Copyright( 1972 American Society forMicrobiology Printed in U.S.A.
Plasmid-Controlled
Variation in
the Content of
Methylated Bases
in
Bacteriophage
Lambda
Deoxyribonucleic Acid
STANLEY HATTMAN
Departmentof Biology, University of Rochester, Rochester, New York 14627
Received for publication 12 June 1972
TheN6-methyladenine (MeAde) and 5-methylcytosine (MeG) contentsin deoxy-ribonucleic acid (DNA) ofbacteriophage lambda has beenanalyzed as afunction of host specificity. Thefollowing factshaveemerged: (i) lambdagrown onstrains
harboring the PI prophage contain ca. 70 more MeAde residues/DNA molecule
thanlambda grown either in thePl-sensitive parent, or in aP1 immune-defective
lysogenwhichdoesnotconfer PI modification; (ii)lambda grownonstrains
harbor-ing the N-3 drug-resistance factor containca.60 moreMeCresidues/DNAmolecule than lambdagrown on the parental strainlackingthefactor; (iii) lambdagrown in
Escherichia coliBstrains is devoid ofMeC, whereas lambda grownin a B (N-3)host
contains a high level of MeC; (iv) the MeAde content in lambda DNA is not affectedby the N-3 factor. These results suggestthatPl controlsanadenine-specific DNA methylase, and that the N-3 plasmid controls a cytosine-specific DNA
methylase. The N-3 factor has been observed previously todirect cytosine-specific
methylationofphageP22 DNAand E. coli B DNA invivo;invitrostudiespresented
here demonstrate thisactivity.
Previous results from our laboratory (18, 19)
showed that the N-3 drug-resistance factor in-creased thelevelofmethylcytosine (MeG) in the
deoxyribonucleic acid (DNA) ofphage P22 and
hosts Salmonella typhimurium and Escherichia
colistrainB;E.coliB isnormallydevoidofMeC
(11, 12, 13, 15, 19, 25). In contrast, presence of
the plasmid appeared not to exert any influence
onthe level ofmethyladenine (MeAde)inDNA. These observations suggested that specific
meth-ylation of cytosine residues may be responsible
for the host-controlled modification determined
by theN-3 plasmid. Sincebacteriophage lambda
isalso susceptible tothisrestriction-modification
system (hs II; 7, 8) it was reasonable to expect
thatweshouldobserve similar effects of the
plas-mid onthecontentofmethylatedbases inlambda DNA.
The present communication presents in vivo and in vitro data to support this prediction. In
addition, we describe a specific increase in the
MeAde content oflambda DNA controlled by
P1 prophage; thisplasmid is alsoresponsiblefor
restriction-modificationofDNA
(1, 22).
MATERIALS AND METHODS
Phageandbacterial strains.PhagesXc1857
in&
and XcI857 ind- sus S7 were generously provided by B.Dottin;XcandXvir werefrom S. E. Luria. The bac-terial strains used in this study are summarized in Table 1.
Media and chemicals. Growth media were as
described earlier (17-19). [2-3H] adenine (Schwarz/ Mann), S-adenosyl-L-[methyl-'HJmethionine (SAM) and L-[methyl-3H]-methionine (Amersham/Searle), N6-methyladenine and S-adenosyl-L-methionine-iodide (repurified in our lab) (Calbiochem), 5-methyl-cytosine (Mann), calf thymus DNA (Worthington)
wereobtained from the firmsindicatedin parentheses. SSC consisted of 0.15 M NaCl plus 0.015 M sodium citrate, pH 7.0. Sonication buffer was 0.02 M potassium phosphate, pH 7.2, plus 0.005 M Na2 ethylenedi-aminetetraacetic acid (EDTA) plus 0.05 M NaCl. Dialysis buffer was sonically treated buffer plus 0.001 M
2mercaptoethanol plus 5% glycerol (v/v).
Preparation of labeled XcI857 ind DNA. Bacterial strains lysogenic for XcI857 ind- were grown to ca.
