Copyright©1975 AmericanSocietyforMicrobiology Printed inVol. 16, No. 4U.S.A.
Analysis of
Expressions of the rII Gene Function
of
Bacteriophage T4
L. J. HEERE* AND J. D. KARAM
Department of Biochemistry,MedicalUniversity of South Carolina, Charleston,SouthCarolina29401
Received forpublication21 May 1975
Temperature-sensitive (ts)mutantsofthe T4 phage rIIgene were isolatedand
used intemperatureshiftexperiments that revealedtwodifferentexpressions for
the normal rII (rII+) gene function in vivo: (i) an early expression (O to 12min postinfection at30C) thatpreventsrestriction of T4 growthinEscherichia coli hostslysogenic forXphage, and (ii) alaterexpression (12to18minpostinfection
at30C) that results in restriction of T4 growth when the phage DNA ligase (gene 30) is missing. The earlier expression appeared to coincide with the period of synthesis of the protein product of the T4 rIlA cistron, whereas the later expression occurred afterrIIA protein synthesishadstopped. The synthesis of the protein product of therIIB cistron continues for several minutes after rIIA protein synthesisceases (O'Farrell andGold, 1973). ThetworII+geneexpressions might require different molar ratios of therIIA andrIlBproteins. It ispossible that the
separate expressions of rII+ gene function are manifestations of different
associations between thetworII proteinsand otherT4-inducedproteins thatare
synthesized oractivated at differenttimes after phage infection.
The rII gene ofbacteriophage T4 consists of two contiguous cistrons, rIIA and rIIB, the functions of which have not been explained.
Mutations in either cistron can result in a
number of effects on the physiology ofphage
growth in infected Escherichia coli hosts. Two types ofobservations suggest that the rII gene products play roles in T4 DNA metabolism. (i) T4rIImutantsdonotpropagate inE. coli hosts that are lysogenic for phage X (2). It appears that normal T4 rII (rII+) gene function is required to prevent the inhibition ofT4 DNA replication that is caused, at least in part, by the rex gene function ofX prophage (1, 10, 11, 22, 26). (ii) T4 rII mutants can replicate their DNA andproduce high yieldsofphagewithout the DNAligasecodedbyT4 gene 30(4,6, 12).It appears that rII mutations result in decreased levels of DNase activity in infected cells and that this leads to decreased dependence on
repairbyligase (13, 16, 23). T4 mutants that are defective in both gene 30 and the rII gene can utilize the hostDNA ligaseforgrowth (9).
Not all rII mutants showthese two types of effects. For example, some rII mutations can
rescue gene 30 defective phagewithout leading to restriction ofphage growth in X lysogens (4,
16).Also,somerIImutants are notrestrictedin X lysogens although they exhibit the
"rapid-lysis" phenotype that ischaracteristic ofthese mutants (5). In this communication we report
on evidence that differences in phenotype among different rIImutants may be caused by alterations in temporal expression of the rII gene productsin infected cells.
We isolated several temperature-sensitive
(ts)rIImutantsand usedthesein temperature-shift experiments that revealed two separable
expressions for the rII+genefunction in vivo. (i) Anearlyexpression (Oto 12minpostinfectionat 30 C)thatpreventsrestriction ofT4growthin X lysogens. This expression seemed to coincide with the time ofsynthesis ofthe rIIA protein. (ii) A later expression (12 to 18 min postinfec-tion at 30C) that resultsinrestriction ofphage
growth when the T4 gene 30 DNA ligase is missing. This expression seemed to require a higher rIlBtorIlA proteinratiothan the earlier expression. Thetwoseparateexpressionsof rII+ gene function could be shown to occur inde-pendently of each other and could be demon-strated by using ts mutations of either rII cistron. Differentexpressions of rII+ gene func-tion maybemanifestations of different associa-tions between thetworII proteins andbetween rII proteins and otherT4-inducedproteins that are synthesized or activated at different times afterphage infection.
