Vol.52, No.3 JOURNAL OFVIROLOGY, Dec. 1984,p.966-972
0022-538X/84/120966-07$02.00/0
Copyright C 1984, American SocietyforMicrobiology
An
Accessory Role for
Escherichia coli Integration Host Factor:
Characterization of
a
Lambda
Mutant
Dependent Upon
Integration
Host Factor
for DNA
Packaging
SUSAN E.
BEAR,'
DONALD L.COURT,'*
ANDDAVID I. FRIEDMAN2Laboratory of MolecularOncology, National Cancer Institute, Frederick Cancer Research Facility, Frederick, Maryland
21701,'
andDepartment of Microbiology and Immunology, University of Michigan School of Medicine,AnnArbor,Michigan 481092
Received4June1984/Accepted 18 August 1984
Bacteriophage lambda grows lytically on Escherichia coli defective forintegration host factor, a protein involved in Asite-specific
recompbination
and theregulationof geneexpression. Wereportthe characterizationof amutant, A coslS4, that, unlike wild-type X, is defective for growth in integration hostfactor-defectiveE. coli. The cis-dominant mutation in A cosl54 is a single base pair change in a region of hyphenated dyad symmetry close to the k left cohesive end; this mutation prevents DNA packaging. We propose the following twoalternative roles for this site in K DNA packaging: (i) to bind an E. coli accessory protein required in the absence ofintegration host factor or (ii) to bind the phage-encoded terminase protein that is essential for DNA packaging.
The bacteriophage lambda is encapsidated as a linear
48.5-kilobase duplex DNA molecule by cleavage ofa long
concatemerformedlate ininfection byrolling circle replica-tion(Fig. 1)(reviewedinreferences6and14).This cleavage
occurs at a unique site called cos and requires the phage terminase protein encoded bythegenes NulandA(16).The terminase protein introduces two nicks in the DNA sepa-rated by 12 base pairs (bp) and generates 5' single-strand extensions known as the A cohesive ends (2, 42). On infection of Escherichia coli by X, the linear DNA is
in-jected, the cohesiveendspair, and thecircularized DNA is ligated to form a covalently closed molecule to serve as a
substrate forreplication (39).
Thescanning
model
ofDNApackaging (8)suggeststhat,after the initial cleavage at cos, the terminase protein
re-mains boundtotheprohead as theX DNA isencapsidated.
When terminase encounters a second cos on the DNA
moving into the prohead, itcleaves the DNAagain (Fig. 1). Recent studies have defined the substrate surrounding cos
thatis requiredfor in vivo cleavage and packaging.A160-bp region extending from about bp -40from the left ofthe cos
junctiontobp +120 to therightappears both necessary and
sufficient for efficient encapsidation (9, 19, 33).
Both the cos cleavage reaction and the encapsidation of phage DNAcan beaccomplished in vitro (1,21, 40). These
reactionsrequire phage DNA, terminaseprotein, ATP, and extractsof E. coli
(2).
Thissuggests theparticipation
of hostprotein(s) in the reaction. A22-kilodalton (kd)protein
puri-fiedfromuninfectedcellular extracts promotes coscleavage
in vitro (16). Recent findings indicate that purified integra-tion host factor (IHF) also supports this in vitro reaction (16).
IHFbinds to DNA andis isolated in activeformas a 1:1 multimer of theproducts ofthe E. coli himA andhip(himD)
genes(32, 36;E.Flamm, personal
communication).
IHFwasidentified as an accessory factor in the site-specific
recom-bination reactions for integration and excision of the X genome (23). These recombination reactions, which occur
* Correspondingauthor.
between unique sites on theKand E. coliDNA, require the phage-encoded Int protein (3, 35). Although IHF has no DNAcleavageactivity,itbindstospecificsequences on the A DNA near the sites of recombination and enhances the
binding of the Int protein (36). In another capacity, IHF controlstheexpression of avariety of genes (11, 22, 26, 30). Does. IHF participate in the K DNA packaging reaction? Although it has been shown that IHF promotes cos cleavage in vitro (14), in vivo studies demonstrate that A can grow lytically on either himA or hip mutants (27). This indicates
that,in vivo,Adoesnotutilize IHF, oralternatively,that K uses either IHF or another protein(s) in the encapsidation
process. Ifthe latter is true, the 22-kd protein identifiedin vitro might be the alternative to IHF in the cos cleavage
reaction in vivo. A role for IHF in packaging is clearly
indicated bystudies with the lambdoidphage 21. UnlikeA, this phage does notgrow lytically on himA or hip mutants and requires IHF for packaging in vitro (M. Feiss, S.
Frackman, and J. Sippy, personal communication). In this
work, we characterize a mutant of A, A coslS4, whose properties suggest that IHFparticipates inDNApackaging
in vivo.
MATERIALS ANDMETHODS
Bacterial and phage strains and media. The bacterial and
phage strains usedare shown inTable 1. Nodifference was observed in the efficiency ofplating (EOP) ofany phage
between C600 and N99. Thestrainused in the redplaquetest (5) to assay for IntactivitywasRW842. Singleandmultiple A lysogens (X coslS4 cI857 or Xcos+ c1857) in N99 were constructed asdescribed previously (13, 18). TheAhimA82
mutation was introducedinto these strainsby P1
transduc-tion (24).
The A coslS4 int6 c1857 phage was obtained from the
National Institutes of Health phagecollection. It had been treatedpreviouslywithnitrosoguanidinetogenerate the int6
mutation (17). The strains used for
phage
constructions(DC7, SA297, andN4830) arerepresented in
Fig.