3 X 108/ml at 32 C in minimal medium. The
cul-tures were transferred to 42 C and incubated for
15 min with aeration, after which time they were placedat 37 C (t = 0). After 10 min of incubation, 5.0-ml sampleswereplaced in flasks (at 37 C) contain-ingthe following: (i) 0.2mlof amixture of adenine, thymidine,and uridine (1 mg of each/ml) plus 0.01 ml ofmethionine (1 mg/ml) plus 0.10 ml of [methyl-3H]methionine (1 mCi/ml), or (ii) 0.05 ml of adenine (0.1 mg/ml) plus0.05 ml of[2-3H]adenine (0.5 mCi/ ml). Thecultureswere incubated with vigorous aera-356
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Escherichia coli strains and relevant properties
Parental straina Host-specificityphenotypeb Source and remarks
Bc251 rB+mB+ J. W.Wiberg (2)
B834 met-gal- rB-mB- H. Revel (33) derived from Bc251
1100 rgl-su+ B1-endol- rK-mK+ H. Revel
C600 rK+mK+ R. Yuan
C600 (P1) rK+mK+rpl+mpl+ R. Yuan
C600 (PIdef) rK+mK+rpF-mpl- Ourlab from C600 (P1)
aAbovestrains servedashostsforlysogenizationwith Xc1857 ind or
P1,
and forreceipt of the N-3 factor. Forbrevity,all derivatives used in thisstudyare notlistedin theabove table.bTheterminology for host specificity is that proposed earlier (4); rB and mB refer to B-specific restriction and modification, respectively.
tion until spontaneous lysis occurred 60 to 90 min later. The labeledphagewaspurifiedbytwoalternate cycles ofhigh- andlow-speedcentrifugations,nuclease digestion (17), and by centrifugation to equilibrium in a CsCl gradient. The peak fractions were pooled anddialyzedagainst SSC,and the DNAwasextracted
as described earlier (17).
Determination ofMeC/MeAdeand MeAdecontent.
Phage lambda DNA was precipitated overnight in absolute ethanolat-20Cand harvestedby centrifuga-tion; the resulting pellets weredried under a stream
ofair. For determination of the MeC/MeAde ratio, the dried pellet was hydrolyzed in 0.1 ml of 70% perchloric acid and subjected to descending paper chromatography as previously described (19). The MeAdecontentof lambda DNAwasdeterminedafter hydrolysis in 1 N HC1 and paper chromatography
(17). TheMeC, MeAde, and Aderegions wereeach identified (bymeans of theultraviolet absorbance of co-chromatographed authentic markers), cut out, eluted,andcountedasbefore(17).
Preparation of crudeextracts. Cultures of rE-mB-andrB-mB-(N-3)weregrownto3 X 108to5X 108/ml
at 37 C in broth. The cells were harvested at low speed and resuspended in sonication buffer. After sonictreatmentandclarificationatlowspeed, the
ex-tract wascentrifugedfor 30 minat50,000 rev,/minin
an International A-321 rotor. The supernatant fluid
was collected anddialyzed overnight at 4C against dialysisbuffer.
Determination of bases methylated in vitro. The
basic methylase stock mixture (1.0 ml) contained the following: tris(hydroxymethyl)aminomethane (Tris)-hydrochloride, pH 8.0 (200 ,umoles); SAM
(10 nmoles); [mnetll-3H]SAM (ca. 0.5
ACi);
(Na2 EDTA, pH 7.6 (20 umoles); 2-mercaptoethanol (25 jumoles); NaOH to adjust the pH to neutrality. The crudeextract (ca. 1 mg ofprotein) andvarious DNA species (ca. 100to 150,g) wereadded tothe stock mixture (total volume = 2.5 ml) and incubated at 37Cfor2hr(thiswassufficienttosaturatethedifferent substrates, as shown by parallel kinetic analysis of 3H-CH3 label incorporation into acid-insoluble material). Sarkosylwasadded to0.3%(w/v) and the DNA species were extracted by phenol. Hydrolysis and chromatographic analysis ofMeC/MeAde wereperformedasabove.