MATERIALS AND METHODS
Bacterial andphage strains. E. coli K38strr and
BE arenonpermissive (su-) for T4 amber (am)
mu-974
on November 10, 2019 by guest
http://jvi.asm.org/
tants. E. coli KllOstrr and CR63 are permissive (su+3, tyrosine,andsu+1, serine, respectively). Some ofthe E. coli K-12derivatives used had been cured of
Aphageby heteroimmune superinfection with 21hyb2
phage and relysogenized with a wild-type strain of X(A++). The properties of these and other bacterial strainsused have beendescribed (14, 15).
Twodouble-am mutants ofT4gene 30wereused:
amXE30andamXC30.The genetic compositions and the growth properties of these mutants are
summa-rized inTable 1.TheT4 rIImutantsthat wereused forgenetic mappingexperiments,wereprovidedby S.
Champe and have beendescribed byBenzer (3). Media and growth conditions. The liquid and solidmedia thatwereusedinmostexperimentswere asdescribed previously (14, 15). Theeffects ofMg2+ on the rII mutantphenotypeswerestudied by using
the growth conditions described by Garen(8)with the following modifications. Cells were grownto 10' per
ml in 1% tryptone broth (Difco) containing 0.1 M NaCl and infected with phage at a multiplicity of infection(MOI)of 5at30C. Anti-T4serum(finalk=
10) was added at 5 min postinfection to remove
unadsorbed phage and after2minthe infected culture
wasdiluted 104-foldinto 0.2%tryptonebroth contain-ingeither 0.1 M MgCl2or0.1MNaCl. Phagegrowth
was continued until the diluted cultures were lysed with chloroform at 2.5 h after infection. Infective center titers (at 7 min postinfection) and average
phage yields per infected cell (burst size at 2.5 h postinfection) weredeterminedonE.coli CR63.
The media and methods thatwereusedforassays
ofT4DNA andprotein synthesis and for characteriza-tion of T4-induced proteins by sodium dodecyl sul-fate-polyacrylamide gel electrophoresis have been described (13, 14). [Methyl-3H]thymidine and l4C-labeled mixture of .-amino acids were purchased
fromNewEngland Nuclear Corp., Boston,Mass. Isolation oftrsllmutants.We searchedforT4rII mutations that permitted the growth of gene 30-defectivephageat42C, andrestrictedthisphageat 30C. StocksofamXE30 and amXC30were diluted
and plated on E. coli K110 strr and CR63, respec-tively.Theplaquesthat formed afterovernight incu-bation at30Cwereeach pickedand suspendedin 1 mlofM9S growthmediumcontaining afewdropsof chloroform. These plaque solutions represented our
random sample of double-am mutant phage. The
averagephagetiter for theplaquesolutionswasabout
5 x 107 PFU/ml. An appropriate amount of eachof the plaque solutions was then plated on E. coli K38strr and theplateswereincubatedat42C.Under thesesu-conditions tsrII mutations, aswellasother typesofrnI mutations, permit amXE30andamXC30 phage toform plaques. Theplaques which didform afterovernightincubationwerepickedandtested for growth on E. coli K38strr at 30 and 42 C. Isolates thatgrewat42C butnot at30ConK3&trr occurred
at an average frequency ofabout 5%. Thefirst such isolate tobe encountered from each of theamXE30
oramXC30 plaque solutions examined was purified
by replating on E. coli K38strrat 42 C and a high
titer phage stockwas subsequently prepared froma
sampleofasingle plaque thatwasgrownat30C on
E. coli KllOstrr for amXE30 plaques and on CR63 foramXC30 plaques. SomeoftheamXE30/tsrll iso-lates that were obtained were backcrossed to
wild-type phage and the single tsrII mutations were iso-lated. The single tsrII mutants were then crossed to amXC30 and the respective amXC30/tsrIl mutants were isolated. These genetic purification procedures werecarried out to remove secondary mutations that aroseduring the isolation of some of theamXE30/tsrII
mutants that we studied.
