JOURNAL OFVIROLOGY,June 1973,p.848-855
Copyrighti 1973 American Society for Microbiology Printed inVol. 11, No. 6U.SA.
Replication
of
Bacteriophage
qX174 DNA
in
a
Temperature-Sensitive dnaE
Mutant
of
Escherichia
coli C
LAWRENCE B. DUMAS AND CHRISTINE A. MILLER
Department of Biological Sciences, and Program in Biochemistry and MolecularBiology, Northwestern University, Evanston, Illinois60201
Received forPublication 17January1973
Bacteriophage 4X174 cannot grow in a temperature-sensitive dnaE (DNA polymerase III) mutantofEscherichia coliC atthe nonpermissivetemperature.
The inability to grow is the result of inhibition of virus DNA synthesis. The
synthesis of the parental replicative form is unaffected, but the replicationofthe replicative form and the synthesis, of the single-stranded virus DNA are
inhibited.
Several DNA synthesis and DNA polymerase mutants ofEscherichia coli have recently been isolated. Mutants defective in genes polA and polB, which code for DNApolymerasesI andII, respectively, are ableto replicate (2, 7). Thus, neither of these polymerases is essential for bacterial chromosome replication. Conditional
lethal mutants in other loci have been isolated and mapped (19). Mutantsat oneoftheseloci, dnaE, synthesizedefective DNApolymeraseIII molecules (6, 14). This enzyme is required for replication of the bacterial chromosome.
Inexaminingthe rolesoftheproductsofthese DNA synthesis loci in DNA replication, it is
desirable to follow the replication of a small DNA molecule whose replicting intermediates
can be isolated intact. The bacteriophage
4X174
DNA molecule anditsreplicatingforms can easily be isolated and characterized. This virus carries a limited amountofgeneticinfor-mation.Itcodesfor noknown DNApolymerase. Since it must use the E. coli DNA synthesis
system to replicate its DNA, it should be a
useful probeinthis examination.
DNA polymerases I and II are not essential
for
OX174
DNA replication (2). We haveiso-lated a temperature-sensitive dnaE mutant of
E. coli C, the normal 4X174 host strain. We report herethe effects ofthis mutation on the replicationofthe DNA of this virus in vivo.
MATERIALS AND METHODS
Bacteria and phage strains. LD301 (uvrA-,
thyA-, endI-, dnaE")is anitrosoguanidine-induced,
temperature-sensitivemutantofH502 (uvrA -, thyA -,
endIl) isolated in our laboratory. The temperature-sensitivelocus cotransduces with tonA at a frequency of 60%,thus identifying this marker as dnaE (19). The mutant bacteria grow well at 30 C but cannot form visible coloniesonagarmedium at41C. The details of the isolation of this mutant and its characterization will be published elsewhere.
Strain C is the standard host for4X174usedhereto prepare all phage stocks. HF4714 is an amber sup-pressor 4X174 host strain. OX174am3 is an amber mutant in gene E (lysis defective) and is the phage used in all experiments described in this report.
E.coliC, HF4714, and H502, and phage
OX174am3
wereobtained from R. L. Sinsheimer.
Media and buffers. TPGA medium is TPG me-dium (17) with1.0gofKH2PO4and10g ofCasamino Acids per liter. Starvation buffer has been described
(3).
Growth and purification of4X174. Nonradioac-tivephagewerepreparedaspreviouslydescribed(4). 32P-labeled phage were grown as described by Sin-sheimer et al. (17), and purified as described by
Francke and Ray (5). The specific activity ofthe purified phagewasapproximately 10-4countpermin perplaque-forming unit.
Celllysis and Pronasedigestionof the lysates.