2. TheK coslS4int'
c1857phage was madebyinfecting
SA297 with KcoslS4 int6cI857 andselectingforint+ recombinant phage 966on November 10, 2019 by guest
http://jvi.asm.org/
Head
A
Tail
5' NulA a
3' LeftCohesive End
Empty Phage Head
B 5' SR i lNu1A attP SR NulA
DNAReplication SiteSpecific
Recombination Immunity Lysis
m - r m n I I
3ttPintxis NclcrocllOP SR3'
5'
RightCohesive End
ttP SR Nu1A
3, c
cos
C
NuIAcos
_ 5'
FIG. 1. A, The 48.5-kilobase linear A DNA molecule. The cohesiveendsare a12-bp5' extension. Thegenomecanbe divided intoareas
involved in head and tailprotein synthesis,site-specific recombination (integrationandexcision), immunity, DNAreplication,and celllysis. Selected genetic markers, including the leftmost genes Nul and A and the rightmost genes Sand R, are indicated. B, The A genomic
concatemerproduced by rolling circle replication of the circularized DNA. Topackageaunitlength of DNA, the phage-encoded terminase
protein bindstothe DNAatcosandtotheemptyphage head. Single-strandnicks separated by 12 bpareintroducedinto the DNAatcos
by terminase (vertical arrows). C, The left end of the DNA(indicated by thegenesNul andA)entersthephage head first.Terminasecleaves the DNA againuponencounteringasecondcossite. The cohesive endsarethus generated, andaunitlength ofX DNAis encapsidated.
bythe redplaque testwith RW842.This kcoslS4 int+cI857
phage exhibited thesamephenotype inIHF-defective (IHF-) E. coliasitsint6parent.ThekcoslS4 int6 imm21phagewas madeby infectingDC7 withAcosl54int6 c1857 andselecting for imm21 by platingon N4830.
The mediumwasLuriabrothcontaining1% Difcotryptone broth, 0.5% NaCl, and 0.5% yeast extract. Tryptone broth
agar plates contained 1% Difco tryptone, 0.5% NaCl, and 1%agar.Phagewerediluted in 10 mM Tris(pH 7.4), 10 mM MgSO4, and 0.01% gelatin. Tryptone broth top agar con-tained 0.7%agar. Whennecessary, ampicillin wasadded to
a final concentration of 50 ,ug/ml. Indicator agar plates for the red plaque test were tryptone broth with 1% galactose and2,3,5-triphenyl-tetrazoliumchlorideasdyeindicator(5). Bacteriophagecrosses.C600 cellsweregrownto2x 108/ml in Luria broth supplemented with 0.2% maltose. The cells were centrifuged and concentrated 5x in 10 mM MgSO4. Cells (5 x
107)
were infectedwithphage ata multiplicityof 3. After 10min ofadsorptionat roomtemperature, themix ofphageandcells(50 ,ul)wasexposedtoUV irradiation(300ergs per mm2). Luria broth (10 ml) with 5 mM CaCI2 was added, and the infections were incubated with shaking at 38°Cfor 90min.After the addition ofchloroform, thelysate
was vortexed, and the debris was removed by centrifuga-tion.
Marker rescue experiments. C600 was transformed to ampicillin resistance with either pBR322 vector DNA or
pHC79cosmid DNA, andN4830 was transformedto ampi-cillin resistance withpKC30int (D. Court, unpublished data) bystandardprocedures (38).Note that N4830 doesnotcarry
thewild-typeallele of int6. ThepKC30int plasmidcarries the entire int gene. The phage infections were carried out as describedabove, with the exception thatthemultiplicity of phage infectionwas0.5. The supernatantwas removed toa fresh tube, diluted, and used to score forplating efficiency on K1942 (AhimA82 gyrB230). To screenfor int+
recombi-nant phage, the diluted supernatant was used to infect RW842.
DNA sequencing studies. Phage DNA was treated with bacterial alkaline phosphatase to remove the 5'-phosphate (BAP- MATE; Bethesda ResearchLaboratories, Inc., Gai-thersburg, Md.). These endswere then labeled with 32Pby using [-y-32P]ATP and polynucleotide kinase. Alternately,
the 3' endsofthephageDNAwerelabeled with[a,-32P]dATP
by using the Klenowfragmentof E. coli DNA polymerase. An HpaIrestriction fragment containing thefirst 734bp of
the A left-end terminus was isolated and purified. The
modification and cleavage reactions of Maxam and Gilbert wereused (25), and the resulting preparations ofDNA were
[image:2.612.138.470.78.223.2]electrophoresedin8%and6%polyacrylamide gels.Thegels
TABLE 1. Bacterial and phage strains
Strain Description Sourceor reference
Bacteria
C600 supE tonA thr leu thi NIH" strain collection N99 suporpsLgaIK2 NIH strain collection
K634 N99 himA42 (28)
E268 N99 hip-157 E. Flamm
JD12 N99 himA42hip-157 K. Abremski K1299 N99 AhimA82 H. Miller (11)
E444 N99Ahipl E. Flamm
K1870 N99gyrB230 (12)
K807 N99gyrBI14 (29)
K1942 N99 AhimA82 (11)
gyrB230
RW842 HfrH (5)
gaiT: :(XA[int-FII])
K159 N99(Ximm434) D. Friedman
K124 N99(Xc+) D.Friedman
Phage
AcoslS4int6 NIH phage collection
c1857 (18)
XcoslS4int+ This work
c1857
XcoslS4int6 This work
imm21
Xint6 red3 NIH phage collection
imM21C
Kimm21 c NIH phage collection
Xc1857 NIH phage collection
Aimm434 NIHphage collection
XAamllc1857 NIH phage collection
XLam63c1857 NIH phage collection
Xc1857Pam3 NIH phage collection
Xc1857RamS NIH phage collection
aNIH, National Institutes ofHealth.
attP SR Nul A
:=t-I
i
g _qon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.314.556.401.717.2]968 BEAR, COURT, AND FRIEDMAN
Gal BOP'
-ntxi
cos
r-i
cill N cl cro cil OP 0 S R A
Po0' Bio
J b
BOP
N4830 lHlBam
BOB'
DC7 3bio10 imm21
BOP'
SA297 AJ1297
FIG. 2. Representation ofanintegratedkprophage (top line) and the deletions orsubstitutions in strainsused inthis work. N4830(M. Gottesman) isaKc1857lysogenwithadeletionfromtheBamHIsite intheintgenetoaBamHIsitenearclll.Prophagedeletion AHi removes
DNAfromcro into thebiooperonofE.coli, deleting all interveninggenesofX. BothDC7 andSA297 areF- rpsLhisrelA(D.Court and S.Adhya, unpublished data). The prophagein DC7is the result of the integrationofanimm2'bio transducingphageattheXattachmentsite. The A443endpointwasdeterminedtobewithingene0(S. Adhya, personal communication). The endpointof theprophage deletioninSA297
wasmappedbetween croandcll. The strain isCro+,but CII-.
were autoradiographed, and the DNA sequence starting at theA left cohesive end was determined.