Genetic mapping of tsrII mutants. Many of the tsrII mutants we isolated failed to grow on X lysogens at42 C and were mapped on EHA (Enriched Hershey agar) platesby using the deletion mapping methods described by Benzer (3). Some of our tsrII mutants, however, were not restricted in X lysogens when grown onEHA agarplates at 42 C, but were restricted at this temperature on the lysogens in liquid media. So, it was possible to assign these mutants to the two rII cistrons by complementation tests that were carried out in liquidculture at 42C. These tests consisted of
coinfectingE. coliK38(X++)str'cells at anMOIof5 with each of the amXE30/tsrII under examination and r638 (a deletion covering the entirernIB cistron)
or rH88 (a partial rILA deletion mutation). Under theseconditions, complementation between the tsrIl
under examination and the rII deletion resulted in rII+ phenotype and in phage growth. Lack of com-plementation resulted in an rII- phenotype and in restriction of phage growth on the X lysogen. All of
amXE30/tsrII mutations we isolated complemented
one orthe otherrIldeletion used inthetests.
RESULTS
Genetic characterization of T4 tsrll
mu-tants. All of the T4 tsrll mutants that are
described here were isolated for theirabilityto grow in the absence of the T4 gene 30 DNA ligase at 42C, but not at 30 C. These mutants were also examined for growth properties in X
TABLE 1. Growth characteristicsof the T4 gene 30 amber mutants used
Growtha in response to
Mutant su su+1 su+3
30C 42C 30C 42C 30C 42C
amXE30(double _ - - +
-mutant:amH39X /amE605)
amXC30 (double - _ + - + +
mutant:amH39X
/amC104)
amH39X _ - + + + +
amE605 _ - + - + +
amC104 + + + +
a, No growth; +, growth in spot tests on EHA plates.
16,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.244.438.484.658.2]lysogenichosts. Table2summarizes the results that were obtained on a collection of tsrII mutants which were isolated in three different attempts at mutant isolation. The mutants could beclassified into threegroups onthebasis of differences in their growth responses in X lysogens. Group I mutants (Table 2) were re-strictedin X lysogens at 42C, butnot at30C, group II mutants were not restricted in A lysogens at either temperature, and group III mutants were restricted in X lysogens at both temperatures. The existence of the tsrII mu-tantsrepresented bygroupsII and III confirms that restriction in Xlysogens andrescueofgene 30-defective phage are two separable pheno-types of rII mutations (4, 16). It should be
emphasized that the results shown in Table 2
describe the growth characteristics ofthe tsrII mutants on agarplates. Insome cases,different growth responses wereobtained by using liquid
culture instead of agar plates. For example, in
M9SandinEHAliquid media, all thegroup II mutants we isolated were restricted in A lyso-gens at both 30 and 42 C. These effects ap-pearedtobe independent of thecistron inwhich the mutations occurred, and probably stem from some relationship between rII gene func-tion and cell membrane permeability of T4-infectedE.coli (7, 8, 18, 21, 25, 27).
Figure1shows the sites within the rII cistrons atwhich thetsrIImutationsfromexperiment 1, Table2werefoundto map.The results from the somewhat limited number of mutations that havebeen examinedsofarsuggestthe existence of "hot spots" for temperature sensitivity within the rII gene, especially within the rIIA cistron. Few tsrIIB mutations have sofarbeen
isolated and mapped, but it maybe
significant
tomention that all the group III mutations we have obtained mapped inrIIB. Group II
muta-TABLE 2. Growtha characteristics of isolated tsrII mutants
No. oftsrII Growthof
30-/tsrIIC
Growth of30+/tsrIICmutantsr
isolated phage on E. coli phage on E. coliGroup pergroup K-12 K-12(X) K-12 K-12(X)
Exp1 Exp2 Exp3 30 42 30 42 30 42 30 42
I 30 71 64 - + - - + + +
-II 10 45 105 - + - + + + + +
III 1 3 3 - + - - + + _ _
a-, Nogrowth; +,growth inspot tests on EHAplates.