32P-phage infected cell samples were collected by
centrifugation, washed threetimes in 0.05 Msodium tetraborate,0.006MEDTA,pH9,toeluteradioactive phage particles still attachedtothe cellsurface(12),
andfinallyresuspendedin 0.5mlofthesamebuffer. Lysozyme was addedto 0.2 mg/ml, and the suspen-sion wasincubatedat0Cforatleast1h.Incubation wascontinuedforseveralmorehoursat0Cafterthe addition of 1% Sarkosyl. After addition of 2 mg of
self-digested Pronase per ml the suspension was
incubatedfor 8h at 37 C. 848
on November 10, 2019 by guest
http://jvi.asm.org/
4X174DNAREPLICATIONINdnaEMUTANT
Cell samples from cultures infected with nonradi-oactivephagewerewashed andlysedin0.05MTris, 0.003MEDTA, pH 8.1. All other conditionswereas
described above.
Sucrose density gradient analysis. Lysed, Pro-nase-digested samples of phage-infected cells were
layeredonto38-ml linear gradientsof 5to20%sucrose
in 0.05 M potassium phosphate, 2mM EDTA, 1 M NaCl, pH7.These gradientswerecentrifugedfor 16h at25,000rpmat4C inaSpinco SW27rotor.Fractions of approximately 0.7 ml were collected from the bottoms of the tubes byusingtheperforatordescribed by Noll (13) (MICO, P.O. Box1652,Evanston, Ill.). Portions of each fractionweredilutedwithwaterand mixed with 3 ml of scintillationfluid in 1-dramglass shell vials. Theradioactivity ineach vialwas deter-mined inaliquid scintillationspectrometer. 5H radi-oactivitywascorrected for32poverlap.The scintilla-tion fluid consisted of 4 g of 2,5-diphenyloxazole
(PPO) perliter in toluene-Triton X-100(2:1). Measurement of acid-insoluble radioactivity. Portions(0.1 ml) of cell culturesweretransferredinto 1ml ofcold 5% trichloroacetic acid. A200-,ggamount ofhighly polymerized calf thymus DNAwasaddedas carrier. TheprecipitateswerecollectedonWhatman GF/A glass fiber filters andwashed three times with cold 5% trichloroacetic acid and finallywith metha-nol. The air-dried filters were placed in 3 ml of scintillation fluid (4 gof PPO per liter intoluene). Radioactivity was measured in aliquid scintillation
spectrometer.
Measurement of intracellular plaque-forming units.Samples (0.1 ml) of the phage-infected cultures were transferred into 1.2 ml oflysis buffer (0.4 M sodium tetraborate, 8 mM EDTA, 0.8% Brij 58, 1.7 mg/mllysozyme, pH 9). The mixtureswereincubated overnightat0C. Afterfreezing and thawing twice, the lysates werediluted with 0.05 M sodium tetraborate andassayedby theagaroverlay technique using host strainHF4714asindicator.
Chemicals. Thymidine-methyl-3H, 10 to 25 Ci/
mmol, and thymine-methyl-3H, 12.5 Ci/mmol, were purchased from Intemational Chemical and Nuclear Corp. and Amersham/Searle. 32P-phosphoric acid, carrier-free, was purchased from New England
Nu-clearCorp. Mitomycin C, chloramphenicol,eggwhite lysozyme, and Pronase (protease type VI) were pur-chased fromSigma Chemical Co.
RESULTS
OX174
growthat 41 C.Totestwhether hostmutant LD301 wouldsupport
OX174
infection,aliquid culture of bacteria grown at 30 C was
infected with *X174am3. Portions of the in-fected culturewere incubatedat30C and41C. Figure 1 shows that progeny phage are not
synthesizedat41C inthis host. Mature phage
are synthesized at 30C. After 3 h of infection
theyield isapproximately 50 PFU percell.
Phage synthesis at 30 C in the parent host strainH502occurs ata morerapidrate than in
mutant LD301, and the total yield after3 h of
a
1fI
- - _8.0
0
0
7.0-0
6.0 _ _
0
2
3
TIME
(hours)
FIG. 1. Growth of bX174am3 in mutant host LD301at30and41C. The bacteriaweregrownat30 Ctoacell densityof approximately2 x10' cellsper
ml on TPGA medium supplemented with 10 ug of thymineperml. The cells werecollected and resus-pended in0.1volumeof starvationbuffer. 4X174am3 wasaddedtoamultiplicity of3.After15min without aeration, 1 volume of growth medium was added. Portions of the culturewereaeratedat30C and41 C. At 30and 60min after aeration was resumed (zero time), portions of the 30 C culturewereshiftedto41 C. At theindicated times0.1-ml portions were with-drawn into lysis buffer. Total PFUwere then deter-mined. Symbols: 0, PFU per ml from a culture incubated at 30 C; 0, PFUper mlfrom a culture incubated at 41 C; 0, PFUper ml from a culture shifted to 41 C at 30 min; A, PFUper ml from a
cultureshiftedto41 Cat60min. Arrows indicate the times of shifts from30to41 C.