RESULTS
Effect of the coslS4 mutation on A lytic growth in IHF-hosts. The mutant phage was found in a A strain that had undergone nitrosoguanidine mutagenesis (17) and had the characteristic of poor lytic growth on IHF- E. coli. The
phenotype of the mutant phage was demonstrated by the
EOP ofboth A cos+ and A coslS4 on a series of isogenic strains with mutations in IHF genes (Table 2). The cosl54
phage forms tiny plaquesonmostof the IHF-strains, and in the case ofsome IHF mutants shows a markedly reduced EOP.
IHF has beenidentifiedas amultimerof the himA and hip
geneproducts (32, 36; E. Flamm, personal communication).
E. coli with mutations in himA or hip are defective in A site-specific recombination (27, 29), suggesting that each
gene product does notfunction independently. In contrast, single point mutations in either himA orhip do not have a
severe effect on the EOP of X coslS4 (Table 2). This also
appearstobetrueforahip deletion andtoalesserextentfor
a himA deletion. However, the X coslS4 exhibits an EOP
that is 0.01% thatofX cos+ in E. coli with the double point
mutations himA42 and hip-157. These data suggest that either IHF subunit mayfunction independently to support lytic growth ofX cosl54.
Mutations in gyrB (himB) alter the ,B subunit of DNA
gyrase and reduce X DNA supercoiling. Because of this,
site-specific recombination is reduced in gyrB mutants (11, 15). A cos+ and X coslS4 were examined for their EOP on
[image:3.612.151.483.76.146.2]twostrainseach withasingle gyrB mutation andon astrain witha himA deletion and agyrBpoint mutation (K1942). A
TABLE 2. EOP ofX coslS4 on IHF- E. coli
EOPofphage
Strain Relevantgenotype X
cosl54
int6X
imm2
cimm.1
N99 him+ 1.Oa 1.0
K634 himA42 0.8 0.7
E268 hip-157 0.5 0.5b
JD12 himA42 hip-157 0.1 10-4
E444 Ahipl 0.9 0.6b
K1299 AhimA82 0.6 10-2
K1870 gyrB230 1.0 0.8
K807 gyrBI14 0.9 0.8
K1942 AhimA82gyrB230 0.4 10-3
aThe titerofeachphageonN99 at38°Cwasgivenavalue of 1. The EOPs onotherstrainsareexpressedasfractions of thisnumber.
bPlaqueswereverysmall withraggededges.
coslS4forms normal-sized plaqueson hosts with either the gyrB230orgyrB114mutation (Table 2). However, the EOP ofA coslS4on K1942 is 10% that of the EOP ofA coslS4on
E. coli witha himA deletion. The fact that the gyrB muta-tions alone exhibitno observable effecton the growth ofk
cosl54 leadsustoconclude that the gyrBmutation inK1942 amplifies the specific effect of the himA deletion. Recent findings indicate that the gyrB mutation decreases the burst of A (12). This may be the effect of the gyrB mutation in K1942.
coslS4, a cis-dominantmutation. Does the mutation have
an effect on a cis- or trans-acting component? This
question was addressed by performinga dominance test. A COS+ and coslS4 were used to infect N99 and K1942 (/himA82 gyrB230), eachat amultiplicity of infection of 5. The resulting burst sizes arepresented in Table 3. In N99, the burst size of each phage was comparable and not substantially altered upon mixed infection. In K1942, the
burstsize ofA cos+wasnotloweredsignificantly, but that of A coslS4 was decreased and did not increaseas aresult of mixedinfection. Thisindicates that thedefect of A cosl54 is notinatrans-acting function andrepresentsamutation ina site on the A chromosome or, less likely, a mutation in a cis-acting protein.
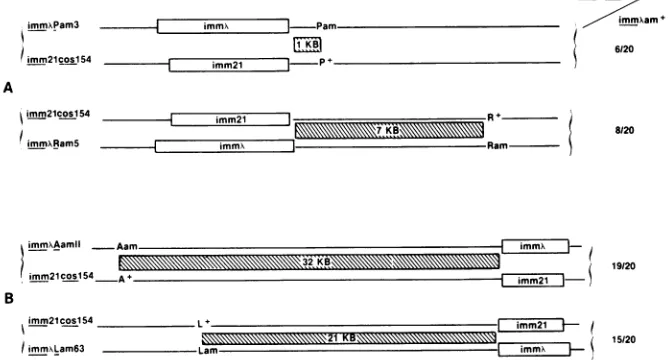
Genetic location of the coslS4 mutation. Phage crosses were used toposition the cosl54 mutationto either the left or right of the immunity region (imm). A diagram of the
crosses and the results are shown in Fig. 3. In cross A, A
coslS4 int6 imm21 phage were crossed with either A immX
Pam3 or
kimm'
RamS phage. Recombinants immX am+, inwhich the crossoverhad occurred between imm and either geneP or geneR (represented by the hatched area), were
selected by the ability to grow on DC7, a nonsuppressor
strainimmune toinfectionby phage 21. These recombinant
phagewerethenscreened for thecoslS4 mutationbytesting
for reduced EOP on K1942. No correlation was observed between the length ofthe crossover region (hatched area)
TABLE 3. Complementationstudies withX cos+ and kcoslS4 Burstofphageat32°C' Strain Genotype
Acos+imm434 X cosl54 cI857
N99 himA+ 67
N99 himA+ 54
N99 himA+ 60 17
K1942 AhimA82 19
K1942 AhimA82 1
K1942 AhimA82 32 1
aTheburst sizes at32°C after120 minweremeasuredbydeterminingthe
titerof thephageon K159(forburst ofimm'phage)andonK124(forburstof
iMM434phage). Theburst wascalculatedas thephage titerdivided bythe
number ofinfected bacteria.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.67.308.579.700.2] [image:3.612.326.565.611.695.2]immAPam3 imm Pam.
