Vol.65,No. 6 JOURNAL VIROLOGY, June 1991,p.3007-3014
0022-538X/91/063007-08$02.00/0
CopyrightC 1991, American Society for Microbiology
Expression of Active
Human
Immunodeficiency
Virus
Type
1Protease
by
Noninfectious Chimeric
Virus Particles
NANCYE.
KOHL,'*
RONALD E. DIEHL,' ELAINE RANDS,' LENORA J. DAVIS,' MICHELLE G.HANOBIK,1BOHDANWOLANSKI,2 AND RICHARD A. F. DIXON'
Departments ofMolecular Biology' and Virus and Cell Biology,2 Merck Sharp and Dohme Research Laboratories, West
Point,
Pennsylvania
19486Received 24October 1990/Accepted5 March 1991
To generate nonpathogenic viral particles which express active human immunodeficiency virus type 1 (HIV-1) protease (PR), plasmids containing sequences from the genomes of HIV-1 and Moloney murine leukemia virus (M-MuLV) were constructed. Either the PR coding region alone; thegag, PR, and reverse transcriptase protein-coding regions; or the completegag andpol protein-coding regions from HIV-1 were substituted for the corresponding regions of a full-length M-MuLV clone to yield the chimeric plasmids pMoHIV-I, pMoHIV-III, and pMoHIV-IV, respectively. Cell lines which expressthe viral gag polyprotein wereisolated for hybrids pMoHIV-I and pMoHIV-III. These cells produced viral particles which contained processedcoreproteins. Cleavage of thegagpolyproteinintheviralparticleswas inhibited by the HIV-1 PR inhibitorL-687908, indicatingthat the viral PR is responsible for the observed processing. The hybrid virions were not infectious; analyses indicated that the viral particles contained little or no reverse transcriptase activity.Inaddition,particlesproduced by pMoHIV-III transfectants failedtopackagetheviral genomicRNA. The cell linewhichexpressesandprocessestheHIV-1gagpolyprotein isasafe andeffectivereagentforthein vivoevaluation ofpotentialinhibitors of the HIV-1 PR.
The human immunodeficiency virus type 1 (HIV-1) is a member of the lentivirus subfamily of retroviruses. The HIV-1 genome consists ofthe three majorretroviral genes: gag, which encodes the viral structural proteins MA, CA, NC, and p6; pol,which encodestheviral enzymes protease
(PR), reverse transcriptase (RT), and integrase (IN); and env,which encodes the viralglycoproteinsSU and TM (20). Inaddition,theHIV-1genomeencodes numerous regulatory
proteins (Rev, Tat,Nef, Vif, Vpr, and Vpu [20]).
Typical ofallretroviruses, HIV-1initially synthesizes the
structuralproteinsand enzymes in the formoflarge precur-sorpolyproteins, whicharecleavedby the viral PR toyield
the matureproteins. Biochemical and structural characteri-zation ofHIV-1 PRaswell asanalysis ofmutantformsof the
protein indicatethat the enzyme isamember of theaspartic
acidfamilyof proteases(9,23, 38). HIV-1 PR isanessential
viral enzyme (25, 32) and therefore an attractive target for
antiviral therapy. Indeed, several potent inhibitors ofthe enzymehaverecentlybeen reportedwhich exhibit
antipro-tease activityin vitro and prevent the spread ofthevirus in
susceptible cells in culture(10, 29, 30, 33).
The intense interest in the development ofinhibitors of HIV-1 PRhas created a needfor safe andeffective in vitro and invivo systemsin which toevaluate such compounds.
While inhibition of processing of the gag polyprotein in
HIV-1-infectedcells in culture has been usedextensivelyto evaluate thespecificity and potency ofHIV-1 PRinhibitors (29, 30, 33), the useof livevirus in the assay posespotential
risks. The successful use of monkey cells infected with a recombinant vacciniavirus which expresses and processes the HIV-1 gag-polprecursor for the evaluation of
antipro-tease activity has recently been described (29). While this assay eliminates the riskof infectionbyHIV-1,precautions
mustbe taken toavoid infection with vaccinia virus.
* Corresponding author.
In an attempt to produce nonpathogenic viral particles
which express active HIV-1 PR, we haveconstructedhybrid
clonesbetweenMoloneymurine leukemiavirus(M-MuLV)
and HIV-1. Following transfection ofthese DNAsinto NIH 3T3 cells, stable cell lines wereisolated whichrelease viral
particles which express and process the gag polyprotein.
Theseparticleswere notinfectious;all lackeddetectableRT
activity. Inaddition, particles produced bycellsexpressing
one ofthe hybrid DNAs failed to package viral
genomic
RNA. Proteolytic processing in the particles was inhibited byaknown HIV-1 PRinhibitor. Thus, this systemprovides
a safe andeffectivemethod for the evaluation ofanti-HIV-1 PR activity.
MATERIALS ANDMETHODS
Cells and viruses. NIH3T3 cells andXC cellsweregrown in Dulbecco's modified Eagle's medium (Mediatech, Inc.) supplemented with 10% fetal bovine serum (Armour).
M-MuLV releasedby NIH3T3 cells after transfectionwith
plasmidclone pNCA(6) wasthe source ofwild-type virus. Viralinfectivitywasdeterminedby the XCplaqueassay(34)
orby infection of fresh NIH 3T3 cells. Virus infectionswere
carried out in the presence of 8 ,ug of Polybrene per ml. Stableproducers ofhybridviralparticleswere
generated
by cotransfection (41) of NIH 3T3 cells with a mixture of chimeric plasmidDNA and pSV2neo DNA in a 10:1 or5:1ratio, followed by selection in G418 (GIBCO) at 2 mg/ml.
Producer cell lines weremaintained in G418at 1 mg/ml.
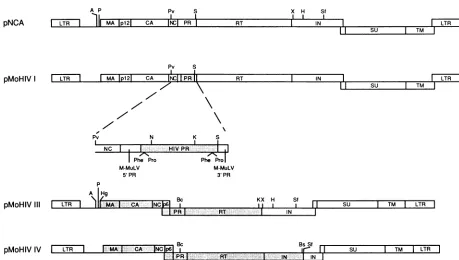
Construction of M-MuLV/HIV-l chimeric plasmids. The
infectious, full-length M-MuLV clone pNCA (6),
kindly
supplied by S. Goff, was used as a substrate for the con-struction of the chimeric
plasmids.
InpMoHIV-I,thecoding
regionforHIV-1PRreplacesthatfor the viral PRof
pNCA.