IThe mutants for exp1wereobtained from stocks of amXE30; those for exp 2 and exp 3 were from amXC30. c30-,amXC30on su- host; 30+,am+forgene30 or amgene 30phage mutanton su+host.
1
I
<
I
I*
la,
III9
a
-
iII
l\1IIgo.,I
i'
l
AlI A2 I As I A4 I AX AS
I
III I ICISTRON II
FIG. 1. Genetic map positions of some ofthe tsrIImutants that were isolated in this study. The map
positions ofthe30groupImutants(solid bars)andthegroupIIImutant(openbar) fromexperimentIofTable 2weredeterminedbydeletionmapping.Themutants atdeletionsegmentsA6a2a, A6cl,andBMalbmappedat twosites within eachofthesesegments.
8
7
6
I-z5 Ui.
0
f 4
3
8
7
6 5
4
3
2
I
silli941164,ag Booon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.119.411.451.625.2]T4 rIIGENE FUNCTION 977
tions (notshown in Fig. 1) mapped in both rII cistrons (seeMaterials and Methods).
Other evidence for separable expressions of T4rII+ gene function. It hasbeen demon-strated thatthe restrictionof T4rII mutants in X-lysogenic hosts can be reversed in the pres-enceof0.1M Mg2+ (8). The resultswe showin Table 3 confirm these observations, but also show thatMg2+ does not reverse theeffects of
rIlmutationsonthe growthof gene30-defective T4 phage. In the X lysogen, E. coli K38(X++)strr, the T4 rII deletion mutantr1605 failed to grow in 0.1 M NaCl medium but
exhibited a considerable degree of growth in medium containing0.1MMgCl2 inplace of the NaCl. This phenotypic reversal of rH- to rIl+ did not result in restriction of ligase-defective phage [amXC30/r1605 on E. coli K38(X++)strl
and K38strr hosts, Table 3]. These results suggest that the two phenotypes of rII muta-tions which we examined are due to different postinfection metabolicevents.
Time of expression of T4rIIgenefunction. Weused the tsrII andamXC30/tsrIIderivatives that we isolated (Table 2and Fig. 1) in alarge
number oftemperature-shift experiments that wereintendedto measurethetime of expression of the rII gene in infected cells. Mutants of group I could be usedto measure theeffects of
[image:4.493.244.433.347.523.2]shuttingoffrII+genefunction onboth, restric-tion in X lysogens, and survival of amXC30 under su- conditions. Mutants ofgroup IIand III were only informative with regards to the effects on amXC30. Qualitatively similar re-sults wereobtained among mutants of any one of the three groups. The quantitative differ-ences wedidobserve within groups were due to the different degrees of leakiness exhibited by
TABLE 3. Effect of magnesiumon the T4 rIImutant
phenotypes
Burst size in medium containing Phage strain E. coli host
0.1M 0.1M Na+ Mg2+
T4D2+ K38strr 42 31
amXC30 K38strr <0.5 0.36
amXC30/r1605a K38strr 28 46
r1605 K38strr 36 35
T4D2+ K38(X+ +)strr 15 31
amXC30 K38(X++)strr 0.36 0.36 amXC30/r1605 K38(X++)strr <0.36 10 r1605 K38(X++)strr <0.36 30
ar1605is anrIIgene deletionthat covers portions of
bothrIIcistrons (3).
thedifferent mutants. Thatis, some mutations were more temperature-sensitive than others. Weemphasize here the qualitativeaspectsofT4 rII+ gene expression.
