JOURNAL OF VIROLOGY, Mar. 1977, p. 1159-1169 Copyright©D1977 American Society for Microbiology
Vol. 21, No. 3 Printed in U.S.A.
Characterization of
a
Temperature-Sensitive
Mutant
of
Human
Adenovirus Type 7
MARY K. ESTES AND JANET S. BUTEL*
Department of Virologyand Epidemiology, Baylor College of Medicine, Houston, Texas 77030
Received for publication 19 November 1976
Theproperties ofanaturally occurring temperature-sensitive (ts) mutant of
human adenovirus type 7 (Ad7) were studied. Mutant Ad7 (19), or E46-, was the nonhybrid adenovirus component derived from the defective simian virus 40
(SV40)-Ad7hybrid (PARA). Growth of the mutant was restricted at40.50C, and
the ratios ofvirusyieldsin KB cells at 40.5 and 330C were 10-2 to 10-3. Viral
DNA synthesis and the synthesis of adenovirus-specific antigens (tumor, capsid,
hexon, and penton antigens) appeared normal at the restrictive temperature.
The assembly of virus particles was aberrant, as determined by thin-section of
infected cells. Theinfectivity of mutant virions was heat labile at 50'C,
suggest-ing a tsdefect in a structural component of the virion. Analysis by
polyacryl-amide gel electrophoresis of [35S]methionine-labeledpolypeptides synthesized in
mutant-infected cells suggestedthat at least the major virion polypeptides were synthesized at the restrictive temperature. A lack of inhibition of host protein
synthesis lateinmutantinfections,ascomparedwithwild-type (WT) infections atboth thepermissive andnonpermissive temperatures, madequantitation of
infected-cell polypeptides difficult. Analysis of theassemblyofcapsomeres from
cytoplasmicextracts of infectedcells on sucrose gradients and by
non-dissociat-ingpolyacrylamide gel electrophoresis suggested that hexon capsomeres were
made at 40.5°C. The hexon capsomeres made by the mutant at either 33 or
40.5°C displayed a decreased migration in the non-dissociating gels compared
with the WThexon capsomeres. The molecular weights of the mutantandWT hexon polypeptides were identical. These results suggest that the ts lesion of this
group B human Ad7 mutant may bereflected in altered hexons. The mutant
Ad7 interferedwith the replication of adenovirus types 2 and 21 at the elevated temperature.
Conditional-lethalmutantsofbacteriophages and animal viruses have been valuable tools for analysis of the viral genes expression inboth theproductive and transforming cycles ofvirus
infection(10). The currentisolation of viral and
host cell mutants is allowing a more precise
analysis of the molecular mechanisms that
reg-ulateviral synthesis and cellular alterations in
eukaryotic cells. Temperature-sensitive (ts)
mutantsofadenovirustypes 2, 5,and12,
repre-senting groups A and C human adenoviruses,
and of avian orphan (CELO) adenovirus were
reported previously (9, 11, 14, 16, 20, 31-36,
39-41). The biochemical andgenetic
characteriza-tionsof these mutants haveprovided
informa-tion on the organization ofthe adenovirus
ge-nomeand have permitted identification of the
biological functions of some of the viral gene
products. Thisreport characterizes a naturally
occurringts mutantof adenovirustype7
(Ad7),
agroup B adenovirus.
MATERIALS AND METHODS
Cells. Human embryonic kidney (HEK) cells were obtained from either HEM Research, Rock-ville, Md., or Flow Laboratories, Inc., Rockville,
Md. The cellswere grown in Melnick lactalbumin
hydrolysate medium containing 10% fetal bovine
serum,0.075% sodiumbicarbonate, 100Uof penicil-lin, and 100 ,ugofstreptomycinper ml. Theywere maintainedinthesamemediumsupplementedwith 2%fetal bovineserum. HEKcellswereutilized up to passage three in tissue culture. KB cells were growninEagle medium supplementedasdescribed above.
Viruses. The prototype strain of Ad7wasobtained from the American Type Culture Collection. The strain waspassed nine times in KB cells. An Ad7 isolated from a fatal case ofpneumonia (3) was designatedAd7 (Hu) and waspassed fivetimes in HEK cells.The titerof the Ad7 (Hu) stock used for allexperiments described herewas6 x 107PFU/ml.
Mutant Ad7(19),E46-,wasderived from the paren-tal PARA-Ad7 stockby three successiveplaque pu-rificationsinHEK cells andwasthenpassed eight
1159
on November 10, 2019 by guest
http://jvi.asm.org/
1160 ESTES AND BUTEL
additionaltimes inHEKcells. Theplaque-purified progeny nolonger induced simian virus 40(SV40) antigens (4)and couldnotreplicate in green monkey kidney cells inthe absence ofSV40 (5). The titer of the Ad7 (19) stockused for allexperiments described in this report was 7 x 108 PFU/ml. Other Ad7 strains were kindly provided by H. Ginsberg and Wyeth Laboratories, Philadelphia, Pa., and were designated as such. Adenovirus type21 (Ad21) was obtained from W. Parks and was used after one passagein KBcells andtwopassagesinHEK cells. Adenovirus type 2 (Ad2) was obtained from M. Ben-yesh-Melnick and had undergone six passagesinKB cellsand one passage in HEK cells.
The adenoviruses were assayed by the plaque techniquein HEKcells as previously described (7). Virus assays wereperformed at370C. The relative plaquing efficiency of the mutant at370C/330Cwas
0.95to1.0,andat40.50C/330Cit was10-2 to10-3.
Growth curves. Growth studies wereperformed withMagniwhirl waterbaths to assure precise tem-perature control. HEK andKB cell monolayers in tubeswereinfected withAd7 (19) or Ad7 (Hu) at a multiplicity of infection (MOI) of 2 to 5 PFU/cell. After adsorption at370C, monolayers were washed with Tris-buffered saline (TBS), maintenance me-diumwasadded, and replicate tubes were incubated
at33 or40.50C. Replicate tube cultures were
har-vested at various intervals postinfection (p.i.), and theclarified lysates were assayed for virus yields as described above.
Antisera. Antisera to purified Ad2, Ad7, and Ad2M were prepared in rabbits as previously de-scribed (8). Adl2 tumor antibody was obtained from tumor-bearing hamsters. Antisera to Ad7 hexon and penton antigens, kindlydonated by G. Dreesman, were prepared in rabbits against purified hexons and pentons isolated from infected cells by gel filtra-tion onSepharose 2B, followed by sucrose gradient centrifugation (manuscript in preparation).
Immunofluorescence techniques. Monolayers of HEK cells on cover slips in Leighton tubes were infected with Ad7 (19) orAd7(Hu) and incubated in waterbaths at the permissive (33°C) or
nonpermis-sive(40.5°C)temperature.At24and48h, the cover
slips were harvested, fixed for 3 or 10 min with
acetone, and stained for tumor, capsid, hexon, or penton antigen, respectively, with the sera de-scribedabove. Thisprocedurewasfollowed by add-ing anti-rabbit globulin baboon globulin labeled with fluorescein isothiocyanate as previously de-scribed (27).
