0022-538X/90/073471-15$02.00/0
Copyright© 1990, AmericanSociety for Microbiology
The Repressing and
Enhancing Functions of the
Herpes
Simplex
Virus Regulatory Protein ICP27
Map
to
C-Terminal
Regions
and
Are Required
To
Modulate Viral Gene
Expression Very Early in Infection
LINDAMcMAHAN AND PRISCILLA A. SCHAFFER*
Laboratory of TumorVirus Genetics, Dana-Farber Cancer Institute and Department of Microbiology and
MolecularGenetics, Harvard Medical School, Boston, Massachusetts 02115 Received 7 February 1990/Accepted 12 April1990
The phenotypic properties ofICP27temperature-sensitiveanddeletionmutantsand the results of transient expression assayshave demonstratedthatICP27hasamodulatory effectonviralgeneexpression induced by ICPs0and 4.Inordertoidentify the regions of theICP27molecule thatareresponsible for itsenhancing and repressing activities, 10nonsenseand 3 in-frame deletion mutationswereintroduced into the codingsequence
ofthe cloned ICP27 gene. Thesemutantgenes were tested in transientexpressionassaysfor their ability to complementanICP27 nullmutantandtoenhance andrepressexpression fromaspectrumof herpes simplex virus type 1promotersinreporterCATgeneswhen expressionwasinducedbyICPOorICP4. Theresultsof assayswithclonedmutantgenesdemonstrate that theICP27polypeptide containstworegions, located between
amino acidresidues 327and 407 and residues 465 and 511, that contributetoitsrepressing activity. The amino acid region located betweenthetworepressing regions (residues 407to465) is ableto interfere withICP27 repressingactivity. None of themutantgenesexhibitedefficient enhancing activity foranyof the herpes simplex type 1 promoterstested, demonstrating that amino acidscomprising thecarboxy-terminal half of theICP27 molecule, including theterminal phenylalanine residue,arerequired for wild-type enhancementaswellasfor efficient complementation of an ICP27 null mutant. Phenotypic characterization of an in-frame deletion mutant, vd3, and a previously isolated null mutant, 5dl 1.2 (A. M. McCarthy, L. McMahan, and P. A. Schaffer, J.Virol. 63:18-27, 1989), demonstratedthatICP27 is requiredtoinducetheexpression ofall classes of viralgenesveryearlyininfection and confirmedtherequirementfor ICP27laterin infection(i)torepress earlygeneexpression, (ii)toinducewild-type levels of delayed-earlyor-ylgeneexpression, and (iii)toinduce truelateory2geneexpression. The vd3mutant,whichspecifiesanICP27 peptidelacking the repressing region
betweenresidues 327and407, isableto(i)repressearlygeneexpression, consistent with the repressing ability
of thed3 mutation in transient expressionassays, (ii) inducethesynthesisofsignificant but reduced levels of delayed-early(yl)proteinsandnoy2 proteins (thus vd3 exhibitsalateprotein phenotype intermediatebetween
thatofthewild-type virusand 5dl1.2),and(iii)confer alteredelectrophoretic mobilityonICP4,demonstrating arole forICP27in theposttranslationalmodification of this essentialregulatory protein.Takentogether,these observations support a model in whichICP27 performsits regulatoryactivities differentiallyovertime and mediates these activities indirectly via interactions with, and modifications of, ICP4 andperhapsother viral and cellularproteins.
The genome ofherpes simplex virus type 1 (HSV-1) has
beencompletely sequenced(33-35, 42)and encodes 72 viral
proteins. These proteinsare divided into fourmajorkinetic classesonthebasisofrequirementsfor their
expression
and the times of their maximum rates ofsynthesis:
immediateearly, early, delayed early
(sometime
referred to as"leaky
late" oryl
proteins), andtruelate (-y2) (5, 25, 27,reviewed in reference 56). Viral genes of all four kinetic classes aretranscribedbythecellularRNA
polymerase
II(1, 6)andareexpressed inacoordinate and
sequential
fashionduring
the course ofproductive
infection(26).
Immediate-early
genes are the first to be expressedduring
thereplicative cycle,
followed by the early genes, whoseexpression requires
functionalimmediate-early
proteins (13, 26, 43).
Finally,
thedelayed-early andtrue late genes are
expressed.
Maximumexpression
of these lattertwo classes of genesrequires
theactivitiesof both
immediate-early
andearly
geneproducts
aswellasviral DNAsynthesis (24, 38).The
primary
difference*
Corresponding
author.betweendelayed-early (-yl)andtruelate(-y2)genesis that
-yl
gene
expression
does notrequire viral DNAsynthesis
(al-though DNAsynthesisisrequired for maximumexpression of -yl genes),whereas
expression
of-y2genes isstringently
dependentonviral DNAsynthesis (24, 38).
Immediate-early
genes encode the major HSV regulatoryproteins,
early
genescode forproteinsinvolved in viralDNAsynthesis,
anddelayed-early and late genes specify virion structural pro-teins.
Immediate-early genes are
expressed
in the absence ofprior viral
protein
synthesis (5).
Thesegenes aretranscrip-tionally activated by a
protein
(VP16) present inincoming
virus particles (2, 4). VP16, together with cellular factorsincluding the
octomer-binding protein,
formsprotein-DNA
complexes with the consensus sequence
TAATGARAT,
located in the 5'
promoter-regulatory
regions
of the fiveimmediate-early genes (4, 34, 42). The
immediate-early
genesencode infected-cell
polypeptides (ICPs) 0, 4, 22,
27,and47(41). Thelocationsof the genes
encoding
immediate-early proteinsin the HSV-1 genome are indicated in
Fig.
13471
on November 10, 2019 by guest
http://jvi.asm.org/
(top line). All immediate-early proteins except ICP47 are
nuclearphosphoproteins (57) and act toregulate their own
synthesisaswellasthe synthesisofproteinsof later kinetic
classes (9, 10, 13, 19, 31, 38, 39, 44).
Insight intothe functional roles of each of the immediate-early proteinsintheregulationof viralgeneexpressioncame initially from studies of viral mutants. Characterization of temperature-sensitive (ts) and deletion mutants in the five immediate-early geneshas shownthatonly ICP4 andICP27
are absolutely essential for virus replication (9, 10, 13, 29, 32, 43, 47, 48, 50, 54). Studies ofmutantshave also shown that ICP4 is involved in the down regulation of its own
expression as well as that of other immediate-early genes and isrequired throughoutthe viral replicative cycle to up regulate the expression of early, delayed-early, and late genesatthe level oftranscription (9, 10, 12, 13, 43). Studies ofmutantsin thegeneencoding ICP27have shown that this
protein is requiredtorepressexpression of certain
immedi-ate-early andearly genes and is absolutely requiredfor the
transcriptional activation of late genes, a function that is
independent of early protein and viral DNA synthesis (32, 46, 47). Compared with the wild-type virus, ICPO deletion mutants grow well and induce the synthesis ofwild-type
amounts of viral proteins at high multiplicities of infection but they grow poorly and induce reduced levels of viral
proteins atlow multiplicities (48, 54). Thus, ICPO playsan
important although nonessentialrole inproductiveinfection. ICP22appears tobe involved in the activation of lategene
expression and has been shownto be essential for efficient virus replicationinsomecell types but notinothers (50;C. Bogard, unpublished observations). ICP47isapparentlynot essentialfor viral growth, atleast in cell culture (29).
Transient assays that measure the regulatory effects of
individual immediate-early proteins on the expression ofa
reporter gene (chloramphenicol acetyltransferase [CAT]) controlled by various HSV promoters have confirmed the findingsobtained with viralmutants. Thus,ICP4was shown toactivateCATexpressioncontrolledby earlyand late viral promotersandtorepressexpressionfrom its ownpromoter (i.e., ICP4 is autoregulatory) by bindingto a specific hexa-nucleotide (ATCGTC) near the ICP4 transcriptional start site(10, 15, 19, 31, 39, 44). Although phenotypic analysisof ICPO deletion mutants has shown that this protein is not
absolutely requiredfor viralgeneexpression during produc-tive infection, transientexpression assayshave shown that (i)ICPO isapotenttransactivator of all four kinetic classes of HSVpromoters, as wellas a variety of cellularpromoters, and(ii)thetransactivating activityof ICPOforthese
promot-ersis increasedsynergistically inthepresence of ICP4(15, 19, 31, 38, 39, 44, 51, 52). The mechanism of ICPO-mediated transactivation is unknownat present.