infection is several-fold higher. At 41 C phage synthesis is faster than at 30 C (unpublished observations).
Figure 1 also shows that temperature shifts from30to41 Cat30and 60 min after infection result in a slowly decreasing rate of phage synthesis, allsynthesis havingterminated by30 to60min afterthe shift.
Theseresultsindicate the dnaEgeneproduct
VOL. 11, 1973
849
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.257.446.51.376.2]DUMASAND MILLER
ofthehost bacteriumis required for the
synthe-sisof progeny phage.
4X174DNA
synthesis
at41C. The tempera-ture-sensitive mutation in the host bacteriumLD301 specifically affects host DNA synthesis.
The effect of this mutation on kX174 DNA
synthesis was examinedin phage-infected cells
by measuring the incorporation of 3H-thymine intophage DNAat 30and 41 C.
Bacteria grown at 30 C were treated with
mitomycin C prior to infection to specifically
inhibit host DNA synthesis (11). Infected cells were then incubated at the two temperatures.
Figure 2 shows that the level of radioactive
DNA synthesizedat 41C in phage-infected cells is no higher than that in uninfected cells
in-cubatedat30C and41 C. No detectablephage
DNA synthesis occurs.
When 30 C cultures of infected cells were
shiftedto 41 C at30and60min afterinfection, thephage DNA synthesisrateslowly decreased, reachingzeroby30 to 40minafterthe shift. The termination of net phage DNA synthesis was
followedbyaperiodofapparent degradationof
phage DNA. Under similar experimental
condi-tions, cultures ofuninfected mutant cells, not
treated with mitomycin C, cease net DNA
synthesis30 to 40minafterashift from30 to 41
C (unpublished data).
These data indicate that the product ofthe host dnaE gene is required for 4X174 DNA synthesis.
Parentalreplicative formsynthesisat 41C. The replication of
OX174
DNA occurs inthree distinct stages (16). In stage one the single-stranded DNA carriedbythe virus is convertedto the double-stranded parental replicative
form(RF)inthe hostcellby polymerizationof a
complementary strandonthe virus strand
tem-plate. During stage twothese double-stranded
replicative forms serve as template for the
synthesisofapproximately20 progenyRF mole-cules. In stage three theprogeny RFmolecules
serve as template for the asymmetricsynthesis
of many copies of the single-stranded virus DNA. Wehave examined the effect of thednaE mutation in host strainLD301 oneach of these
threestages of
kX174
DNA replication.To test the affect of this mutationonparental
RF synthesis, mutant host cell cultures grown at 30 C and preincubated at either30 or 41 C
were infectedwith 32P-labeled
4X174am3.
The conversion ofthe single-strandedvirus DNAtoRF moleculeswasexaminedbyzone
sedimenta-tion oflysates ofthecultures insucrose
gradi-ents. In high ionic strength buffer the
single-stranded DNA and the two double-stranded forms, RFI andRFII (closed, circularsupercoils
and open, nicked circles, respectively),
sedi-ment at rates of 23s,
21s,
and 16s, respectively(1).