Co-infection studies. Tube cultures ofKB cells wereinoculatedsimultaneously in mixed infections with3 to 5 PFUper cell ofeach of the two viruses to be tested. Singly infected control cultures received the same multiplicity ofeach virus. After adsorp-tion, the cells were washed three times with TBS, incubatedinwater baths at33°C for 96 h or at 40.5°C for 72 h, and harvested by three cycles of freeze-thawing. The cell debris was removed by low-speed centrifugation; the supernatant fluids were pooled, treatedwiththe appropriate antiserum at37°C for 1 h, and analyzed for viral content at 37°C in HEK cells.
Viral DNA. Cultures ofHEK cells were mock
infected with TBSorinfected with virus atanMOI of 10 PFU/cell. After a 90-min adsorption period, depleted medium (12)wasadded and cultures were incubated at 33 or 40.5°C. Since initialexperiments with 2-h pulses showed that peak labeling of viral DNAoccurred atapproximately 32hp.i. at40.5°C and at 36 h p.i. at 330C, 5 ,uCi of [3H]thymidine (TdR) per ml was addedtothe cultures from 24 to48 h p.i. At48h p.i., DNAwasextractedby the Hirt procedure (13), and DNA in the supernatant fraction was purified by treatment with RNase, Pronase, andphenol. DNA wasalso extracted frompurified virionsby sequentialtreatmentwithsodium dode-cyl sulfate (SDS), RNase, and Pronase, followed by extractions withphenol. Samples containing 1 ,ug of DNAwerecentrifuged for72hat35,000 rpmat200C toequilibrium in gradients ofCsCl with an average density of 1.70 g/ml by usingaTi5O rotor. Gradients were fractionated by bottom puncture of the tube; refractive index of each fraction was determined with a Bausch and Lomb refractometer. Each frac-tion was precipitated with cold trichloroacetic acid onglass-fiber filters, dnd theradioactivity was de-terminedin aBeckman LS250liquid spectrometer. Electronmicroscopy.Cultures of HEK cellswere mock infectedorinfected with Ad7 (19)orAd7(Hu) at anMOI of10PFU/cell. After incubationat33or
40.50C for 96 or 72 h, respectively, the cells were
washed withTBS,trypsinized, pelleted by low-speed centrifugation, and fixedat 40C in3% glutaralde-hydeinMillonigsphosphate buffer forembeddingin Spurr low-viscosity media. The embedded cultures werestained with saturateduranyl acetatein50% ethanol andReynoldslead citrate, sectioned on an MT2-B ultramicrotome, and examinedon an RCA EMU 3 microscope operated at 100 kV. Duplicate cultureswereharvestedby freezingandthawingto monitor virus yields.
Heat inactivation. Virus stocks werediluted
10-fold in TBS andsonically treated for30 s. Aftera
further 100-fold dilutionincoldTBS,0.2-mlportions were dispensed into replicate tubes that were im-mersedina500Cwaterbath. Atspecified intervals, atubewasremoved, and thecontents werediluted withcold TBSandassayed for surviving infectivity by the plaquetechnique.
Analysis of viral polypeptides and capsomeres.
Polypeptides synthesized in infected cells were
la-beled byadding [35S]methionine (Amersham/Searle Corp., ArlingtonHeights, Ill.)at afinal concentra-tionof 5,Ci/mlto a culture mediumcontaining0.1 the normal concentration of methionine. Infected cultures wereexposedtothe radiolabeled precursor forvaryinglengths oftime.
Infected-cell extractswereprepared from100-mm petri dishes. The cellswereremovedby scrapingat
40C andweresedimentedat1,000 xg for5min; the cell pack was washedtwicewith ice-cold TBS. The cell pellet was suspended again in 1 ml ofTBS containing300,ugofphenylmethyl sulfonyl fluoride per ml and frozenat -20°C untilanalysis. Atthat time, the cellpelletsweredisruptedinsample buffer
(0.0625MTris,2%SDS, 5%2-mercaptoethanol,30%
sucrose, and 0.005% bromophenol blue, pH 6.8), heated to 1000C for 2 min, and analyzed by the discontinuousSDS-polyacrylamide gel
on November 10, 2019 by guest
http://jvi.asm.org/
ts MUTANT OF ADENOVIRUS TYPE 7 1161 sis (SDS-PAGE) system of Laemmli (15). Samples
were subjected to electrophoresis on slab gels 1.5-mmthickfor4h at 100 V.Separating gels contained 12%acrylamide, 0.12% bisacrylamide, 0.1% SDS, 0.5 M urea, and 0.375 M Tris-hydrochloride (pH 8.8); stacking gels contained4%acrylamide, 1% bisacryl-amide, 0.1% SDS, 0.5 M urea, and 0.05 M Tris-hydrochloride (pH 6.8).
Intact viral capsomeres were analyzed either on non-dissociating slab gels containing 5% acrylamide and 0.13% bisacrylamideinTris-glycine buffer (pH 8.3)with a spacerpreparedby the method of Maizel (21), or bysedimentation of [35S]methionine-labeled cytoplasmic extracts in 5 to 20% neutral sucrose gradients inan SW41 rotor at 39,000 rpm for 16 h (22). Cytoplasmic extracts were prepared from washed cells suspended againin 0.14 MNaCl-0.01 MTris-hydrochloride (pH 7.2)-1.5 x 10-3 MMgCl2, andlysedby the addition of NonidetP-40(NP-40)to afinalconcentrationof0.5%.After5min at40C,the nuclei were removed by centrifugation at 200xg for 5 min (6). The cytoplasmic and nuclear fractions weredialyzed against the electrophoresis buffer con-tainingphenylmethylsulfonyl fluoride priorto elec-trophoresis. Insomecases, thecytoplasmicextracts werecentrifuged at45,000 rpm at4°C for 2h in a Ti5Orotor to remove aggregated material (24), but this wasfoundtobeunnecessary. After
electropho-resis, the gels were fixed with 5% methanol-7.5% aceticacid, driedonfilterpaper, andexposedto
X-TABLE 1. Relativeyields ofseveral Ad7 isolatesfrom KBcells atthe permissive(33°C) and nonpermissive
(40.5°C) temperatures
Virus yield(PFU/ml)a from Virusyield Ad7isolate cultures incubated at: at
40.5°C/vi-rusyieldat
33°C 40.5°C 33°C
Prototype 4.0 x 107 4.0 x 107 1.0 Wyeth 4.0 x 106 8.0 x 106 2.0 Ginsberg 4.5 x 107 6.9 x 107 1.5
Hu 1.4 x 107 3.4 x 107 2.4
19 2.0 x 107 6.0 x 104 3.0 x 10-3
aVirusyields from infectedKBcellcultures
incu-bated at 33 or 40.5°C for 96 or 72 h, respectively,
weredetermined by plaque assay in HEK cells at 37°Casdescribedinthetext.
rayfilm. The developedautoradiograms were
ana-lyzedwithaCanalco model G densitometer.