Incontrast totheactivitiesof ICPOandICP4,ICP27 alone has littleregulatory activityfor HSVpromotersin transient expressionassays(9, 15, 51). Anexceptiontothisrule is the observation by Rice and Knipe that ICP27 by itself can
transactivate the delayed-early glycoprotein B (gB)
pro-moter (45). Despite its limited regulatory activity alone, ICP27 can both repress and enhance expression from a varietyof HSV-1promoters whoseexpression is induced by ICPOorICP4orboth (16, 45,51, 55).Thus, ICP27 actsas a modulator ofthe regulatory functions of ICPO and ICP4. Availabledata havenotestablished whether ICP27mediates its modulatory effects by direct interaction with ICPO and ICP4, indirectly through interaction withormodificationof
viral or cellularproteins, orby direct orindirectbindingto DNA. Althoughless wellstudied,neitherICP22 norICP47,
aloneorin combination with otherimmediate-early proteins,
has been shown to exert
regulatory
effects on viral geneexpression in transientexpressionassays.
Thus, of the five HSV-1 immediate-early proteins, only ICPO, ICP4, and ICP27 havebeen demonstrated to exhibit
significant trans-regulatory activity. Systematic
mutationalanalyses of the
coding
sequences for ICP4 and ICP0 haveidentifiedspecific regionsof theseproteinsthatareinvolved in their
respective
transcriptional regulatory
andDNA-binding activities,
sites ofphosphorylation,
and nuclear andintranuclear localization (3, 12, 17, 40). As noted above, ICP27 has at least two
major
regulatory
functions. It acts both as a repressor of selected immediate-early and earlygene
expression
andas anenhancer of late geneexpression.LikeICP27, the
immediate-early protein
ElA of adenovirusactsbothas a
transcriptional
repressorand activator of viralgene expression (37). Moreover, these activities are
speci-fiedby different
regions
of thepolypeptide (28).
In order to
assign
the functional activities ofrepression
and enhancementtospecific regions
of the ICP27molecule and togain
furtherinsight
into thesignificance
of these activities in the cascade of viral gene expression, we haveintroduced nonsense and in-frame deletion mutations into
the
coding region
of the cloned ICP27 gene. We havecompared
mutantICP27proteins
with thewild-type protein
fortheir
ability
tocomplement
anICP27 nullmutantandto repress and enhanceexpression
ofreporter genescoopera-tively
with ICPs 0 and 4. We have also characterized amutant
virus,
vd3,
containing
an in-frame deletion that has lost itsability
to enhancebut retained itsability
to repress HSV-1 geneexpression.
The results ofstudies ofmutantplasmidsin transient assays have
(i)
confirmedtheability
ofICP27 to trans repress and trans enhance
expression
from HSV-1 promotersactivatedby
ICPOandICP4, (ii)
localizedtwofunctionaldomains
responsible
for therepressing
activ-ity ofICP27 within the
hydrophobic carboxy-terminal
one-third of theICP27
molecule,
(iii)
demonstratedarequirement
for the
carboxy
half of the ICP27 molecule forenhancing
activity,
and(iv)
shown that theintegrity
ofthecarboxy-terminal
phenylalanine
residueofICP27isrequired
forbothenhancing activity
and efficientcomplementation
of anICP27 null mutant. Studies of the mutant virus vd3 have confirmed
previous
findings
that ICP27isresponsible
for adistinct shift in the
mobility
of ICP4 in sodiumdodecyl
sulfate(SDS) gels, implicating
ICP27in theposttranslational
modification of ICP4. Theability
ofICP27 tomodify
ICP4 was also shown to be distinctfrom its repressing activity.Finally,
studies of vd3 have revealed thatICP27performsitsrepressing
andenhancing
functions in a manner that isdependent
upon the time course of viral infection. Thus,ICP27 functionsas anenhancerofviral geneexpressionat a veryearly stage in infection and as repressorof early gene
expression
andan enhancer of late geneexpression atlaterstagesin the HSV-1 replication cycle.
MATERIALSANDMETHODS
Cells and viruses. African green monkey kidney cells
(Vero; ATCCCCL81)andICP27-expressing Verocells (3-3 cells[32])werepropagatedaspreviouslydescribed (47). The
wild-type
KOS strain of HSV-1, the KOS-derived ICP27deletionmutant5dl1.2(32),and amutant, vd3, generated in these studies were propagated and assayed as described
previously
(49).Plasmids. (i) Immediate-early
effector
plasmids. Effectorplasmids
containingtheimmediate-earlygenesforICPs0, 4,on November 10, 2019 by guest
http://jvi.asm.org/
and 27 of strain KOS were used in these studies. Plasmid pW3, which contains the ICP0 gene, was obtained from
Wendy Sacks (Dana-Farber Cancer Institute, Boston, Mass.) (48), andpnllND, which contains the gene for ICP4, wasobtained from Neal DeLuca (Dana-Farber Cancer Insti-tute) (11). (Please note that we have designated the ICP4-containing plasmid pnll, originally described by Neal De-Luca,as pnllND to distinguish it from theICP27-containing
plasmid pnll generated in these studies.) Plasmids pW3 and pnllND have been shown to transactivate HSV-1 early genes in transient expression assays (11). Plasmid pKHX-BH, which contains the ICP27 gene, was provided by
Stanley Person (Pennsylvania State University, University Park, Pa.).
(ii) CAT reporter plasmids. Chimeric CAT plasmids
pICP4CAT (also know as pIE3CAT [45]), ptkCAT, pVP5
CAT, and pL42CAT, which contain the CAT gene under controlof promoters for the genes encoding ICP4 (an
imme-diate-early protein), thymidine kinase (an early protein), VP5 (a delayed-early or -yl protein), and L42 (a true late or -y2 protein), respectively, were obtained from Neal DeLuca (10). pgBCAT, which contains the promoter of the g B gene (adelayed-early gene), wasprovided by Steven Rice (Har-vardMedical School, Boston, Mass.) (45). All enzymes and
restrictionenzymelinkers used to construct ICP27 nonsense and deletion mutants were obtained from New England
BioLabs,Inc. (Beverly, Mass.)and used as suggested by the
manufacturer. The promoters inserted into all of these
plasmidswerederived from the KOS strain of HSV-1. DNA isolation. Bacterial plasmid DNAs were isolated and
purifiedasdescribedpreviously (3). CsCl-banded KOS- and vd3-infected cell DNAs were isolated as described by De-Lucaetal. (8, 48).
Transfection.Fortransient expressionassays,transfection
of Vero cells was carried out by the calcium phosphate precipitation procedure followed by glycerol shock,as
pre-viously described (3). The amounts and types ofplasmid DNAsused in thesetests areindicatedin thetextandfigure legends. Contransfection ofICP27-expressing 3-3 cellswith 2 ,ug ofinfectious KOS DNA and 4
jg
oflinearized pd3mutant plasmid during efforts to generate the vd3 mutant virus was carried out as described previously for marker rescue (47).
Construction of ICP27 mutant plasmids. A series of 10 nonsense and 3 deletion mutations were introduced into
coding sequences of the cloned ICP27 gene in
plasmid
pKHX-BH. The locations of HSV DNA sequences inpKHX-BH relative totheviral genome and the locations of the mutationsin pKHX-BH are shown in
Fig.
1. Nonsense mutations in the ICP27 gene were generated as describedpreviously by
inserting
asynthetic
oligonucleotide
contain-ing a HpaI site and translational termination codons in all three
reading
frames5'GGCTAGTTAACTAGCC3'
HpaI
3'CCGATCAATTGATCGG5'
into five NaeI sites, four SmaI
sites,
and theunique Sall,
StuI, NsiI,EcoNI, andSacl sites in the cloned ICP27gene
(Fig. 1) (11). Since the
ICP27-containing
plasmid pKHX-BH
has no HpaI
cleavage
sites, HpaI
digestion
ofplasmids
containing
thesynthetic
oligonucleotide
results in linearmolecules,
demonstrating
the presence of theoligonucleo-tide in theseplasmids.
In-framedeletion mutations in theICP27
coding
sequence were constructedby
restricting pKHX-BH
with theindi-cated enzymes(Fig. 1), filling in or trimming the protruding ends, and adding appropriate synthetic linkers at the indi-cated sites. Forexample, the 5' protruding end of plasmid pdlwastrimmedand the 3' protruding ends of plasmids pd2 andpd3 were filled in with Klenow, and 12-base-pair (pdl), 8-base-pair (pd2), and 10-base-pair (pd3) BglII linkers were blunt end ligated to the respective plasmids. The ligated plasmids were then cleaved with excess BglII to ensure that each plasmid contained only one copy of the linker (pKHX-BH hasno BglII cleavage sites). The cleaved plasmids were religated with T4 ligase. The locations of the HpaI insertion mutations and of the limits of deletion mutations (Fig. 1) were confirmed by restriction enzyme analysis.