Figure3A showsthe results of such an analy-sis of a lysate from mitomycin C-treated cells harvested 15 min after infection at 30 C. The
32P-labelis seen ina single-stranded DNA band centered about fraction 23, an RFI band
cen-tered about fraction 32, and an RFH band centered about fraction 36. 3H-labeled RFI and RFII bands are also present, representing progeny RF molecules synthesized during the
o
0)
I0 x
%-2-
3
TIME
(hours)
FIG. 2. Radioactive thymine uptake into phage
DNA in kX174am3 infected LD301. Bacteria were grown at 30 C in 10 ml of TPGA medium
supple-mented with 5 ugof thyminepermltoacelldensityof approximately 3 x 108 cells per ml. The cells were collected, washed in starvation buffer, and
resus-pendedin1 mlofthesame.MitomycinCwasadded to 0.1 mg/ml.After20min in the dark at 30C, the cells were collected and resuspended in 2 ml of
starvation buffer. Phagewereaddedto 1.3mlofthis suspensionat amultiplicityof3.After15minat30C withoutaeration,3mloffreshTPGA medium
supple-mented with 1 Mg ofthymineper ml and15MCi of 3H-thymine were added to 0.3-ml portions of the
uninfectedandinfectedcultures(zerotime).Portions of eachwereincubatedat30C and41 C. One eachof
theinfected culturesincubatedat30Cwasshiftedto 41 Cafter 30 min and 60 minofaeration at 30 C. Acid-insoluble radioactivity was measured in 0.1-ml
portions. Symbols: 0, 30 C, infected; 0, 30 C, uninfected;A,41C, infected;A,41 C, uninfected; 0,
shiftedto41 Cat 30min; *,shiftedto41 Cat60min. Arrowsindicate the timesofshiftsfrom30 to 41 C.
850 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.263.450.204.448.2]V1X174 DNAREPLICATION IN dnaE MUTANT
N
'-0
x
%..
A
6
4
2^ to
I0
X
a-N 6cm
4)
2
I
10 20 30 40 50 10 20 30 40 50
[image:4.493.54.447.46.310.2]FRACTION NUMBER
FIG. 3. Zone sedimentationof parental phage DNVAfrominfectedLD301. Cultures (50ml) ofbacteriawere grown and treated withmitomycin Casdescribed inFig. 2.Portions (25 ml) ofthetreated cells in TPGA medium supplemented with 1 ggof thymineper ml wereaeratedfor 15 min at 30 C and 41 C (A and B, respectively) priortoinfection.Inaseparateexperiment25-mlportionswereincubatedfor30minat 30C and 41C (C andD,respectively) priortoinfection.32P-OX174am3wereaddedtoeach cultureat amultiplicity of10. Thespecific activity ofthe phagewas approximately 1 x 10-4 32pcountper min perPFU in A and B and
approximately5 x 10-532p countper min perPFUin C and D.3H-thymidine (0.1mCi)wasaddedatthetime
ofinfection.Each culturewasincubatedfor5minwithoutaeration,15min withaeration,andrapidlychilled. The infected cells were collected, washed, lysed, anddigested with Pronase. The lysates weresedimented through linear sucrosegradientsasdescribed inMaterials and Methods. Totalradioactivitywasdetermined in 0.2-mlportions ofeachfractiondiluted with 0.1 mlofwater.Symbols: ,'H;__, Sp.
15-mininfection period.The "P-labeled
single-stranded DNA band isdue, at least in part,to
DNA released during the lysis and Pronase
treatments from phage particles still attached
tothe cell surface. The relative size of thisband decreases with the number of borate-EDTA washes prior tolysis.
A comparison of Fig. 3Bto 3A shows that a 15-min preincubation of the mutant host
cul-ture at 41 C prior to infection followed by a 15-min infection period at 41 C caused no
reduction in the amount of 32P-labeled phage
DNAconvertedtoRFrelativetothatat 30C. In fact, there is 2.3-fold more 3"P-DNA in the RF
bands. In contrast, the amount of 3H-labeled
progeny RF made at 41 C is only 40% that at 30
C.
Similarly, preincubation ofthe mutant host culture at 41 C for 30 min prior to infection causes no reduction in the conversion of
pa-rental virus DNAlabel toRFat 41 C (compare
Fig. 3D to 3C). The amount of
'H-labeled
progenyRF madeat 41 C inthisexperiment isonly30%that at 30C.
We have extended thepreincubationsat41C
to 1 and 2 h and find no decrease in the
efficiency of conversion of parental DNA to
parental RF at 41 C comparedtothat at30C. The 3H label in RF DNA from these 41 C cultures is barely detectable and represents at most5%ofthat from the30C cultures (unpub-lished data).