RESULTS
Isolation and growth characteristicsof
mu-tant virus. Ad7 (19) was isolated by plaque
purification asthe nonhybrid adenovirus
com-ponent from the defective SV40-Ad7 hybrid
(PARA) population (4, 5). It has been used in
many subsequent studies of the defective
hy-bridvirus (26). Thets natureof thisisolatewas
investigated after the observation that ats
le-sion exists inthe adenovirus regionofthe
hy-brid PARAgenome thataffects the replication
of PARA (M. K. Estes, M. J. Guentzel, and J.
S. Butel, Abstr. Annu. Meet. Am. Soc.
Micro-biol. 1973, V36, p. 200).
Comparisons of virus yields from the
permis-sive (33°C) and restrictive (40.5°C)
tempera-tures withseveral strains of Ad7 are shown in
Table 1. All the wild-type (WT) Ad7 isolates
tested (i.e., prototype, Wyeth, Ginsberg, and
Hu)exhibited similar virus yieldsatboth
tem-peratures. In contrast, the Ad7 (19) isolate
showedatleasta300-fold reduction in yieldat
40.5°Cascompared with thatat33°C,
illustrat-ingthetsphenotype of this virus. An analysis
of thekinetics of growth of Ad7 (19)inKBcells
confirmed themarkedinhibition of virus
multi-plication at40.5°C (Table 2). The growth cycle
was essentially complete by 48 h p.i. at both
temperatures. Alsoconfirmed in Table2is the
fact that Ad7 (Hu), the WT virus used in all
furtherexperiments, didnotrevealany
restric-tion ofgrowthat40.5°C.
The Ad7 (19) mutant typically exhibits a
ra-tio ofyieldsin KBcells at40.5°C/33°Cof 10-2to
10-3. The plaquing efficiency of the uncloned
adenovirus component in theparental
PARA-Ad7 population at 40.5°C/33°C is 10-3 or less.
The cloned Ad7 (19) mutant displays greater
leak and/or reversion during growth in HEK
cells, such that the relative yield at 40.5°C to
that at33°C occasionally approaches 10-1.
Be-cause ofthis, experiments performed in HEK
TABLE 2. Kineticsofgrowth ofmutantAd7inKBcellsa Incubation Virus yield (PFU/ml) at hours p.i.:
Virustemp (~C) 6 24 48 72 96
Ad7 (19) 33 5.0 x 103 5.0 x 105 1.5 x 107 2.2 x 107 2.4 x 107
40.5 2.8 x 103 1.2 x 103 1.1 X 105 6.4 x 104 6.2 x 104
Ad7(Hu) 33 NDb ND ND ND 6.0 X 106
40.5 ND ND ND ND 6.0 x 106
aVirusyieldsfrom infected KB cultures incubated at 33 or40.5°Cforindicatedtimesweredeterminedby
plaque assayin HEKcellsat37°C, asdescribedinthe text.
bND, Not done. VOL. 21, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.57.246.395.487.2] [image:3.501.60.452.555.635.2]1162 ESTES AND BUTEL
cells always included duplicate cultures
har-vestedtomonitor theamountof leak and
rever-sion; no results were considered valid if the
ratio of virus yield at 40.5°C/33°C wasgreater
than 10-2.
Induction of viral DNA synthesis. The abil-ityoftsmutantAd7 (19)toinduce thesynthesis
ofviral DNA in HEK cells under
nonpermis-sive conditions was examined by using a 2-h
pulse of [3H]TdRatvarious timesp.i. (Table 3).
The incorporation of [3H]TdR by mock-infected
control HEK cells was greatly decreased at
40.5°C compared with that at 33°C; markedly
enhanced incorporation of label did occur at
40.5°C, however, when the cellswere virus
in-fected. These datasuggested that the majority
ofthe DNA being synthesized at40.5°C by 34h
p.i. was probably virus specific. However, to
confirm that the DNA synthesized at 40.5°C
was viral in nature, samples from infected
HEK cells wereanalyzed by equilibrium
centi-fugation. The DNA in Hirt supernatant
frac-tions obtained from infectedHEKcells, which
had been labeled with [3H]TdR from24 to 48 h
p.i., was characterized. A similar,
homogene-ous peak of labeled material that bandedat a
density of 1.710 g/ml was obtained from
mu-tant-infected cells incubated under either
per-missive or nonpermissive conditions (Fig. 1).
DNA with a similar buoyant density was
ex-tracted from WT-infected cells and from
puri-fied Ad7 (Hu) virions (datanot shown).
Induction of virus-specific antigens.
Mu-tant-infected cells weremonitoredby
immuno-fluorescence for the production of virus-specific
antigens. HEKcells infectedwith Ad7 (Hu) or
Ad7(19)atanMOI of 2PFU/cellwerefixed and
examined for the presence of tumor, capsid,
hexon, andpentonantigens.As shown inTable
4, all these virus-specificantigenswere
synthe-TABLE 3. Incorporationof[3H]TdRinto trichloroaceticacid-precipitablematerialbyHEK
cells infectedwith Ad7(19)at33or40.5oCa
cpm/,g of DNA Time of
pulse a- 33°C 40.5°C
bel(h
p.i.) Mock in- Ad7 (19) Mockin Ad7 (19)
fected fected
0-2 3,460 3,500 1,966 900
24-26 6,871 6,666 49 4,200
32-34 5,640 10,750 50 11,549 36-38 4,465 21,989 50 4,223
48-50 1,708 7,404 100 787
aControlorvirus-infected HEK cellswerelabeled
with 5
ACi
of[3H]TdRpermlfor the indicated timeintervals p.i. Cellextracts wereprepared and
pre-cipitatedwithtrichloroacetic acidatthe endof the pulse timeasdescribedinthetext.
4
C-)
x
I
3
2
1
1.80
1.70 gm/ml
1.60
FRACTION NUMBER
FIG. 1. Densitygradient analysis of DNA
[image:4.501.261.452.61.256.2]synthe-sized in Ad7 (19)-infected HEK cells at 33 and
40.50C. Cultures of HEK cells were infected withan
MOI of 10 PFUlcell and labeled with 5puCi of [3H]TdR per ml from24to 48h p.i.Viral DNA was extractedby the Hirt procedure,purified,and centri-fugedtoequilibriuminCsCl for72hat35,000 rpm at200Cin a TiSO rotor. Symbols: *, DNA synthe-sizedat330C; 0,DNAsynthesizedat40.50C. Arrow indicates positionatwhich marker-cell DNA bands. TABLE 4. SynthesisofvirusantigensinHEKcells
at40.50Ca
Synthesis of virus-specific antigen: Virus
Tumor Capsid Hexon +
fiber
Ad7 (19)b + + + +
Ad7(Hu) + + + +
a
HEK
cells growingon coverslipswereinfectedwithAd7 (19) orAd7 (Hu) at anMOI of2PFU/cell. At48 hp.i., the cellswerefixed withacetoneand stainedby the indirect immunofluorescence test as describedinthetext.