Complementation. Complementation of theICP27 deletion mutant 5dl 1.2 by peptides expressed transiently from cloned, mutated ICP27 genes was conducted in a manner similar to that described by Sacks et al. (47), except that Vero cells wereused instead of CV-1 cells and thefollowing proceduralmodifications were employed: 0.5 ,ug (instead of 0.3 ,ug) ofplasmid DNA and 4.5 ,ug (instead of 4.1 ,ug) of salmon testis DNA in 0.3 ml (instead of 0.2 ml) ofHEPES (N- 2-hydroxyethylpiperazine-N' -2-ethanesulfonic acid)-buffered saline was used to transfect -2 x 105 cells per 35-mm dish. Mutant 5dl 1.2 is an ICP27 deletion mutant that fails to replicate in Vero cells and expresses no detectable ICP27polypeptide (32).Transfected cells were infected with 5dl 1.2 24 h after glycerol treatment. After 18 h postinfection, infected cellswereharvested and progeny viruswasassayed on 3-3 cells toquantify total virus (i.e., virus generated by
complementationaswellasanyrecombinantsthat may have beengenerated) and on Vero cells to quantify any
replica-tion-competent recombinants. 5dl 1.2 DNA provides no sequence homology to the left of the BamHI site in the
ICP27-containing BamHI-HpaIinsert inplasmidpKHX-BH (32). As expected, no replication-competent recombinants between 5dl 1.2 and any of the plasmids tested were de-tected.
CAT assays. The in vitro assay for CAT activity was performed byamodification(10) of the method of Gorman et al. (21).
Southern blot analysis. DNA restriction fragments sepa-rated by agarose gel electrophoresis were transferred to
nitrocellulose and hybridized to labeled probes by the method ofSouthern(53). Theprobeused forSouthern blots wastheplasmid pKHX-BH
(Fig.
1).Theprobewaslabeled with [32P]dCTPand -dGTP (Dupont, NEN Research Prod-ucts, Boston,Mass.) by nicktranslation(30).Analysis of infected-cell polypeptides. Infected Vero cells werelabeled with [35S]methionine (Dupont, NEN Research
Products)asindicated infigurelegends. Lysatesof
radioac-tivelylabeledinfected-cell cultureswere
prepared
andelec-trophoresed on
SDS-polyacrylamide gels
as describedpre-viously (48).
RESULTS
ICP27mutantplasmids. Ten nonsenseand three deletion mutationswere introduced into the codingsequenceof the clonedICP27genein
plasmid pKHX-BH (Fig.
1).The genescontaining nonsense mutations
specified
ICP27peptides
composedof from 26(pn2)to511(pnll)amino acid residues of the 512-residue
wild-type protein.
Thesynthetic
linker used to generate nonsense mutations was also introduced into sequences downstream of the 3'transcription
termina-tion site of the ICP27 gene. Theresulting plasmid,
pnl4, specifies the wild-type ICP27 protein. The three deletionon November 10, 2019 by guest
http://jvi.asm.org/
ICPO ICP4 ICP47
IC PO _____---- ICP27 -. P22 ICP4
N N SNS BSI N BpSt NS EN S ScH
100bp, N N Nl- 8 N Na E S Sc
ICP27mRNA
ICP27codingsequence
UMMM/ 7/77777 /"//"/ 77 /"//
COMPLEMENTATION
INDEX 161
97
1.2 1.2 1.1 0.8 1 .o 1.5 1.9
9.2
9.7
X
*
[
c
0
z
I II II 1
345 6 7 8 910 1112 13
2
n14
iKHX-BH ,512
n2 - 26
n3 153
n4 163
nS 178
n. 261
n7 327
nS 407
ng nlO nIl
1.5 r dl
1.1 o d2
1.1 L d3
.460
-465
- 511
St Ns
I 512
407 460
Bp St
- I- 512
375 407
as St
-_ - 512
258 407
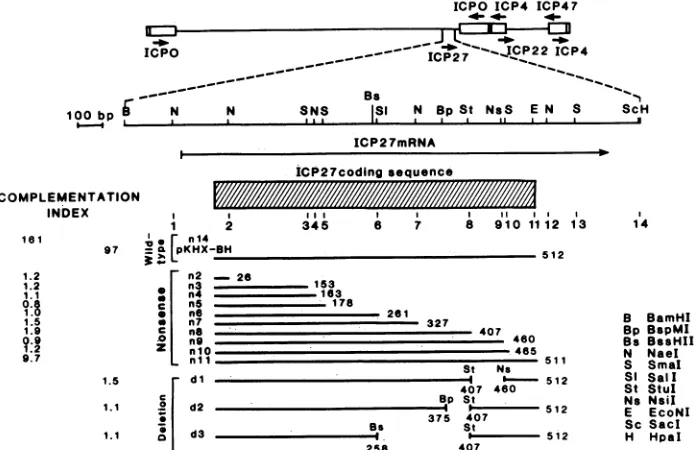
FIG. 1. Physical mapofICP27nonsense(n)anddeletion(d)mutationsandcomplementation of5dl1.2byICP27mutantplasmids. The locations of theimmediate-earlygenesencoding ICPs 0, 4, 22, 27, and 47areshownas arrowsontheHSV-1genomeatthe top of thefigure. Below thegenomeis anexpanded restrictionmap of theBamHI-HpaIviralDNAinsertinplasmid pKHX-BHusedtogeneratemutations in theICP27gene. Beneath theexpandedmap areshownthe limits ofthesequencesspecifyingICP27mRNA,thedirectionoftranscriptionof this mRNA, andthelocations of theICP27coding sequence. The numbered vertical linesbelow theICP27codingsequence indicate the locations ofnonsensemutations introduced into theICP27geneinpKHX-BHtoyield thenonsenseinsertionmutantplasmidsnlthroughn14. Theheavylines in diagrams ofnonsense mutantsindicatetheICP27 amino acidsequenceencodedbymutantplasmids. Theheavylines in deletionmutantdiagramsindicate the DNA sequences that encodein-frameICP27 amino acids.To the left ofmutantdiagramsareshownthe complementationindices(CI) ofmutantpeptides for5dl1.2.(CIequalstheratio oftheyieldof5dl1.2inVerocells transfectedwithamutant plasmidtotheyield of5dl 1.2inVero cellstransfectedwithpBR322.)
mutations removed viral DNA sequences that encode 53 (pdl)-, 32 (pd2)-, and 149 (pd3)-amino-acid residues in the
carboxy-terminalhalfof the ICP27 molecule.
ComplementationofanICP27 deletion mutant (5d11.2)by mutantplasmids. Inordertodeterminewhetherthe mutated ICP27 genes specify peptides with ICP27-associated activi-ties, mutant plasmids were first tested for their ability to
supply these activities by complementation of the ICP27
deletion mutant 5dl 1.2 (32). As shown in Fig. 1 (far left),
with the exception of the wild-type ICP27 protein, which produced complementation indices of 161 and 97 in two
independent tests, peptides specified by nonsense mutant plasmids pn2 through pnlO and deletion mutant plasmids
pdl, pd2, and pd3 failed to complement 5dl 1.2. Mutant plasmid pnll specifies a peptide in which the
carboxy-terminal
aminoacidresidue,
phenylalanine, was replaced bythree amino acid residues (Leu, Ala, and Ser) following
linker insertion. Plasmid pnll was able to complement 5dl 1.2 to only 6% of the wild-type level (complementation index, 9.7). These results demonstrate that one or more essential
ICP27-associated
activities were affected by eachofthe mutations in our series of mutant plasmids and that
replacementofthecarboxy-terminal-most amino acid,
phen-ylalanine, significantly reduced complementing activity for 5dl 1.2.
Repressing and enhancing activities of mutant plasmids. In order to identify regions of the ICP27 molecule that are
requiredfor its repressing and enhancing activities, mutant plasmids were tested for their ability to modulate the up
regulatory effects of ICPO or ICP4 on CAT reporter genes driven by various HSV-1 promoters. Reporter plasmids tested included ptkCAT, pgBCAT, pICP4CAT, pVP5CAT,
and pL42CAT. The mutant effector plasmids tested were
pn2, pn4, pn6, pn7, pn8, pn9, pnlO, pnll, pnl4, pdl,
pd2,
and pd3. pKHX-BH, which contains the wild-type ICP27 gene, andpnl4wereusedaspositivecontrols. Plasmidpnl4
contains a nonsense insertion mutation at the Sacl site outside the ICP27 coding sequence and, therefore, like pKHX-BH, specifies the wild-type ICP27 protein (see Sacl sitedownstream of the transcription termination site, Fig.1).(i) Repression of ptkCAT activity. Results of cotransfec-tions ofptkCATtogether with the various ICP27 alleles and
plasmids expressing either ICPO (pW3) or ICP4
(pnllND)
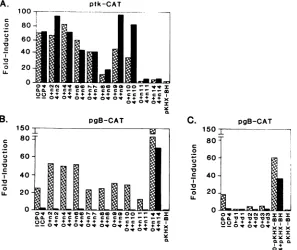
areshown inFig. 2A. Consistent with previous findings(10,
19, 31, 38, 44), both ICPO and ICP4 induced ptkCATactivity significantly above(-70-fold) the uninduced level, whereas wild-type ICP27 (pKHX-BH) exhibited no significanten-hancingactivity for the thymidine kinase (tk) promoter. Also consistent withprevious findingswastheability ofwild-type ICP27 (specified by pnl4) to repress ptkCAT activity in-duced by ICPO or ICP4 (51). The pn8 and pnll forms of ICP27 also exhibited significant repression, but repression
by pn8 was less efficient than that observed with the wild-type protein. The ICP27peptides encoded by pn6 and pn7
exhibited modestrepression of both ICPO- andICP4-induced ptkCAT activity, whereas neither the pn2 nor the pn4 form of ICP27 exhibited repression of either ICPO- or ICP4-induced activity. Interestingly, the pn9 and pnlO forms of ICP27 reproducibly exhibited no repression ofICP4-induced
activity, butthey repressed -30 and -40% ofICPO-induced ptkCAT activity, respectively.