HIV sequences immediately N terminal and C terminal to the 5' and 3' PRcleavagesites,
respectively,
wereincluded to facilitate autocatalytic cleavage of the PR from thepol3007
on November 10, 2019 by guest
http://jvi.asm.org/
3008 KOHL ET AL.
A P Pv S
'II I I
7
Pv
7
N K S
I I
NC ...P...P....R .:..
Phe Pro M-MuLV
5PR P
Phe Prol M-MuLV
3PR
BC Bs~~~~~~~~~Sf
I
LTR
I A
MIN
PRlEti:Et
lEL: RT!VESA iiE iE le III
r
SUI
T I
LTR
I
I
LTRIM
CA
NCp6
1
PR;
SU
TMI
LTRnr
1
IN
IFIG. 1. Structure of M-MuLV/HIV-1 chimeric plasmids. The chimeric plasmids were constructed by replacing sequences from the
infectious, full-length M-MuLV clone pNCA, shownatthetop,withcorrespondingHIV-1sequences,asdescribed in Materials and Methods.
Open boxes, M-MuLV sequences; shadedboxes, HIV-1 sequences. Restriction enzymes: A, AatlI; Bc,Bcll; Bs, BsmI; H,Hindlll; Hg, HgaI; K, KpnI; N, NotI; P, PstI; Pv, PvuI;S, Sacl; Sf,Sfil;X, XmnI. Only sites usedinthe construction of the plasmidsareindicated.
precursor protein. pMoHIV-I was constructed by isolating
the NotI-KpnI fragment spanning the coding region for amino acids 9 to 73 of HIV-1 PR from syn7, a bacterial expression plasmid containing the chemically synthesized HIV-1 PR gene from the NY5 isolate (24a). A synthetic oligodeoxynucleotide was ligatedto the 5' end of this frag-ment to encode M-MuLV sequences from the PvuI site at the 3' end ofgag (residues 2117 to 2122; numbering as in reference 36) (Fig. 1) through the active-site aspartic acid residue of the PR(residues 2317to2319), the sixamino acid residues from the HIV-1 polgene N terminal to the 5' PR cleavage site,andtheeightN-terminal residues of HIV-1 PR and to regenerate a NotI site. Another synthetic oligode-oxynucleotide wasligated to the 3' end of thisfragment to regenerateaKpnI siteandtoencodeaminoacid residues73 to99 of HIV-1PR, sevenadditional residues of HIV-1polC terminaltothe3' PRcleavagesite andtoaddaSacl site. The resulting fragment was used to replace the sequence of pNCA between the PvuI site atthe 3' end ofgag and the SacIsiteatthe 3' of PR(Fig. 1).
InpMoHIV-III,theentiregag,PR, andRTcoding region ofpNCA wasreplaced withthe corresponding region from HIV-1. For construction of pMoHIV-III, the 4.5-kb AatII-HindIII fragmentfrompNCAwassubclonedinpUC19.The resultingDNAwas digested withPstI andXmnI toremove the M-MuLV gag, PR, and RT coding regions (Fig. 1). Oligodeoxynucleotides encoding M-MuLV sequences be-tweenthePstI site and thegaginitiation codon, unique ClaI
andKpnI sites,HIV RTsequences3'of the 3'KpnIsite,and M-MuLV IN sequences 5' of the XmnI sitewere inserted. The DNA was digested with ClaI, treated with mungbean nuclease, digested withKpnI, andligatedtothe4-kb HgaI-KpnI fragment from the full-length HIV-1clonepNL4-3 (1),
spanning gag, PR, and most of RT. The resulting AatII-HindIII fragmentwas then reinserted intopNCA.
In pMoHIV-IV, the entire gag and pol coding regions fromHIV-1replacethecorresponding regionsof M-MuLV. The 3' M-MuLV IN sequence, containing the env splice
acceptor and the 5' coding region ofenv, was retained to facilitate synthesis of the envelope proteins. For construc-tion of pMoHIV-IV, the BclI-BspMI fragment ofpNL4-3 spanning the3' end ofPR, HIV-1RT, and most of IN was isolated. Asyntheticoligodeoxynucleotidewasligatedtothe 3' end of thisfragmenttoregenerateaBspMI site,toencode the remainder of the HIV IN sequence, and toadd an
Sfil
site. The resulting fragment was used to replace these-quenceinpMoHIV-ITT betweentheunique BclIand
Sfil
sites(Fig. 1).
Analysisof viralproteins. Monolayers of transformants at 50to 100% confluence in 10-cm dishes were washed twice with 5 ml of cold phosphate-buffered saline (PBS) and scraped into 1.2 ml of cold PBS. Cells were collected by centrifugation, resuspended in sample buffer(0.125 M Tris-chloride [pH 6.8], 4% sodium dodecyl sulfate [SDS], 20% glycerol, 1.4 M 3-mercaptoethanol) andheated at100°Cfor 3 to 5 min. Proteins were separated on 12.5% denaturing polyacrylamide gels and electroblotted onto Immobilon membranes (Millipore). Membranes were incubated with anti-HIV antibody-positive antiserum(1:500 [25])for detec-tion of HIV-1-specific proteins or with goat anti-Rauscher p30 serum (1:250; NCI 77S-174) for detection of M-MuLV capsid protein. Immunoreactive proteins were detected by autoradiographyafter incubation with1251I-labeledproteinA (Amersham). For analysis of proteins in viral particles, particles were pelleted from clarified culture supernatant through a glycerol cushion (35), resuspended in sample pNCA
pMoHIV I
X H Sf
I l
pMoHIVIII
pMoHIVIV
I I I
I LTR
I
ih MA IP121 CAI.Ej
PR I RT I INSu I
Pv s
:1I
I LTR
I
I MAIP121
CA INCI [RR: RTE--
IN I LTRI
11-11'
I Su TM I1%
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.80.539.69.329.2]buffer, boiled, and analyzed by polyacrylamide gel electro-phoresis (PAGE) and immunoblotting as described above.
Electron microscopy. Cell monolayers were fixed in 1% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, washed in cacodylate buffer, postfixed in 1% osmium tetroxide, and dehydrated in an ethanol series (50 to 100%). The cells were then scraped from the dishes, pelleted, and infiltrated in Spurr's resin (Polysciences, Inc.)-ethanol (1:1) for 1 h, placed in fresh Spurr's resin at 4°C overnight, transferred to freshresin inBeemcapsules, and polymerized at 70°C for 48 -h. Ultrathin sections were cut, stained with methanolic uranyl acetate and leadcitrate, and subsequently examined
in aPhilips 300transmission electron microscope.