RESULTS
Effect of N-3 and P1 plasmids on methylation of phage lambda DNA. Bacteriophage lambda is subject to the restriction and modification en-zymescontrolledby the N-3 and P1 plasmids (1, 4, 5, 7, 8, 22, 30, 32). The N-3 factorhas been shown to determine a cytosine-specific DNA
methylation of P22 DNA (19); in contrast, no
effect of P1 on lambda DNA methylation has
beenobserved (15). We investigated the possible influence of the N-3 and P1 plasmids on the in
vivo methylation of bases in lambda DNA.
Various bacterialstrains lysogenic forprophage
Xcl857ind were heat-induced (by thermal
inac-tivation of the repressor); during DNA replica-tion, theceUls were labeledwith either
[2-3H]ade-nine (for measurement of the MeAde/Ade plus
MeAde ratio) or [methyl-3H]methionine (for
measurement of the MeC/MeAde ratio). The
dataobtainedbythefirst method allows
calcula-tion of thenumber ofMeAde per lambda DNA molecule; along with data obtained by thesecond
method,the numberof MeCper DNAmolecule
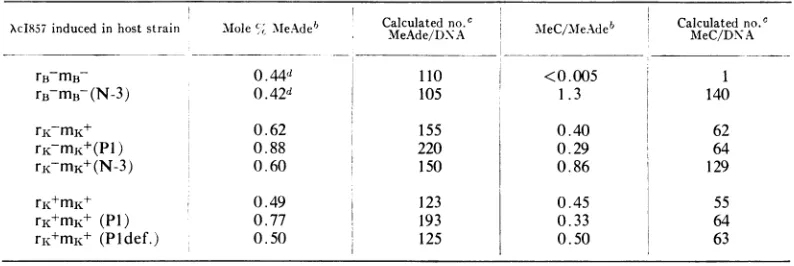
canbecalculated.The results of such analyses are listed in Table 2. The important findings are summarized as follows: (i) lambda DNA con-tains no MeC after growth in rB-mB cells; in contrast, growth of lambda in cellscontainingthe N-3 plasmid leads to extensive methylation of
cytosine, without affecting the level of MeAde;
(ii) in agreement with others (15, 23), lambda DNAcontains MeC aftergrowth in E. coli K12
strains; however, presence ofthe N-3 factor still
leads to a twofold increase in the MeC/MeAde
ratio, whereas the MeAde content remains un-altered. Theseresultssuggest that the N-3 plasmid controls an enzyme which recognizes additional
sequence(s) on lambda DNA not methylated by theK12cytosine-specific enzyme; (iii) presence of theP1 prophage leads to a 40 to50%increase in the level of MeAde, (ca. 70 more MeAde per DNAmolecule) as well as aparallel decrease in 357
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Analysis of methylatedbases in phageXcI857 ind-deoxyriboniucleic acid afterthermalinduiction
in various host strainisa
XcI857 induced in host strain Mole MeAdeb Calculated no. MeC/MeAdeb Calculatedno.
MeAde/DNA Me/eAe eC/DNA
rB-mB- 0.44d 110 <0.005 1
rB mB (N-3) 0.42d 105 1.3 140
rK-mK 0.62 155 0.40 62
rKmK+(P1) 0.88 220 0.29 64
rKmK+(N-3) 0.60 150 0.86 129
rK+mK+ 0.49 123 0.45 55
rK+mK+ (P1) 0.77 193 0.33 64
rK+mK+ (Pldef.) 0.50 125 0.50 63
Procedures described inMaterials and Methods.
bValues presented are themean values obtained from several independent DNA preparations; in
somecases,duplicate analyses wereperformedon agiven preparation. Variationwasgenerallyless than i10% of themeanvalue.
cThenumber ofMeAde/DNA andMeC/DNA wascalculated on thebasis of25% Ade and105total nucleotides/DNA.
dResults ofa single determination.
the MeC/MeAde ratio; (iv) when lambda is propagated in an immune-defective P1-lysogen (immune to P1, butproducesnoactivephageand does not restrict or modify lambda [16]), the
DNA exhibits the normal MeAde level. These
results suggest that the P1-plasmid controls an
adenine-specific DNAmethylase.