imm2lcos154 _ m
mK
imm2lcosl54 mm2l
imm\RamS =mm
immxAamll
imm21cos154
immXcosl54
/mmam+
6120
R7K
-AamiImmn
32K;B \ \ \\ \M3 19120
5imm21 |- )
B
imm2lcosl54 L
IimmxLam63 L
imm2l 21 K B
-l immA i
FIG. 3. GeneticmappingofAcoslS4.(A) Representationof therighthalfof thephagegenomeinphagecrossA.XcoslS4int6imm2' was
crossed with either Ximm'Pam3 or XimmX RamS.Twentyrecombinant phagefrom eachcrossthatwere imm"am+ werescreened forthe coslS4phenotype by examiningtheEOPonK1942.The number of coslS4phageisexpressedas afractionof the 20 immX am' phagethat
werescreened. (B) Representation of theleft half of thephagegenomeinphagecrossB.,XcoslS4 int6imm21wascrossed with eitherAAamll imm'orkLam63 immX. 20 recombinantimm'am' phagewerescreened for the coslS4phenotype by examiningthe EOPonK1942. Ineach
case the hatched lines indicate the area (given in kilobases) in which homologous recombination can occur to yield the immX am' recombinants.
and the frequency of k coslS4 recombinants. Thus, the
coslS4 mutation ismostlikely locatedtothe left of the imm region. To test this inference, cross B was done. A coslS4 int6
imm21
phagewerecrossed with either kimmX Aamll,orKimm'
Lam63 phage. Phage resulting from recombinationevents in either cross were selected as
imm' am'
by the ability togrow inDC7 as above. These recombinant phagewerethen screened for the cosl54phenotype byinabilityto
grow lytically on K1942. A direct correlation was found between the length of the crossover region and the
fre-quency ofK cosl54 recombinants. The high percentage of
recombinants with cosl54 in bothcrosses indicates that the
mutation is tothe left of the amber markers used and more tightly linkedtotheAgene.The rarercos+ recombinants(1 of20,5of20) presumably resulted fromasecond crossover between the selected amber markers and the cosl54 muta-tion. In cross A, double crossovers account for the rarer cosl54 recombinants foundin bothcases.
TheAgene isnearthe left terminusof the Kgenome,the
openreading framebeginningatbp +641 (4, 37). To position the cosl54 site more exactly, the cos+ allele was rescued from a cosmid recombinant molecule, pHC79 (20). This cosmid contains thewild-typeK cosregion inserted into the plasmidvectorpBR322. The first 415 bp from the left end of the A genome are included in this DNA molecule. C600, transformed with eitherpHC79 DNAorpBR322 DNA (asa control),wasinfected withA coslS4 int6imm2.Ifthe cosl54
site is within the first 415 bp of the A genome, then cos+ phage could arise by homologous recombination with the cosmid, but not with the pBR322 plasmid. To detect the recombinants, phage resulting from the infectionwereplated on K1942. In two independent experiments, the EOP of phage recovered from C600 containing pHC79was 10-fold higher than that of phage recovered from the infection of C600containing pBR322, indicating rescue ofthe cos+ site from thepHC79 DNA. To establishanindependentmeasure of marker rescue frequency from a plasmid, N4830 cells transformedwithaplasmidcarrying the intgene(pKC30int,
seeabove)wereinfected withA coslS4 int6
imm21.
A10-foldincrease inint+ phagewasdetected afterinfection, similarto thelevel ofrescue ofcos+ phage.
Molecular identification of the basechanged by the coslS4 mutation. The results of the genetic mapping experiments located thecosl54 site closetotheA left end. To identify the mutation at the nucleotide level, we determined the DNA sequence ofK coslS4 and the K cos+ recombinant phage generated in theK cosl54 int6 imm21x Aimmx Aamll phage
cross (Fig. 3, crossB).
Polynucleotide kinase and the Klenow fragment of DNA polymerase were used to 32p label the 5' and 3' ends of A DNA respectively. An Hpal restriction fragment that con-tains the first 734 bp of the A left end was isolated and purified from each labeled preparation. The base specific modification and cleavage reactions of Maxam and Gilbert (25) were employed, and the first 350 bp of each strand of eachphage wasdetermined.
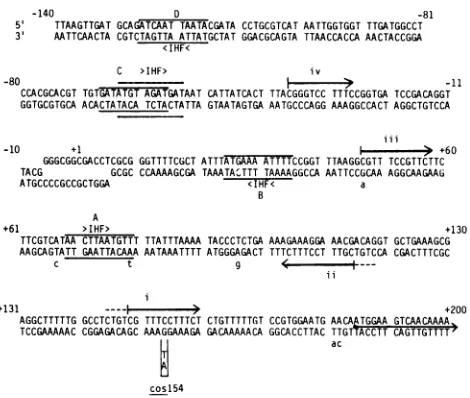
The DNA sequence of the cos+ recombinant was identicaltothepublished Awild-typesequence(4, 37).Only onechange in thesequenceof nucleotides from theA coslS4
phagewasdetected. This isindicated in Fig. 4atbp +154as aGC-to-AT transition(hence thename cosl54). The muta-tion is in a noncoding region upstreamof the open reading
frameof theleftmost Agene,Nul (37; bp+185 inFig. 4), in agreement with its cis-dominant phenotype. This change occurs within one of two 16-bp repeated sequences that comprisealarger palindromic region betweenbp +47 and bp +160 (Fig. 4, i and ii). Two variations of this repeated
sequencearefound closertothecosregion andarenoted in Fig. 4(iii andiv). Itappearsthatasingle transition event at bp +154isresponsible for theinability of themutant phage togrowlytically in IHF- E. coli.
Effect of cosl54onDNAencapsidation. The location of the mutationsuggested that the coslS4 mutationmight be defec-tivein someaspectof the DNApackaging reaction in IHF-hosts. We examined DNA packaging using a biological
assay, the Ter test (18, 34). The Ter test is based on the observation that a full-length genome can be packaged from tandem prophage in the E. coli chromosome after A
-i 15/20
A.-4-A+.
n ;s L
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.134.468.79.259.2]970 BEAR, COURT, AND FRIEDMAN
infection with an int- heteroimmune phage. Under these
conditions, theprophage remainrepressed and nonreplicat-ing. The heteroimmune infecting phage provides the prod-uctsnecessaryfor efficient cleavage andpackagingof the K
genome between thetwo cos sites in therepressed tandem prophage. In thecase ofa single prophage, packaging from thesinglecossite cannotencapsidateacomplete genome.