In vitro HIV-1 PR processing assay. Either 1 x 106 MoHIV-I-33, 5 x 105MoHIV-III-4A, or 1 x 105 pNCA-20 cells were seeded in medium without G418 and allowed to attach. Protease inhibitor in dimethyl sulfoxide (DMSO) was added at theindicated concentration; the final concentration ofDMSO was 0.1%. After 24 h at 37°C, the cultures were washed three times with 5 ml of medium, and fresh medium
containing inhibitorwasadded. Followinganadditional 24-h
incubationat37°C, the medium was harvested. Virions were
prepared from clarified supernatant and analyzed by
SDS-PAGE andimmunoblotting asdescribed above.
RT assay. Viral particles were pelleted from clarified culture medium collected from producer cells in 10-cm
dishesat50 to100%confluence. RT activity was determined in a standard assay mix containing
Mg2+
(14). Quantitative resultswereobtainedbycounting radioactivity in the DEAE papertowhichthe DNAproduct was bound in ascintillation counter. RT activity associated with the MoHIV-III parti-cles wasalso assayed in the presence ofMn2+
asdescribed before (15).Analysis of viral RNA. Preparation of total cellularRNA
from transfected cells, fractionation through formaldehyde-agarose gels, and transfer to nitrocellulose were done as
described before (27). Viral RNA was detected by hybrid-ization with a DNA fragment spanning the M-MuLV long
terminalrepeat (LTR)radiolabeled by therandom priming
method (Boehringer). For preparation of viral packaged
RNA, culture mediumwascollected twice over aperiod of 24 h from monolayers of transformants at 75 to 100% confluence in 15-cm dishes. RNA was purified from virus
pelleted bycentrifugation essentiallyasdescribedbyBender etal. (4)and analyzed asdescribed above.
RESULTS
Construction of M-MuLV/HIV-1 chimeric plasmids. The
similarityin genome structureamongretrovirusessuggested that it might be possible to generate nonpathogenic viral
particles which express active HIV-1 PR by creating
chi-meric viruses. The murine ecotropic type C retrovirus
M-MuLV provides an ideal system for the construction of
such chimeras. M-MuLV can be passaged in vivo and
replicates in culture. In addition, the virus has been well characterized at the molecular and biochemical levels, and
infectious, full-length molecular clones havebeenisolated. Asafirst step ingeneratingsuchchimericviruses,aseries ofhybridcloneswereconstructed inwhich HIV-1 sequences were substituted for the corresponding regions of the
full-length M-MuLV clone pNCA. In pMoHIV-I, the coding regionfor thePRfromHIV-1replacesmostof theM-MuLV PRcoding region (Fig. 1). Althoughthe viral PRfrom both HIV-1 and M-MuLV catalyzes its release from thegag-pol
precursor protein, autocatalytic cleavage ofHIV-1 PR
oc-curs at Phe-Propeptide bondsflanking themature enzyme, whereascleavage ofM-MuLVPRoccurs atLeu-Thrbonds. Toanticipatethepossibilitythat HIV-1 PRmightnotcleave the Leu-Pro bond which would result from the precise
substitution of the HIV-1 PR coding region for that of M-MuLV, sequencesencodingthe sixamino acids N-termi-nal to Pro-1 and theseven amino acids C terminal to Phe-99
ofthematureHIV-1 PR wereincludedin theclone.
Synthe-sis of the gag-pol polyprotein occurs by a translational frameshift in theregionof the gag andpol overlapin HIV-1
(21) and by in-frame suppression ofa terminationcodon in M-MuLV (42). Since sequences C terminal tothe
termina-tion codon at the gag-pol junction in M-MuLV may be
required for synthesis of the viral enzymes, sequences
encoding the N-terminal 32 amino acids of the mature M-MuLV PRwereretained in the chimera.
Despite identification ofall ofthe HIV-1 PR gag andpol cleavage sites, the exact requirements for a PR substrate havenotbeenpreciselydefined. Consequently,itis difficult topredictwhether the HIV-1 PR iscapable ofmakingall of the cleavages in the M-MuLV gag and pol
polyproteins
whicharenecessaryfortheproductionof infectious virions.
Therefore,twoadditional chimeric cloneswereconstructed: pMoHIV-III, which contains the gag, PR, and RT coding regions of HIV-1 in place ofthe corresponding regions of M-MuLV;andpMoHIV-IV, which containsallofthe HIV-1 gag andpol coding regions (Fig. 1). In order to retain the
splice acceptor used to generate the M-MuLV env
tran-script,
sequencesencoding
the C-terminalportion
of M-MuLV IN which contain the splice acceptor were in-cluded inpMoHIV-IV.Toestablish stable cell linesexpressing pMoHIV-I, pMo-HIV-III, orpMoHIV-IV,eachof the DNAswas
separately
cotransfected into NIH 3T3 cells together with a gene
conferring resistance to the antibioticG418. G418-resistant
colonies were selected andexpanded to mass culture. Analysis of viralproteins. Totestfor viral geneexpression
in the transfected cell lines, cell lysates were
analyzed
by SDS-PAGE andimmunoblotting.
Goat anti-Rauscher p30serum was used to detect the M-MuLV CA
(p30)
and its precursor, p65, expected to be expressed in pMoHIV-Icotransfectants. As shown in
Fig.
2A,whereasno immuno-reactiveproteinsweredetected in the lysatesof cells trans-fected with theneomycin
resistancegenealone(lane 1), p65,
CA, and processing intermediates were detected in cells transfected withpMoHIV-I (8of 13 lines
tested;
2areshown in lanes 3 and5)and in cellstransfected withpNCA
(lane 7).
The presence ofCA in the
pMoHIV-I
celllysates
suggests that HIV-1 PR is produced in these cells andiscapable
ofprocessingthe M-MuLVgagprecursor
protein.
Anti-HIV
antibody-positive
antiserum, which ishighly
reactive with the HIV-1 CA(p24)and its precursor,
p55,
was usedto testfor the presence of intracellular viralproteins
in cells transfected withpMoHIV-III
andpMoHIV-IV.
pS5was detected in the celllysatesof 13 of the 14pMoHIV-III
transfectants examined
(data
notshown).
While low levels ofp55 processing
intermediatesweredetected in many of theselines, a
significant
level of CA was seen inonly
oneline,
MoHIV-III-4A(Fig.
2B,lane3).
FourpMoHIV-IV
transfec-tants were analyzed for the presence of intracellular viralproteins.
Althoughp55
was detected in twoof theselines,
CA was not present in
either,
indicating
that the gag precursorwas notbeing processed.