Finally, we observed small, but reproducible,
differences in the MeAdecontentoflambda DNA
propagatedin different K12strains;thiswasalso
noted by others (C. Hidalgo andH. A. Nash, J.
manuscriptinpreparation) and mayreflect
strain-specificdifferences in host DNA methylase (14).
In vitro methylationofvarious DNA species by extracts of N-3 containing cells. Reports from
other workers have shown thatextracts ofE.coli
B bacteriaexhibitDNA-methylating activity spe-cific for adenine residues only (13). We have
confirmed thesedata and extended themtostudies
on strain B(N-3). Since the latter strain has been
shown to methylate lambda DNA-cytosine in
vivo (Table 2),weattemptedtodemonstrate such
anactivityin vitro using crudeextractsprepared
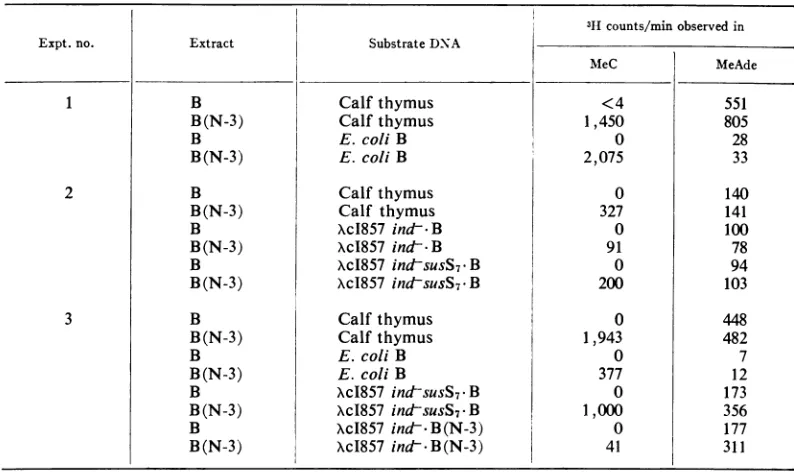
from uninfected cells. The resultsof several such experiments are summarized in Table 3. First, it can be seen that the methylation of adenine residues is similar in both B andB(N-3) extracts for each DNAtested. However, thereare differ-encesin acceptoractivity among the variousDNA species; e.g.,E. coli B DNA was arelatively poor acceptor compared to calf thymus and lambda DNAspecies.
Incontrast, it is clearthat the ability to
meth-ylate cytosine residues is exhibited only by the
B(N-3) extract, and all of the DNAspecies tested in these experiments were substrates for in vitro cytosine methylation. In this respect, morethan
95%cl
of the in vitromethylation
of E. coli B DNA was on cytosine residues; thus, this DNA would be a particularly useful substrate to moni-tor the N-3 methylase during its purification.Finally, when XcI B(N-3) DNA was used as a
substrate in the B(N-3) extract, relatively few
methyl groups were transferred to cytosine; this
is to be expected, since most of the N-3 specific sequences werealready methylatedduringinvivo replication of thelambda DNA.
Weshould comment here on thebehaviorof the N-3restriction and methylation enzymes in crude extracts.Theexperiments summarized inTable 3 wereall carried out inthe absence of added Mg2+
ionand in the presence of EDTA. When EDTA wasomitted and Mg2+ added,we observed little
or no 3H-CH3 incorporation into acid-insoluble
material. This is presumed to result from the
degradation of the acceptor DNAby the
restric-tion nuclease(s). Thus, it would seem that the
restriction nuclease, but notDNA methylase, re-quires Mg2+. Similar ion requirements have been observedfor the P1,K12,andB-specific modifica-tion andrestrictionenzymes(21, 24, 26, 28).