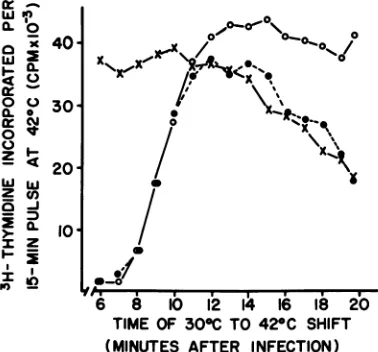
The experiment described in Fig. 2 demon-strates typical responses that were obtained withgroupI mutants. Inthisexperiment oneof theamXC30/tsrII isolates from groupI and the corresponding single tsrII mutant were used to infect E. coli K38strr and E. coli K38(X++)strr cultures at 30C, and at various times after infection the cultures were diluted in growth medium at 42 C (to inactivate the rII+ func-tion). The rates of thymidine incorporation afterthetemperature-shiftwerethen measured. The results (Fig. 2) demonstrated thefollowing properties for the rII+ gene function under the experimental conditions used: (i) rII+ gene function is not required for growth in the X lysogen after 12 min postinfection [tsrll-infected K38(X++)strr curve, Fig. 2], (ii) rII+ expression before the 12th min postinfection does not lead to restriction ofamXC30 (curve for amXC30/tsrII-infected K38strr, Fig. 2), and
0:
-o40I
o0
C0 /
a rO 401 < <o-o /
,_
, Ix
ZXJi.
o 0. 30.
IJW *
Z
q)
IzX 10
/
a ~of
6 8 10 12 14 16 18 20 TIME Of 30°C TO 42°C SHIFT
(MbINUTES AFTER INFECTION ) FIG. 2. Time of expression of the T4 rII+ gene. Log-phase E. coli cells (eitherK38strrorK38(A\++)strr) at 108 per ml were infected with the phage under examination at MOI 10 in M9S at 30 C. At various times postinfection, samples of infected culture were diluted 10-fold in M9S at 42 C.Fifteen minutes after the temperature shift, 1.61i Ci of (methyl-'H]thymi-dine (at 2.5MlCiof3lI/ygof thymidine) were added per ml of heated culture and thymidine incorporation was permitted to occur for an additional 15 min before it was stopped with 5 ml of cold 10% trichloroacetic acid per ml of incorporating culture. Symbols: (0)
amXC30/tsrII-infected K38(XA++)strr; (0) tsrII-infected K38(AX+ +)str'; (X) amXC30/tsrII-infected
K38strr. The tsrIImutation used for this experiment mapped in T4rIIAsegmentA6a2a.
16,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.36.230.493.652.2]978 HEERE AND KARAM
(iii) inrII+-infected cells, theT4 gene 30ligase is required for phagegrowth after the 12th min postinfection [compare tsrII-infected
K38(X++)-strr and amXC30/tsrIl-infected K38(X++)strr]. Similar results were obtained by using M9 mediumthatwas notsupplemented with amino acids and E. coli B, instead of E. coli K-12
hosts (data not shown). For example, at 30 C and in M9 (instead of M9S) medium, an
amXC30/tsrll-infectedE. coli B(X) culturegave lower levels of thymidine incorporation than was obtained in theexperiment forFig. 2, but the timing ofrII+ expression was the same for thetwo experiments.
In other experiments, such as the one
de-scribedinFig. 3, wedetermined thatthymidine incorporation was linear during the period of pulse labeling thatwasusedfortheexperiment inFig. 2. Cumulative DNA synthesiswas mea-sured in
amXC30/tsrII-infected
K38(X++)strrcells. When these infected cells were shiftedto 42 C, there was an immediate effect on the amount of DNA which was subsequently syn-thesized. The accumulation oflabel into DNA
40 A12min
/v 14 min f ol6 min
o.1 Vo 0min
0
30-Owvo /KI18min
A
X20min 0~~~~~~~~
0
w
z x8min
io-6min
TIME (MIN) AFTER INFECTION
FIG. 3. Cumulative DNA synthesis in amXC30/ tsrII-infectedE.coliK38(X ++)strr. Log phasecells(at 108perml)wereinfectedatMOIof10 inM9Sat30C. At various times postinfection samples of infected culture were diluted 10-fold into [methyl-Hjthymi-dine(20 ,uCi of3H/ml;27,Ciof3H//g of thymidine) inM9Sat42C. The timesoftemperature shiftsare
indicated on the thymidine incorporation curves.