ThepMoHIV-IV
trans-fectantswere notanalyzed
further.Todetermine whether viral
particles
wereproduced by
thepMoHIV-I
orpMoHIV-III
transfectants which showedin-VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
3010 KOHL ET AL.
A. B.
2 3 4 5 6 7 8
68P
43
-29,
A* 71 ._..
4*~-0- D3C
i5;,t
.i'
:
1 2 3 :
97 68
-43 .4
29 18
18k-14
FIG. 2. Intra- and extracellular expression of viral proteins in MoHIV-I andMoHIV-III producer celllines. Proteins from trans-fectedcells(odd-numberedlanes) orfrom pelletedvirions released bythecells(even-numbered lanes)wereseparated by SDS-PAGEin 12.5% gels, transferred to Immobilon membranes, and incubated withgoatanti-Rauscherp30serum (A) or humananti-HIV antibody-positive antiserum(B).Immunoreactiveproteinswerevisualizedby incubationwith'25I-labeledproteinAfollowed byautoradiography. Molecularmass values ofprestained markers (Bethesda Research Laboratories) are indicated (in kilodaltons). (A) Cells were trans-fectedwithpSV2neo(lanes 1 and 2),pMoHIV-I-26 (lanes3 and4), pMoHIV-I-33(lanes 5 and 6), orpNCA (lanes 7 and 8). (B) Cells weretransfectedwith pSV2neo (lanes 1 and 2) orpMoHIV-III-4A (lanes3 and 4).Inactivated HIV-1antigen(5.5F.g)(Cytotech)(lane 5) was run as a control.
tracellularexpression of the viral gagpolyprotein, superna-tant from these cultures was clarified and placed over a sucrose cushion. Following centrifugation, the pellet was resuspended in sample buffer and analyzed by SDS-PAGE andimmunoblottingasdescribed above. Viralproteinswere
detected in the supernatants of all lines which showed
intracellular viral protein expression. Whereas CA was the most predominant immunoreactive protein in the superna-tant fromthe pNCA-transfected cells (Fig. 2A, lane 8), the most predominant proteins in cells transfected with pMo-HIV-I were either the gag precursorprotein(Fig. 2A, lane 6) orprocessingintermediates(Fig.2A, lane4).Thus, although HIV-1 PRiscapable ofcleaving the M-MuLV gag precursor to release CA, the reaction is less efficient than that
cata-lyzedby M-MuLV PR. Supernatantfrom theMoHIV-III-4A cells contained primarily the HIV-1 CA, although trace amountsofthe gag precursor andprocessing intermediates were also detected (Fig. 2B, lane 4).
Electron microscopy. To further characterize the viral
particlesreleasedby theMoHIV-Iand
MoHIV-Ill
producer cell lines, the cells were analyzed by electron microscopy. Retroviruslike particles were detected in thin sections of theMoHIV-I-26, MoHIV-I-33, andMoHIV-III-4Acells (Fig. 3).
Particles fromMoHIV-IandMoHIV-IIIcells ranged in size from80to 120 nm. Althoughimmature and mature particles were observed in both cell types, immature particles pre-dominated. In the MoHIV-III-4A cells, approximately 25% of theparticles exhibited the condensed core characteristic ofmature retroviral particles. The number of particles re-leased from the MoHIV-I-26, MoHIV-I-33, and MoHIV-III-4A cells was low (0.5, 1.1, and 0.2 virions per cell, respectively) relative to the number released by cells trans-fected with pNCA (2.95 virions per cell) or cells persistently infected with M-MuLV (7.5 virions per cell).
Maturation of chimeric viral particles is inhibited by L-687908. To demonstrate that HIV-1 PR was responsible for theproteolytic processing of the gag precursor protein in
r.
$ s *.
.,4..
FIG. 3. Electron micrograph ofMoHIV-III-4A cells showinga
virion with a partially condensed core (left) and one with an
electron-dense core,typical ofmaturevirions(right). Bar,100nm.
the chimeric viral particles,we assayed for the ability of a known HIV-1 PRinhibitorto block thiscleavage. L-687908 is apeptide mimetic which inhibits in vitro cleavage ofan HIV-1 PR substrate peptide with a 50% inhibitory concen-trationof 0.03nM(40a). FollowingtreatmentwithL-687908
for 24h,thecellswerewashed and grown foran additional 24hin the presence of thedrug.Viralparticleswere
pelleted
from thecollected medium, andthe protein components of theparticleswereanalyzedby SDS-PAGE and immunoblot-ting. As shown inFig.4A, theproduction of CAin
MoHIV-1-33 viral particles was almost completely inhibited by
L-687908at10FLM(lane3) butwasrestoredat1FLM(lane4). Incontrast, production ofCA in viralparticlesproduced by pNCA transfectants was not significantly affected by
L-687908at10 ,uM(Fig. 4A, lane 7). However, the presence of a low level of the p40 processing intermediate and an increased ratio of gag precursor to CA in particles treated with L-687908 (Fig. 4A, lanes 7 and 8) suggest that the
compound mayweakly inhibit the M-MuLV protease.
Pro-duction ofCA in the treated MoHIV-III viralparticles was
completelyblocked at concentrations of the drug aslowas 100 nM (Fig. 4B, lanes4and5). Production of CAwas first observedat 10nM L-687908(lane 6) andremained
partially
blockedat1 nML-687908(lane7).Analysisofaliquotsof the
clarified culture medium from the treated MoHIV-III-4A
cells in an HIV p24 antigen assay (Coulter) indicated that L-687908 inhibited HIV-1PRwith a50% inhibitory concen-tration of 4.7nM(19a). These data indicate that HIV-1 PR is
responsible forproteolytic processing of the M-MuLV gag
polyprotein in MoHIV-I particles and theHIV-1 gag poly-protein in MoHIV-III particles.
