0022-538X/84/030731-10$02.00/0
Copyright C) 1984, American Society for Microbiology
Deletion and Insertion Mutations
in Early Region
la
of
Type
5
Adenovirus That Produce Cold-Sensitive
or
Defective
Phenotypes
for
Transformation
LEE E. BABISS, P. B. FISHER, ANDH. S. GINSBERG*
Departmentof Microbiology, Cancer CenterlInstitute of Cancer Research, Columbia University College of Physicians and
Surgeons, New York, New York 10032
Received19 August 1983/Accepted9November 1983
On the basis of earlierfindings showing that HShrl (hrl) is cold sensitive fortransformation, a seriesof
mutantswereconstructedsothat theycontained deletionsorinsertions indifferentsitesofearly region la
(Eta)toascertain: (i) whether the cold-sensitive phenotype ofhrlwasthe result of theidentifiedsingle-base
pair deletion of nucleotide 1,055orduetoamissense mutationatanother site and(ii)what regionand how
much of the Ela 51-kilodalton protein is actually required to produce cell transformation. A mutant,
HSdllOl (d1101), was constructed to contain a 5-base pair deletion ofnucleotides 1,008 to 1,012, which
producedaframeshift andasubsequentstopcodonatnucleotide1,241. Thismutant,whichshould encodea
truncated 33-kilodalton protein in place of the wild-type 51-kilodalton protein, had a cold-sensitive
phenotype for transformation essentially identicalto hrl. Consonantwith thisfinding, amutant (HSinlO6)
engineeredtocontaina16-basepair insertioninitiated after nucleotide 1,009,withastop codonbeginningat
thenewly inserted nucleotide 1,013, also had acold-sensitive phenotype like hrl anddllOl. It is striking,
however, thatamutant(H5dllOS)witha69-basepair deletion beginningatnucleotide 1,003,and havinga
stop codon atnucleotide 1,544, was totally defective for transformationat anytemperature. Transfection
studies withplasmids containing the ElaorEla and Elbgenesofsub3O9, hrl, anddllOl further revealed
that the cold-sensitive transformation phenotype observed couldbe exhibited in the absence of viral Elb
geneexpression.
Theearlyregionla(Ela) geneof adenovirus5 (AdS) (0to
4.5 mapunits [MU]) has been shown to serve an essential
role ininitiating both the viralproductive cycleinpermissive cells (3, 24) and the transformation of rodent cells (15, 20,
43). Whereas a number of deletion, insertion, and point mutations have been introduced into thisgene(7, 17, 25, 39),
very littleis known about the mechanism(s) bywhich
Ela-encoded proteins may interact with cellularfactors (31, 32) or virus-encoded macromolecules or both to elicit their
biological effects.
During
the
viralproductive infectiontwospliced mRNAs are synthesized early, 12S and 13S (4, 10, 26, 41), andMontelletal. (30)have shown that the proteins encoded by
the smaller 12S mRNA are notobligatory for viral replica-tion. Viral host range mutants containing alterations in the
13SmRNAcoding
capacity
havethusfar been showntobedefective in transformation and viral replicative functions, which suggests that the acidic 51-kilodalton (kd) protein
encoded
by
this mRNA (35) plays a critical role in bothprocesses.
The requirement for the 51-kd protein in the process of cellular transformation has been demonstrated by using a
viralhost range mutant, hrl (21), whichproduces a truncated
form of thisprotein (35). Recentinvestigations indicatethat
hrl is cold sensitive forthe maintenance of transformation
(1, 22). By using a cloned rat embryo fibroblast cell line,
CREF, it was demonstrated that clones ofhrl-transformed
cells display the transformed phenotype in a
temperature-dependent manner; i.e., at 32°C they reflect the normal
phenotypeof CREF cells and when shifted to 37°C as long as
3 weeks after infection they convert to the transformed
phenotype (1). These observations in conjunction with DNA *Correspondingauthor.
731
transfer-filterhybridization studiessuggest that thoseevents
leading to initiation, i.e., integration, are not conditionally
regulated in hrl-infected cells.
Although it has been demonstrated that hrl contains a
single-base pair deletion at nucleotide 1,055 (35), the entire
sequence ofthe Ela gene was not determined. Since this
mutant wasisolatedwith nitrousacid, itwaspossible thatan
additional missense mutation was present and that this mutation produced the conditionally lethal phenotype. To
approachthisproblemand toprobethefunction(s) ofthe
51-kd protein further, threeAdS mutants were constructed so that they contain deletions or insertions of nucleotide
se-quences aroundtheSmaI restrictionendonuclease cleavage
site (2.8 MU). Two of the viral mutants exhibited a
cold-sensitive transformationphenotypewhichwassimilarto that
found with hrl and confirmed that the single-base pair deletion in hrl was in fact responsible for the observed phenotype. In addition, DNA sequence analyses of the altered viral Ela gene sequences from the three viral
mu-tants revealed that coding sequences varying by no more
than 5 base pairs (bp) coulddetermine whetheratruncated
protein produced by the 13S mRNA was conditionally functional or inactive at 32 and 37°C. DNA transfection
studies described in this paper demonstrate that the Ela
gene sequences from the mutant viral genomes are capable
ofeliciting theconditional phenotypein the absence of Elb
genefunctions.