Samples (0.05 ml)werewithdrawnfromeach diluted
cultureat10-minintervalsafterdilution intoisotope
at 42C and were assayed for trichloroacetic
acid-insoluble radioactivity. The tsrII mutation used for thisexperiment mappedin T4rIIA segmentA6a2a.
was linearfor atleast 50min. Therateof DNA synthesis (slopes of curves in Fig. 3) depended onthe timeoftemperature-shift suggesting that there is a directrelationshipbetweentheextent of rII+ expression and (i) the degree of restric-tion on a X lysogen and (ii) the degree of restriction ofligase-defective phage.
Time of synthesis of rII proteins. It has been shown thatinT4-infected E. coli, the rIIB protein is translated from two mRNA species that are the result of two different modes of transcription (20, 24). We compared the ki-neticsofsynthesis of therIIAandrIIBproteins inanattempt tocorrelate thetwoexpressions of rII+ with the relative dosage of the two rII proteins.The levelsofsynthesesofthe rIIA and rIlB proteins in amXC30-infected E. coli BE cellsweremeasuredby using gel electrophoretic assays, the resultsofwhich areshowninFig.4. These results (Fig. 4) confirm the observation made by O'Farrell and Gold (20) that, in T4-infected cells, the rIIB protein continuesto be synthesized for several minutes after rIIA protein synthesis has stopped. Under the growthconditions used inour study,the period ofsynthesis of the rIIA protein (Fig. 4) seemed to coincide with the period of rII+ gene expres-sionthat isrequired topreventrestrictionofT4 growthin Xlysogens (Fig. 2).Soit appearsthat the two expressions of the rII+ gene function occur atdifferent rnIB torIIAprotein ratios.
DISCUSSION
Many different effects of T4 rII mutations havebeen described (2,4, 12, 17, 18, 23,25), but ithasnotbeen possibletorelate allthese effects to one metabolic deficiency. The restriction of T4 rII mutants in X-lysogenic E. coli hosts appears to be due to metabolic events that occur during the firstfew minutes after phage infection. This has been determined in two types of study. (i) Certain concentrations of
Mg2+(8)reversethe restrictionofrIImutants in X lysogens only when added before 10 min postinfection at 37 C. (ii) Inactivation of ts T4 rII gene function prior to 10 to 20 min post-infection (at 30C) results in restriction ofphage growth in X lysogens (19). Our studies confirm these observations and extend themby showing
that a different T4 rII+ gene
expression
is involved in the restrictionofDNAligase
(gene30)-defectivephage.
Inthestudy thatwasdescribedinthis report we examined the relationship between two rII mutant phenotypes to each other and to the patterns ofsynthesisofthephagerIIA and rIIB proteins in vivo. Our experiments defined two
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.63.255.339.550.2]T4 rIIGENEFUNCTION 979
_- -
__
- i-. ...._0 ~ ~
C - -_ n _ . n
o e 1 co fla.
C-~~~~~~~~~~~~~~~~~C
---
>ae
*0 XNco 0 2 * 0 N
0 aa7 7 (N
(N0
(NTimem) after i-nf Time (min) after infection
WIA
co
z -s
0 = CL IL-0 0
_ l
CU)
0 a)
ux
5 7 9 11 13 15 17 19 21 23
TIME (MIN)AFTER INFECTION
FIG. 4. Patterns of rIIA andrIIB protein synthesis in T4amXC30-infectedE. coliBE. Cellsgrown in M9 medium toadensity of2x 108 mlwereinfectedatMOIof10withamXC30at30 C. At varioustimeintervals postinfection, 1-ml samples of infected culture werepulse-labeled with 1 UCi of "4C-labeled amino acids. Extracts of the isotopically labeledcellswere thenpreparedandanalyzed bysodium dodecyl sulfate-polyac-rylamide gel electrophoreticassays byusingthe methodspreviouslydescribed(13). Thefigureshowsapicture
ofanautoradiogramofa7.5%gelandthe resultsofdensitometrictracingsthatwerecarriedoutontherIIAand rIIB bands in the autoradiogram. The "amount of protein synthesized per pulse" refers to the weight (milligrams) oftherIIAandrIIB bands thatwererecordedonchartpaperinthe densitometricscanning ofthe autoradiogram.
separate expressions ofthe T4 rII+ gene func-tion that are not dependent on each other. In M9S medium at 30C, the rII+ gene function that is required toprevent restriction of T4 in E. coli strains lysogenic for X phage was ex-pressed before the 12th min postinfection.