Chimeric viral particles are not infectious. To determine whether the viral particles released by the MoHIV-I and MoHIV-III producer cell lines were infectious, culture su-pernatants were applied to fresh NIH 3T3 cells in the presence ofPolybrene. Successful infectionwasmonitored
by immunoblot analysis. Whereas expression of M-MuLV CA was detected in NIH 3T3 cells exposed to supernatant frompNCA producer cellsat1weekpostinfection, noviral
proteins were detected in the lysates of NIH 3T3 cells
exposedtosupernatantfrom eitherMoHIV-I or
MoHIV-Ill
producercells(data notshown). NIH 3T3 cells whichwere J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.319.557.72.252.2] [image:4.612.61.302.75.198.2]M-MuLV/HIV-1 HYBRID VIRAL PARTICLES 3011
A. B.
1 2 3 4 5
97
A5>-
__
- _~~~~~~~~43
'¢68-
*
-2E5
2-9
[image:5.612.59.292.75.206.2]18
FIG. 4. Proteolytic processing of the gag polyprotein in
MoHIV-I and MoHIV-III viral particles is inhibited by L-687908. Stable producer linesweretreated with L-687908 for 24 h, washed
extensively, and incubated in the presence of the drug for an
additional 24 h. The protein componentof viral particles pelleted fromthe 24- to 48-h clarified supernatant was analyzed by SDS-PAGE and immunoblotting. Molecular mass values of prestained
markers areindicated (in kilodaltons). (A) Goat anti-Rauscher p30
serum wasusedtodetect the M-MuLV CA in MoHIV-I and pNCA viral particles. The viral particles were from MoHIV-I-33 cells
whichwerenottreated (lane1),treated with0.1%DMSO (lane2),
ortreated with 10 ,uM (lane 3)or1 ,uM (lane 4) L-687908 and from
pNCA-20 cells whichwere nottreated (lane 5), treated with 0.1% DMSO (lane 6), ortreated with 10 p.M (lane 7)or 1 p.M (lane 8)
L-687908. (B) Human anti-HIV antibody-positive antiserum was
usedtodetect the HIV-1 CA in viral particles released by MoHIV-III-4A cells which were not treated (lane 2), treated with 0.1% DMSO (lane 3), ortreated with 1 ,uM (lane 4), 100 nM (lane 5), 10
nM(lane 6),or1 nM(lane 7) L-687908. Lane 1, 5.5 pgof inactivated HIVantigen (Cytotech)runas acontrol.
exposedtoMoHIV-III supernatant and passaged continued tobenegative for viral protein expression forupto1 month postinfection. The chimeric particles also failed to form plaquesin an XCassay(datanotshown).
To determine why the chimeric viral particles were not infectious, we first examined the level of RTactivity asso-ciated with theMoHIV-IandMoHIV-IIIviralparticles. The level of RT activity in the MoHIV-III particles was tested separately inthepresenceof
Mg2"
andMn2"
to accountfor thepreference of HIV-1 RT forMg2".
As shown in Table 1, littleor noRTactivitywasdetected in either theMoHIV-Ior MoHIV-III-4A viral particles. Attempts to detect RT in MoHIV-I andMoHIV-III cell lysatesandvirion particles by immunoblot analysis wereconsistently negative under con-ditions in which the protein was readily detected in thecontrols(pNCAcells and virions for M-MuLV RTexpected to be presentin MoHIV-I and inactivated HIV antigen for HIV-1 RT in MoHIV-III; data notshown), suggestingthat mature RTwasnotproducedin either celltype.
The lack ofinfectivityof the chimeric viralparticles could also be dueto afailureto packagethe viralgenomic RNA. To examine this possibility, particles werepelleted by cen-trifugation from clarified supernatant collected from MoHIV-I-26 or MoHIV-III-4A producercells. Analysis of the clarified supernatant from the MoHIV-III-4A cells by
enzyme immunoassay (Coulter) indicated that it contained
particles equivalent to 154 pg of CA. Nucleic acids were extracted from two-thirds of the pelleted viralparticlesand analyzed byNorthern (RNA) blotting.As shown inFig. SA, whereas MoHIV-I-26 and pNCA virions contained signifi-cantamounts ofhybridizable RNAcorrespondingin sizeto genomic RNA (lanes 2 and 3), no genomic RNA was
TABLE 1. RTlevels in particlesreleased byMoHIV-Iand MoHIV-III stableproducercellsa
RT activity (cpm) Cell line
Mn2- ([32pJTrp) Mg2+([3HITTP) MoHIV-I
neo-20 429
neo-21 613
MoHIV-I-26 1,451
MoHIV-I-30 6%
MoHIV-I-31 976
MoHIV-I-33 1,143
pNCA-4 43,198
pNCA-20 38,537
pNCA-21 46,036
MoHIV-III
neo5-2 313 75
MoHIV-III-4A 250 141
pNCA-B 23,418 4,128
HIV/RTr 61,668
a Viral particleswerepelletedfrom10mlof clarifiedsupernatantcollected
from the indicated cells in 10-cm dishes at 50 to 100%1 confluence and
resuspendedin PBS.RTactivity associated withtheentire pellet(MoHIV-I)
orhalfof the pellet (MoHIV-III)wasdeterminedasdescribed in Materials and
Methods. As acontrol,2.16 nMpurified recombinant HIV-IRT(HIV/RTr;
15)wasusedintheMg2+reaction.
detected in the MoHIV-III-4Aparticles (lane 8). Exposureof the membrane for a more than 40-fold longer time beyond whenbands could easily be detectedinthe pNCA viral RNA still revealed no bandsinthe MoHIV-IIIviralRNA(datanot
shown), suggesting thatfailure todetectRNAwas notdue
simplytothesmall number ofparticles released bythecells.
The protein composition of the viral particles in the
remaining one-third of thepelletwasexaminedby
immuno-blotting. As shown in Fig. 5B, lane 6, both the HIV-1 gag
polyproteinandCAwerepresentintheMoHIV-III-4A viral
lysate. Thus, despite thefailure todetect viral RNA inthe MoHIV-III-4A particles, the particles contained the
ex-pectedviral proteins.
To examine the possibility thatfailure to detect RNA in the MoHIV-III-4A particles was due to a defect in the
expression of viral sequences in the transfectants, cellular
RNA from the producer cells was analyzed by Northern blotting.Asshown inFig. 5A, thepatternof viraltranscripts expressed in the MoHIV-III-4A cells was similar to that observed in thepNCA producercells(comparelanes 11 and
12).Moreover, the leveloftheseviraltranscriptswassimilar
in the MoHIV-III-4A andpNCA producercells. These data suggestthat the lackofRNAin the MoHIV-III-4Avirionsis not due to a decrease in the level of viral RNA in the
producercells.
DISCUSSION
This report describes the construction and
characteriza-tionoftwoM-MuLV/HIV-1chimericplasmidswhich,when transfected into NIH 3T3cells,direct theproductionof viral
particles which express active HIV-1 PR. Transfection of NIH 3T3 cells with pMoHIV-III DNA yielded cell lines which expressed active HIV-1 PR at only a very low
frequency. Thus, although 13 of the MoHIV-III transfec-tants tested
expressed
the gagpolyprotein, only
one line exhibited significantprocessingof this precursor. It may be that a mutation which allows theproduction
ofthe PRby
another mechanism may have occurred in the MoHIV-VOL.65, 1991
I M.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.310.552.96.269.2]3012 KOHL ET AL.
