Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Functional Analysis of the CD4
⫹
T-Cell Response to
Epstein-Barr Virus: T-Cell-Mediated Activation of Resting B Cells and
Induction of Viral BZLF1 Expression
ZHENG FU†ANDMARTIN J. CANNON*
Department of Microbiology and Immunology, University of Arkansas for Medical Sciences, Little Rock, Arkansas 72205
Received 18 January 2000/Accepted 26 April 2000
In contrast to the major role played by Epstein-Barr virus (EBV)-specific CD8ⴙcytotoxic T-cell responses
in immunosurveillance, recent studies have offered the apparently paradoxical suggestion that development of EBV-driven human B-cell lymphoproliferative disorders and tumors in SCID/hu mice is dependent on the presence of T cells, in particular CD4ⴙT cells. This study presents a functional analysis of the CD4ⴙT-cell
response to EBV and shows that while CD4ⴙT cells may be cytotoxic, they also express Th2 cytokines and
CD40 ligand (gp39) and possess B-cell helper function. We show that EBV-specific CD4ⴙT cells can provide
non-HLA-restricted help for activation of resting B cells via a gp39-CD40-dependent pathway and are able to induce expression of BZLF1, a viral lytic cycle transactivator in latently infected resting B cells, ultimately resulting in rapid outgrowth of transformed B-cell colonies. These results support the proposal that CD4ⴙT
cells may play a key role in reactivation of latent EBV infection and may thus contribute to the pathogenesis of EBV-driven lymphoproliferative disorders.
There is a general consensus that HLA class I-restricted CD8⫹cytotoxic T-cell responses play a major role in the con-trol of latent Epstein-Barr virus (EBV) infection, by virtue of their ability to recognize viral antigens associated with B-cell transformation (9, 19). While EBV-specific CD8⫹T-cell re-sponses have thus been widely studied, little is known of the CD4⫹T-cell response to EBV. Virus-specific CD4⫹ T cells that lyse in vitro-transformed lymphoblastoid cell lines (LCL) have been described (12–14, 17), and it has recently been shown that CD4⫹T cells can induce Fas-mediated apoptosis in LCL (23), suggesting a possible role for CD4⫹T cells in EBV-specific immunosurveillance.
In contrast to the acknowledged importance of EBV-specific cytotoxic T-cell responses in maintenance of asymptomatic latent infections in healthy individuals, recent studies have presented the apparently paradoxical finding that EBV-driven tumorigenesis in SCID/hu mice is dependent on the presence of T cells (5, 22). The work of Veronese and colleagues showed that both CD4⫹and CD8⫹T cells were capable of fulfilling the helper function necessary for the development of EBV-in-duced tumors in SCID/hu mice, with CD4⫹T cells being the more efficient (22). These observations support the proposal that helper CD4⫹T cells can play an important role in the pathogenesis of EBV-associated disease, through non-HLA-restricted bystander activation of resting B cells and resultant reactivation of latent EBV infection. In this report, we show that EBV-specific CD4⫹T-cell function in a healthy individual covers a broad spectrum, encompassing cytotoxic responses and expression of both Th1 and Th2 cytokines. In particular, we demonstrate that EBV-specific CD4⫹T cells can provide
highly efficient help for activation of resting B cells, and we further show that CD4⫹-T-cell-mediated activation of latently EBV-infected peripheral blood resting B cells through a CD40-dependent pathway induces expression of BZLF1, an immediate-early transactivator of viral lytic cycle replication.