Protection against N-3 restriction conferred by host cytosine-specific DNA methylase. In Sal-monella, N-3 restriction directed against phage P22is a strong one; i.e., ca. 1 in 103 unmodified P22 is capable of producing a plaque on an (N-3)-containing host. Although P22 is a sub-strate in vivo for both N-3 and host cytosine
HATTMAN J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
methylases, there is no indication that either
methylation gives any protection against the
heterologous restriction-nuclease (19). In E.
coli,
N-3 restriction against phage lambda appears to be arelatively weak one (5, 7, 8). However, the degree oflambda restriction may actually be a function of the presence or absence of MeC on
the infecting lambda phage DNA. For example,
Xc
rB-mB-
andXcrB+MB+
(bothlack MeC)are seento bestrongly restricted in (N-3)-containinghosts (Table 4); in contrast,Xc.rK+mK+(contains
MeG) is onlyweaklyrestricted. Asimilarpattern has also been observed by Arber and Morse (5).
Thissituationcanbeinterpretedinthelight ofour
new knowledge concerning the MeC content of
phage lambda DNA; viz. when lambda DNA
contains MeC, added by the E. coli K-12 meth-ylase, it is partially protected against the N-3 restriction nuclease. In contrast, N-3 directed methylation doesnotprotectlambda against the
hs Kor hsB restriction;e.g. Xc-rB_mB-(N-3) is fully restricted in
rK+
andrB+
cells, respectively (Table 4).DISCUSSION
The N-3 plasmid controls the restriction and modification (host-specificity II) of P22 DNA in
S. typhimurium, and ofX, TI, and P1 DNA
TABLE 3. Methylation ofvariousdeoxyribonucleic acid species incrudeextracts ofEscherichia
coli B and B(N-3)a
3Hcounts/minobserved in
Expt. no. Extract Substrate DNA
MeeC MeAde
1 B Calf thymus <4 551
B(N-3) Calfthymus 1,450 805
B E. coliB 0 28
B(N-3) E. coliB 2,075 33
2 B Calf thymus 0 140
B(N-3) Calf thymus 327 141
B XcI857 ind-B 0 100
B(N-3) XcI857 ind-B 91 78
B XcI857 indsusS7*B 0 94
B(N-3) XcI857 indsusS7-B 200 103
3 B Calf thymus 0 448
B(N-3) Calfthymus 1,943 482
B E.coli B 0 7
B(N-3) E. coli B 377 12
B XcI857
ind&susS7-B
0 173B(N-3) Xc1857 ind-SUSS7-B 1,000 356
B XcI857 ind--B(N-3) 0 177
B(N-3) XcI857
indt-
B(N-3) 41 311a Procedures are as described in Materials and Methods. The crude extracts, as well as the
phage
and bacterialDNAspecieswereobtained from therB-mB- andrB-mB-(N-3)
strains.TABLE 4. Efficiency of plating of phage lambda grownand testedoni varioushost strainsa
Host strain Phage
rBmBBBrir+Nr~~~m-(N-3)B r+m+(N-3)B BicKrKmBXiKKr+ m; m (N-3)
Xc-rBmB 1.0 10' 5 X 10' 10-6 2 X 10' 1.0 2 X
10-Xc*rB+mB+ 1.0 1.0 6 X 10' 5 X 10-5 3 X 10' 1.0 2 X 104
Xc-rB-mB (N-3) 0.8 10 1.0 10o- 2 X
104
1.0 1.0).c-rB+mB+(N-3) 1.0 0.9 1.0 1.0 2 X 104 1.0 1.0
Nc-rK+mK+ 1.0 8 X 10-' 0.13 8 X 10-5 1.0 1.0 0.19
Xvir*rK+mK 0.6 1--,' 0.10 104 1.0 1.0 0.05-0.10
aLog-phase cultures
takenas 1.0.