These observations indicate that the 80 amino acids present in the pn8 protein but absent from the pn7 protein (Fig. 1) specify significant repressing activity for the tk
14
B BamHI
Bp BspMI
Bs BasHII
N NaeI S SmaI
Si SalI
St Stul
Ns Nail
E EcoNI
Sc SacI
H HpaI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.133.482.77.302.2]A. ptk-CAT
0 80 -i
~0
.E 40 o 20
U-0t CYCY t*CD r.- Q0a0a0oo -- t,*x
cc cc cc cc cc cc -_--m
--- Ot ow ov0) ovov ++ + +++ x
ov OVov x 9
QL
pgB-CAT
C.
C 0 ~0
r--I
0 LA.
-OV CM NV VZD IDf.- N octlD X O0 -IVV x
COCL
C cc4c
cccc---- --m20++ +0+++ ++ 0+0+ +C CC C
0V 0v X
ot0
tIC
OL150
80
60
40
20
0
pgB-CAT
22-ox*0
l+
+0I+
1m
m mSII
+
O1*
FIG. 2. Effect of wild-typeandmutant ICP27 peptides on tkCAT and gBCAT expression induced byICP0orICP4.Monolayers of Vero cellswerecotransfectedwith 2 ,ug of ptkCAT or 4 ,ug of the pgBCAT reporter plasmids and with 2 ,ug of pBR322, 3 ,ug of either pW3 (encoding
ICP0)orpnllND (encoding ICP4), 2
jig
of the indicatedICP27mutant plasmid, or 2jig
ofpKHX-BH orpnl4.(Note: ThepnllNDplasmid encoding ICP4isdistinct from thepnllplasmid encoding theICP27genemutated in the C-terminal phenylaline residue.) All transfections were brought up to 25 ,ug with salmon testis DNA. Transfected cells were harvested 2 days posttransfection, and cell extracts were assayed forCAT activity.Theuninduced level of ptkCAT or pgBCAT activity was determined bycotransfectionwithpBR322 alone with no effector plasmidadded (not shown). This uninduced level was arbitrarily set at 1.0. The "fold" induction of CAT activity in cotransfections with effector plasmidswascalculated relative to the uninduced value.promoter. Notably, addition to the pn8 peptide of 53 (pn9) and 58 (pnlO) amino acids interfered with the ability of the pn8 peptide to repress. Likewise, the addition of 46 amino
acids to the pnlO peptide to produce the pnll peptide
relievedthe interference with repression exhibited by pnlO
such that the pnll peptide exhibited wild-type repressing
activity. Thus, studies with nonsense mutations have iden-tified an 80-amino-acid repressing region (327 to 407), a
58-amino-acid region (407 to 465) that interferes with the repressingactivity of the 80-residue region, and a
46-amino-acid repressing region (465 to 511). Results with pnll also
demonstrate thatinsertion of the synthetic nonsense linker
into the codon specifying the carboxy-terminal
phenylala-nine residue did not affect the ability of ICP27 to repress
ICPO- orICP4-induced activity.
Deletion plasmids pdl, pd2, and pd3 all specify the
46-amino-acid repressing region of ICP27 (Fig. 1). The pdl plasmid also specifies the 80-amino-acid repressing region.
Bycontrast,pd2 lacks48(357to407) ofthese 80(327to407) amino acids and pd3 lacks all 80 residues. Notably, the
protein specified by pdl was more efficient than
wild-type
ICP27inrepressing ICPO-inducedptkCATactivity,whereas thepd2 protein was asefficientasthewild-typeproteinand
pd3waslessefficientthan
wild-type
ICP27in thisregard
(see Fig. 4, right-hand panel). The increasedrepressing activity
ofpdl relativetowild-type ICP27is,
bydefinition,
duetothe absence of residues 407 to 460. Such an effect is notunexpected whenone recalls thatthese residues interfered with therepressingactivity ofthepn8protein. The absence of48 residues of the 80-amino-acid
repressing region
inpd2
hadnoeffectontherepressing
activity
ofICP27(seeFig. 4,
right-hand panel),implying that the activity associated with this region is likely specifiedby residues 327 to 375. Sup-porting this view, the pd3 protein, which lacks the entire 80-residue region, was significantly less efficient than wild-typeICP27 inrepressingICPO-inducedptkCAT activity.The
residual repressing activity retained by the pd3 protein is
likelyduetothe fact that itcontains 45 of the 46 residues in the carboxy-terminal-most region that specifies ICP27 re-pressing activity. Taken together, these data suggest that repression of ICPO- and ICP4-induced ptkCAT activity in-volves residues 327to 375 and 465to 511.
(ii) Enhancement of pgBCAT activity. Previous studies haveshown thatICP27is abletofurther enhanceexpression ofpgBCATactivityinducedbyeither ICPOorICP4(45).We therefore attempted to identify the ICP27 amino acid se-quences responsible for this enhancement. The results of transient expression assays in which the gBCAT reporter gene was cotransfected with
wild-type
and mutant ICP27plasmidsandplasmidsexpressingICPOorICP4areshown in
Fig. 2B and C. In these tests, the wild-type form of ICP27 alone (specified by pKHX-BH) had no enhancing effect on thegB promoter, whereas
pgBCAT
activity
wasinduced 20-to25-fold by ICPO.ICP4 alone exhibited minimalenhancing
activity for the gB promoterunder the conditions used in thesetests. Althoughthe
wild-type
form ofICP27 specified by pnl4 enhanced both ICPO- and ICP4-induced pgBCAT activitysignificantly,
none of the nonsense(Fig.
2B)
or deletion (Fig. 2C) mutant forms of theprotein
enhanced ICP4-inducedactivity
andonly
pn2,pn4,
andpn6
enhancedICPO-induced
activity
(-twofold
above the control level).The failure ofpnlltoenhance
pgBCAT
activity
mustbe due150j
80 -B.
C
0
0
C
0I
0 LiL
60
-40
-20
-L
--
lttStE
E
tERI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.170.467.74.324.2]A.
c 0
0
r-10
IL
B.
C
0
0
r-70 UI.
C.
C 0
~0
U.
plCP4-CAT 30
-20
10
0 N v 0 N 0 a 0
0. c c c c c c -
-+ + + + + c c
0 0
I I
- CD
C x
0 X
°lI
O0 NVt VWDNCDCD0e C**00-- Vtt I
CL.0 c c c CC CC CC C C- --- -- m
22.+++t++ + c
__ 0t0t 0t 0t 0t 0t X
0C
FIG. 3. Effect of wild-type and mutant ICP27 peptides on
pICP4CAT,pVP5CAT, and pL42CATexpression induced byICPO
orICP4. Forexperiments withpICP4CAT, 2jigofpICP4CATwas
coprecipitated with either1
,ug
of pBR322, 1 pLgof pW3 (encoding ICPO),1jigof the indicatedmutantICP27plasmid inthepresenceof 1 pig of pW3, or 1Fg
of cloned pKHX-BH. Experiments withpVP5CAT and pL42CAT were performed and quantified in a
manneridenticaltothatdescribedfor ptkCAT andpgBCATinthe
legendtoFig. 2.
toeither the absence oftheterminalphenylalanine residueor
to its replacement by three amino acids brought about by linker insertion.
Taken together, these observations suggest that the
car-boxy terminus ofICP27isrequired for the efficient
enhance-ment of ICPO- and ICP4-induced transactivation of the gB
promoter.
(iii) Repression of pICP4CAT activity. ICPO is able to
induce expression from the ICP4 promoter (10, 39), and
ICP27
is able to repress this activity (38). Similarly, in ourhandsICPO enhanced pICP4CAT activity byapproximately
25-fold (Fig. 3A) and the wild-typeICP27protein specified
by pnl4 as well as the mutant peptide specified by pnll
repressed this activity with nearly equal efficiency.