Derivation and characterization of polyclonal EBV-specific CD4ⴙT cells.CD4⫹T cells were purified from the peripheral blood lymphocytes (PBL) of a healthy, EBV-seropositive, adult donor (donor 1) by positive selection with anti-CD4-coupled magnetic beads (Dynabeads; Dynal A.S., Lake Suc-cess, N.Y.). EBV-specific CD4⫹T cells were selected by co-culture with autologous, irradiated (7,500 cGy) LCL (grown in the presence of 10% human AB serum, rather than fetal calf serum, to avoid stimulation of a CD4⫹ T-cell response to bovine proteins) and maintained as a polyclonal line by peri-odic restimulation with LCL in RPMI 1640 plus 10% human AB serum, supplemented with 50 U of recombinant human interleukin 2 (IL-2) (provided by the Biological Response Modifiers Program, National Cancer Institute, Bethesda, Md.) per ml. Donor 1 CD4⫹T cells strongly proliferated in response to stimulation with autologous donor 1 LCL but failed to respond to allogeneic LCL from an unrelated HLA class II-mismatched donor (donor 2). Furthermore, donor 1 CD4⫹T cells failed to recognize autologous, normal B lymphoblasts activated with anti-CD40 and IL-4 according to the protocol of Banchereau et al. (1). Collectively, these results confirmed that the donor 1 CD4⫹T-cell response was EBV specific. Donor 1 CD4⫹T cells were cytotoxic to autologous LCL (21% specific lysis at an effector/target ratio of 20:1) but showed minimal cytotoxicity to autologous anti-CD40-activated normal B lym-phoblasts, allogeneic LCL, or NK-sensitive K562 cells (data not shown).
Intracellular cytokine expression by EBV-specific CD4ⴙT
cells.Two-color flow cytometric analysis allows determination of cytokine expression at the single-cell level (15). CD4⫹ T cells (7.5⫻105/ml) were activated by incubation at 37°C for 6 h
in RPMI 1640–10% human AB serum plus 50 ng of phorbol myristyl acetate (PMA) per ml and 500 ng of ionomycin per ml. * Corresponding author. Mailing address: Department of
Microbi-ology and ImmunMicrobi-ology, Mail Slot 511, University of Arkansas for Medical Sciences, 4301 W. Markham, Little Rock, AR 72205. Phone: (501) 296-1254. Fax: (501) 686-5359. E-mail: cannonmartin@exchange .uams.edu.
† Present address: Department of Microbiology and Immunology, University of Michigan Medical School, Ann Arbor, MI 48109.
6675
on November 9, 2019 by guest
http://jvi.asm.org/
Brefeldin A at 10g/ml was added for the final 3 h of incu-bation. Control, nonactivated cultures were incubated in the presence of brefeldin A only. The cells were harvested, washed, and fixed with 2% paraformaldehyde in phosphate-buffered saline (PBS) for 20 min at room temperature. For intracellular staining, the cells were washed and permeabilized by incubation in PBS plus 1% bovine serum albumin (BSA) and 0.5% saponin (S-7900; Sigma, St. Louis, Mo.) for 10 min at room temperature. Monoclonal antibodies (MAb) used were fluorescein isothiocyanate (FITC)-conjugated anti-gamma in-terferon (anti-IFN-␥), FITC–anti-IL-2, FITC–anti-tumor ne-crosis factor alpha (FITC–anti-TNF-␣), phycoerythrin (PE)-conjugated anti-IL-4, and PE–anti-IL-13 (all from Becton Dickinson, San Jose, Calif.). After staining for 30 min at room temperature, cells were washed twice with PBS plus 1% BSA and 0.5% saponin, washed once with PBS plus 0.5% BSA, and fixed a second time with 2% paraformaldehyde in PBS.
Coordinate analysis of IFN-␥and IL-4 expression revealed three distinct but overlapping subsets (Fig. 1A); 35% of CD4⫹ T cells were IFN-␥hiIL-4lo, akin to classically described Th1
cells, and 12% were IFN-␥loIL-4hi, typical of Th2 cells.