In our experiments, tandem multiple prophage of either
cos+ cI857 or coslS4cI857 were established in N99. The
AhimA82 deletion wasintroducedinto these lysogens by P1 transduction. If coslS4 interferes with packaging in IHF-hosts, the yield of this prophage compared with the cos+
prophage should be reduced.
Theresults of the Tertestwith int6red3
imm21
casthe heteroimmune infectingphage areshown in Table 4. In thehimA+ strain, no difference is found between thecos+ and
coslS4prophage yields. In the AhimA82 strain, the number of cos+ cI857 phage from the multiple lysogen is not significantly reduced. However, in the AhimA82 strain, the prophage yield from the coslS4 c1857 multiple lysogen
decreases to 10% of that from thehimA+ strain. Moreover, it is5% of thatfromthe cos+ cI857 multiplelysogen. We conclude from these data that coslS4 is dependent upon
IHFfor efficientpackaging ofits DNA.
Revertants of A cosl54. The plaques that arise at a
fre-quencyof
10-4
onthe himAhip double-pointmutant(Table-140 D -81
5S TTAAGTTGATGCAGATCAATTAATACGATA CCTGCGTCAT AATTGGTGGT TTGATGGCCT
3' AATTCAACTA CGTCTAGTTAATTATGCTATGGACGCAGTATTAACCACCA AACTACCGGA
<IHF<
C >IHF> iv
-80 >-11
CCACGCACGT TGTGATATGT AGATGATAATCATTATCACT TTACGGGTCC TTTCCGGTGA TCCGACAGGT
GGTGCGTGCAACACTATACA TCTACTATTAGTAATAGTGAAATGCCCAGGAAAGGCCACTAGGCTGTCCA iii
-10 +1 +60
GGGCGGCGACCTCGCGGGTTTTCGCT ATTTATGAAAATTTTCCGGTTTAAGGCGTTTCCGTTCTTC TACG GCGC CCAAAAGCGATAAATA'TTT TAAAAGGCCA AATTCCGCAA AGGCAAGAAG
ATGCCCCGCCGCTGGA <IHF< a
A
+61 >IHF> +130
TTCGTCATAACTTAATGTTT TTATTTAAAA TACCCTCTGA AAAGAAAGGAAACGACAGGTGCTGAAAGCG AAGCAGTATT GAATTACAAA AATAAATTTTATGGGAGACT TTTCTTTCCT TTGCTGTCCA CGACTTTCGC
c t g
ii
+131 ----l +200
AGGCTTTTTG GCCTCTGTCG TTTCCTTTCTCTGTTTTTGTCCGTGGAATG AACAJTGGAAGTCMCAACAAA
TCCGAAAAACCGGAGACAGC AAAGGAAAGAGACAAAAACA GGCACCTTACTTGTTACCTTCAGTTGTTTT7 ac
[image:5.612.329.570.98.170.2]cos154
FIG. 4. k cos+ DNA sequence surroundingthejunction of the
cohesiveends frombp -140tobp +200. The firstbptotherightof thecenterof the 12bpcohesive endjunctionis referredtoasbp+1. This DNA sequenceis identical to the published Asequence(37). The DNAsequenceof the first 350bpof the left end ofAcosl54was
determined. AsingleGC-to-AT transitionwasdetectedatbp+154.
Thismutationoccursinoneoftwo16-bpinvertedrepeatsequences
betweenbp +102 andbp+160(iandii).Ashorter12-bpderivative of thissequencebetweenbp+47andbp +58 is also marked(iii).A lessconserved11-bpderivative frombp-37tobp-27canbeseen aswell(iv).Potential IHFbindingsitesarealsoindicated(A through D). The IHFconcensusbindingsite is 5' AANNNNTTGAT3'(N. Craigand H.Nash, personal communication). Twopotential sites, each with 2bp changes, arenotedatbp -54andbp-57(C).Two
other derivatives of the IHF binding sequence, each with 1 bp change, are marked at bp +25 and bp +69 (B andA). The small letters indicate the additionalchangesdetected whenthesequence
ofthe bottom strand ofa cosl54revertantphagewas determined. Thestartoftheopenreadingframe for Nul isbp+185accordingto
[image:5.612.76.311.345.544.2]thissystemofbp numbering.
TABLE 4. Effect ofcosl54on k DNA packaging-results of the Tertest
Relevanthost Prophagea Prophageyieldper
genotype bacterium
himA+ (Xc1857),, 1.25
himA+ (A coslS4c1857),, 0.47
AhimA82 (Acl857),, 0.72
AhimA82 (A coslS4c1857),, 0.04
AhimA82 (A coslS4 c1857)1 0.0006
a Multiplelysogensareindicatedwiththesubscriptn. Ayield of1fromaX
cos+ c1857 tandem double prophage in himA+ E. coli is expected. The
prophageyieldfromasinglelysogen isshown for theAhimA82host.
2) or at afrequency of
10-3
onK1942(Table 2) are of normal size. However, the high frequency suggests that they do not represent true revertants.Four spontaneous "revertants" ofK cosl54 int6 imm21 were isolated from single plaques arising on K1942. These phage form plaques with equalefficiency on N99 and K1942. Eachphagewascrossed withKcos+Aamll immX (as in Fig. 3, cross B). Twenty-four immX am+ phage were isolated from eachcross(growthon asup°
Kimm21
lysogen,DC7)and were examined for the cos+ phenotype. All recombinant phage were cos+, indicating a tight linkage between the site of "reversion" in each phage andgene A.The DNAfromonecos+ "revertant" phage was isolated, and the3' endswere 32plabeled withthe Klenow fragment of DNA polymerase. An HpaI restriction fragment of the first 734 bp of the K left end was isolated and subjectedto DNA sequence analysis as described above. The DNA sequence from bp +13 to bp +226 was determined. The DNAfromthe phagecontainedthecos+ sequence(GC at bp
+154). In addition, it contained four transitions and two
transversions, noted by small letters at the appropriate
locationin Fig. 4. These numerous differences suggest that the DNA sequenceofthis phagederives fromrecombination with acryptic prophage, and thisrecombinationaccountsfor
thehigh "reversion frequency" ofKcosl54(Table2). Other
examplesof recombinationalrescueofcrypticcossites in E. coli have beenreported (10).