wereused asindicator bacteria. The efficiency of plating onE. coli rK-m,K+ is
359
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.44.441.248.484.2]species in E. coli. We reportedearlier(18, 19)that thepresence of the N-3 specifically increases the MeC content in the DNA species of phage P22 and host cells S. typhimurium and E. coli B. We
concludedthatthe N-3factorcontrols a
cytosine-specific DNA methylase, although there still is
nodirect evidence thatthisactivityisresponsible
forthe host-specificity modification. The present
communication confirms andextends ouroriginal
observations; in addition, wehavedemonstrated
that the Pl-plasmid also affects phage lambda
DNA methylation. The results are summarized
asfollows: (i)invivolabeling with [2-3H]adenine
or [methyl-3H]methionine showed that the N-3
factor specifically increases the MeC content of
the double-stranded DNA in phage lambda, by
ca. 60 residues per DNA molecule (Table 2);
(ii) invitro studies using crudeextractsofE.coli
bacteria whichharborthe N-3factorhas further
demonstrated the cytosine-specific DNA
meth-ylase activity; this activity (missing in E. coli
strainB) does not require Mg2+ ion, whereas it
appears that the N-3 restriction nuclease does;
(iii) the Pl-plasmid specifically increases the
number of MeAde residues in phage lambda
DNA by approximately 70 MeAde/DNA; the
normal MeAdelevel wasobserved when lambda
was propagated on an immune-defective lysogen
in which the P1 prophage was unable toconfer
P1-specific
modification.These resultsstrongly support the notion that
theN-3 (hs II) andP1-directed modificationsare
mediated by DNA-methylating enzymes which
transfermethyl groups
(from
S-adenosylmethio-nine) to specific cytosine and adenine residues,
respectively. Whilethismanuscriptwasin
prepa-ration, direct evidence for a P1-controlled
ade-nine-specific DNA methylase has been reported
(10). Our findings appear to be at variance with
thoseof GoughandLederberg (15), whodid not
observe anydifference in the MeAde content of
lambda growninE. coli strains B andB(P1);we
do not yet understand this discrepancy. On the
other
hand,
weobservelevelsofMeCand MeAdein lambda grown on C600rK+mK+thatare simi-larto theirs(15)andothers (C.Hidalgo and H. A. Nash, manuscript inpreparation).
Accepting that the host-specific modification is mediated by specific DNAmethylation, it is rele-vant to ask whether all the MeC and MeAde residuesproduced by the
plasmid
methylases are essential for protection against the restriction enzymes. In other words, are all the additionalmethylgroups added by theP1 and N-3
methyl-ases locatedatrestriction-modification sites?
P1,
forexample, appears to increase theMeAde
con-tent of lambda by about 70 residues/DNA. If
the host-specificity site possesses twofold
rota-tionalsymmetry (4, 9, 20), thenthere are at least
ca. 35sites per lambda DNA.However, the num-ber ofP1-sites appears to be considerably lower (26), suggesting that the majority of methylated bases added by P1 are not essential for protection against the restriction enzyme. This raises the question of whether the PI-adenine methylase recognizes more than one nucleotide sequence(or there is more than one methylase). In this respect, the single-stranded DNA phages fd and M13 are not subject to restriction by the N-3plasmid, but they are subject to plasmid-specific methylation in vivo (Hattman, manuscript in preparation). Thus, this could mean that the modification enzyme is, in fact, capable ofgratuitously
methy-lating certain nucleotidesequences notrecognized
by the restriction enzyme. Although we cannot rule out the possibility that a (potential) single hs II restriction site is always modified before re-striction, thispossibility does not seem likely, inas-much asfdvariants containing onlya single-site
sensitive to hs B restriction have been isolated
(3,29).
Thus, we consider the most likely explanation of the above observations to be that modification is mediated by a DNA methylase which recog-nizes more than one kind of nucleotide sequence.
This situation, in fact, is not unusual; e.g., the
DNAmethylase specifiedby phageT2 wild-type,
or its uPI-mutants, appears to recognize more than a single sequence of bases (17, 27, 31; R. Hehlmann and S. Hattman, J. Mol. Biol., in
press). Although Arber and his co-workers have
presented data that restriction and modification
enzymesact atthesamesite (6,21,) 29,
nonethe-less,it cannot beprecludedthatthemodification
methylase could still recognize additional
se-quences as well.