How-ever, the major population (49%) was of an intermediate, double-positive IFN-␥hiIL-4hiphenotype. Broadly similar
pat-terns were revealed by coordinate analysis of IL-4 expression versus TNF-␣and IL-2 expression, with a double-positive pop-ulation being the predominant phenotype in each case (Fig. 1C and E). We also recorded the distribution of CD4⫹T cells that expressed IL-13, a Th2 cytokine that has many of the func-tional characteristics of IL-4, including the ability to induce proliferation of B cells activated by CD40 engagement (4, 7). Two-color analysis of IL-13 and IFN-␥expression by CD4⫹T cells again suggested three distinct functional subsets (Fig. 1B), as seen with the IFN-␥–IL-4 analysis, but in this instance only 22% of T cells possessed an IFN-␥hiIL-13loTh1 phenotype,
whereas the double-positive IFN-␥hiIL-13hipopulation
con-stituted 62% of the total. Overall, 75 to 80% of CD4⫹T cells were IL-13 expressers, and a majority of these cells also ex-pressed TNF-␣and IL-2 (Fig. 1D and F).
CD40 ligand (gp39) expression by EBV-specific CD4ⴙ T
cells.T-cell-mediated activation of resting B cells requires T cell expression of gp39 and subsequent engagement of B cell CD40. Coordinate expression of cell surface gp39 and intra-cellular IL-4 or IFN-␥was measured by flow cytometry, using the activation protocol described above for intracellular cyto-kine assays. In this case, activated and control (nonactivated) T cells were stained with FITC–anti-gp39 MAb or an isotype-matched FITC-conjugated control MAb (both from Ancell Corp., Bayport, Minn.) following PMA and ionomycin activa-tion but prior to the fixaactiva-tion and permeabilizaactiva-tion steps nec-essary for detection of intracellular cytokine expression. Anti-cytokine antibodies used were IL-4 and PE–anti-IFN-␥(both from Becton Dickinson). EBV-specific CD4⫹T cells expressed gp39 in an activation-dependent manner (Fig. 2). The gp39⫹IL-4⫹subset had a mean fluorescence intensity for IL-4 expression of 37.8, whereas the gp39⫺IL-4⫹subset had an IL-4 mean fluorescence intensity of only 20.9, suggest-ing that gp39 expression may coincide with high IL-4 expres-sion (Fig. 2A). A subset (30%) of IFN-␥-expressing CD4⫹T cells also expressed gp39 (Fig. 2B).
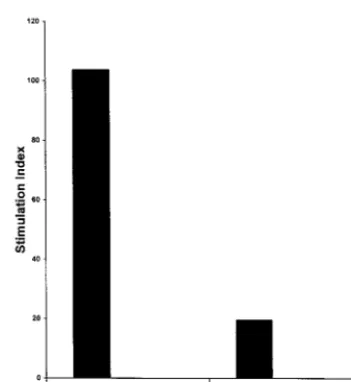
Activation of resting B cells by EBV-specific CD4ⴙT cells. The high frequency of IL-4 and IL-13-expressing EBV-specific CD4⫹T cells, coupled with activation-dependent gp39 expres-sion, suggested that these T cells were likely to provide efficient help for activation of resting B cells. Purified resting B cells from the donor 1 or 2 were prepared by positive selection from PBL using anti-CD19 Dynabeads (Dynal A.S.). B cells were of 95 to 98% purity, as determined by flow cytometric analysis, and residual T-cell contamination was less than 1%. EBV-specific CD4⫹ T cells (5 ⫻ 104/well) were irradiated (3,000 FIG. 1. Two-color flow cytometric analysis of intracellular expression of IL-4
versus IFN-␥, IL-2, and TNF-␣, and intracellular expression of IL-13 versus IFN-␥, IL-2, and TNF-␣by EBV-specific donor 1 CD4⫹T cells. T cells were
stimulated for 6 h with PMA and ionomycin, with brefeldin A added for the final 3 h. In all cases, control, unstimulated T cells failed to stain for intracellular cytokine expression.