Under the same conditions, the expression of rII+ gene function that results in restriction of DNA ligase-deficient phage occurred predomi-nantly afterthe 12th minpostinfection.
The pleiotropic effects of T4 rII mutations may be manifestations of the participation of
VOL. 16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.91.387.79.509.2]980 HEERE AND KARAM
rIIA and rIIB proteins in several different phage-induced metabolic processes. It is likely
thatboth therIIA and the rIIB proteins
partici-pate in the same metabolic processes since all therIImutantphenotypes that havesofarbeen
discovered can be exhibited by mutations in
either rII cistron. The relative contributions of the two proteins in these processes, however,
may differ andmaybe controlled bythe differ-entmodes ofsynthesisof theseproteinsinvivo.
The two effects of rII+ gene function that we
examined probably arise from two separate
metabolic events since (i) notallrII mutations affect both rII+ gene expressions (4, 16; Table 2) and (ii) a change in physiological
condi-tions, i.e., the addition of 0.1 M Mg2+ to T4 rII-infectedcells,affectedonerIImutant
pheno-type (restriction in X lysogens) but not the
other (growth of gene 30-defective phage)
(Table 3). The molar ratioof the rIIA andrIIB proteins changes progressively during early times after phage infection (20; Fig. 4). It is
interestingto notethatthe time ofsynthesisof
the rIIA proteins seems to parallel the rII+ expression that prevents restriction of T4
growth inX lysogens,whereas the restriction of
DNA ligase-defective phage occurs at a later
time, duringaperiod ofcontinuedrIlBprotein
synthesis. Conceivably, the two expressions of
rII+ gene function that we examined require
differentrlIBtorIIA molar ratios.
It is possible to explain the two separate expressionsofrII+gene functiononthe basis of
both (i) the interaction of rIlA proteins with
rIlB proteins, and (ii) the interaction of rII proteins with otherT4-induced proteins.It has
been shown thatboththerIlAandrIlBproteins
are membrane bound (7, 21, 27), but no free
rIIA-rIIB protein complexes have yet been
iso-lated. Since many of the reactions that are
involved in macromolecular biosynthesis in
'14-infected cells are thought to be membrane
associated, it can be postulated that the rII proteins exert their effects on these processes
either directly by acting as anenzyme (e.g., a
controlled endonuclease [4, 12, 23]thatactson
membrane-bound DNA) or indirectly by
in-fluencing the activities of various components inmembrane-protein complexes.
ACKNOWLEDGMENTS
We thank B. Baggett, L. Chao, B. Ledford, and J. Speyerfordiscussions andcommentsonthe manuscript.
This work was supported by a Public Health Service
grant(GM18842)andaResearchCareer Development Award
(GM-70-725) to J. D. K. from the National Institute of General Medical Sciences, and by the South Carolina AppropriationforBiomedical Research.
LITERATURE CITED
1. Astrachan, L., and J. F. Miller. 1972. Regulation of Xrex expression after infection of Escherichia coli K by lambda bacteriophage. J. Virol. 9:510-518.
2. Benzer, S. 1955. Fine structure of a genetic region in bacteriophage. Proc. Natl. Acad. Sci. U.S.A. 41:344-354.
3. Benzer, S. 1961. On the topography of the genetic fine structure.Proc.Natl. Acad. Sci.U.S.A.47:403-415. 4. Berger, H.,and A. W.Kozinski.1969.SuppressionofT4D
ligase mutations by rIIA and rIlB mutations. Proc. Natl.Acad. Sci.U.S.A. 64:897-904.