These results show that the product of the host dnaEgeneisnotrequired for the synthesis
of kX174 parental RF.
4X174RF
replication
at41C. The resultsofthe experiments described in Fig. 3 indicate that kX174 RF replication is inhibited during the first15minof infectionat 41C. To examine
this inhibition in moredetail, we pulse-labeled
$X174-infected LD301 cultures at 15 and 30 min after ashiftfrom 30 to 41 C andcompared the rates of RF DNA synthesis at the two temperatures. The cultures were treated with
mitomycin Cprior to infection and infected in
thepresence of 30
,g
ofchloramphenicolperml.This concentration of chloramphenicol allows
RFreplicationtoproceed forseveral hoursand
VOL.11,1973 851
on November 10, 2019 by guest
http://jvi.asm.org/
DUMASAND MILLER
prevents single-stranded virus DNA synthesis
(17).
Lysates ofthese pulse-labeled cultures were
then sedimented through sucrose gradients. Figure4Ashows thesedimentationprofile of the
lysate of the 30 C culture, and Fig. 4B shows
that of the41C culture, pulse-labeledat 15 min
after the shift. The radioactive label inthe RF
region ingradient B isonly about 17%ofthatin
gradient A.
In a separate experiment a portion of a
similarly infected culture was incubated for30 minafterthe shiftto 41C before pulse-labeling. Figure 4D shows that theamountof 3Hlabel in RF DNA from the 41 C culture is again only about one-fifth that from the30C culture (Fig. 4C).
When4X174-infectedcultures ofLD301were
pulse-labeled during the period ofRF
replica-tion immediately after the shift to 41 C, we
observed 2.4-fold more pulse-label inRF DNA
6 A
4
0
K 6
fromlysatesof41Ccultures thanfromidentical
cultures at 30 C (datanotshown). The rate of RFreplicationat 15and30minaftertheshift is therefore approximately 7 to 8% of the initial
rate at 41C.
The results of these experiments show that
theproductofthe host dnaEgene isrequiredfor thereplicationofkX174RFDNA.
Single-stranded phage DNA synthesis at 41 C. A culture of LD301 was infected with
kX174am3
andincubatedat30 C for 45 min.By
this time single-stranded virus DNA is being synthesized andencapsulated in mature phage particles (see Fig. 1). Label incorporated into RFmolecules during short pulsesatthisstageis found predominantly in the viral strand and chases into virusparticles (10).
Half ofthe culture wasthen shiftedto 41 C. At 10,20,and30 minafter theshift, portionsof
the 30 C and 41 C cultures were pulse-labeled with3H-thymidinefor 1min.The labeledphage
10
20
30
40
50
10
20
30
40
50FRACTION NUMBER
FIG. 4. Zone sedimentationofphageDNAfrom X174-infectedLD301pulse-labeled duringRFreplication.
Bacteria cultures were grown and treated with mitomycin C as described in Fig. 2. Treated cells were resuspended in TPGA mediumsupplementedwithI .tgofthymine and30,ugofchloramphenicolper ml.Phage
wereaddedat a multiplicity of3. The cultures were incubated at 30 C. At20 minafteradditionofphage,
one-half of the culture was shifted to 41 C. After 15 min 10 ml ofthe 30 C and the 41 C cultures were
pulse-labeledfor 1 min with125MCi of8H-thymidine(AandB,respectively).Inaseparateexperiment,similar 10-mlcultures of30 C and41 Cwerepulse-labeledfor1 minat30minafter theshiftto41 C. The labeled cultureswererapidlychilledbytheadditionofanequalvolumeofacetonechilled inamethanol-dryice bath. The cellswerecollected,washed,lysed,anddigestedwith Pronase.TheIysatesweresedimentedthroughlinear sucrose gradients as described in Materials and Methods. Arrows indicate the peak positions of added
'2P-labeled4X174virus DNA marker. Totalradioactivity wasdetermined in0.4-mlportionsofeachfraction
dilutedto 1ml with water.