MATERIALSANDMETHODS
Constructionand characterizationofAd5 Eladeletionand
insertion mutants. The HpaI-EDNA fragment ofAdS (0 to
4.5 MU) was cloned into the PstI siteof
pBR322
(5),
usingtheproceduredescribedbyStow(42). A 2-,ugportionof this
on November 10, 2019 by guest
http://jvi.asm.org/
XboI 0 3.8
XmoI XbaI 0 2.8 3.8 4.5
I a
Xmo I
Sl
orBal 31Ligate
XbaI
0
3.8 4.5
I I L
XboI,
Ligote>
XbaI
0
8Xba I
Isolate
3.8-100
fragment
I
Transfect
293 cells
100 5 0
° H5dllOl
H5dtlOS
SmaI XboI 0 2.8 3.8 4.5
Xbol linkers
Ligate
XbaI XbaI 0 2.8 3.8 4.5
I I
' mix
XbaI XbaI 3.8
0,2-8
/ 100FIG. 1. Diagramof the methods usedto construct mutantsin the ElA region of Ad5. The diagram shownrepresentstheconstruction of dIlOl anddllOSby ligation of the mutated0-to3.8-MUfragmenttothe3.8-to100-MUfragmentof sub309 and theformation of in106 by
over-lap recombination betweenthemutated0-to4.5-MUfragment and the3.8-to100-MUfragmentofsub309. The0-to4.5-MU DNAfragment of Ad5 was cloned into the PstI site in pBR322. Open parenthesis represent areas ofdeleted DNA sequences, andopen triangles denote
restrictionenzymecleavage sitespresentontheviralgenomes.
plasmid DNAwascleaved with XmaI(2.8 MU) and digested
with either 6.25 Uornuclease S1 (P-L Biochemicals) for 30
minat30°Cor0.001 U of nuclease Bal31 (BethesdaResearch
Laboratories) for 30 minat 37°C. After the recircularization
of thesemolecules with T4ligase and cleavage with SmaIto
remove any molecules that escaped nuclease digestion,
Escherichia coli HB101 cells were transformed to
tetracy-cline resistance. Plasmids containing the desired deletions
were isolated, and their DNAs werepurified by the
Sarko-syl-lysate procedure of Clewell and Helinski (11).
Toprepareinsertions of linker DNA into the SmaI restric-tionenzymesite(Fig. 1), 1p.gofplasmid DNAwasdigested
with this enzymeandligated witha200-fold excessof XbaI
linker ends (CTCTAGAG; Collaborative Research). After
digestion with SmaI, the ligated molecules were used to
transform E. coli HB101 cells, selecting for
tetracycline-resistant colonies containing plasmids withtwo XbaI sites
2.8 and3.8 MU.
The modified viral Elagene contained withinthese
plas-midswereintroduced into the viralgenomeofsub3O9(25) by
aprocedure described by Stow (42) for dIlOl and d1105orby
overlap recombination (9). Toenhance thefrequency ofthe
overlap recombination event, subconfluent cultures of 293
cells (19) maintained in Dulbecco modified Eagle medium
supplemented with 10% calfserum wereexposed to 15 J of
UV light (254 mm; General Electric germicidal lamp), and
theirradiated cells were then incubated for 16 h at 37°C in
thedark. After the transfection of these cells (19) with the
overlappingDNAs (1 jig ofsub3O9 DNAcleaved with ClaI
and XbaI and 1 [tg equivalent of plasmid DNA containing Elagene sequences cleaved with PstI), viralplaques were
isolated after incubation for11days at 37°C.
All viruses were plaque purified three times before
pro-ducing stocks for characterization anduse. Mutant viruses
were identified by their SmaI digestion pattern (Fig. 2),
which revealed the loss ofone enzyme siteat2.8 MU. The
size of the deletions generated and the position of XbaI
linker in inlO6wasdeterminedby cloning the ClaI (2.6
MU)-HpaI (4.5 MU) DNA fragment from each virus into plasmid
vector pDR33 (37) containing unique ClaI and HpaI sites.
All of the plasmids constructed were propagated in E. coli
strain GM33 (27), which is deficient in adenine methylase.