Relative yield of mutant
(40.50C/330C)
= 1 x 10-2.e +,Presence ofantigen.Approximately40 to 50% of the cellsin a culture were positive for a given antigen. Allfluorescentreactions were observedin the nucleus.
sized at 40.50C, with approximately the same
percentage of cells showing positive
fluores-cence at40.50C as at 330Cfor eachgiven
anti-gen. All the virusantigens detectedby
immu-nofluorescence werelocated in the nucleus.
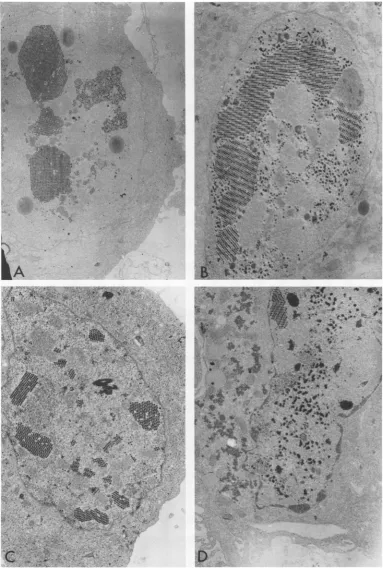
Production ofvirus particles. Electron
mi-croscopic examination of thin sections of
in-fected HEK cells wasperformed. The nuclei of
WT Ad7 (Hu)-infectedcellsrevealed large
crys-talline arrays of virus at both temperatures.
Typical WT-infected cells at 33 and40.5°C are
showninFig.2Aand B. Thecytoplasmic
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.262.452.378.440.2]MA
An'~~~~~~~~~~~~~~r
.. ;.- ~W 'P O
FIG. 2. Electronmicrographs of thin sections ofHEK cells infected with Ad7 (Hu) or Ad7 (19) for 72 or 96 hat33 or40.5 C,respectively. (A) Ad7 (Hu)-infected HEK cell at 330C. x4655. (B) Ad7 (Hu)-infected HEK cellat40.50C. x7130. (C) Ad7 (19)-infected HEK cell at 330C. x8060. (D) Ad7 (19)-infected HEK cell at 40.50C. x6080.
1163
-31
;.". a
... ..-t.
-:. ..-v".", 1; 't
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.59.443.51.620.2]1164 ESTES AND
phous material seen in mutant-infected cells at
40.5'C, and describedbelow, waslackinginthe
WT-infected cells,
although
somenuclear inclu-sionbodieswere seen(Fig. 2B). Thepercentageof cells infected at 40.50C with Ad7 (Hu) was
approximatelyhalf that observed at330C, even
though the WT virus produced 108 PFU/ml at 40.50C.
Cells infected with Ad7 (19) revealed that
intact virusparticleswereapparentlynotmade
as efficiently at 40.50C as at 330C. In cells
in-fected at330C, 80 to 90% of the nuclei contained
vast crystalline arrays of virus (Fig. 2C). In
contrast, virus particles were observed inless
than50%of thecells maintainedat40.50C,and very few crystalline arrays were detected.
In-stead, large areas of nuclear inclusion bodies
were seen, and amorphous substances were
prominent in the cytoplasm of cultures from
40.50C (Fig. 2D). TypeIandtype IIinclusions,
asdescribed for Ad12 infections (23), were sel-dom seen at 330C but were common atthe
re-strictivetemperature. Nuclear membrane
frag-mentation occurred late after infection (not
shown) at
330C,
but itwas seldomobserved at40.50C
in mutant-infected cells. The relativeyield of the ts mutant in these experiments
(40.50C/330C)
was7.5 x 10-3.Heatstability ofmutant virions. That Ad7
(19) particle assembly appeared altered at the
nonpermissive temperatureraised the possibil-ity that a structural component of the virion might beaberrant. Therefore,the effect of
heat-ing at500Contheinfectivityof boththe mutant
Ad7 (19) and the WT Ad7 (Hu) virus grown at
37°C was examined (Fig. 3). The infectivity of
Ad7 (19) was inactivated more rapidly than
that of theWT virus; only 1%of theinfectious
mutant virus survived after 9 min at 500C,
whereas about 26min ofheatingwasrequired
toreduce theWTinfectivitytoasimilar level. The heatlability of the mutant virus suggests
that thetsdefectmay lie in astructural compo-nent of thevirion.
Synthesis of polypeptides in infected cells.
The technique of PAGE is a powerful tool for
probing events in infected cells at the
poly-peptide level. The synthesis of polypeptides
in WT-and Ad7 (19)-infectedcellslabeledwith
[35S]methionine wasmonitored todetermineif
any structural polypeptides were lacking that
might account for the aberrant virion
assem-bly. Analysis by SDS-PAGE of cells labeled
from 18to 42 h p.i. at 40.5°C or from 24 to 48 h
p.i. at 33°C revealed that at least the major
structural polypeptides were synthesized in
both mutant- and WT-infected cells at both
temperatures (Fig. 4). Polypeptides II, III, IV,
V, andVI, asdesignatedby Maizel (22), can be
100
0
r
(L-C., a
-n
(f)
I--a
LLI
Q~
10
10 20 30
MINUTES AT 50 C
FIG. 3. Heat inactivation of Ad7 (Hu) and Ad7 (19)at50°C.Portionsofsonically treated virus stocks were immersed ina waterbath at50°C forvarying intervalsoftime and thenassayedfor surviving
in-fectivity, asdescribed in thetext.
detected, and the patterns in mutant- and
WT-infectedcells were similar when compared at 33
or40.5°C. Becausethese experiments withlong
labeling periods did not reveal differences in the polypeptide patterns between the Wt- and
mutant-infected cells at the nonpermissive
temperature,noattempts were made to
investi-gate the processing ofpolypeptides.
Oneinteresting observationwasthat the
mu-tant virions did not appear to shut off host
proteinsynthesis aseffectivelyasWTvirus,as
evidencedbythehigher levels of background in
the mutant-infected cells at both thepermissive
and nonpermissive temperatures. All the
ex-tracts for polypeptide analysis were prepared
from cells infected with 5 PFU of virusper cell,
and 100,000 counts of radioactivity was
ana-lyzed with eachsample. Experiments designed
tofurthercharacterizethe lack of inhibition of
host protein synthesis were hampered by the
leakiness exhibited by this mutant at higher multiplicities of infection. The lack of
inhibi-tion ofsynthesis of host proteins late in
infec-tionobscures thequantitation of the
virus-spec-ified proteinsin the mutant-infected cells.