DISCUSSION
We have characterized a mutant of K, K cosl54, that exhibits reduced efficiency of plating on E. coli mutants
defectiveinaDNA-binding protein, IHF.Thiscis-dominant mutationwas genetically positioned close to the left cohe-sive end of the K genome. DNA sequence determination revealedasingle GC-to-AT transition event atbp +154ina
noncoding region between the A left cohesive end and the firstA gene, Nul. The mutation results ina defect in thein
vivopackagingof KcoslS4DNAinIHF-E.coli. Successful
packaging of X DNA is the result of multistep processes
requiring an initial cleavage at the cos site by A-encoded
terminase andsubsequent uptakeof the DNA intoanempty phage head. The coslS4 mutation may cause a defect in eitherorboth of these processes.
Factors involvedin coscleavage.The terminaseprotein(the
productof thephageNulandAgenes)binds andcleavesthe phage DNAat cos(9). Invitrothisreactionalsorequiresan E. coli-encoded protein(s) (2, 16). The requirement for E.
coli accessory factors can be satisfied either by a 22-kd
protein orby IHF,anE. coliproteininitially identified and
purifiedas ahostfactorrequiredforKsite-specific
recombi-nation. (The separateidentity ofthese two
proteins
has not J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
been
confirmed.)
Our observation that IHF is notrequired
for
packaging
ofKDNA invivo,
but isnecessary when thephage
carries thewcosl54mutation,
is consistentwith twomodels.
(i)
Hostproteins
areessential forDNApackaging.
However,
eitherthe22-kdprotein
orIHFcanbe used in thepackaging
reactionin vivo and in vitro.(ii)
Bacterialproteins
are not necessary for the
packaging
reaction. Invivo,
terminase alone may be sufficient for the entire
reaction,
whereasin
vitro,
hostproteins
arerequired
because condi-tions forpackaging
may besuboptimal.
In
considering
the role of the site definedby
the cosl54mutation
inthepackaging reactions,
it isimportant
tokeep
in mind the
position
of this mutation: a transition 154bp
tothe
right
ofthecenterof thecossite.Significantly,
theregion
of
hyphenated dyad
symmetryidentifiedby
coslS4 isoutside theboundariesoftheminimum substraterequired
forin vivoand in
vitro
coscleavage
and DNApackaging (9, 19, 33).
These boundaries extendfrom
bp positions
-40to +120ofthe cos
region (Fig. 4). However,
theexperiments
that havedefinedthese boundaries utilize assay systems that contain
IHFin vivoandin vitro.
Therefore,
itisnotknownwhether
terminase can cleave and
package
the minimum substrate DNA in the absence ofIHF.Role for E. coli proteins in DNA packaging. If an E.
coli-encoded
protein(s)
isrequired
forpackaging
invivo,
ourexperiments
withk cosl54 demonstrate that IHFisalikely
candidate.
However,
thefact
that k cos+phage
growlytically
in cells that cannotsupply
IHF suggests thatanother
protein
cansatisfy
thisrequirement.
An obvious candidate is the22-kdprotein
thatpromotescoscleavage
invitro.
According
to thisidea,
XcoslS4fails togrow in IHF mutantsbecause
thecosl54 mutationalters thebinding
site for the 22-kdprotein.
Thus,
IHFisrequired
to support theterminase reaction in the presenceofthe coslS4mutation.
If the
16-bp
sequence alteredby
coslS4 isrequired
forrecognition
by
the 22-kdprotein,
then we suggest that theminimum
substrate,
which lacks thissite,
alsorequires
IHFfor efficient DNA
packaging. Additionally,
wepredict
that the minimum substrate containsIHF-binding
sites. Exami-nation ofthe nucleotide sequence reveals that the DNA ofthe minimum substrate includes two
potential IHF-binding
sites(Fig.
4;
sites A andB)
each 1bp
different from the consensus sequence derivedby Craig
and Nash(personal
communication).
Moreover,
deletions ofthe minimumsub-stratethatremoveIHFsiteAdecrease
packaging efficiency
by
afactorof10(19).
Alternative role of the
cosl54
site. Examination of theregion flanking
the coscleavage
site reveals numerousrepeated
sequences(33)
(Fig. 4).
Infact,
the16-bp
sequencealtered
by
the cosl54 mutation isrepeated
in inverted orientation 26bp
tothe left(Fig. 4, ii).
Aderivative ofthis sequence is found closer to cos between the twopotentialIHF-binding
sites(Fig. 4,
iii between A and B). Anotherrelated sequence less conserved is
located
tothe left ofthecos
cleavage
site(Fig.
4,iv).
In the absence of directevidence foranE. coli
protein interacting
attheserepeatedsites,
it ispossible
thatthey
are instead thebinding
sitefor
terminase itself. The results of DNA footprinting
experi-ments
(6)
andsubstrate
binding
competition experiments(9) have demonstrated that terminase binds and protects thisgeneral region.
We
suggested
above that IHF may be required for coscleavage
orpackaging (or both) only
under conditions in which terminaseinteractionsareweakened(apossibleeffect of the cosl54mutation).
In these cases, IHF might berequired
to strengthen these interactions. A relationshipbetweenterminaseand IHF is suggested from studies ofthe
lambdoidphage 21. Phage 21 requires IHF for DNA
pack-aging in vitro and in vivo (Feiss, Frackman, and Sippy,
personal communication). This phenotype resembles that of theKcosl54 mutant and may be explained by the model that IHFstrengthensaweakerphage21-terminaseinteraction.In
fact,her mutants ofphage21 that revert thisphenotype and arepackaged in IHF- hosts have alterations ingene 1. This gene isanalogoustoNul and encodes a subunit ofphage 21 terminase(Feiss, Frackman, and Sippy, personal
communi-cation).