Signifi-cant repressive activity was also seen with pn8. The other
mutantforms ofICP27 (pn2, pn4, pn6, pn7, pn9, andpnlO) exhibited no significant repression of ICPO-induced
pICP4CAT activity. These results are similar to those
de-scribed above for repression of the tkpromoterand support
the existence ofan80-amino-acid region between residues
327(pn7) and 407(pn8) anda46-amino-acid
region
between residues 465 (pnlO) and 511(pnll)
thatspecify
repressing
activity.
(iv) Repression of pVP5CAT and
pL42CAT
activity.
In order toidentify theregion(s)
of the ICP27peptide
respon-sible for the modulatory effectsofICP27 ontheexpression
ofrepresentative-ylandy2genes(51),
mutantpeptides
were tested for their ability to affect theexpression
of a-yl
(pVP5CAT) and a -y2(pL42CAT)
reporter gene.In these studies, ICP27 alone
(pKHX-BH)
exhibited noenhancing orrepressing
activity
for either gene, ICPO exhib-ited a strong enhancing effect on theexpression
of both reporter genes, and ICP4exhibitedmodestenhancing
activ-ity for pVP5CAT but not forpL42CAT
(Fig.
3B andC).
Wild-type ICP27, specified
by
pnl4,
and thepnll
mutantpeptide repressed the
enhancing
activities inducedby
both ICPO (for pVP5CAT and
pL42CAT)
and ICP4(for
pVP5CAT) to uninduced control levels. Theability
tore-press ICPO- and ICP4-induced activation of
pVP5CAT
wasalso a property of the pn8 mutant
peptide
and to a lesserextent of thepeptides
specified
by
pn7
andpnlO
(for
ICPO-induced activity). The relative
strengths
oftherepressing
activities of the various ICP27 mutantpeptides
for ICPO-induced pL42CAT activity wasnearly
identical tothat seenwith ptkCAT and pICP4CAT.
(v) Verification ofthe repressing activities ofmutant pep-tides. In order todetermine
definitively
whether the repress-ingactivity exhibitedbywild-type
plasmids
pKHX-BH
and pnl4 and mutant plasmidspn8,pnll(Fig.
2A),
pdl,
pd2,
and pd3 is aconsequence oftheactivities ofthecorresponding
ICP27 peptides, additional tests were conducted with
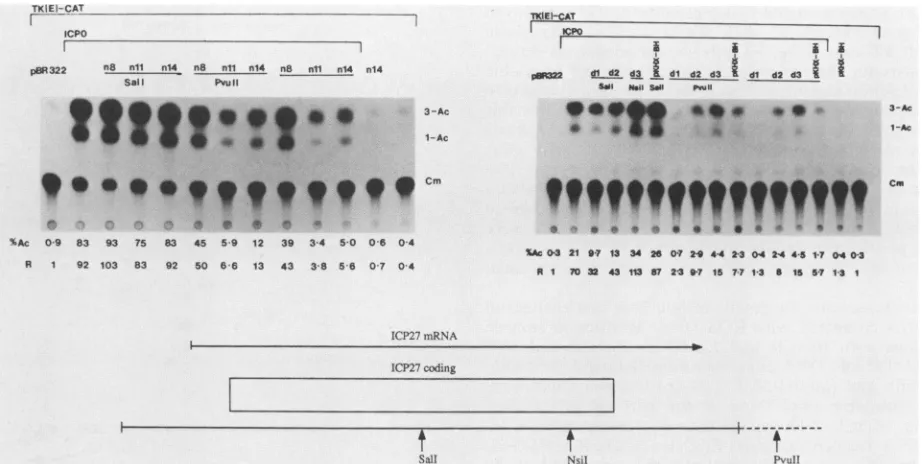
ptk-CAT (Fig. 4). Inthesetests, effector
plasmids
werecleaved withinICP27 coding sequences(Sall
forpnl4,
pKHX-BH,
pn8, pnll, pdl, and pd2; NsiI for
pd3)
or inplasmid
sequences with PvuII, before cotransfection of cells with ptkCAT and pW3 (see Fig. 1 for relevantcleavage
sites).
Cotransfection of SalI- orNsiI-digested
plasmids
with the ICPO plasmid did not result inrepression
ofICPO-inducedptkCATactivity (i.e., CAT
activity
similar tothatproduced
incontrol cellstransfected withthe ICPO
plasmid
alonewas observed). Therefore,interruption
ofICP27
coding
se-quences in the wild-type and mutantplasmids
impaired
the ability of the peptides they encode torepress ICPO-inducedptkCAT activity. In contrast, cotransfection of
PvuII-di-gestedplasmids with the ICPOplasmid
resulted inrepression
of ICPO-induced ptkCAT
activity (i.e.,
CATsignals
were similar to those of theundigested
plasmids).
Thus,
the repressing ability of the ICP27proteinspecified
by
wild-type
plasmids pKHX-BH and n14 and mutant
plasmids
pn8,
pnll, pdl, pd2,andpd3requiresthe
expression
oftheICP27
proteins they encode.One may argue that the
repressing
activity
of the mutantforms of ICP27 is not the residual
activity
ofICP27 but atrans-dominant negative phenotype converted from a
posi-tive activity. If this were the case, this trans-dominant negative activity should have a negative effect on the
gB
promoter, in contrast to wild-type
ICP27,
whichdisplays
anenhancing effect. We didnot observe sucha
negative
effect,in that mutants which lost the ability to activate
pgBCAT
(e.g., pn8) did not acquire theability
to represspgBCAT
(Fig. 2B). On the other hand, the mutants resembled wild-typeICP27 in that theyrepressedexpression
from the ICP4, tk, VP5, and L42 promoters.Therefore,
the mutantpheno-types reflect the repressing activity ofthe
wild-type
ICP27
molecule and can be used legitimately to map theregion
involved inrepression.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.91.258.70.423.2]TKIEI-CAT
F----ICPO
pBR322 nB8 nil n14 fnB nil n14 n8 nll M4 n14
Sa Pvuli
IIIf*lag
0 0 it0
1p0
3-Ac
1-Ac
It
* * *t *tit
t
t9
9
Cmle " I.. - -I 0 0e e e .
%Ac 09 83 93 75 83 45 5-9 12 39 3-4 5-0 0-6 0-4 R 1 92 103 83 92 50 6-6 13 43 368 56 0 7 0 4
ii'
i'
i_322 dt d2 d3 1 dl d2 d3 I dl d2 d3 1 I
Soli 1 WSI PWII
*--3
*,*
?
Con
# .- .*e 4 a * . * * *0 Cv *
%Ac03 219713 34260-7 2 44 23 04244517 0403 R 1 7032 43 113 87 2 t-7 157-7 1-3 t51557 1-31
ICP27inRNA ICP27coding
Sall NTsl Psull
FIG. 4. Effectof intra- and extragenic cleavage of ICP27 mutant plasmids on ptkCAT expression induced byICP0.Prior to transfection, plasmids eitherwere cleaved withSall,NsiI,or PvuII or were not cleaved. Shown below the autoradiogram is the ICP27coding sequence andthe locations ofthe relevantrestrictionenzyme cleavage sites. This experiment wasperformed in a manner identical to that described inthelegend to Fig. 2 for ptkCAT.
Collectively, the results of transient expression assays
demonstrate that ICP27, which alone had little or no effect ontheexpression ofanyof the HSV-1 reporter genes tested, acts as arepressorofICPO- and/orICP4-inducedexpression
ofpICP4CAT, ptkCAT,pVP5CAT,andpL42CATandas an
enhancer ofpgBCAT activity induced by ICPOorICP4. The resultsof transient expressionassayswith theICP27mutant
plasmids are summarized in Fig. 5. The following
conclu-sionscanbedrawnfromthesestudies: (i)tworegionsof the
ICP27 molecule exhibit repressing activity-one region be-tweenamino acid residues327 and 407 andasecond between
residues 465 and 511, (ii) residues 407 to 460 are able to
interfere with the repressing activity specified by residues 327to407,(iii)manyregionsof thecarboxy-terminalhalfof the ICP27 protein are critical for the enhancing effect of
ICP27onthe gB promoter andforefficientcomplementation ofanICP27 deletionmutant, 5dl 1.2,and(iv) substitution of
a nonsense codon specifying three amino acids for the
carboxy-terminal phenylalanine codon destroyed the en-hancing activity of ICP27 but did not affect its repressing
activity.