FIG. 2. Two-color flow cytometric analysis of CD40 ligand (gp39) expression versus intracellular expression of IL-4 and IFN-␥. T cells were stimulated for 6 h with PMA and ionomycin, with brefeldin A added for the final 3 h.
on November 9, 2019 by guest
http://jvi.asm.org/
rads) and cocultured with B cells (2 ⫻ 104/well) in 96-well
flat-bottom plates (Falcon; Becton Dickinson, Lincoln Park, N.J.) precoated with anti-CD3 MAb (50g of OKT3 per ml in PBS, overnight at 4°C). Where appropriate, blocking anti-gp39 MAb was used at 50g/ml. B-cell activation and proliferation were measured by [3H]thymidine ([3H]TdR) incorporation
af-ter 4 days. The results illustrated in Fig. 3 represent one of two experiments conducted with EBV-specific CD4⫹T cells from donor 1. The same results were also seen for B-cell activation experiments with EBV-specific CD4⫹T cells from two other donors. Coculture of T cells with resting B cells in the presence of plate-bound anti-CD3 MAb showed that EBV-specific CD4⫹T cells were capable of providing efficient help for B-cell activation and proliferation (Fig. 3). The presence of anti-gp39 blocking MAb markedly reduced the B-cell stimulation index (significant at aPvalue of⬍0.01, Wilcoxon two-sample test), indicating that non-HLA-restricted T-cell-mediated B-cell ac-tivation was strongly dependent on gp39-CD40 interaction. B-cell activation was also dependent on helper CD4⫹-T-cell activation, as coculture of T and B cells in the absence of
plate-bound anti-CD3 MAb failed to induce B-cell activation (not shown).
Does CD4ⴙ-T-cell-mediated activation of latently infected
resting B cells result in reactivation of the EBV lytic cycle?
Purified resting B cells (5⫻106/well) prepared following
leu-kapheresis of donor 1 were cultured with EBV-specific donor 1 CD4⫹ T cells (5 ⫻ 106/well) for 48 h in 8 ml of RPMI
1640–10% human AB serum per well in six-well plates (Costar Corp., Cambridge, Mass.) precoated with anti-CD3 MAb (50
g of OKT3 per ml in PBS, overnight at 4°C). The cells were then spun down, and the RNA was extracted with TRIzol (1 ml/well; Gibco BRL, Grand Island, N.Y.) and subsequently prepared as described previously (16). T-B coculturing was conducted in the presence or absence of blocking anti-gp39 MAb. Purified B cells were also incubated with anti-CD40 plus IL-4 as a positive control for B-cell activation (1). As the frequency of resting B cells latently infected with EBV is in the range of only 1 to 100 per 106 B cells, we utilized highly
sensitive reverse transcription (RT)-PCR assays capable of detecting very rare viral mRNA species. Nested sets of primers were used for amplification of BZLF1 and EBNA2 sequences from cDNA, as described by Prang et al. (16) and Chen et al. (3), respectively. The outer BZLF1 primer sequences were 5⬘-ATTGCACCTTGCCGCCACCTTTG-3⬘and 5⬘-CGGCAT TTTCTGGAAGCCACCCGA-3⬘, while the nested BZLF1 primer sequences were 5⬘-GACCAAGCTACCAGAGTCTA T-3⬘and 5⬘-CAGAATCGCATTCCTCCAGCGA-3⬘. The outer EBNA2 primer sequences were 5⬘-ACTTTGAGCCACCCACA GTAACCA-3⬘and 5⬘ -TGGAGTGTCTGACAGTTGTTCCTG-3⬘. The nested EBNA2 primer sequences were 5⬘-CATAGAAG AAGAAGAGGATGAAGA-3⬘ and 5⬘-GTAGGGATTCGAG GGAATTACTGA-3⬘. Viral mRNA transcripts are spliced, and each primer pair utilizes sequences from different exons. This strategy enables us to distinguish between PCR products ampli-fied from cDNA and PCR products ampliampli-fied from possible con-taminating genomic DNA, which would include intron sequences (3, 16) and thus give rise to significantly larger PCR products (Table 1). For qualitative and quantitative control of RNA and cDNA preparations, we used RT-PCR for histone 3.3 (a house-keeping gene), as described previously (16).