It now seems fairly clear that the
restriction-modification enzymes act at specific sites; the
sequence of bases within these sites may or may notpossess twofoldrotational symmetry
perpen-dicular totheDNAhelix axis(4, 9, 20).However the case may be, it is evident that the specific
sequence can be presentonlyinlimited numberon each DNAmolecule. Furthermore,for the mole-cule not to bedegraded,themodificationenzyme
must recognize, at least, all the sites recognized
bytherestriction nuclease. (Itremainsto be seen
whetherthe sequences arecongruent or are two
distinct neighboringregions.) If, within the
modi-ficationsequence, base changes could be tolerated
withoutaffectingitsmethyl acceptor activity (i.e.,
the methylation sequence would be degenerate)
while abolishing nuclease activity, then such a
situationallows for gratuitous methylation of all
similar sites located elsewhere on the DNA. In
other words, due to a relatively less-stringent
on November 10, 2019 by guest
http://jvi.asm.org/
PLASMID-CONTROLLED DNA METHYLATION
recognition pattern, the modification methylase mightactatsitesnotrecognized bythe restriction nuclease (but not vice versa). This question
re-mainsopenfor futureinvestigation.
ACKNOWLEDGMENTS
Theexcellenttechnical assistance of Diane Kuharikisgratefully acknowledged. We thank Bob Dottin for strains andadvice.
This work was supported by Public Health Service grants
AI-08738 and AI-1 0864fromtheNational Institute ofAllergy and Infectious Diseases.
LITERATURE CITED
1. Arber, W., and D.Dussoix. 1962. Host-specificityof DNA
producedbyEscherichia coli. I. Host-controlled
modifica-tion ofbacteriophageX.J Mol. Biol.5:18-36
2 Arber, W.,and C. Lataste-Dorolle. 1961.Erweiterung des
WirtsbereichesdesBakteriophagen X auf Escherichia coli B. Pathol. Microbiol. 24:1012-1018.
3. Arber, W., andU.Kuhnlein. 1967.Mutationeller Verlust
B-spezifischer Restriktion des Bakteriophagen fd. Pathol. Microbiol.30:946-952.
4. Arber, W., and S.Linn. 1969. DNAmodificationand restric-tion.Annu. Rev. Biochem.38:467-500.
5.Arber, W., andM. L. Morse. 1965.Host-specificity of DNA produced by Escherichia coli.VI.Effectsonbacterial
con-jugation.Genetics 51:137-148.
6.Arber, W., A.Rifat, D.Wauter-Willems, andU. Kuhnlein. 1972. Host-specificity of DNA produced by Escherichia coli.XVI. PhageXDNAcarriesasingle site of affinity fcr A-specificrestriction andmodification. Mol. Gen. Genet. 115:195-207.
7 Bannister, D.,and S.W.Glover.1968.Restrictionand Modifi-cationofbacteriophages byR+ strains of Escherichia coli K12. Biochem.Biophys.Res.Commun.30:735-738. 8.Bannister, D., and S. W. Glover. 1970. Theisolation and
propertiesofnon-restrictingmutantsoftwodifferent host
specificitiesassociatedwithdrugresistancefactors.J.Gen.
Microbiol.61:63-71.
9. Boyer, H.W. 1971. DNA restriction andmodification
mecha-nisms in bacteria.Annu. Rev.Microbiol.25:153-176.
10.Brockes, J. P.,P. R.Brown,and K.Murray. 1972.The deoxy-ribonucleicacid modificationenzymeofbacteriophageP1.
Purification andproperties.Biochem.J. 127: 1-10. 11. Doskocil, J., and Z. Sormova. 1965. The sequences of5.
methylcytosineinthe DNA ofEscherichia coli. Biochem. Biophys. Res.Commun.20:334-339.
12. Doskocil, J., and Z. Sormowi. 1965. Theoccurrence of
5-methylcytosine in bacterial deoxyribonucleic acids.
Biochem.Biophys. Acta95:513-515.
13. Fujimoto, D.,P. R.Srinivasan, andE.Borek.1965. Onthe
natureof thedeoxyribonucleicacidmethylases.Biological evidencefor themultiplenatureoftheenzymes.Biochem. 4:2849-2855.