The DNA was sequenced, according to the Maxam and
Gilbert (28)technique, after the plasmids werelinearized at
the ClaI site, and the 5' end of the plasmid DNA was
phosphorylated with T4polynucleotide kinase.
Viral transformation and DNA transfection assays with
CREF cells. CREF cells were usedfor viral transformation
assays aspreviously described (1, 12)at amultiplicity of
10-PFU of H5sub3O9, H5dllO1, or H5inlO6 or 30 PFU of
H5dllO5 percell. DNAtransfection assays wereperformed
with7 ,ug of recombinant plasmid DNA, with 3
jg
of salmonspermDNA ascarrier(19). Approximately 10 CREFcells
inmonolayers in 10-cm platesweretransfected, and after 4 h
at 37°C the plates were treated successively with 15%
glycerol in phosphate-buffered saline, plain
phosphate-buff-ered salinealone, andtrypsin(0.125%)-EDTA (0.05%). The
cellswerereplatedat105 cellsina5-cm culturedish and fed
with low-Ca2+ medium twiceaweek for 8 weeks at37°C or
11 weeksat32°C,afterwhichtransformed fociwerecounted
and isolated. In all assays, each group contained eight
cultures ofreplated cells (i.e.,
105
cellsperculturedish).Construction of recombinantplasmidscontaining earlygene sequencesfromhrl,dllOl,and sub3O9. The steps involved in
thecloning ofthe Elagene(HpaI-Efragment, 0to45MU)
or Ela and Elbgenes (XhoI-C fragment, 0 to 15.5 MU) of
hrl, sub3O9, and dllOl areoutlined and described in Fig. 3.
pLB209wasconstructed withpACYC177 plasmid DNA (8)
100
H5sub 309
H5 in 106
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.134.485.75.326.2]A. o ) °0
ro - c:
0 LO)
t -r
-6
I
B.
2-1c
1r() C
rn U_
-o
f___I
-A
-B
-c -
F/K'
-DE-F -G
-H
-I
-K
-L-FIG. 2. Electrophoretic analysisofmutantviralgenomes,using
SmaIandXbaI restriction endonucleases. A1-pRgamountof DNA
from each virus described was digested with 4 U of (A) SmaI
(sub3O9, dIlOl,d11O5,andinlO6)or(B)XbaI(sub3O9andinlO6),and thefragmentswereseparatedon a0.6%agarosegel containing1 pg
of ethidium bromide perml. The bands were visualized with UV lightandphotographed.The F/K* bandgenerated bySmaIcleavage ofdIlOl, dllO5, and inlO6 represents afusion of the SmaI-F + K bands since the enzyme site at 2.8 MU is no longerpresent. The
presence of an additional XbaI site at 2.8 MU is detected by
cleavage of inlO6 with this enzyme to yield two unique DNA fragments, Bi (O to 2.8 MU) and B2 (2.8 to 3.8 MU). The DNA fragmentlabeledB inthe sub3O9lane extends from0 to3.8 MU.
and the AdS XhoI-C fragment instead ofthe Ad2 XhoI-C
fragment byaprocedure previously described (2). All DNA
fragmentsusedforligation and transformationwere purified
withlow-melting-pointagarose(Bethesda Research
Labora-tories). For the dIlOl plasmids pLB211 and PLB214, the
presence of the deletion at 2.8 MU was confirmed by
restriction enzymeanalysis.
Characterization of cells transformedby dIlOl, hrl, inlO6, and sub3O9. Two transformed fociresultingfrom viral infec-tion orDNAtransfection from each virus type werecloned
by seeding 100 cells per 90-mm plate and isolating
well-separated clones by using steel cloning cylinders. The ability of the cloned cell linestogrowinananchorage-independent
manner was assayed in agar as previously described (13).
Briefly, 103, 104, or 105 cells in low-Ca2+ medium
supple-mented with 0.4% Noble agar were layered on 0.8% agar bases prepared in low-Ca2+ medium. The plates were fed once aweek with 3 mlof 0.4%agarinlow-Ca2+ medium,and colonieswere countedat 3 weeks for37°Cand5to6weeks for 32°C cultures.
The presence of viral or plasmid DNA sequences in the cloned transformed cell lineswasdeterminedbyDNA filter-transferhybridization analyses, as previously described (12, 36, 40, 45).