A model that proposes the cosl54 site to be the
termin-ase-bindingsite does not excludethepossibility of a function for the 22-kd protein isolated by Gold and Becker (16). It may also functionduringcoscleavage, butmay beunable to support the packaging of the cosl54 mutant DNA in the absence of IHF. Other repeated DNA sequences found in this region (33) may represent interaction sites of this
protein.
Substrate similarity between cos and alt. The evidence shows that IHF is involved in reactions that require a
site-specific cleaveage ofK DNA, i.e., integrative and excis-iverecombinationat thephageattachment site(att)and the
packagingreaction at cos. These events have similar prop-erties. Inboth,aphage-encoded proteinbinds and nicksthe
DNA; Int binds and cleaves the DNA at att (3), and terminase binds and cleaves the DNA at cos(9). Thegenes
encodingtheseproteinsarelocatedadjacenttothe sitewith whichthey interact (Fig. 1).
A comparison of the substrates used in these reactions reveals similarities. Bothcontainacoreregioninwhich the
nickingreactionsoccur.Inthe DNAsurroundingthe core of
att,
IHF-binding
sites are interspersed among bindingre-gions for Int(reviewed in reference41). Outside of the cos
core
region, potential IHF-binding
sites are situated amongproposed
terminase interaction domains (Fig. 4, A, B, C,and D and
i, ii, iii,
and iv). The recognition sites fortheseDNA-binding proteins
arelocated inboth arms surroundingtheatt(41)andcos cores
(Fig.
4). Intalso bindstothe core ofatt, but thebinding
is weaker andthesequence is differentthaninthe sites
surrounding
thecore(reviewed in reference 41). At the cos core,terminase presumably bindsto cleave the DNA. This recognition sequence for terminase is dif-ferentfromthebinding
sequence outside the core(7,9,33).IHFis anaccessoryprotein in K site-specific recombina-tion. Ithasno demonstrated cleavage activity, but bindsto
specific
sequencesneartherecombination site(N. CraigandH.
Nash,
personal communication) and promotes thebind-ing
ofthekIntprotein
nearby (36). Wesuggested abovethatIHF may also strengthen terminase binding in the cos
region.
Terminase protein from wild-type K may be veryefficient andnot
require
theenhancing
effectofIHF.Lambdasite-specific
recombination and phage 21 DNA packagingboth
require
IHF, but an independence of IHF can bedemonstrated in these systems as well. int mutants have been isolated that are
independent
ofIHF for site-specific recombination invivo,
e.g.,int-h3(31). Analogousterminasemutants(her)ofphage21 do notrequire IHFforpackaging.
Thereforethe
possibility
exists toisolate terminasemutants in theK coslS4background thatwill beindependentof IHF for DNApackaging.ACKNOWLEDGMENTS
Wethank D. Watson,M. Psallidoulos,and T.Robins for
assist-ance in determining DNA sequence; R. Weisberg for identifying potentialIHFbindingsites withincos; E. Flamm,K. Abremski, S.
on November 10, 2019 by guest
http://jvi.asm.org/
972 BEAR, COURT, AND FRIEDMAN
Adhya, and M. Gottesman for bacterial strains; and M. Feiss, S. Frackman, J.Sippy, N. Craig, H. Nash, and E. Flamm for furnishing results before publication. We appreciate the help of E. M. Olson with selectedexperiments. We also thank N. Craig for a critical review of this manuscript.
D.I.F. was supported by Public Health Service grantsfromthe NationalInstitutes ofHealth.
LITERATURECITED
1. Becker, A., and M. Gold. 1975. Isolation ofthebacteriophage lambda A-gene protein. Proc. Natl. Acad. Sci. U.S.A. 72: 581-585.
2. Becker, A., and M. Gold. 1978. Enzymatic breakage of the cohesive end site of phage X DNA: terminase (ter) reaction. Proc. Natl.Acad. Sci. U.S.A. 75:4199-4203.
3. Craig, N.L., and H. A. Nash. 1983. Themechanism of phageA site-specific recombination: site-specific breakage ofDNAby Inttopoisomerase. Cell35:795-803.
4. Daniels,D., J.Schroeder,W.Szybalski, F. Sanger, A. Coulson, G.Hong, D.Hill,G. Peterson, and F. Blattner. 1983. Complete annotated lambda sequence, p. 521. In R. W. Hendrix, J. W. Roberts, F. W. Stahl, and R. A. Weisberg (ed.), LambdaII. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y. 5. Enquist, L. W., and R. A. Weisberg. 1976. The redplaque test:
arapidmethodforidentification of excision defective variants of bacteriophagelambda. Virology72:147-153.
6. Feiss, M.,andA. Becker.1983. DNApackaging andcutting,p. 305-330. In R. W. Hendrix, J. W. Roberts, F. W. Stahl, and R. A. Weisberg (ed.), LambdaII. Cold SpringHarbor Labora-tory, Cold SpringHarbor, N.Y.
7. Feiss, M.,I. Kobayashi, and W. Widner. 1983.Separatesitesfor binding and nicking of bacteriophage X DNA by terminase. Proc.Natl. Acad. Sci. U.S.A. 80:955-959.
8. Feiss, M., and W. Widner. 1982. Bacteriophage X DNA pack-aging:scanning for the terminal cohesive end siteduring pack-aging. Proc. Natl. Acad. Sci. U.S.A. 79:3498-3502.
9. Feiss, M., W. Widner, G. Miller, G.Johnson,andS. Christian-sen. 1983. Structure of the bacteriophage lambda cohesive end site: location of the sites of terminasebinding(cosB)andnicking (cosN).Gene 24:207-218.
10. Fisher, R., and M. Feiss. 1980.Reversionofacohesiveendsite mutantof bacteriophage lambda by recombination witha defec-tive prophage. Virology107:160-173.
11. Friedman, D. I., E. J.Olson, D. Carver, and M. Gellert. 1984. Synergistic effect of himA and gyrB mutations: evidence that Him functions control expression of ilv and xyl genes. J. Bacteriol. 157:484-489.