A. B.
Virions Cells
1 2 3 4 5 6
.P
f
Mo HIV-28S I...i"'
7 ~~ - 18S
7 8 9 10 1112
III
S28
* -0F' w <18S
1 2 3
.
-4 5 6
-'97
-FIG. 5. (A)Expression of viralRNA instableproducer cellsand viralparticles. RNAwasisolatedfrom viralparticles pelleted from
the clarified supernatants of neo5-2 (lanes 1 and 7), MoHIV-I-26 (lane 2), pNCA-B (lanes 3 and 9), and MoHIV-III-4A (lane 8) cultures and from neo5-2 (lanes 4 and 10), MoHIV-I-26 (lane 5), pNCA-B (lanes 6 and 12), and MoHIV-III-4A (lane11) cells. Ten micrograms of cellular RNA and two-thirds ofthe particle-associ-ated RNA were fractionated on a 1% formaldehyde-agarose gel, transferredtonitrocellulose,andhybridized witha32P-labeled DNA
fragment corresponding to the M-MuLV LTR. Ethidium bromide staining of thegel indicatedthattheamountof pNCAcellular RNA (lane 12)loadedwasapproximately twofold lessthan theamountof MoHIV-III-4Acellular RNA(lane 11).The positions of the 28S and 18S rRNA bands are indicated. (B) Analysis of proteins in viral particles. Proteins inone-third of theparticles produced by neo5-2 cells(lanes1 and5), MoHIV-I-26cells(lane 2), pNCA-B cells (lane 3),andMoHIV-III-4Acells(lane 6)wereseparated by SDS-PAGE
in12.5%gels, transferredtoImmobilonmembranes, andincubated withgoatanti-Rauscherp30serum(lanes 1to3)orhumananti-HIV antibody-positiveantiserum (lanes 4to6).Immunoreactive proteins
werevisualized by incubation with125I-labeled proteinAfollowed by autoradiography. Lane 4,2.75 ,g ofinactivated HIV-1antigen (Cytotech) run as a positive control. Molecular mass values of
prestained markersareindicated (in kilodaltons).
III-4Acell line. Althoughsequence analysis of the provirus in theMoHIV-III-4A cells might elucidate suchamutation,
identification ofthe provirus which is capable of this
proc-essingfromamongthemany copies of theprovirus in these cells(approximately 100 copies, asdetermined by Southern analysis [9a]) would be difficult.
Although expression of active HIV-1 PR was detected
only at a low frequency in the pMoHIV-III transfectants, expression of the gag polyprotein was observed in the majority ofthese cells. Expression of the HIV-1 structural proteins both in the virus and from eukaryotic expression
vectors requirescoexpression of the viralrev gene(8,11-13, 17, 19, 24, 40). It isinterestingthatgagexpressionoccursin the absence of Revin theMoHIV-IIIviralparticles. Perhaps an activity analogous to Rev is encoded by the M-MuLV genome.
Analysisof theproteincomponentsof the MoHIV-Iviral
particles indicated that HIV-1 PRwas capable ofcleaving the M-MuLV gag polyprotein to yield the mature core
protein.This observation is consistent with a previous report that bacterially expressed HIV-1 PR is capable of cleaving the Gazdar MuLV gag polyproteininvitro(5). Reaction in
vitro ofpurified, Escherichia coli-produced HIV-1 PR with
synthetic peptide substrates followed by sequencing of the
cleavage productsdemonstrated that the enzymewas capa-ble ofaccurately processingall of theM-MuLVgagandpol
cleavage sites with the exception of the NC/PR site, for which cleavage was observed to occur one amino acid N-terminaltothepredictedsite. For the CA/NC and PR/RT
substrates, an additional cleavage site was also observed (7a). These results suggest that HIV-1 PR might correctly cleave the M-MuLV gag andpol polyproteinsin vivo at all butone site.
We have observed that the protease inhibitor L-687908 shows differential inhibition of CA production in the MoHIV-I andMoHIV-III particles. Since theprimary
anti-bodyused in the immunoblotanalysisis theonly
distinguish-ing feature in the assay, it is possible that the observed difference results from differential affinity of the two
anti-bodies forthe precursor and matureforms of therespective antigens. Alternatively, the difference in sensitivity ofthe MoHIV-I and MoHIV-IIIparticlesto the inhibitor could be due to the presence of different polyprotein substrates. L-687908isacompetitiveinhibitor of the PR and mayexhibit a range of potency in the context of various substrates.
Indeed, the L-687908 class of protease inhibitors exhibit differential inhibition of the PR in thecontextofthe various HIV gagcleavage sites(lla).
NIH 3T3 cells stably expressing either pMoHIV-I or
pMoHIV-III
released viral particles which were notinfec-tious. Inboth cases, the lack ofinfectivity couldbe attrib-uted to failure to produce active RT. Although neither matureRTnor its precursorpolyprotein could be detected
by immunoblot
analysis
in eithercell type,failuretodetect thelargeprecursor may be duetopoor transfer of theprotein tothemembrane.Itseemslogicaltoexpectthat thegag-polpolyprotein should be synthesized because the viral PR, which is synthesized as part of the precursor, is active in both MoHIV-I and MoHIV-IIIparticles. Furthermore,
nu-cleotide sequenceanalysis of theRT coding regioninboth
pMoHIV-I and pMoHIV-III confirmed the expected se-quence (data not shown). Thus, premature termination
seems an
unlikely
explanation
for failuretodetectRTunless a mutation was introduced duringthe establishment of the cell lines. If the gag-polprecursor was synthesized in the MoHIV-I and MoHIV-III cells, RT activity should have beendetected, sinceprocessingof the gag-pol precursortoyield
thematureform ofRTisnotnecessaryforactivation of the enzyme(7).It is notable that packaging of the viral genomic RNA occurs in the MoHIV-I but not the MoHIV-III particles.