Al-though the major virionpolypeptides appeared
tobe synthesized in mutant-infectedcells,SDS
gelsseparate polypeptides onlyonthe basis of
molecularweight, and,therefore, these
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.285.426.64.315.2]ts MUTANT OF ADENOVIRUS TYPE 7 1165
4*
.4 E n
-1*0 ma
A
i
w
II
S
-_F
A_
vCi
1 _ _r~~-
11
i - 94K
-_-
-III
...A
Imokl'
--
iia
50K--v
- 45K
-
VI
- 23.5K_W- ..- Vil
vIi*
A.B
A
B
C
D
E
F
G
H
I
FIG. 4. SDS-PAGE autoradiogram of Ad7 (Hu)- or Ad7(19)-infected KB cells labeled from18to 42hat
40.50Corfrom24to 48h at330Cwith 5pLCiof[35S]methionineper ml.Plates (100 mm)infected withanMOI
of5PFU/cellwereprocessed as describedin the text, and approximately 100,000cpm wasappliedtoeach
samplewell. Thesampleorder isasfollows: (A) mock-infectedcellsat330C; (B)Ad7(Hu)-infectedcellsat
33C; (C) Ad7(19)-infected cells at 33 C; (D) mock-infected cells at40.50C; (E)Ad7(Hu)-infected cells at
40.50C; (F) Ad7 (19)-infected cells at40.50C; (G)purified [35S]methionine-labeled virus; (H) hexon band
elatedfrom the non-dissociating polyacrylamide gel ofAd7 (Hu)-infectedKBcell cytoplasmic extractshown inFig. 6C; and (I) hexon band elated from the non-dissociatingpolyacrylamide gel ofAd7 (19)-infected KB cell cytoplasmic extract shown in Fig. 6B. Wells (A) through (G) were subjected to electrophoresis inthe same slab gel. Roman numerals indicate the viral polypeptides (nomenclature ofMaizel[22]) and the arabic numbers indicate the location of molecular weight markers detected by Coomassie brilliant blue staining: phosphorylase A, 94,000; heavy-chain gamma globulin, 50,000; SV40 polypeptide I, 45,000; and light-chain gammaglobulin, 23,500.
ments could not determine ifassembly of the
polypeptides intocapsomereswasaberrant.
Synthesis of viralcapsomeres.Todetermine
ifthe viral polypeptides were assembled into
capsomeres, cytoplasmic extracts of WT- and
mutant-infected cells labeled with
[35S]methio-ninewereanalyzedonsucrosegradientsandon
non-dissociating polyacrylamide slab gels (pH
8.3). From the sucrose gradients, it was
ap-parent that, although the extent of
incorpora-tion of radioactivity was reduced at 40.50C,
capsomereswereinfactformed(Fig. 5).
Analy-sis of fractions from the sucrose gradient by
SDS-PAGE showed that they contained
pre-dominately the hexon andpentonpolypeptides.
This observation is in agreement with earlier
work with Ad2 (17).
An analysis of infected-cell cytoplasmic
ex-tracts (viral capsomeres) on 5%
non-dissociat-ing polyacrylamide gels (pH 8.3) revealed the
presenceofonemajorandoneminorband that
were not present in extracts ofmock-infected
cells (Fig. 6). The major band from both WT-andmutant-infected cellextracts wasidentified
asthe hexon
polypeptide by eluting
the band andanalyzingitby SDS-PAGE (Fig.4HandI). The Ad7 (19) hexon had a molecularweight
identicaltothat ofWThexon whenanalyzedon5or12%SDS-PAGE. The minor band could not
beanalyzedfurther due toinsufficient
incorpo-rationofradiolabelcounts. Thematerialatthe
top of thegels, seen inFig6BandC, contained
primarilythe corepolypeptidesVand VII and
specifically lacked hexon
polypeptide.
Thebands at the bottom represent free
[35S]_
methionine.A comparison of the mobility of the hexon
capsomeres from extracts of WT- and
mutant-VOL. 21, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.53.451.74.345.2]1166
ESTES AND BUTEL0o
m
CD
x
z
2
LLJ
AAd7 (19)-33 C AAd 7 (19)-40.5 C oAd 7 (Hu)-33 C
*Ad7(Hu)-40.5 C
5 10 15 20 25
FRACTION NUMBER
FIG. 5. Sucrose gradientcentrifugation of capso-meresfrom cytoplasmicextractsof KB cells infected with Ad7(Hu)orAd7 (19) atanMOI of 5 PFU/cell
and labeled with [35S]methionine from 24 to 48 h (330C) or from 18 to 42 h (40.50C). Cytoplasmic
extractsweremade andanalyzedasdescribed in the
text.Thecapsomereregion(fractions 16 through 19) contained both hexon and pentonpolypeptides, as
shown bySDS-PAGE analysis. The variousextracts
areindicatedby the following symbols:0,Ad7
(Hu)-infected KB cells, 330C; 0, Ad7 (Hu)-infected KB
cells, 40.50C; A, Ad7 (19)-infected KB cells, 330C; and A,Ad7(19)-infected KB cells, 40.50C.
infected cellsinnon-dissociating gels, however,
revealed a decrease inthe mobilityof the
mu-tant hexon (Fig. 6B through E). This altered
mobility wasevident in hexons extracted from
KB or HEK cultures incubated at the
per-missive (330C) as well as the nonpermissive
(40.50C) temperatures. Two bands are evident
when themutantand WTcapsomeres are
sub-jected toco-electrophoresis (Fig. 6F).
Hexons extracted from cells infected with
Ad7prototypeorAd7 (Ginsberg) showed
identi-calmigrationtothe Ad7(Hu)WThexons(data
not shown). These resultssuggest that the
de-fective structural protein of the Ad7 (19)
mu-tantmaywell be the hexonpolypeptide.
Interference with replication of
heterolo-gous adenoviruses. Co-infection experiments
in KB cells were performed as described in
Materials and Methods to ascertain whether the mutantlesioninAd7(19)exertedanyeffect
on the replication of heterologous Ad2 and
Ad2l. In the presence ofco-infecting WT Ad7
(Hu), less thanatwofoldreductioninthe yields
of Ad2l orAd2 at either the permissiveorthe
nonpermissive temperature took place (Table
5). Similarly, in the presence of co-infecting
Ad7 (19) under permissive conditions (330C),
minimal reductions in the yields of
heterolo-gousvirusesoccurred(1.8-foldfor Ad2land
7.8-foldwith Ad2.). However, under conditions of
incubation that block the production of
infec-tious mutant viruses (40.50C), there were
marked reductions in the yields of both Ad2M
(18-fold) and Ad2 (19-fold). The basis for this
interference exerted by mutant Ad7 (19), and
notby WTAd7, is not known.Onepossibilityis
that a mutant protein bearing the Ad7 (19)
lesion is used by the co-infectingheterologous
adenoviruses andrenders them either
noninfec-tious orunabletoreplicate efficiently.