12. Friedman, D.I., L. C. Plantefaber, E. J. Olson, D. Carver, M. H.O'Dea,and M.Gellert. 1984.MutationsintheDNAgyrB gene that are temperature sensitive for lambda site-specific recombination, Mugrowth, and plasmid maintenance. J. Bac-teriol. 157:490-497.
13. Friedman, D. I., and M. B. Yarmolinsky. 1972. Prevention of the lethality of induced X prophage byan isogenic X plasmid. Virology 50:472-481.
14. Furth,M., andS. Wickner. 1983. LambdaDNAreplication, p. 145-174. In R. W. Hendrix, J. W. Roberts, F. W. Stahl, and R. A.Weisberg (ed.), Lambda II. ColdSpring Harbor Labora-tory,Cold Spring Harbor, N.Y.
15. Gellert, M. 1981. DNAtopoisomerases. Annu. Rev.Biochem. 50:879-910.
16. Gold, M., and A. Becker. 1983.Thebacteriophage Xterminase: partialpurification and preliminary characterization of proper-ties. J. Biol. Chem. 258:14619-14625.
17. Gottesman, M., and M. Yarmolinsky. 1968.Integration-negative mutantsof bacteriophage lambda. J. Mol. Biol.31:487-505. 18. Gottesman, M. E., and M. B. Yarmolinsky. 1968. The
integra-tionand excision of the bacteriophage lambda genome. Cold Spring HarborSymp. Quant. Biol. 33:735-747.
19. Hohn, B. 1983. DNA sequences necessary for packaging of
bacteriophage X DNA. Proc. Natl. Acad. Sci. U.S.A. 80:7456-7460.
20. Hohn, B., and J. Collins. 1980. A small cosmid for efficient cloning of large DNA fragments. Gene 11:291-298.
21. Hohn, B., and T. Hohn. 1974. Activity of empty, headlike particles for packaging of DNA of bacteriophage X in vitro. Proc.Natl. Acad. Sci. U.S.A. 71:2372-2376.
22. Hoyt, M. A., D. M. Knight, A. Das, H. I.Miller, andH. Echols. 1982. Control ofphageXdevelopment bystability and synthesis ofcll protein: role ofthe viral clll and host hflA, himA and himD genes. Cell 31:565-573.
23. Kikuchi, Y., and H. Nash. 1978. Thebacteriophage X int gene product. A filter assay for genetic recombination, purification of Int, and specific bindingtoDNA. J.Biol. Chem. 253:7149-7157. 24. Lennox, E. S. 1955. Transduction of linked geneticcharactersof
the host bybacteriophage P1. Virology 1:190-206.
25. Maxam, A., and W. Gilbert.1980. Sequencing end-labeled DNA with base-specific chemical cleavages. Methods Enzymol. 65:499-560.
26. Miller, H. I. 1981. Multilevel regulation of bacteriophage X lysogeny by the E. coli himA gene. Cell25:269-276.
27. Miller, H. I., and D. I. Friedman. 1977. Isolation of Escherichia colimutants unable to support lambdaintegrative recombina-tion, p. 345-356. In A. Bukhari, J. Shapiro and S. Adyha (ed.), DNAinsertion elements, plasmids, and episomes. Cold Spring HarborLaboratory, ColdSpring Harbor, N.Y.
28. Miller,H. I., and D.I. Friedman. 1980. An E. coli gene product required forX site-specific recombination. Cell 20:711-719. 29. Miller, H. I., A. Kikuchi, H. A. Nash, R. A.Weisberg,and D.I.
Friedman.1979.Site-specificrecombination of bacteriophage X: the role of host gene products. Cold Spring Harbor Symp. Quant. Biol. 43:1121-1126.
30. Miller,H.I.,M. Kirk, andH.Echols. 1981. SOSinductionand autoregulation ofthehimA geneforsite-specific recombination inEscherichia coli. Proc.Natl.Acad. Sci.U.S.A. 78:6754-6758. 31. Miller,H.I., M. A.Mozola,and D. I.Friedman. 1980.int-h:an int mutation of phageX thatenhances site-specific recombina-tion.Cell 20:721-729.
32. Miller,H.I.,and H. A. Nash. 1981.Direct role ofthehimA gene product in phageX integration. Nature(London) 290:523-526. 33. Miwa, T., and K. Matsubara. 1982. Identification of the
se-quences necessary for packaging DNA into lambda phage heads.Gene 20:267-279.
34. Mousset, S., and R. Thomas. 1968. Dilysogenic excision: an accessoryexpression of the termination function? Cold Spring HarborSymp. Quant. Biol. 33:749-754.
35. Nash, H., K. Mizuuchi, L. Enquist, and R. Weisberg. 1981. StrandexchangeinX integrative recombination: genetics, bio-chemistry,andmodels. ColdSpring HarborSymp. Quant. Biol. 45:417-428.
36. Nash, H.,andC.Robertson. 1981.Purification andproperties of theE. coliprotein factorrequired forX integrative recombina-tion.J.Biol. Chem. 256:9246-9253.
37. Sanger, F., A. R. Coulson, G.F. Hong, D. F. Hill, andG. B. Petersen. 1982. Nucleotide sequenceofbacteriophageX DNA. J. Mol. Biol. 162:729-773.
38. Silhavy, T.J., M. L. Berman, and L. W. Enquist (ed.). 1984. Experiments with genefusions, p. 169-170.ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
39. Tomizawa, J.,and T.Ogawa.1969.Replicationofphagelambda DNA. ColdSpring HarborSymp. Quant. Biol. 33:533-551. 40. Wang, J. C.,and A. D. Kaiser.1973.Evidencethatthe cohesive
ends ofmature X DNAaregenerated by the geneA product. Nature(London) NewBiol.241:16-17.
41. Weisberg, R.,and A. Landy. 1983. Site-specific recombination in phage lambda, p.211-250. In R. W. Hendrix,J. W.Roberts, F. W.Stahl,and R. A.Weisberg (ed.),LambdaII. ColdSpring HarborLaboratory, ColdSpring Harbor,N.Y.
42. Wu, R.,and E. Taylor. 1971. Nucleotidesequence analysis of DNA.II.Complete nucleotide sequence ofthecohesive ends of bacteriophageX DNA. J. Mol. Biol. 57:491-511.
J. VIROL.