[image:3.612.84.260.71.262.2]Nested RT-PCR analysis of RNA harvested after 48 h showed that BZLF1 mRNA expression was readily detectable in B cells cocultured with activated CD4⫹ T cells, whereas BZLF1 expression could not be detected in B cells cultured alone (Fig. 4A). The fidelity of the BZLF1 RT-PCR product from the T-B coculture was confirmed by TA cloning (Invitro-gen, San Diego, Calif.) and subsequent DNA sequencing (data not shown). CD4⫹-T-cell-induced expression of BZLF1 was completely blocked in the presence of 50g of anti-gp39 MAb per ml. While blockade of gp39-CD40 interaction inhibited
FIG. 3. Bystander activation of resting B cells by EBV-specific donor 1 CD4⫹
T cells. Resting B cells were purified from peripheral blood by positive selection with anti-CD19-coupled magnetic beads and then cultured for 4 days with irra-diated CD4⫹T cells in the presence of plate-bound anti-CD3, in the presence or
absence of blocking anti-gp39 MAb (50g/ml). Stimulation indices were calcu-lated as the ratio of [3H]TdR uptake by B cells cultured with T cells divided by
[3H]TdR uptake by B cells cultured alone. Background counts in control wells of
irradiated T cells were subtracted prior to calculation of stimulation indices for B-cell proliferation.
TABLE 1. PCR conditionsa
Primer (mmol/liter)Mg2⫹ Annealingtemp (°C) No. ofcycles
Size (bp) of: cDNA
product DNA productGenomic
BZLF1 outer 2 59 40 608 732
BZLF1 inner 2 60 45 442 566
EBNA2 outer 2 61 45 338 702
EBNA2 inner 2 58 45 271b 635
aDenaturation times and temperatures: 20 s at 94°C for the BZLF1 primers and 15 s at 95°C for the EBNA2 primers. Annealing times: 40 s for the BZLF1 primers and 30 s for the EBNA2 primers. Extension times and temperatures: 40 s at 72°C for the BZLF1 primers and 30 s at 72°C for the EBNA2 primers.
bBased on the EBNA2 cDNA sequence for EBV B95-8, published by Sample et al. (21). Using the same primers, Chen et al. have predicted a PCR product of 267 bp (3).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.52.552.614.695.2]T-cell-mediated induction of viral lytic cycle activation in la-tently infected B cells, we also found that BZLF1 expression could be strongly induced by the combination of an agonist anti-CD40 MAb (Immunotech, Westbrook, Maine) and re-combinant human IL-4 (Fig. 4A). EBNA2 mRNA expression was not detected by nested RT-PCR in either resting B cells cultured alone or CD4⫹-T-cell-activated B cells (data not shown). RT-PCR for histone 3.3 mRNA sequences gave ho-mogeneous band intensities for all samples, indicating that variations in the levels of BZLF1 expression could not be attributed to variations in RNA or cDNA sample sizes (Fig. 4B).
Activation of resting B cells with irradiated, anti-CD3-acti-vated CD4⫹T cells also resulted in rapid outgrowth of EBV-transformed B cells, i.e., “spontaneous” LCL, within 21 days (data not shown). As B-cell activation initially induces BZLF1 expression rather than transformation-associated transcripts such as EBNA2 mRNA, this observation supports the two-step model of EBV transformation (18, 24), involving viral replica-tion and secondary infecreplica-tion rather than direct outgrowth of activated B cells.