Titration of the Ad7 yields from the doubly
infected cellsrevealedthat, in some instances,
the presence ofAd2 orAd2M hadadeleterious
effecton the replication of human Ad7 (Table
6). Ad2M co-infection resulted in a marked
de-creaseinyields (112-fold)of Ad7(Hu) at40.50C
and of Ad7 (19) at 330C (84-fold). Ad2M had
A
B
C
D
E
F
G
FIG. 6. Non-dissociating PAGE autoradiogram of Ad7 (Hu) or Ad7 (19) viral capsomeres. KB cells infectedat an MOIof 5PFU/cell werelabeled with
5,uCiof[35S]methionineper ml from 24 to 48 h(330C)
or 18 to 42 h (40.50C). Cytoplasmic extracts were preparedasdescribed in the text. One hundred thou-sand counts weresubjectedtoelectrophoresis atpH 8.3 inTris-glycine buffer. The anode is at the bottom. The cytoplasmic extracts were from the following samples:(A) mock-infected KB cells,330C;(B) Ad7 (19)-infectedKB cells,330C; (C) Ad7 (Hu)-infected KB cells, 330C; (D) Ad7 (19)-infected KB cells,
40.50C;(E) Ad7(Hu)-infectedKBcells,40.50C; (F) mixture ofequal amounts ofsamples (D) and (E) and
(G)mock-infectedKBcells, 40.5°C. (A), (B), and (C)
were from one gel analysis and can be compared directly; (D), (E), (F), and(G) were from a separate experiment.
J. VIROL.
IC
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.56.245.63.244.2] [image:8.501.264.452.282.499.2]ts MUTANT OF ADENOVIRUS TYPE 7 1167
TABLE 5. Interference by mutant Ad7 on replication of heterologous adenoviruses at elevatedtemperatures
Yieldof heterologousvirus inpresenceof:
Ad7 (Hu) Ad7(19)
Heterol- Ad7
ogous vi- pres- 33°C 40.5aC 33°C 40.5aC
rus ent
PFU/ml duction'Fold re- PFU/l Fold re- PFU/ml Fold re- PFU/ml
reduc-a m duction duction P
tion
Ad2M 0 1.0 x 108 - 3.9 x 107 - 3.6 x 106 - 1.0x 107
-+ 8.0 x 107 1.2 2.2 x 107 1.7 2.0 x 106 1.8 5.5 x 105 18
Ad2 0 6.0 x 107 - 7.2 x 105 - 3.9 x 107 - 3.9 x 105
-+ 3.6 x 107 1.6 4.1 x 105 1.7 5.0 x 106 7.8 2.0 x 104 19
a Fold reduction=yieldofheterologous virus in absence of Ad7/yield of heterologous virus in presence of
Ad7.
TABLE 6. Effect of heterologous adenoviruses on replication of human Ad7 Yieldof:
Heterolo- Ad7 (Hu) Ad7 (19)
gousvirus 33°C 40.5°C 33°C 40.5°C
present_________________________________________________
PFU/ml ductionFold re- PFU/ml Fold re-duction PFU/ml Fold re- PFU/ml Fold
re-duction duction
None 4.7 x 107 - 2.7 x 107 - 4.2 x 107 - 1.5 x 105
-Ad2M 7.3 x 106 6.4 2.4 x 105 112 5.0 x 105 84 2.4 x 105 0
Ad2 4.3 x 105 109 1.7 x 106 15.8 5.9 x 106 7.1 7.0 x 104 2
a Fold reduction =yield of Ad7inabsence of heterologous virus/yield of Ad7 in presence of heterologous virus.
considerably less effect on replication of Ad7
(Hu) at 330C. Co-infection with Ad2 caused a
decrease in Ad7 (Hu) yields at both 33 and
40.5°C (109-fold and 15.8-fold, respectively).
Ad2 did not inhibit Ad7 (19) replication to as
great an extent (7.1-fold at 33°C). Asexpected,
since Ad7 (19) was ts for replication, the
co-infecting heterologous adenoviruses had little effect on yields at 40.5°C. The basis for the
inhibitory effects on Ad7 replication by Ad2M
and Ad2 iscurrentlyunknown.
DISCUSSION
This report characterizes a naturally
occur-ring ts mutant of
weakly
oncogenic humanAd7. Thismutant, Ad7 (19), appearstobe de-fective in a late gene function that interferes
with the assembly ofvirus particles. Particles
made at the permissive temperature exhibit
greater thermolability than do WT virions,
which suggests that the defect may involve a
virion structural polypeptide. Analysis by
PAGErevealedthat the major structural poly-peptides (hexon, penton, fiber, and core) are
synthesizedat40.5°C.
Precise quantitation ofvirus-specific protein
synthesis was difficult, because the inhibition
ofhost protein synthesis by the mutant, was
notefficienteven atlate stagesafter infection.
This apparent lack of inhibition of cellular
mac-romolecular synthesis has not been described
forassemblymutantsof otheradenovirus
sero-types.Thepossible regulatory role of the hexon
polypeptide is intriguing. Previous studies (2)
suggestedarelationship between the synthesis
of capsid proteins and the inhibition of host
proteinsynthesis. Although the suggestion was
made thatthe fiber antigenmight perform this
function(2, 18), ithas been shown that detecta-ble levels of fiber are not present at the time
hostprotein synthesisbegins to decline (1, 29,
38). Hexonpolypeptides, however, are
detecta-ble at that time (1, 29), and our results are
consistent with the possibility that the hexon
may be involved in inhibition ofhost protein
synthesis.
Levine and Ginsberg (19) showed that both
fiber and hexon antigenscan bind to KB cell
and adenovirus type5 (Ad5) virus DNAin
vi-troandinhibitDNA-dependentRNA
polymer-aseactivity. However,the functioningofthese
proteinsinvivoremains unproven. Thelack of
bindingto cell DNAby the aberrant Ad7 (19)
hexonwould substantiate these theories.
Analysisonnon-dissociating gelsof
cytoplas-mic extracts from infected cells revealed an
alteredmobilityfor the hexon capsomeres,
sug-gestingthat thets lesionresidesin the hexon
VOL. 21, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
1168 ESTES AND BUTEL
gene. The altered
mobility
could be due to achange in the amino acid composition of the hexonpolypeptide such that its chargewas al-tered.Slight variationsinthe
migration
ingels
of hexons from three different adenovirussero-types (types 2, 3, and 5) have been reported;
concomitant amino acid analyses revealed a
lower glutamic acid
composition
in the leastelectrophoretically
mobile adenovirus type 3 (Ad3) hexons (25).The possibility that the Ad7 (19) mutation that affects assembly might involve a lack of
viral polypeptidetransportisunlikely, but the
accumulation in the cytoplasm of large
amounts of
amorphous
material detectedby
electron microscopy remains
unexplained.