Interaction ofavirionproteinwithasiteontheviral genome is thoughttodirect the packagingof the RNA. Mutational
analysis
ofthecysteine
arrayof thenucleocapsidprotein
of M-MuLV (16, 31) and, morerecently, HIV-1(3) suggests a rolefor this viralproteinin RNAencapsidation. Regionsof the M-MuLV genome between the 5' LTR and the gag J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.70.300.73.374.2]M-MuLV/HIV-1 HYBRID VIRAL
initiation codon
(*,
sequence) as well as within the gag coding regionarerequiredincis for efficientpackagingof the genome and subsequentproduction of infectious virions (2, 4, 28). Recently, sequences in the corresponding region of HIV-1 were shown to have a similar function (3, 26). Although these sequences are highly conserved among dif-ferent HIV-1strains (26), they differ from the corresponding sequences inM-MuLV. It is likely that this sequence differ-ence may prevent the interaction of the HIV-1-derivednucleocapsid protein with the specific packaging site onthe
M-MuLV-derived RNA in the MoHIV-III-4A cells. Several characteristics of the MoHIV-III-4A cell line make itanideal reagent for the evaluation of potential HIV-1 PR inhibitors. As demonstrated in this report, proteolytic
processing of the HIV-1 gag polyprotein in the particles can be inhibited in a dose-dependent fashion, and this inhibition is easily monitored. In addition, the MoHIV-III-4A cell line
providesastablesourceof viralparticles whichexpressand processthe HIV-1 gagpolyprotein. This property confersa distinct advantage over the transient systems which have recently been developedforthe expression and processing
of the gagpolyproteinin viruslikeparticles (18, 22, 37, 39). Finally, the viral particles released by the MoHIV-III-4A cells are not pathogenic, enabling large-scale assays to be runwithout concern for workersafety.
ACKNOWLEDGMENTS
We thankS. Goff forthegift of pNCAandMalcom Martin for supplying pNL4-3. We are grateful to Julie Bowen and Mark Goldman for performingRT assays, toRobert Keysforhelp with DNAsequencing,and to Paul Darkefor performinginvitroprotease assays.
REFERENCES
1. Adachi,A., H. E. Gendelman,S.Koenig,T.Folkes,R.Willey, A. Rabson, and M. A. Martin. 1986. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J.Virol. 59:284-291.
2. Adam,M.A., and A. D. Miller.1988.Identification ofasignal in
amurine retrovirus that issufficient forpackaging of
nonretro-viralRNAinto virions. J.Virol. 62:3802-3806.
3. Aldovini, A., and R. A. Young. 1990. Mutations ofRNA and protein sequencesinvolved inhuman immunodeficiency virus type1 packaging result in production of noninfectious virus. J. Virol. 64:1920-1926.
4. Bender, M. A., T. D. Palmer, R. E. Gelinas, andA. D.Miller. 1987. Evidence that the packaging signal of Moloney murine leukemia virus extends into thegagregion. J. Virol. 61:1639-1646.
5. Bu, M., S. Oroszlan, and R. B. Luftig. 1989. Inhibition of bacteriallyexpressedHIV proteaseactivitydeterminedbyanin vitro cleavage assay with MuLV Pr65gag. AIDS Res. Human Retroviruses 5:259-268.
6. Colicelli, J.,and S. P. Goff.1988.Sequenceandspacing
require-mentsofaretrovirusintegration site.J. Mol. Biol. 199:47-59.
7. Crawford,S.,andS. P. Goff.1985. Adeletion mutationin the 5' partof the polgeneofMoloney murine leukemia virus blocks proteolyticprocessing ofthe gagandpolpolyproteins.J.Virol. 53:899-907.
7a.Darke,P. Unpublisheddata.
8. Dayton, A. I., E. F. Terwilliger, J. Potz, M. Kowalski, J. G. Sodroski, and W. A. Haseltine. 1988. Cis-acting sequences responsive totherevgene productof the human immunodefi-ciency virus.J. AcquiredImmuneDefic. Syndrome 1:441-452. 9. Debouck, C., and B. W. Metcalf. 1990. Human immunodefi-ciency virus protease: a targetfor AIDS therapy. Drug Dev. Res. 21:1-17.
9a.Diehl, R.E.,and N. E. Kohl. Unpublisheddata.
10. Dreyer, G. B.,B. W.Metcalf,T. A.Tomaszek, Jr.,T.J.Carr, A.C. ChandlerIII,L.Hyland,S. A.Fakhoury,V. W.Magaard, M.L.Moore, J.E.Strickler,C.Debouck,and T. D. Meek.1989. Inhibition ofhumanimmunodeficiencyvirus 1 protease invitro: rational design of substrate analogue inhibitors. Proc. Natl. Acad. Sci. USA 86:9752-9756.
11. Emerman, M., R. Vazeux, and K. Peden. 1989. The revgene productof the humanimmunodeficiencyvirusaffects envelope-specificRNAlocalization. Cell 57:1155-1165.
11a.Emini,E. Unpublisheddata.
12. Feinberg,M.B.,R. F.Jarrett,A.Aldovini,R.C.Gallo,and F. Wong-Staal.1986.HTLV-IIIexpressionandproductioninvolve complexregulation at the levelsofsplicingand translation of viralRNA.Cell 46:807-817.
13. Felber,B.K.,M.Hadzopoulou-Cladaras,C.Cladaras,T. Cope-land,andG. N. Pavlakis. 1989. revproteinofhuman immuno-deficiencyvirustype 1affectsthestabilityand transport ofthe viralmRNA. Proc. Natl. Acad. Sci. USA 86:1496-1499. 14. Goff, S., P. Traktman, and D. Baltimore. 1981. Isolation and
propertiesofMoloneymurineleukemia virusmutants: useofa rapidassayfor releaseof virionreversetranscriptase.J. Virol. 38:239-248.
15. Goldman,M.E.,G. S.Salituro,J.A.Bowen,J.M.Williamson, D. L.Zink,W. A.Schleif,and E. A. Emini.1990. Inhibition of humanimmunodeficiency virus-1reversetranscriptase activity byrubromycins:competitiveinteractionatthetemplate-primer
site. Mol. Pharmacol. 38:20-25.
16. Gorelick,R. J.,L. E. Henderson, J. P. Hanser,and A. Rein. 1988.PointmutantsofMoloneymurine leukemia virus that fail
topackageviralRNA:evidence forspecificRNArecognition by
a "zinc finger-like" protein sequence. Proc. Natl. Acad. Sci. USA 85:8420-8424.
17. Hadzopoulou-Cladaras, M.,B. K.Felber,C.Cladaras,A. Atha-nassopoulos,A.Tse,andG. N.Pavlakis.1989. Therev(trslart) protein ofhuman immunodeficiency virus type 1 affects viral mRNAandproteinexpressionviaacis-actingsequencein the
envregion.J.Virol. 63:1265-1274.
18. Haffar, O.,J. Garrigues, B.Travis, P. Moran,J. Zarling,and S.-L. Hu.1990. Humanimmunodeficiency virus-like,
nonrepli-cating, gag-env particles assemble in arecombinant vaccinia
virusexpressionsystem.J.Virol. 64:2653-2659.
19. Hammarskjold, M.-L.,J. Heimer, B. Hammarskjold, I. Sang-wan, L. Albert, and D. Rekosh. 1989. Regulation of human
immunodeficiencyvirusenvexpression bytherevgeneproduct.