Reactivation of latent EBV and outgrowth of transformed LCL have long been held to be spontaneous events (18, 24). However, more recent studies with the SCID/hu mouse model
of spontaneous EBV-associated lymphoproliferative disorders have revealed a T-cell requirement for tumorigenesis. SCID/ hu mice reconstituted with T-cell-depleted PBL (5, 22) or in which T-cell activation was inhibited with cyclosporine (2) failed to develop EBV-associated human B-cell tumors. Col-lectively, these observations strongly suggest that spontaneous reactivation does not take place in vivo and that T-cell inter-action with latently infected B cells is a requirement for viral reactivation. As the work of Veronese and colleagues showed that CD4⫹T cells were highly effective at restoring the gen-eration of EBV-driven human B-cell tumors (22), we con-ducted a functional analysis of interactions between CD4⫹T cells and B cells. Although we show that EBV-specific CD4⫹T cells can be cytotoxic to autologous LCL (and potentially con-tribute to EBV-specific immunosurveillance), we also found that they can express Th2 cytokines, such as IL-4 and IL-13, and that they are efficient mediators of bystander activation of resting B cells. Furthermore, we found that helper CD4⫹ -T-cell-mediated B-cell activation induces expression of viral BZLF1, an immediate-early transactivator of EBV lytic cycle replication. Our subsequent finding that CD4⫹-T-cell activa-tion of latently infected resting B cells leads to rapid establish-ment of LCL transformed by endogenous virus supports the proposal that T cells play a potentially important role in viral reactivation and tumorigenesis in vivo.
EBV-specific helper CD4⫹T-cell responses to productively infected or transformed B cells may result in bystander activa-tion of other latently infected resting B cells, which would lead to further viral reactivation and progression of disease. How-ever, T-cell interaction with latently infected resting B cells, and consequent viral reactivation, is non-HLA restricted and noncognate, i.e., there is not a requirement for EBV-specific antigen recognition. In the SCID/hu mouse, activation of CD4⫹ helper T cells of any specificity (e.g., through graft-versus-host responses), and concomitant bystander activation of latently infected B cells, is likely to be sufficient for viral reactivation and subsequent tumorigenesis. Conversely, non-specific blockade of T-cell activation would cut off the required help for B-cell activation and thus prevent tumorigenesis, as is the case in SCID/hu mice treated with cyclosporine (2).
CD4⫹-T-cell-mediated B-cell activation and consequent ex-pression of BZLF1 could be blocked with a MAb specific for T-cell-expressed gp39 (CD40 ligand). We also show that direct engagement of B-cell CD40 with an agonist anti-CD40 MAb (in conjunction with IL-4 treatment) results in strong induction of BZLF1 expression. CD40 early signaling events involve pro-tein kinase C (8), the activation of which can contribute to viral lytic cycle induction (6). Engagement of CD40 may thus play a direct role in BZLF1 transactivation. However, induction of BZLF1 expression may also be a downstream event following T-cell-mediated B-cell activation. Further investigation will be needed to distinguish between these possible pathways.
In conclusion, we show that CD4⫹-T-cell-mediated activa-tion of latently infected resting B cells leads to expression of BZLF1, a key immediate-early lytic cycle transactivator. This event is followed by rapid in vitro outgrowth of transformed B cells. Coupled with earlier reports demonstrating a require-ment for T-cell function in EBV-driven tumorigenesis in the SCID/hu mouse model (2, 5, 22), these results point to an important role for T cells in the pathogenesis of EBV-associ-ated lymphoproliferative disorders. Interestingly, it has re-cently been shown that transplant patients have elevated levels in serum of IL-4 (10, 11), a cytokine which promotes develop-ment of Th2 CD4⫹T-cell responses with strong helper func-tion for B-cell activafunc-tion and proliferafunc-tion, i.e., the type of T-cell response which would favor EBV reactivation from
la-FIG. 4. Induction of BZLF1 mRNA expression in resting B cells following CD4⫹-T-cell-mediated activation. Purified resting B cells from the peripheral
blood of donor 1 were cultured alone or with EBV-specific donor 1 CD4⫹T cells
(in the presence of plate-bound anti-CD3) or were activated with anti-CD40 MAb plus IL-4. Total RNA was extracted after 48 h, followed by nested RT-PCR for BZLF1 (A) and RT-PCR for histone 3.3 (B). RNA harvested from B95-8 cells was used as a positive control for BZLF1 expression. Nested RT-PCR for BZLF1 yielded a 442-bp product from the inner primers (Table 1), and RT-PCR for histone 3.3 gave a 215-bp product.
on November 9, 2019 by guest
http://jvi.asm.org/
tency. A Th2 bias in CD4⫹T-cell responses may thus contrib-ute to an increased viral load, which is known to be a risk factor for development of posttransplant lymphoproliferative disor-ders (20).
