz, R. J., M. B. Feinberg, D. Trono, and D. Baltimore. 1990. Lipopolysaccharide is a potent monocyte/macrophage
stimulatorof humanimmunodeficiencyvirus type 1expression.
J. Exp. Med. 172:253-261.
35. Pomerantz, R.J., D. Trono, M. B. Feinberg, and D. Baltimore. 1990. Cells nonproductively infected with HIV-1 exhibit an
aberrantpatternof viral RNAexpression:amolecular modelfor latency.Cell61:1271-1276.
36. Robert-Guroff,M.,M.Popovic, S. Gartner, P. Markham, R. C. Gallo, and M. S. Reitz. 1990.Structure andexpressionof tat-, rev-, and nef-specific transcripts ofhuman immunodeficiency virustype 1 ininfectedlymphocytes and macrophages.J.Virol. 64:3391-3398.
37. Selby,M.J.,E.S.Bain, P. A.Luciw,and B. M.Peterlin. 1989. Structure, sequence, andposition of the stem-loopintar deter-minetranscriptional elongation by tatthrough the HIV-1long terminalrepeat.GenesDev.3:547-558.
38. Siekevitz,M., S. F.Josephs, M. Dukovich,N.Peffer,F.
Wong-Staal, and W. C. Greene. 1988.Activation ofthe HIV-1 LTRby T cell mitogens and the trans-activator protein of HTLV-1.
Science 238:1575-1578.
39. Sodroski, J., R. Patarca, C. Rosen, F. Wong-Staal, and W. Haseltine. 1985. Locationof the trans-activating regiononthe genomeofhuman T-celllymphotropic virus type III. Science 229:74-77.
40. Tong-Starksen, S.,P. A. Luciw,and B. M. Peterlin. 1987. The HIV LTR responds to T-cell activation signals. Proc. Natl.
Acad. Sci.USA 84:6845-6849.
41. Zack,J. A., A. J. Cann, J. P. Lugo,and I. S. Y. Chen. 1988.
HIV-1 production from infected blood T cells after HTLV-I inducedmitogenic stimulation. Science 240:1026-1029.
on November 10, 2019 by guest
http://jvi.asm.org/
ERRATA
Expression
of
Collagenlike Sequences by a Tumor Virus,
Herpesvirus Saimiri
PETERGECK,SCOTT A. WHITAKER, MARIA M. MEDVECZKY, AND PETER G.MEDVECZKY
Departmentsof Pharmacologyand Molecular Genetics and Microbiology, University of Massachusetts MedicalSchool, Worcester, Massachusetts 01655
Volume 64, no. 7, p. 3512 and 3513: The nucleotide sequence in Fig. 5A, between nucleotide positions 1101 and 976, and the
corresponding
amino acidsequence inFig.
6,
between aminoacidpositions
64 and105,
should readas shownbelow:CCC CCA GGA CCT CCA GGA CCT TCA GGA CTG CCA GGA TTG TTT GTA ACA AAC TTA
P P G P P G P S G L P G L F V T N L
TTG CTT GGA ATC ATA ATT TTA CTC TTA TTA ATT ATA GTT GCG ATC TTA CTG GTG
L L G I I I L L L L I I V A I L L V
TCT AAA TTA GTA GTA AAC TAA
S K L V V N
Induction of
Human
Immunodeficiency
Virus Type 1
Expression
in
Chronically
Infected Cells
Is
Associated
Primarily
with
a
Shift in RNA
Splicing Patterns
NELSONL. MICHAEL, PAULMORROW,JOSEPH MOSCA, MARYANNEVAHEY, DONALDS.BURKE,ANDROBERT R. REDFIELD
Departmentof RetroviralResearch and DivisionofRetrovirology, Walter Reed Army Instituteof Research,
13TaftCourtSuite 200, Rockville, Maryland20850,andSRA Technologies and
Henry M.JacksonFoundation, Rockville, Maryland20850
Volume 65, no. 3, p. 1292, "Oligodeoxynucleotide synthesis and sequences," line 9: Insert NEF-3 as shown: NEF-3, 5' TTGCTACTTGTGATTGCTCCATGTTTTTC3' (8529through 8501);
Lines 11and 12, FPS-3 should read:
FPS-3,5' CGCACGGCAAGAGGCGAGGG 3' (260 through 279);