Introduction ofthe d3 mutation intotheviral genome. As shown in Fig. 4, the mutant peptides encoded by deletion
plasmids pdl, pd2, and pd3 were able to repress
ICP0-induced ptkCATactivity in transientexpression assays. Of the three mutants, therepressing activity of pdlwasgreater thanthatofthewild-type ICP27protein and that ofpd3was somewhat less than thatofwild-type ICP27 (Fig. 4).Because the d3 mutation exhibited a repressing phenotype midway
between thatofwild-type ICP27and the nullphenotype (Fig.
4,right-handpanel),thed3 mutationwastransferred into the viral genome by marker transfer. Since the pd3 mutant
plasmid failed to complement the ICP27 deletion mutant
5dl1.2 in transient complementation assays (Fig. 1, left
ICP27coding sequence R
I I 2
512
3 407 1 1
repression | |
Iepression
Of tk,ICP4 Activation (P5 andL42 ofgB
promoters promoter
! + +
J5i1 4+
repression
Complementation
of 5dl 1.2
+
(t)
6%dII~-512 +9
407 460
Deletion d2 1 512 +
375 407
d3 2- 512 +
-L ~~~~258 407
FIG. 5. Locations oftheICP27primaryaminoacid sequencesresponsibleforrepressionof CATactivityfromreportergenes. Beneath the boxrepresentingtheICP27codingsequenceareshown thelocations of thenonsenseinsertion(n)and deletion(d) mutations in mutant plasmidsthatdefinefunctionallyimportantregionsof theICP27molecule. Therepressiveandenhancingactivities of mutantICP27plasmids areshowntotheright ofthefigure.
3-Ac I-AC
Wild-type
n7
n8 Nonsense
nil
nli 1.
-6
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.74.535.73.305.2] [image:7.612.136.488.558.684.2]column), it was assumed that replacement of the wild-type
ICP27
gene with the d3 allele would result in a nonviablevirus.ICP27-expressing 3-3 cells used previouslyas
permis-sive hosts for the isolation and propagation of nonviable
ICP27
deletion mutants (32)weretherefore used inefforts totransfer the d3 mutation into the viral genome. For this purpose, 3-3 cellswerecotransfected with thelinearizedpd3 mutant plasmid and infectious KOS DNA. Progeny virus from the cotransfected culture was plated on 3-3 cells, and
individual plaqueswerepickedand screenedfortheir ability to grow in 3-3 and Vero cells. Plaque isolates that exhibited impaired growthinVerocellsbutgrewwell in 3-3 cellswere
plaque purified three times and amplified in 3-3 cells, and a
single isolate, designated vd3, was chosen for further
anal-ysis.
Figure 6 presents the results of Southern blot analysis of vd3 DNA compared with KOS DNA. Restriction enzyme
digestions with BamHI and EcoNI or BamHI and NsiI showed that vd3 DNA contained BamHI-EcoNI (1.50-kilo-base [kb]) and BamHI-NsiI (1.34-kb) fragments that were
indistinguishable from those of the parental pd3 mutant
plasmid, being 0.45 kb smaller thanthe corresponding wild-type DNA fragments (BamHI-EcoNI, 1.95kb;BamHI-NsiI, 1.79 kb). This analysis demonstrates that viral DNA ofthe vd3 mutant contains the deletion present in the parental plasmid.
Phenotypic analysis of ICP27 deletion mutants vd3 and
5dll.2.(i) Growth properties. The results of single-cycle growth experiments shown in Table 1 demonstrate the following points. (i) Whengrown and assayed inVero cells, no infectious vd3 orSdll.2 was detected, reflecting the nonpermissivenessofVerocellsforthesemutants. Thisisin contrast to KOS, which grew and platedwell in Vero cells. (ii) When grownin Verocells and assayed in 3-3cells, both vd3 and 5dl 1.2 yielded low titers of virus. This virus was likely residual unadsorbed mutant virus, since the plaques were noticeably smaller than KOS plaques on 3-3 cell
monolayers. (iii)TheICP27-transformed cell line, 3-3,
com-plementedboth mutants efficiently, as demonstrated by the
titers and plating efficiencies of the mutants on these cells.
(iv) When vd3 was grown in 3-3 cells and assayed on Vero cells, no detectable wild-type recombinants were observed. This demonstrates that although vd3 DNA contains
se-quences flanking the deletion that are homologous to the ICP27-containing fragment resident in 3-3 cells (32), the
superinfecting mutant virus did not recombine detectably
with residentICP27 sequences.
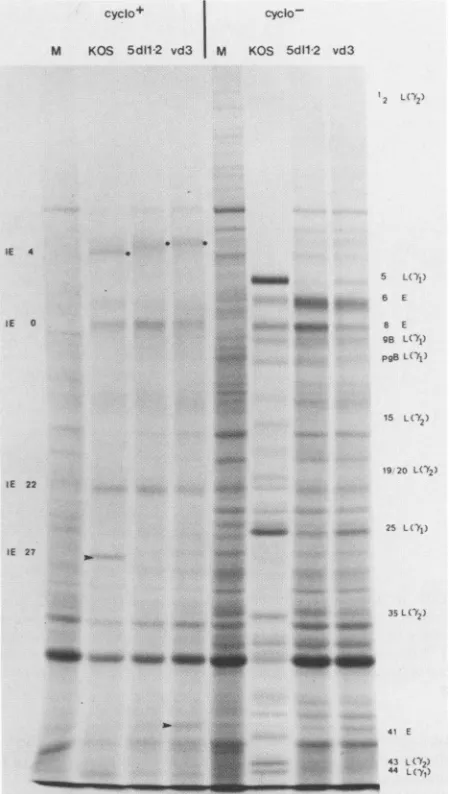
(ii) Immediate-early polypeptide synthesis. Figure 7
(cyclo+) shows the immediate-early polypeptide profiles of KOS-,5dl 1.2- and vd3-infected Verocellsinthepresenceof
dactinomycinfollowing removal ofcycloheximide.Although
KOS-infected cells contained significant amounts ofICP27
(Fig. 7, arrow), 5dl 1.2 produced no detectableICP27 polypeptide. This isconsistent with thegenotype of5dl 1.2,
whichlacks the transcriptional start site as well as portions
ofthepromoterand codingsequence of theICP27gene(32).
As expected, vd3, which contains an in-frame deletion
mutation inICP27 coding sequences (Fig. 1), did not
pro-duceabandthat migrated in the position of wild-typeICP27
butratherproducedaprominentband of the size expected of thetruncated vd3ICP27 polypeptide.
In HSV-infected cells, ICP4 is often present as multiple
electrophoretic forms, presumably as a consequence of different levels of phosphorylation ofthe ICP4 peptide (13, 41, 57). Recently, Riceand Knipe observed achange inthe
electrophoretic mobility of ICP4 in cells infected with an
BamHI+EcoNNI BamH1+NsII
I I
OU) Y Co
0 ;> Y Ml In > Y CL
._,_ _ 441
-4426
-40 -o:1-95
If 4179
*b
- -c1-Sa -a1.34
-Dl3
_
_---JPC P 2
bP8 St Nm E H
TCP27,mRNA
pKHX BH
B BamHl
BsB ssHTI
StStuI
Ns Nsil
E EcoNI
H HpaI
Bs
d3
-258
512 St
- 5 1 2
407
FIG. 6.
Southern
blot analysis of vd3 and KOS DNAs relative to the viral DNA inserts in plasmids pd3 and pKHX-BH. Viral (vd3 and KOS) and plasmid (pd3 and pKHX-BH) DNAs were digested with eitherBamHI
and EcoNI (left-hand lanes) or BamHI andNsiI (right-hand lanes). The digested DNAs were electrophoretically separated on a 0.7% agarose gel, transferred to nitrocellulose, and probed with32P-labeled
pKHX-BH. Sizes of DNA fragments in kilobases are indicated on the right of the autoradiogram. Shown below the autoradiogram in descending order are a diagram of the KOS genome, an expanded diagram of theBamHI-HpaI viral DNA insert in plasmid pKHX-BH, the locations of relevant restriction sites, the location and direction of transcription ofICP27 mRNA, the location ofICP27
coding sequences, and the genotypes of the wild-typeICP27
gene in pKHX-BH and the d3 deletion mutation in plasmid pd3.ICP27
ts mutant (45). This implies thatICP27 may be involved either directly or indirectly in the posttranslational modification of ICP4. In our hands, the ICP4 species syn-thesized in cells infected with5dl
1.2 or vd3 migrated more slowly than the ICP4 species synthesized in KOS-infected cells. This difference was also seen in immunoprecipitation experiments in which monoclonal antibody to ICP4 was used (data not shown). Although the d3 mutation is able to repress viral gene expression as described above, it does not confer wild-type-like electrophoretic mobility on ICP4. This indi-cates that the repressive function associated withICP27 is distinct from the function that affects ICP4 electrophoretic mobility. No significant alteration was detected in the quan-tity of ICP4 or in the quantities or electrophoretic mobilities of ICPs 0 or 22 in mutant infected cells.on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Growth of KOS, vd3, and
5dl
1.2 in Vero and ICP27-transformed cellsVirusyield (PFU/ml) when grownina:
Virus Vero cells 3-3 Cells
Vero 3-3 Vero 3-3
KOS 7.6 x 107 1.0 X 108 1.2 x 108 1.3 x 108 vd3 <101 5.4 x 103 <1o2b 5.1 x 107
5dl 1.2 <10' 4.3 x 101 <102 7.6 x 107
aMonolayers of Vero or 3-3 cells were infected with the indicated virus at a multiplicity of infection of 5 PFU/cell. At 18 h postinfection, infected monolayers wereharvested by scraping cells into medium. Suspensions were thenfrozen, thawed, sonicated, and clarified by low-speed centrifugation. Supernatant fluids were assayed on Vero monolayers.
bWhen vd3 and5dl 1.2 were grown in permissive 3-3 cells and assayed on nonpermissive Vero cells, monlayers of the latter cells exhibited uniform nonspecific cytopathic effects at low dilutions; this was likely due to cell killing at high multiplicity infection.