We thank Randy Noelle (Dartmouth Medical School, Lebanon, N.H.) for kindly providing the anti-gp39 MAb and Rosemary Roch-ford (University of Michigan, Ann Arbor) for many helpful discus-sions.
This work was supported by Public Health Service grant CA 63931 from the National Cancer Institute.
REFERENCES
1.Banchereau, J., P. de Paoli, A. Valle, E. Garcia, and F. Rousset.1991. Long-term human B cell lines dependent on interleukin-4 and antibody to CD40. Science251:70–72.
2.Boyle, T. J., R. E. Coles, A. M. Kizilbash, and H. K. Lyerly.1992. Effects of cyclosporine on human B-cell lymphoma development in vivo. Surg. Oncol.
1:79–86.
3.Chen, F., J.-Z. Zou, L. di Renzo, G. Winberg, L.-F. Hu, E. Klein, G. Klein, and I. Ernberg.1995. A subpopulation of normal B cells latently infected with Epstein-Barr virus resembles Burkitt lymphoma cells in expressing EBNA-1 but not EBNA-2 or LMP1. J. Virol.69:3752–3758.
4.Cocks, B. G., R. de Waal Malefyt, J.-P. Galizzi, J. E. de Vries, and G. Aversa.
1993. IL-13 induces proliferation and differentiation of human B cells acti-vated by the CD40 ligand. Int. Immunol.5:657–663.
5.Coles, R. E., T. J. Boyle, J. M. DiMaio, K. R. Berend, D. F. Via, and H. K. Lyerly.1994. T cells or active Epstein-Barr virus infection in the develop-ment of lymphoproliferative disease in human B cell-injected severe com-bined immunodeficient mice. Ann. Surg. Oncol.1:405–410.
6.Davies, A. H., R. J. Grand, F. J. Evans, and A. B. Rickinson.1991. Induction of Epstein-Barr virus lytic cycle by tumor-promoting and non-tumor-pro-moting phorbol esters requires active protein kinase C. J. Virol.65:6838– 6844.
7.De Vries, J. E.1996. Molecular and biological characteristics of interleukin-13. Chem. Immunol.63:204–218.
8.Faris, M., F. Gaskin, J. T. Parsons, and S. M. Fu.1994. CD40 signaling pathway: anti-CD40 monoclonal antibody induces rapid dephosphorylation and phosphorylation of tyrosine-phosphorylated proteins including protein tyrosine kinase Lyn, Fyn, and Syk and the appearance of a 28-kD tyrosine phosphorylated protein. J. Exp. Med.179:1923–1931.
9.Khanna, R., S. R. Burrows, and D. J. Moss.1995. Immune regulation in Epstein-Barr virus-associated diseases. Microbiol. Rev.59:387–405. 10. Martinez, O. M., J. C. Villanueva, L. Lawrence-Miyasaki, M. B. Quinn, K.
Cox, and S. M. Krams.1995. Viral and immunologic aspects of EBV infec-tion in pediatric liver transplant patients. Transplantainfec-tion59:519–524. 11. Mathur, A., D. M. Kamat, A. H. Filipovich, M. Steinbuch, and R. S. Shapiro.
1994. Immunoregulatory abnormalities in patients with EBV-associated B
cell lymphoproliferative disorders. Transplantation57:1042–1045. 12. Misko, I. S., D. J. Moss, and J. H. Pope.1980. HLA antigen-related
restric-tion of T lymphocyte cytotoxicity to Epstein-Barr virus. Proc. Natl. Acad. Sci. USA77:4247–4250.