852 J.- VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
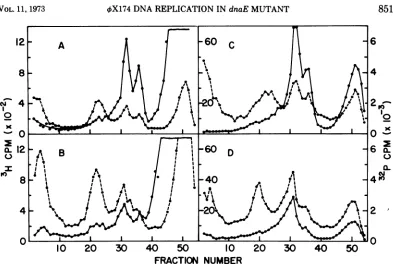
[image:5.493.56.449.290.546.2]VX174DNAREPLICATION IN dnaEMUTANT
DNA from these cultures was thenexaminedby
zone sedimentation.
Figures 5A and 5B show the sedimentation profiles of the DNApulse-labeledat 30Cand41
C, respectively, at 10minafterthetemperature shift. Figures 5C and 5D represent similar analyses of portions of the two cultures
pulse-labeled at 20 min after the shift, and Fig. 5E and 5F at 40 min. Inallofthegradientsthe RFII band is centerednearfraction 30, and the free
single-stranded virus DNA band position, marked by added 32P-DNA, is indicatedby the
arrow. Phage DNA not liberated from phage
particles during the Pronase digestion is found on the dense cushion at the bottom of each gradient.
The amount ofradioactive label incorporated into the phage DNA species during the 30 C
pulses at 10, 20, and 40 min afterthe shift is
approximately the same. The rate of phage DNAsynthesis at 30 C during this time period is therefore constant. The rate at 41 C, however,
decreases. Comparison ofFig. 5B to 5A shows that at 10 min after the shift phage DNA synthesis is occurring at a 3.2-fold greater rate
at 41 C than at 30 C. Ten minutes later the rate at41 C has decreased to 54% of that at 30 C. at
40minafterthe shift the41Crate isonly 26% of that at30C. The rate of virus DNA synthesis at
20 and 40 min after the shift is, therefore, approximately 17 and 8%, respectively, of that
at 10min after the shift.
Single-stranded virus DNA and phage parti-cles aresynthesized at 41C, even after 40 min at thistemperature. But therateofsynthesis has been reducedtoonly8%of the initialrate at41 C.
These results show that the product of the host dnaE gene is requiredforthe synthesis of single-stranded 4X174 DNA.
DISCUSSION
4X174 RF
replication
occurs on the cellmembrane (9),as does E. coli DNAreplication (18).Thereplicationofthe E. coli chromosome requires DNA polymerase II (6, 14). We have shown here that RF replication is inhibited at the nonpermissive temperature in a dnaE mu-tant. The rate of synthesis initially increases after shifts to 41 C, but is approximately 90% inhibitedby 15 min after the shift. Thus DNA polymerase III is also required for kX174 RF replication.
The late synthesis of4X174single-stranded DNA occurs in the cell cytoplasm (8). This
synthesis is alsoinhibited in the dnaE mutant host at 41C. This reaction is approximately 80%
inhibited after 20 min at the nonpermissive
0
4~~~~~~~
10 20 30 40 10 20 30 40 10 20 30 40
FRACTION
NUMBER
FIG. 5. ZonesedimentationofphageDNAfrom<qX174-infectedLD301pulse-labeledduringsingle-stranded
DNAsynthesis.A60-ml cultureof bacteria inTPGA plus thymine was incubated at 30 C until the cell density reached approximately 5 x 108 cells per ml. The cells were collected, washed in starvation buffer, and resuspended in5mlof the same. Phage were added at a multiplicity of 3, and the suspension was incubated for 15min at 30Cwithout aeration. At zero time, 55 ml of TPGA medium plus 2
,gg
ofthymine per ml were added. After45minof aeration at 30 C, one-half of the culture was shifted to 41 C. At 10, 20, and 40minafter the temperatureshift, 100MCiof3H-thymidine were added to 10 ml portions of the 30 C and 41 C cultures. AfterI mineachsample was rapidly chilled and processed as described in Fig. 4. The gradients in this experiment consisted of34mlof buffered5to20%osucroseover a 3-ml cushion of saturatedCsCIinbuffered 60% sucrose. A, 10minat 30C; B, 10minat 41 C; C, 20 min at 30 C; D, 20min at 41 C; E, 40min at 30 C; F, 40min at 41 C. Note that theordinate in gradient B is different from all of the others.853
VOL.11,1973
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.57.445.377.565.2]DUMASANDMILLER
temperature. We conclude that DNA polym-erase III is also required for the synthesis of
OX174 single-stranded
DNA.The synthesis of 4X174 parental RF is not
inhibited in this dnaE mutant. The conversion of parent phage DNA to the double-stranded replicative forms occurs even after 1 to 2 h of preincubation of the culture at 41 C. Little or no
progeny RF replication occurs after such
pro-longed preincubation. We conclude that DNA
polymerase III is not required for parental RF synthesis in this host.