(iii) Viralpolypeptidesynthesis. Previous studies ofICP27
ts anddeletion mutants and the results of transient expres-sion assays have suggested that ICP27 is required to down regulate early gene expression and to enhance late gene
expression. To evaluate the effect of the d3 and 5dl1.2
mutations on the synthesis of early and late viral
polypep-tides,mutantandwild-type infected Vero cells were labeled
fromrr
5 to 17 h postinfection with [35S]methionine (Fig. 7,cyclo-).
The results of this experiment indicate that ICP27 is
requiredtorepress the expression of some early genes and to
activatetheexpressionof others. For example, cells infected
with the ICP27 null mutant 5dl 1.2 overproduced early
proteins (ICP6 and ICP8) but underproduced the early protein ICP41 relative to wild-type virus. Consistent with
previous observations, 5dl 1.2 induced the synthesis of
markedly reduced levels ofproteins of the -yl class (ICP5,
-25, and -44) and no true late (-y2) proteins (ICP1/2, -15, -19/20, -35, and -43) (32). Like 5dl 1.2, vd3-infected cells underproduced ICP41 and failed to produce true late (y2) proteins. The synthesis of pgB and gB relative to KOS-infected cells was notgreatly affected ineither 5dl 1.2- or
vd3-infected cells in these experiments, and host protein synthesis was not efficiently shut offincells infected with
eithermutant. Althoughvd3 isphenotypicallysimilar to5dl
1.2, the two mutantsdiffer intworegards. vd3 induced the
synthesis of early proteins (ICP6 and -8) and especially ICP6,atalevel lower thanthatof5dl1.2but
slightly
higher
than that of wild-type virus, and it induced some(yl)
proteins (e.g.,
ICP5
andICP25) atlevelslower than thatof wild-type virusbut slightlyhigherthan thatof5dl 1.2.Three conclusions canbe drawn with
regard
to the phe-notype of vd3, based on thepolypeptide profiles
shown inFig. 7 (cyclo-). (i)The ability ofvd3torepress
early
geneexpressiontolevelsbetween those of5dl 1.2and
wild-type
virus correlates withtherepressingability ofthepd3mutant
plasmid
for the early target gene tkCAT in transfectionassays (Fig. 4,
right-hand
panel).
(ii)
Unlikepreviously
described ICP27 ts and deletion mutants
(32, 47),
vd3 induces the synthesis ofsignificant
levels of(-yl)proteins
(i.e., ICP5 and ICP25). (iii) Like theICP27 ts and deletion
mutants (32, 47), vd3 is unable toup
regulate
true late(-y2)
geneexpressionand isdefectivein theshutoffof hostprotein
synthesis.Inorder toestablishthat the viral
polypeptide phenotype
ofvd3isaconsequence of the absenceofICP27andnotthe
effect of mutations in other
regions
of the genome, thecyclo+
cyclo-M KOS 5dl12 vd3 PM KOS Sdil-2 vd3
:.- ovo *.s-.
.:....z
:t,'-{Ea
IE22
IE 27
U.-::mm,T
5 L0513
6 E
8 E
96L(31)
pgRL(f1)
15 LO2)
19 20 L.(C2)
25 L(YI)
35L02)
41 E
43
L...
-.
FIG. 7. Polypeptide profilesofKOS-,
5dll.2-,
and vd3-infected cells.Left-handlanes(cyclo+);Verocellmonolayers
werepreincu-batedfor1hat37°Cinthe presenceof75 ,ugof
cycloheximide
per ml. Theywere theninfectedwith 10 PFUoftheindicated viruses percell in thepresenceofthesameconcentration ofcycloheximide.
At 6 h
postinfection,
cycloheximide-containing
medium was re-moved andmonolayers
were washed and overlaid with mediumcontaining
10 ,ug ofdactinomycin
per ml and 40,uCi
of [35S]methionine per 35-mm
plate.
After a further 3 h of incubation,infectedcellswere harvestedand
analyzed by
SDS-polyacrylamide
gel
electrophoresis.
Right-hand
lanes(cyclo-);
Verocellmonolay-ers wereinfectedwith 10 PFU percellat
37°C,
labeledwith thesame concentration of[35S]methionine
indicated above from 5 to 17 hpostinfection,
andprocessed
forSDS-polyacrylamide gel
electro-phoresis
analysis.
The SDSpolypeptides
wereelectrophoretically
separated
in 7%polyacrylamide gels.
Selectedimmediate-early
infected cell
polypeptides (ICP4,
-0, -22,and-27)areindicated intheleft-hand
margin.
Selectedearly
(ICP6,-8, and-41);delayed-early
(yl: ICP5, gB, pgB, -25, and -44); andtrue late
(y2:
ICP1/2, -15,-19/20, -35,and-43)
polypeptide
bands areindicated onthe right-hand sideofthefigure.
mutation in vd3wasrescued
by
standardprocedures
(47)
by
using
theBamHI-HpaI
DNAfragment
inplasmid
pKHX-BH
(Fig.
1).
The rescuedvirus, vd3R,
exhibitedwild-type
polypeptide
profiles
in bothcycloheximide
reversal and22
LC;.f.';:
.fm...:..z ik- ..;!=-11. xs'I..;f:
on November 10, 2019 by guest
http://jvi.asm.org/
B
27~14
.::..
2h |8 6 h | t M KcZS %,%,2wo 3 M. KO6 5drzIA 3 M -CSSd2'ld 3
5 LI-',)
6e:
8F
pg :01:
pgBL:"rl
25; r: 27It.
39F 43e0e-rI
.7~~~~~~~~~~~~~~L*88361
.0 - ||
IF~5..
FIG. 8. Viral polypeptides synthesizedin KOS-, 5dl 1.2-, and vd3-infectedVerocells intheabsence(A)and presence (B)of PAA(400 ,ug/ml)at2, 6,and12hpostinfection.Verocellmonolayers wereinfectedwith 10 PFUof theindicatedviruses andincubatedat37°Cin the absence (A)orpresence(B) of400
pg
ofPAA per ml. At theindicated timespostinfection,cellswerepulse-labeledfor30minwith100puCi of[35S]methionineper 35-mmplate. Monolayerswerethenwashed, andcelllysateswerepreparedandanalyzed bySDS-polyacrylamide gel electrophoresisasdescribed in thelegend toFig. 7. Polypeptide designations on therightaredescribedin greaterdetail in thelegend to Fig.7.long-labeling experiments (datanotshown). Hence, the vd3 polypeptide profiles showninFig.7are aconsequenceof the mutationinICP27 codingsequences.
(iv)Kinetics ofviral protein synthesis. Toascertain whether thealterations in gene expression observed in vd3-infected
cells (Fig. 7) are due to the effect of reduced viral DNA synthesisandtoexamine therates of polypeptide synthesis inKOS-,5dl 1.2-, and vd3-infected cells, KOS- and
mutant-infected Vero cellswerepulse-labeled for30minat2, 6,and 12 hpostinfection with [35S]methionine in the presenceand absence of the DNA synthesis inhibitor phosphonoacetic acid(PAA).