Thetransport of the hexon and penton
antigens
does not appear to be
altered;
these antigensweredetectedinthe nucleus
by
indirectimmu-nofluoresence. PAGE analysis of nuclear
frac-tions from infected cells also showed the
pres-ence of the hexon
polypeptide
in the nucleus.However, it ispossible that the altered hexon
mayhavemodifiedthetransport ofother
poly-peptides
involvedinthe maturationprocess. It should be noted that the hexonantiserumusedin this studywould not detect hexon
polypep-tidesthat might accumulate in thecytoplasm
(32). Since intact hexons are found
predomi-nantly in the nucleus, it is possible that the
hexons being analyzed are actually nuclear
hexonsthat leak intothe cytoplasmduringthe
extractionprocedure. Asimilarprocedure does
extract the intranuclear SV40 tumor antigen
(37).
The characteristics of the Ad7 (19) mutant
arereminiscent of the adenovirustype 5classI
mutants which fallinto four complementation
groups (42). ClassI mutantsareassembly
mu-tants;they shownomajordefectsinthe
produc-tionofcapsid antigens (28) andno marked
re-ductioninthe synthesis ofcapsidpolypeptides
at the restrictive temperature (30).
Thermola-bility of the type 5 mutants and synthesis of
capsomereshavenotbeenreported. Of the class
Itype 5 mutants,some could undergo intertypic
complementation with WTAdl2 inHela cells;
the resultantprogeny of crosses between several mutants with defects in hexon transport or pro-duction exhibited phenotypic mixing. In that
system,theAdl2did not inhibit theAd5
multi-plicationin Hela orhamster embryo cells, but the
effect
of the Ad5 mutants on thereplicationof WTAdl2was nottested (42, 43). These
obser-vationsare of interest, since the Ad7 (19)
mu-tantappears to affect the multiplication of
het-erologous adenoviruses in mixed infections in
KBcells, and phenotypic mixing between these
serotypes also appears to occur (M. K. Estes
andJ. S. Butel,unpublisheddata).
Although theAd7mutantdescribed here was
a natural isolate, its defect appears different
thanthe natural block inassemblyobservedat
42°C with adenovirus 2 (17, 24). In that system,
there was a failure of the hexon polypeptides (monomers) to assemble into capsomeres
(tri-mers) at 42°C (17). The number of capsomeres
detected on non-dissociating gels at 42°C was
much decreased. The trimers and monomers,
however, exhibited identical tryptic peptides,
suggestingthat something other thanjust the primary structure is involved in the assembly
of Ad2at42°C. Our datashow that theAd7(19)
capsomeres are made at 40.5°C, but their
al-tered migration on the non-dissociating gels
suggests that the Ad7 (19) block in assembly
may reflect achange in the primary structure
of the hexon polypeptide, which affects the
hexoncapsomeres. There are obviously
multi-plesteps in assembly; further studies are
neces-sary to determine if the observed biological
changes with Ad7 (19) are due to a direct or indirect effect of the altered hexon polypeptide.
ACKNOWLEDGMENTS
Wewould like to thank Betty Altenburg for performing theelectron microscopic work. We are also grateful to Phil Hopperand Peggy Sansone for excellent technical assist-ance.
This work was supported by Public Health Service re-search grant CA 10,893 from the National Cancer Institute. M.K.E. is therecipient of Public Health Service postdoc-toralfellowship 1-F22-CA 03209 from the National Cancer Institute, and J.S.B. is the recipient of Faculty Research Award PRA-95 from the American Cancer Society.
LITERATURE CITED
1. Anderson, C.W., P. R. Baum, and R. F. Gesteland. 1973.Processingofadenovirus 2-induced proteins. J. Virol. 12:241-252.
2. Bello, L.J., and H.S. Ginsberg.1967.Inhibitionof host proteinsynthesisintype5adenovirus-infectedcells. J.Virol. 1:843-850.
3. Benyesh-Melnick,M., and H. S. Rosenberg. 1964. The isolation ofadenovirus type 7 from a fatal caseof pneumonia and disseminated disease. J. Pediatr. 64:83-87.
4. Boeye,A., J.L.Melnick, and F. Rapp.1965. Adenovi-rus-SV40"hybrids": plaquepurification intolinesin which the determinant for the SV40tumorantigen is lostorretained. Virology 26:511-512.
5. Boeye, A., J. L. Melnick, and F. Rapp. 1966. SV40-adenovirus"hybrids":presence oftwogenotypesand the requirement of theircomplementation for viral replication. Virology28:56-70.
6. Borum, T. W., M. D. Scharff, andE.Robbins. 1967. Preparation of mammalianpolyribosomes with deter-gent NonidetP-40. Biochim.Biophys. Acta 149:302-304.
7. Butel, J. S., and F. Rapp. 1966. Replication in simian cellsof defectiveviruses in anSV40-adenovirus "hy-brid"population.J.Bacteriol. 91:278-284.
8. Butel, J. S., and F. Rapp. 1967. Complementation be-tween a defective monkey cell-adapting component and humanadenoviruses in simian cells. Virology 31:573-584.
9. Ensinger, M. J., and H. S.Ginsberg. 1972. Selection
on November 10, 2019 by guest
http://jvi.asm.org/
ts MUTANT OF ADENOVIRUS TYPE 7 1169 and preliminarycharacterization of
temperature-sen-sitivemutantsoftype5adenovirus.J.Virol. 10:328-339.
10. Fenner,F., B. R.McAuslan, C. A. Mims, J. Sambrook, and D.0.White.1974.Thebiology of animal viruses.
AcademicPressInc., New York.
11. Hama, S., and G. Kimura.1972.Temperature-sensitive
mutantsofhuman adenovirustype12.Jpn. J. Micro-biol.16:337-338.
12. Henry, P., P. H. Black, M. N. Oxman, and S. M. Weissman. 1966. Stimulation of DNA synthesis in
mousecell line 3T3 by simian virus40. Proc. Natl. Acad. Sci. U.S.A.56:1170-1176.
13. Hirt, B. 1967. Selective extraction ofpolyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369.
14. Ishibashi, M.1971.Temperature-sensitive conditional-lethal mutants ofan avianadenovirus (CELO). I. Isolation andcharacterization. Virology 45:42-52. 15. Laemmli, U. K. 1970. Cleavageofstructuralproteins
during the assembly of the head of bacteriophageT4. Nature (London) 227:680-685.
16. Ledinko, N. 1974. Temperature-sensitive mutants of adenovirustype12defectiveinviral DNA synthesis. J. Virol.14:457-468.
17. Leibowitz, J., and M. S. Horowitz. 1974. Synthesis and assembly of adenovirustype 2 polypeptides. II. Re-versible inhibitionof hexonassemblyat42°. Virology 61:129-139.
18. Levine, A. J., and H. S. Ginsberg.1967. Mechanism by which fiber antigen inhibits multiplication oftype5 adenovirus. J. Virol. 1:747-757.