13. Misko, I. S., J. H. Pope, R. Hutter, T. D. Soszynski, and R. G. Kane.1984. HLA-DR-antigen-associated restriction of EBV-specific cytotoxic T-cell col-onies. Int. J. Cancer33:239–243.
14. Misko, I. S., T. B. Sculley, C. Schmidt, D. J. Moss, T. Soszynski, and K. Burman.1991. Composite response of naive T cells to stimulation with the autologous lymphoblastoid cell line is mediated by CD4⫹cytotoxic T cell
clones and includes an Epstein-Barr virus-specific component. Cell. Immu-nol.132:295–307.
15. Nazaruk, R. A., R. Rochford, M. V. Hobbs, and M. J. Cannon.1998. Func-tional diversity of the CD8⫹T cell response to Epstein-Barr virus (EBV):
implications for the pathogenesis of EBV-associated lymphoproliferative disorders. Blood91:3875–3883.
16. Prang, N. S., M. W. Hornef, M. Jager, H. J. Wagner, H. Wolf, and F. M. Schwarzmann.1997. Lytic replication of Epstein-Barr virus in the peripheral blood: analysis of viral gene expression in B lymphocytes during infectious mononucleosis and in the normal carrier state. Blood89:1665–1677. 17. Rencher, S. D., K. S. Slobod, F. S. Smith, and J. L. Hurwitz.1994. Activity
of transplanted CD8⫹versus CD4⫹cytotoxic T cells against Epstein-Barr
virus-immortalized B cell tumors in SCID mice. Transplantation58:629–633. 18. Rickinson, A. B., M. Rowe, I. J. Hart, Q. Y. Yao, L. E. Henderson, H. Rabin, and M. A. Epstein.1984. T-cell-mediated regression of “spontaneous” and of Epstein-Barr virus-induced B-cell transformation in vitro: studies with cy-closporin A. Cell. Immunol.87:646–658.
19. Rickinson, A. B., S. P. Lee, and N. M. Steven.1996. Cytotoxic T lymphocyte responses to Epstein-Barr virus. Curr. Opin. Immunol.8:492–497. 20. Riddler, S. A., M. C. Breinig, and J. L. McKnight.1994. Increased levels of
circulating Epstein-Barr virus (EBV)-infected lymphocytes and decreased EBV nuclear antigen antibody responses are associated with the develop-ment of posttransplant lymphoproliferative disease in solid-organ transplant recipients. Blood84:972–984.
21. Sample, J., M. Hummel, D. Braun, M. Birkenbach, and E. Kieff.1986. Nucleotide sequence of mRNAs encoding Epstein-Barr virus nuclear pro-teins: a probable transcription initiation site. Proc. Natl. Acad. Sci. USA
83:5096–5100.
22. Veronese, M. L., A. Veronesi, E. D’Andrea, A. Del Mistro, S. Indraccolo, M. R. Mazza, M. Mion, R. Zamarchi, C. Menin, M. Panozzo, A. Amadori, and L. Chieco-Bianchi.1992. Lymphoproliferative disease in human periph-eral blood mononuclear cell-injected SCID mice. I. T lymphocyte require-ment for B cell tumor generation. J. Exp. Med.176:1763–1767.
23. Wilson, A. D., I. Redchenko, N. A. Williams, and A. J. Morgan.1998. CD4⫹
T cells inhibit growth of Epstein-Barr virus-transformed B cells through CD95-CD95 ligand-mediated apoptosis. Int. Immunol.10:1149–1157. 24.Yao, Q. Y., H. Czarnecka, and A. B. Rickinson.1991. Spontaneous outgrowth
of Epstein-Barr virus-positive B-cell lines from circulating human B cells of different buoyant densities. Int. J. Cancer48:253–257.