Others have shown that kX174 can grow in mutants defectiveinDNApolymerasesI and II
(2, 7).Presumably parental RF synthesis occurs in these mutants, indicating that neither of thesepolymerasesisrequiredin cellsthat have normal DNApolymerase Ill activity.
Further evidence concerning the possible roles of the three known DNA polymerases in parental RFsynthesis comesfrom experiments usingcell-freeextracts.Extracts ofpolA-polB+ dnaEtsE.coliaredefectiveinthe conversion of added4X174virus DNAtothedouble-stranded
form (15, 20). Extracts ofpolA- polB+ dnaE+ cells are notdefective. Thus, DNA polymerase
III isrequired for the in vitro reaction inextracts
defective in DNA polymerase I activity.
Ex-tractsofpolA- polB- dnaE+ bacteriacan
cata-lyze the in vitro reaction (20). Thus, DNA polymerase II activity is neither required nor
sufficient.
Apossible explanationforthe requirement of DNApolymerase III activity for the conversion of 4X174 virus DNA to a double-stranded molecule in these cell extracts, and no such requirementforparentalRFsynthesisin vivo in LD301, isthat both DNApolymerases I and III might act as catalysts for these reactions. The absence of only one of these two polymerase
activities would not prevent the conversion of virus DNAtodouble-strandedDNA.Sincehost LD301 is defective in DNA polymerase III, but
not DNA polymerase I, parental RF synthesis
occurs. The cell extracts, defective in both
polymerase activities, do notcatalyze the
con-version. This explanation can betestedin vivo in a polAl- dnaEt5 double mutant. Ifcorrect,
parentalRFsynthesisshouldnot occur at41 C.
Iftheinvitro conversion of4X174DNAto a
double-stranded molecule is different from the synthesisofparentalRF invivo,then we cannot
excludeDNApolymeraseIIfromapossiblerole in vivo. We can only say that some DNA
polymerase other than III can catalyze the in vivo reaction in LD301. DNA polymerase III cannot be excluded from participating in
nor-mal hosts. The DNA polymerase catalyzing
parentalRFsynthesisinLD301could be polym-eraseI or II, or anew host cellenzyme.
ACKNOWLEDGMENTS
This work was supported in part by Public Health Service research grantnumber 9882 from the National Institute of Allergy and Infectious Diseases, research grant number GB22867 from theNational Science Foundation, and research grant number 70-21 from the American Cancer Society, ilinois Division, Inc.
LITERATURECITED
1. Burton, A., and R. L. Sinsheimer. 1965. The process of infection with bacteriophage4X174.VII. Ultracentrif-ugal analysis of the replicative form. J. Mol. Biol. 14:327-347.
2. Campbell, J. L., L. Soll, and C. C. Richardson. 1972. Isolation and partial characterization of a mutant of Escherichia colideficient in DNA polymerase II. Proc. Nat. Acad. Sci. U.S.A. 69:2090-2094.
3. Denhardt, D. T., and R. L. Sinsheimer. 1965. The process of infection with bacteriophage 4X174. m. Phage maturationand lysis after synchronized infection. J. Mol. Biol. 12:641-646.
4. Dumas,L.B., G. Darby, and R. L. Sinsheimer. 1971. The replication of bacteriophage OX174 DNA in vitro. Temperature effects on repair synthesis and displace-mentsynthesis.Biochim.Biophys. Acta 228:407-422. 5. Francke, B.,and D.S. Ray.1971.Fate of parental4X174
DNA upon infection of starvedthymine-requiring cells. Virology 44:168-187.