14. Gold, M., and J. Hurwitz. 1963. Theenzymatic methylation of thenucleic acids. Cold Spring Harbor Symp. Quant. Biol. 28:149-156.
15. Gough, M.,andS. Lederberg. 1966.Methylated bases in the
host-modified deoxyribonucleic acid of Escherichia coli and bacteriophageX.J.Bacteriol.91:1460-1468.
16. Hattman,S. 1964.The controlofhcst-inducedmodification
by phage P1. Virology 23:270-271.
17. Hattman,S. 1970.DNAmethylaticnof T-evenbacteriophages andoftheir nonglucosylatedmutants:itsroleinP1-directed resttiction. Virology 42:359-367.
18.Hattman,S. 1971.Variationof6-methylaminopurinecontent
in bacteriophage P22 deoxyribenucleic acidas afunctionof
hostspecificity. J. Virol. 7:690-691.
19. Hattman, S.,E. Gold, and A.Plotnik. 1972.Methylationof
cytosine residues in DNA controlled bya drugresistance
factot. Proc. Nat. Acad.Sci. U.S.A. 69:187-190.
20. Kelly, T. J., Jr., andH.0.Smith. 1970.Arestrictionenzyme
from Heinophilus intfluentzae. II.Basesequenceof the
rec-ognition site. J. Mol. Biol. 51:393-409.
21. Kuhnlein, U., and W. Arber. 1972. Host specificity of DNA produced inEscherichia coli. XV. The role of nucleotide methylation ininvitroB-specificmodification.J.Mol. Biol. 63:9-19.
22. Lederberg, S. 1957. Suppression of the multiplication of
heterologousbacteriophage in lysogenic bacteria. Virology 3:496-513.
23. Ledinko, N. 1964. Occurience of 5-methyldeoxycytidylate in the DNA of phage lambda. J. Mol. Biol. 9:834-835. 24. Linn, S., and W. Arber. 1968. Host specificity of DNA
pro-duced byEscherichia coli. X.In vitrorestriction of phage fd replicative form. Proc. Nat. Acad.Sci. U.S.A. 59:1300-1306. 25. Mamelak, L., and H.W.Boyer.1970. Genetic control of the secondarymodification of deoxyribonucleic acid in Escher-ichia coli. J.Bacteriol. 104:57-62.
26. Meselson, M., and R. Yuan. 1968. DNA restrictionenzyme
form E.coli.Nature(London) 217:1110-1114.
27. Revel, H. R., and S. M.Hattman.1971. Mutants of T2gtwith alteredDNAmethylase activity: relation torestriction by prophageP1.Virology 45:484-495.
28. Roulland-Dussoix,D., andH. W. Boyer. 1969. The Escher-ichia coli BRestriction endonuclease. Biochim. Biophys. Acta195:219-229.
29. Smith,J.D., W. Arber, and U.Ktihnlein. 1972.Host specific-ity of DNA produced by Escherichia coli. XIV. The role of nucleotide methylation ininvivoB-specific modification. J. Mol.Biol. 63:1-8.
30.Takano, T., T.Watanabe, and T. Fukasawa. 1968. Mechanism of host-controlled restriction of bacteriophage X by R-factorsin Escherichia coli K12. Virology34:290-302. 31. Vanyushin, B. F., Ya. 1. Buryanov,and A. N.Belozersky.
1971. Distribution of N6-methyladenine in DNA of T2 phage and its host EscherichiacoliB.Nature N. Biol. 230: 25-27.
32.Watanabe,T., T. Takano, T. Arai, H. Nishida, and S. Sato. 1966. Episome-mediated transfer of drug resistance in Enterobacteriaceae. X. Restriction and modification of phagesbyfi-R-factors. J.Bacteriol. 92:477-486.
33. Wood, W. B. 1966. Host specificity of DNA produced by Escherichiacoli: bacterial mutations affecting the restriction andmodification ofDNA.J.Mol. Biol.16:118-133.
VOL. 10, 1972