The protein profiles shown in Fig. 8A (PAA-) and B (PAA+) demonstrate the following. (i) At 2 hpostinfection, bothvd3 and5dl1.2 failedtoinduce the synthesis of detect-able viralproteins, whereas viral proteins of all classeswere detected inKOS-infected cells (Fig. 8A andB). This obser-vation demonstrates that ICP27 is required to enhance the expression of all viral genes at a very early stage in virus replication. Because both mutantscontain wild-type copies ofICPs0 and 4, the major activators of viral gene expres-sion, the absence of the activating functions of these two proteins at2 h postinfectionimplies thatICP27hasaneffect
on one orboth of these proteins. (ii) At 6 h postinfection, bothvd3and5dl 1.2 induced levels of earlygeneexpression equivalent to those of wild-type virus, whereas -yl gene expression was markedly reduced relative to KOS. These observations indicate that early gene expression, although
slow to beinitiated in mutant-infected cells, reached
wild-typelevelsby6 hpostinfection.This was notthe case for-yl proteins. (iii) At 12hpostinfection, thepolypeptideprofiles of the mutants had changed little from the 6-h profile, whereas in KOS-infected cells, early gene expression was repressed and -yl gene expression was considerably aug-mented. (iv) The mutant profiles were not significantly differentin thepresenceandabsenceofPAA,whereasearly gene expression was repressed more efficiently in the ab-sencethan in thepresenceofPAA,consistent withprevious findings (32). (v)As in thecase oflong-labelingexperiments, themajor difference in thephenotypesof vd3 and5dl1.2 was manifest as aslightly greater repressionofearlygene expres-sionby vd3 than by 5dl 1.2. This differencewasmostnotable at 12h postinfection.
DISCUSSION
ICP27 is a multifunctional regulatory protein that pos-sessesatleasttwodistinctactivities, namely repression and enhancement of ICP0- and ICP4-induced gene expression. Although the mechanism by which ICP27 mediates its regu-latory effects is not clear, the observation that ICP27-stimulated viral geneexpressionoccurs concomitantly with increased rates of the transcriptionstrongly suggests that it exertsits upregulatory effectatthelevel of transcription(16,
32). The ICP27 protein is a nuclear phosphoprotein
com-posedof 512 aminoacids, withanapparentmolecular mass of 63kilodaltons(41). LikeICP4,phosphate has been shown to cycle on andoff the nascent ICP27 protein during
infec-qu:mNMMofiA i omo
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.147.475.74.365.2]tion (57). AlthoughICP27has been reported to bind to DNA (23), it is not known whether this binding is specific or nonspecific or whether it involves interaction with other viral or cellular proteins. The study described herein was designed to begin to relate the specific structural features of
the
ICP27
molecule with its repressing and enhancingactiv-ities.
Functional regions of the ICP27 protein. Although the requirement for thehydrophilic amino portion ofICP27 was not specifically assessed in these studies, Hardwicke et al. have shown that it is not essential for either the repressing or enhancing activity of the protein (22). The importance of the carboxyl portion of the ICP27 molecule to its essential regulatory functions is suggested by the fact that the muta-tions in allICP27 ts mutants mapped to date are located in this region (47) and by the existence of extensive amino acid sequence conservation between this region ofICP27 and the carboxy-terminal halves of the related varicella-zoster virus gene 4 product, the Epstein-Barr virus BMLF1 product, and the IE 52k gene product of herpesvirus saimiri (7, 20; D. J. McGeoch, V. G. Preston, S. K. Weller, and P. A.
Schaffer,
in S. J. O'Brien, ed., Genetic Maps, in press).In an effort to identify regions of theICP27 primary amino acid sequence required for its repressing and enhancing functions, a series of nonsense and in-frame deletion muta-tions were introduced into the ICP27 coding sequence.
Because
they can be used to delete defined stretches ofamino acids, these two types of mutation are especially useful in assessing the activities specified by individual regions of a protein and in identifying functional domains prior to more detailed analysis by amino acid deletion and substitution mutagenesis.
(i) Repression. The results of transient expression assays with mutant plasmids demonstrated that two regions of the
ICP27 primary amino acid sequence specify repressing ac-tivity for immediate-early, early, and late promoters: an 80-amino-acid region between residues 327 and 407 and a 46-amino-acid region between residues 465 and 511. The 80-amino-acid region is not absolutely required for the repressing activity ofICP27, but it is necessary for wild-type levels of repression, since the pd3 peptide which lacks this
region
was able to repressICPO-or ICP4-induced expression from the early tk promoter significantly.The pnll peptide but not the pnlO peptide efficiently represses ICPO- or ICP4-induced expression of immediate-early, immediate-early, and late promoters. This suggests that the 46-amino-acid region between residues 465 (pnlO) and 511 (pnll) also contributes to the total repressing activity of
ICP27. Amino acid sequence analysis has shown that the 46-residue region contains a putative zinc finger motif as well as four serine and two threonine residues which may serve as phosphorylation sites. Similar motifs are present in other transcription proteins.
Lying between the 80- and 46-amino-acid repressing re-gions ofICP27is a 58-amino-acid sequence (407 to 465)that is not requiredforrepressing activity but is able to interfere with the repression specified by the 80-residue sequence. Analysis of the primary and secondary structure ofICP27 reveals that the 58-amino-acid region between amino acids 407 (pn8) and 465 (pnlO)is the most hydrophobic and hence the most rigid portion of the molecule. It ispossible that the additional amino acid residues specified by pn9 and pnlObut not by pn8 in the absence of the remainingcarboxyterminus (amino acids 465 to 512) may cause the pn9 and pnlO peptides to assume a conformation that masksthe activity of the 80-amino-acid repressing region. As in the case of the
mutant peptide specified by pdl (Fig. 4, right-hand panel),
mutations in the 58-amino-acid region would likely yield
peptides with greater repressing activity than thewild-type protein. Such mutants might exhibit a trans-dominant phe-notype (i.e., the ability to inhibit viral growth) even in the presence of wild-type ICP27.
(ii) Enhancement and complementation. NoneoftheICP27 mutant peptides tested in these studies was able toenhance ICPO- or ICP4-induced pgBCAT expression. Thus, specific amino acid sequences lying between residues 258 and 407 (pd3), 375 and 407 (pd2), 407 and 460 (pdl), and 460and 512 (pn9) were shown to be required for pgBCAT enhancing activity. Of special interest was the observation that pnll peptide, which contains 511 of the 512 residues in ICP27 (and hence contains the sequences deleted in mutant peptides specified by pdl, pd2, and pd3), was also unable to enhance gB promoter activity or to complement anICP27 null mutant efficiently. These observations have led us toconclude that unlike its repressing function, which appears to be specified by at least two discrete domains of the ICP27 protein, the enhancing and complementingfunctions ofICP27 involve a larger portion of the carboxy half of the ICP27 molecule. Consequently, enhancing activity would be particularly sen-sitive to mutations thatproduce alterations in the conforma-tion of the protein.
Mutant peptides were tested in this study fortheir ability to modulate ICP0- and ICP4-induced gene expression from five viral promoters representing genes of the four major kinetic classes. A comparison of the relative repressing
activities of mutant peptides for the tk, ICP4, VP5, and L42 promoters induced by ICPO revealed a strikingly similar pattern (compare Fig. 1A and Fig. 2B, C, and D). These relative activities were also evident in the repression of ptkCAT activity induced by ICP4. Together, these observa-tions suggest a common mechanism underlying the repres-sion of both ICP0- and ICP4-induced promoter activity by
ICP27. They further suggest that the mechanism of repres-sion is the same for all fourclasses of promoter.
The results of the transient expression assays conducted in this study confirm and extend the results ofother inves-tigators. Thus, the 80-amino-acid region able to repress
ICPO- or ICP4-induced expression from four classes of
promoter identified in this study is a subset of the 143-residue repressing region (263 to 406)described byRiceetal. for the early ICP8 promoter (46). Similarly, the 46-amino-acid repressing region shown for the same four classes of promoter in this study is asubsetofthe 78-residuerepressing
region (434 to 512) described by Hardwicke et al. for the early tkpromoter (22).Together, these three studies suggest that repression ofICPO- orICP4-induced geneexpression is specified by at least two regions of theICP27 molecule.
Our studies agree well with those of Hardwicke et al. concerning the region of the protein required for enhance-ment ofICPO- or ICP4-induced geneexpression in thatmost
ofall of the carboxyl half of the protein is required for this activity (22). Theseresults differ from those of Riceet
al.,
in which a peptide specifying only the amino terminal 263 amino acids exhibited partial enhancing activity (46). The reasons for this discrepancy are currently unclear.Complexities oftransient expression assays. Investigatorsin severallaboratories haveclearly demonstrated thatICP27 is capable of both enhancing and

![FIG. 8.ofelectrophoresisabsenceFig.,ug/ml) [35S]methionine Viral polypeptides synthesized in KOS-, 5dl 1.2-, and vd3-infected Vero cells in the absence (A) and presence (B) of PAA (400 at 2, 6, and 12 h postinfection](https://thumb-us.123doks.com/thumbv2/123dok_us/1320394.85678/10.612.147.475.74.365/ofelectrophoresisabsencefig-methionine-polypeptides-synthesized-infected-absence-presence-postinfection.webp)