J. Virol. 63:1959-1966.
19a.Hanobik,M. G.,and N. E. Kohl. Unpublisheddata.
20. Haseltine,W. A. 1988. ReplicationandpathologyoftheAIDS virus.J.AcquiredImmune Defic.Syndrome 1:217-240. 21. Jacks, T.,M.D.Power,F. R.Masiarz,P. A.Luciw,P.J.Barr,
and H. E. Varmus. 1988. Characterisation of ribosomal frame-shiftingin HIV-1gag-polexpression.Nature(London) 331:280-283.
22. Karacostas,V.,K.Nagashima,M. A.Gonda,and B. Moss.1989. Human immunodeficiency virus-like particles produced by a vaccinia virusexpressionvector. Proc. Natl. Acad. Sci. USA 86:8964-8967.
23. Kay, J.,and B.M. Dunn.1990. Viralproteinases:weaknessin strength.Biochim.Biophys. Acta1040:1-18.
24. Knight,D.M.,F. A.Flomerfelt,andJ.Ghrayeb.1987.
Expres-sionof the art/trsproteinofHIV andstudyof its rolein viral envelopesynthesis. Science236:837-840.
24a.Kohl,N. E.Unpublisheddata.
25. Kohl, N. E., E. A. Emini, W. A. Schleif, L. J. Davis, J. C. Heimbach,R.A.F.Dixon,E. M.Scolnick,andI. S.Sigal.1988. Active humanimmunodeficiency virusproteaseis
required
for viralinfectivity. Proc. Natl.Acad. Sci. USA85:4686-4690. 26. Lever,A., H. Gottlinger,W. Haseltine, andJ. Sodroski. 1989.Identification ofasequencerequiredfor efficient
packaging
of human immunodeficiency virus type 1 RNA into virions. J. Virol. 63:4085-4087.27. Maniatis, T.,E. F. Fritsch,andJ. Sambrook. 1982. Molecular
cloning: alaboratory manual, p. 187-209. ColdSpring Harbor Laboratory, ColdSpringHarbor,N.Y.
on November 10, 2019 by guest
http://jvi.asm.org/
3014 KOHL ET AL.
28. Mann, R., R. C.Mulligan,andD.Baltimore.1983.Construction ofaretrovirus packaging mutantand ituse to produce helper-free defectiveretrovirus.Cell 33:153-159.
29. McQuade, T. J., A. G. Tomasseili, L. Liu, V. Karacostas, B. Moss, T. K.Sawyer,R.L.Heinrikson,andW. G.Tarpley.1990. A synthetic HIV-1 protease inhibitor with antiviral activity arrestsHIV-like particle maturation. Science 247:454-456. 30. Meek, T. D., D. M. Lambert, G. B. Dreyer, T. J. Carr, T. A.
Tomaszek,Jr., M. L. Moore, J. E. Strickler,C.Debouck,L.J. Hyland, T. J. Matthews, B. W. Metcalf, and S. R. Petteway. 1990.Inhibition ofHIV-1 proteaseininfectedT-lymphocytes by synthetic peptide analogues. Nature(London)343:90-92. 31. Meric, C., and S. P. Goff. 1989. Characterization ofMoloney
murineleukemia virusmutants withsingle-amino-acid substitu-tionsinthe Cys-Hisbox ofthenucleocapsidprotein. J. Virol. 63:1558-1568.
32. Peng, C., B. K. Ho, T. W. Chang, and N. T. Chang. 1989. Role of human immunodeficiency virus type 1-specific protease in coreprotein maturation and viralinfectivity.J. Virol. 63:2550-2556.
33. Roberts,N.A., J. A. Martin, D.Kinchington, A. V. Broadhurst, J. C. Craig, I. B. Duncan,S.A.Galpin, B. K. Handa, J. Kay, A. Krohn, R. W. Lambert, J. H. Merrett, J. S. Mills, K. E. B. Parkes,S. Redshaw,A. J.Ritchie, D. L. Taylor, G. J.Thomas, and P.J.Machin. 1990. Rational design ofpeptide-based HIV proteinase inhibitors. Science 248:358-361.
34. Rowe, W. P., W. E. Pugh, and J. W. Hartley. 1970.Plaqueassay techniques for murine leukemia viruses. Virology 42:1136-1139. 35. Sherr,C. J., and G. J. Todaro. 1979.Purificationand assay of
murine leukemia viruses. MethodsEnzymol.58:412-424. 36. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981.
Nucleotide sequenceofMoloneymurineleukaemia virus.
Na-ture(London)293:543-548.
37. Shioda, T.,and H. Shibuta. 1990.Production ofhuman immu-nodeficiencyvirus(HIV)-like particlesfrom cellsinfectedwith recombinant vaccinia viruses carrying the gag gene of HIV. Virology 175:139-148.
38. Skalka, A.M. 1989. Retroviralproteases: firstglimpsesat the anatomyofaprocessingmachine. Cell56:911-913.
39. Smith,A.J.,M.-I.Cho,M.-L.Hammarskjold,and D.Rekosh. 1990. Human immunodeficiency virus type 1 Pr55gag and
Pr1609'9-P'1expressedfromasimian virus 40 latereplacement vector are efficiently processed and assembled into viruslike particles.J. Virol.64:2743-2750.
40. Sodroski, J., W.C.Goh,C. Rosen, A. Dayton, E.Terwilliger, and W. Haseltine. 1986. A second posttranscriptional
trans-activatorgenerequiredforHTLV-IIIreplication. Nature (Lon-don)321:412-417.
40a.Vacca,J.P.,J.P. Guare,S.J. deSolms,W.M. Sanders,E.A. Giuliani,S.D.Young,P.L.Darke,J. Zugay,I. S.Sigal,W.A. Schleif, J. C. Quintero,E. A.Emini, P.S.Anderson,andJ.R. Huff. 1991. L-687,908, a potent hydroxyethylene-containing HIVproteaseinhibitor. J. Med. Chem. 34:1225-1228. 41. Wigler, M.,R.Sweet,G.K.Sim,B.Wold,A.Pellicer,E.Lacy,
T.Maniatis,S.Silverstein,andR.Axel. 1979.Transformationof mammaliancellswithgenesfrom procaryotesandeucaryotes. Cell16:777-785.
42. Yoshinaka, Y.,I.Katoh,T. D.Copeland,and S.Oroszlan.1985. Murineleukemia virusproteaseis encodedbythegag-pol gene and issynthesizedthrough suppressionof anambertermination codon. Proc. Natl. Acad. Sci. USA82:1618-1622.
J.VIROL.