6. Gefter,M.L., Y. Hirota,T. Kornberg,J. A. Wechsler, and C.Bamoux. 1971.Analysis of DNA polymerases II and III in mutants ofEscherichia coli thermosensitive for DNA synthesis. Proc. Nat. Acad. Sci. U.S.A. 68:3150-3153.
7. Hirota,Y., M.Gefter,and L. Mindich. 1972. A mutant of Escherichia coli defective in DNA polymerase II ac-tivity. Proc. Nat. Acad.Sci.U.S.A. 69:3238-3242. 8. Knippers, R.,A.Razin,R.Davis,and R. L.Sinsheimer.
1969. The process of infection with bacteriophage
4OX174. XXIX. In vivo studiesonthesynthesisofthe single-strandedDNA of progenyfX174bacteriophage. J. Mol. Biol. 45:237-263.
9. Knippers,R., andR. L. Sinsheimer. 1968. The process of infectionwithbacteriophageOX174.XX. Attachment of the parental DNA of bacteriophage 4X174 to a
fast-sedimenting cell component. J. Mol. Biol. 34:17-29.
10. Komano, T.,R. Knippers,and R. L.Sinsheimer. 1968. The process of infection with bacteriophage 4X174. XXII. Synthesis of progeny single-stranded DNA. Proc. Nat. Acad.Sci. U.S.A.59:911-916.
11. Lindquist,B.H.,and R. L.Sinsheimer.1967.The process of infection withbacteriophageOX174.XV. Bacterio-phageDNAsynthesisinabortive infections with a set
ofconditional lethalmutants.J.Mol. Biol. 30:69-80.
12. Newbold,J.E.,and R. L.Sinsheimer. 1969. The process of infection withbacteriophageOX174.XXXII. Early steps in the infection process:attachment,eclipse,and DNApenetration. J. Mol. Biol. 49:49-66.
13. Noll, H. 1969. Polysomes: analysis of structure and function, p.101-179.InJ.Sargent,and P. N.Campbell (ed.),Techniquesinproteinbiosynthesis, vol.2.
Aca-demicPressInc., London.
14. Nusslein, V., B. Otto, F. Bonhoeffer, and H. Schaller. 1971. Function of DNApolymerase IllinDNA
replica-tion.Nature(London) 234:285-286.
15. Schekman, R.,W.Wickner,0.Westergaard,D.Brutlag, K. Geider, L. L. Bertsch, and A. Kornberg. 1972.
Initiation ofDNA synthesis: synthesis ofOX174
rep-854 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
cX174DNAREPLICATION IN dnaE MUTANT licative form requires RNA synthesis resistant to
rifampicin. Proc.Nat. Acad. Sci. U.S.A. 69:2691-2695. 16. Sinsheimer, R. L., R. Knippers, and T. Komano. 1968.
Stages in the replication of bacteriophage *X174 in vivo. Cold Spring Harbor Symp. Quant. Biol. 33:443-447.
17. Sinsheimer, R. L., B. Starman, C. Nagler, and S. Guthrie. 1962. Theprocessofinfection with bacterio-phageOX174.I. Evidence forareplicative form. J. Mol.
Biol. 4:142-160.
18. Smith, D. W., and P. C. Hanawalt. 1967. Properties of the growing point region of the bacterial chromosome. Biochim. Biophys. Acta 149:519-531.
19. Wechler, J. A., and J. D. Gross. 1971. Escherichia coli
mutants temperature-sensitive for DNA synthesis. Mol.Gen. Genet. 113:273-284.
20. Wickner, R. B., M. Wright, S. Wickner, and J. Hurwitz. 1972.Conversion ofOX174and fd single-stranded DNA
to replicative forms in extractsof Escherichia coli. Proc. Nat. Acad. Sci. U.S.A. 69:3233-3237.
VOL.11,1973 855
on November 10, 2019 by guest
http://jvi.asm.org/