5. Crick, F. H. C., L. Barnett, S. Brenner, and R. J. Watts-Tobin. 1961. General nature of the genetic code forproteins. Nature (London) 192:1227-1232. 6. Ebisuzaki, K., and L. Campbell. 1969. On the role of
ligase in genetic recombination in bacteriophage T4. Virology38:701-702.
7. Enis, H. L., and K. D. Kievitt. 1973. Association of the rIIA protein with the bacterial membrane. Proc. Natl. Acad.Sci. U.S.A. 70:1468-1472.
8. Garen, A. 1961. Physiological effects of rII mutations in bacteriophage T4. Virology 14:151-163.
9. Gellert, M., and M. L. Bullock. 1970. DNA ligase mutants of Escherichia coli. Proc. Natl. Acad. Sci. U.S.A.67:1580-1587.
10. Gussin, G. N., and V. Peterson. 1972. Isolation and properties of rex- mutants of bacteriophage lambda. J. Virol. 10:760-765.
11. Howard, B. D. 1967. Phage lambda mutants deficient in rIIexclusion.Science 158:1588-1589.
12. Karam, J. D. 1969. DNA replication by phage T4rII mutantswithout polynucleotide ligase (gene 30). Bio-chem.Biophys.Res. Commun.37:416-422.
13. Karam, J. D., and B. Barker. 1971. Properties of bacterio-phageT4mutantsdefective in gene 30 (deoxyribonu-cleic acidligase)andthe rII gene. J. Virol. 7:260-266. 14. Karam, J. D., and M. G. Bowles. 1974. Mutation to overproduction ofbacteriophage T4 gene products. J. Virol. 13:428-438.
15. Karam, J.D.,and P. V. O'Donnell. 1973.Suppressionof amber mutations of bacteriophage T4 gene 43 (DNA polymerase) by translational ambiguity. J. Virol. 11:933-945.
16. Krisch, H. M., D. B. Shah, and H. Berger. 1971. Replication and recombination in ligase-deficient rII bacteriophageT4D. J. Virol. 7:491-498.
17. Krylov, V. N. 1970. Possible function of rII genes of bacteriophage T4. Virology 41:711-717.
18. Maor, G., andC. Shalitin. 1974. Competence of mem-brane-bound T4rII DNA for in vitro "late" mRNA transcription. Virology62:500-511.
19. Mattson, T.,and M.Russel. 1972. The time ofsynthesis and functionofthe rII+ productofbacteriophageT4. Virology 49:160-167.
20. O'Farrell, P. Z., and L. M. Gold. 1973.BacteriophageT4 geneexpression. Evidencefor twoclassesof prereplica-tive cistrons.J.Biol.Chem. 248:5502-5511.
21. Peterson, R. F., K. D. Kievitt, and H. L. Ennis. 1972. Membrane protein synthesis after infection of Esche-richia coli B with phage T4: therUBprotein. Virology 50:520-527.
22. Ravin, V. K., and G N. Poluchina. 1968. Mutants of phage X unable to exclude phage T4 rII. Virology 35:616-617.
23. Rutberg, B., and L. Rutberg. 1966. Bacteriophage-induced functions in Escherichia coli K(X) infected with rII mutants of bacteriophage T4. J. Bacteriol. 91:75-80.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
24. Schmidt, D. A., A. J. Mazaitis, T. Kasai, and E. K. F. Bautz. 1970.Involvement ofaphageT4afactor andan
anti-terminator protein in the transcription of early T4
genesinvivo.Nature(London)225:1012-1016. 25. Sekiguchi,M. 1966.Studiesonthephysiological defectin
rII mutants of bacteriophage T4. J. Mol. Biol.
16:503-522.
26. Tomizawa, J-I., and T. Ogawa.1971.Inhibition of growth of rlI mutants of bacteriophage T4 by immunity substance of bacteriophage lambda. J. Mol. Biol. 23:277-280.
27. Weintraub, S. B.,and F. R.Frankel. 1972. Indentifica-tion of the T4 rlIB gene product as a membrane protein.J. Mol. Biol. 70:589-615.
VOL.16, 1975
on November 10, 2019 by guest
http://jvi.asm.org/