19. Levine, A. J., and H.S. Ginsberg.1968.Roleof
adenovi-russtructural proteins in the cessationof host-cell biosynthetic functions. J. Virol. 2:430-439. 20. Lundholm, U., and W. Doerfler. 1971.
Temperature-sensitive mutants ofhumanadenovirustype12. Vi-rology45:827-829.
21. Maizel, J. V. 1967. Acrylamide gel electrophoresis of proteins andnucleic acids,pp.334-362.InK.Habel
and N. P.Salzman(ed.), Fundamental techniquesin virology. Academic Press Inc., New York.
22. Maizel, J. V., Jr., D.0.White, and M.D.Scharff.1968.
Thepolypeptidesofadenovirus.I.Evidence for multi-ple proteincomponentsinthevirionand comparison of types2,7Aand12.Virology 36:115-125. 23. Martinez-Palomo, A., J. LeBuis, and W. Bernhard.
1967. Electron microscopy of adenovirus12 replica-tion. I. Fine structural changes in the nucleus of
infectedKBcells. J. Virol. 1:817-829.
24. Okubo, C. K., and H.J. Raskas. 1971. Thermosensi-tive events inthereplicationofadenovirustype2 at 42°. Virology 46:175-182.
25. Pettersson, U.1971.Strcturalproteinsof adenoviruses. VI.On theantigenic determinants of thehexon. Vi-rology 43:123-136.
26. Rapp,F. 1973.ThePARA-adenoviruses, p.104-137. In
L. P. Merkow and M. Slifkin (ed.), Progressin
ex-perimental tumorresearch, vol. 18. Karger, Basel, Switzerland.
27. Rapp, F.,S.Pauluzzi,and J. S. Butel. 1969. Variation
inpropertiesofplaqueprogenyof PARA (defective
simian papovavirus40)-adenovirus7.J.Virol. 4:626-631.
28. Russell, W. C., C. Newman, and J F. Williams. 1972. Characterization oftemperature-sensitivemutantsof adenovirus type 5-serology. J. Gen. Virol. 17:265-279.
29. Russell, W. C., and J. J. Skehel.1972.Thepolypeptides of adenovirus-infectedcells.J. Gen.Virol. 15:45-57. 30. Russell, W. C., and J. J.Skehel.1974.Characterization of temperature-sensitivemutantsof adenovirustype 5:synthesis ofpolypeptides in infected cells. J. Gen. Virol.24:247-259.
31. Shiroki, K., J. Irisawa, and H. Shimojo. 1972. Isolation and apreliminary characterization of temperature-sensitive mutantsof adenovirus12.Virology49:1-11. 32. Stinski, M. F., and H. S.Ginsberg. 1974. Antibodyto type5adenovirus hexonpolypeptide: detection of
nas-cent polypeptides in the cytoplasm of infected KB cells.Intervirology4:226-236.
33. Suzuki, E.,and H.Shimojo.1971. A temperature-sen-sitive mutant of adenovirus 31, defective in viral deoxyribonucleic acid replication. Virology 43:488-494.
34. Suzuki, E.,H.Shimojo, and Y.Moritsugu. 1972. Iso-lation and preliminary characterization of tempera-ture-sensitive mutants of adenovirus 31. Virology 49:426-436.
35. Takahashi, M.1972.Isolationofconditional-lethal
mu-tants(temperature-sensitive andhost-dependent
mu-tants)of adenovirustype5.Virology49:815-817. 36. Takemori, N., J. L. Riggs, and C. H. Aldrich. 1968.
Genetic studies with tumorigenic adenoviruses. I.
Isolationofcytocidal (cyt)mutantsof adenovirustype 12.Virology 36:575-586.
37. Tegtmeyer, P., M. Schwartz, J. K. Collins, and K. Rundell. 1975.Regulationof tumorantigen synthesis bysimian virus 40geneA.J. Virol.16:168-178. 38. Walter, G.,and J. V.Maizel,Jr. 1974. Thepolypeptides
ofadenovirus. IV.Detectionofearlyand late virus-inducedpolypeptidesand their distribution in subcel-lular fractions.Virology57:402-408.
39. Weber, J., M. Begin, and G. Khittoo. 1975. Genetic analysisof adenovirus type 2.II. Preliminary pheno-typic characterization oftemperature-sensitive
mu-tants.J. Virol. 15:1049-1056.
40. Williams, J. F., M. Gharpure, S. Ustacelebi, and S. McDonald. 1971. Isolation oftemperature-sensitive
mutantsof adenovirus type5. J. Gen. Virol.
11:95-101.
41. Williams, J. F., and S. Ustacelebi.1971. Complementa-tion and recombination withtemperature-sensitive mutantsofadenovirustype5. J. Gen. Virol. 13:345-348.
42. Williams,J.F.,C. S. H.Young,and P. E. Austin. 1974. Geneticanalysisof human adenovirus type 5 in
per-missiveandnon-permissivecells. ColdSpringHarbor Symp. Quant.Biol. 39:427-437.
43. Williams, J., H.Young,and P. Austin. 1975. Comple-mentation of human adenovirus type 5tsmutantsby humanadenovirus type 12. J. Virol. 15:675-678. VOL. 21, 1977

![FIG.1.40.50C.sized[3H]TdRfugedatMOIextractedsizedindicates 200C Density gradient analysis ofDNA synthe- in Ad7 (19)-infected HEK cells at 33 and Cultures ofHEK cells were infected with an of 10 PFUlcell and labeled with 5puCi of per ml from 24 to 48 h p.i](https://thumb-us.123doks.com/thumbv2/123dok_us/1548474.107378/4.501.261.452.61.256/tdrfugedatmoiextractedsizedindicates-density-gradient-analysis-infected-cultures-infected-pfulcell.webp)

![FIG. 4.phosphorylasegammasample40.50C;40.50Cofelatedslab33cellnumbersin Fig. 5 C; SDS-PAGE autoradiogram ofAd7 (Hu)- or Ad7 (19)-infected KB cells labeled from 18 to 42 h at or from 24 to 48 h at 330C with 5 pLCi of[35S]methionine per ml](https://thumb-us.123doks.com/thumbv2/123dok_us/1548474.107378/7.501.53.451.74.345/phosphorylasegammasample-cofelatedslab-cellnumbersin-autoradiogram-infected-cells-labeled-methionine.webp)
![FIG.5.andandextractsshownarecells,text.merescontainedwithinfected(330C) Sucrose gradient centrifugation of capso- from cytoplasmic extracts ofKB cells infected Ad7 (Hu) or Ad7 (19) at an MOI of 5 PFU/cell labeled with [35S]methionine from 24 to 48 hor from](https://thumb-us.123doks.com/thumbv2/123dok_us/1548474.107378/8.501.264.452.282.499/andandextractsshownarecells-merescontainedwithinfected-sucrose-gradient-centrifugation-cytoplasmic-extracts-methionine.webp)