RESULTS
Construction and isolation of viral deletion and insertion mutants. The protocol used to construct Ad5 insertion and deletionmutants sothat thecoding capacity of the Ela 13S
mRNA was affected is described in Fig. 1. Briefly, a plasmid
containing a cloned Ela gene (O to 4.5 MU) of AdS was
linearized with XmaI and digested with nuclease Si or Bal3l. By using an approach described by Stow (42), the modified viral Ela gene sequences were introduced back into a complete viral genome, using mutant H5sub3O9 (sub3O9) DNA (24). sub3O9 virus has a phenotype similar to
that of wild-type Ad5 (Ad5wt), but structurally it contains
only one XbaI site at 3.8 MU. Because of this genomic
organization, the 3.8- to 100-MU fragment of this sub3O9 can
be isolated and ligated to the 0- to 3.8-MU mutated DNA
fragments derived from the isolated plasmids. These ligated
molecules were then used to transfect 293 cells in a direct
plaque assay.
Alternatively, SmaI-digested plasmid DNA was ligated to the octanucleotide CTCTAGAG, which contained an XbaI
recognition site. The Ela sequences containing a newly
inserted XbaI site at 2.8 MU were then introduced into
sub3O9 DNA viaanoverlap recombination mechanism (9).
However, the small areaofhomology existingbetween the
twooverlapping DNA fragments (250 bp) resulted in a very
low frequency ofrecombination. To enhance the occurrence
oftherecombinational event,293cells werepretreatedwith UVlightand transfected after a 16-h inductionperiod in the
dark. The UV irradiation effected an enhanced ability to
recombine the overlapping DNAfragments upon
transfec-tion (L. E. Babiss and H. S. Ginsberg, manuscript in
preparation).
To confirm that the viral plaque isolates obtained did
contain alterations in their Ela gene sequences, after two
cycles of plaque purification on 293 cells, viral DNA was
isolated and analyzed by using SmaI or XbaI restriction
endonuclease. The three viral mutants analyzed all
con-tained alterations at 2.8 MU,which prevented SmaI
cleav-age at that site (Fig. 2A). As a result, the SmaI-K and -F
fragmentspresentintheparentalsub3O9 virus were fused in
the mutants viruses and resulted in a new DNA band of
higher molecular weight. H5inlO6 (inlO6)wasalsofoundto
contain an additional XbaI site at about 2.8 MU, which
confirmed the presence of the linker molecule within the viral genome (Fig. 2B).
Todeterminewhether aframeshiftmutation had occurred as aresult ofthedeletionsintroducedat 2.8MUorwhether
the linker sequences ligated properly at the site, DNA
sequence analyses were performed (28). By using acloned
DNA fragment derived from each ofthe mutant viral
ge-nomes,itwasdetermined thatH5dl101
(dlOl)
containeda5-bp deletion extending from 1,007 to 1,013 bp; H5dl105 (dIlO5) contained a 69-bp deletion extending from 1,002 to
1,072 bp; and H5inlO6 contained an insertion of 16 bp
between 1,009and 1,010 bp (Fig.4). Although no alteration
of the reading frame occurred for d11O5, the frameshift mutation that wasintroduced into thecoding region ofthe
13S mRNA for dllO1 resulted in the occurrence of a stop
codonbeginningatnucleotide 1,241 and presumably caused
termination of translation. The 16-bp insertion determined
for inlO6 indicated that two linker molecules were ligated
into the SmaI site in tandem, which produced an in-frame
stop codon at the insertednucleotide 1,013.
Transformation characteristicsofsub3O9 and Ela deletion
and insertion mutants. It was previously shown that hrl
exhibits a cold-sensitivephenotype for transformation, using
bothprimaryratembryocells(22) and CREF cells (1). When
CREFcells wereinfected withdllOl, followedby replating
oftheinfectedcells at32°Candgrowth for6weeks at32°C, morphologically transformed foci could not be discerned
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.57.297.76.301.2]A
BomHI pLB209'
ANhoI
CI %
%s
0.0 2.6 15.5
B
CloI XhoI
0.02.6 15.5
|XhoI aCIoI
H5hrl
or Ta-Z9-
H5dIlO
100 or
H5sub309
I
2I'll
sub309,pLB210W(iR~- H45dl101,pLB211
11 H5hrl,pLB212
0.0 15.5
BomHlIaHpaoI
>|Ligote
(<m HI
-.g-r '. sub3O9,pLB213
Hpol ( H5d1101,pLB214
| l'B H5hrl,pLB215
4.5 0.0
FIG. 3. Methodsfor constructing plasmids containing mutationsin theearly regionsof Ad5 DNA. Theprocedureforcloningthe Ela and Elbgenesof eachvirus indicated ispresented (A).pLB209 containstheXhoI-C(Oto15.5MU)fragmentofAdScloned intopACYC177 (8)
and itsconstructionhas been describedbefore(2). A2-,ugamountof hrl, sub309, ordllOl virus DNAwascleaved with ClaI andXhoIand mixed andligated with100ngofpLB209 cleaved with thesame enzymesand HpaItoinactivatethe Ad5wtfragment extending from2.6to
15.5 MU. After transformation ofE. coliHB101 cells, ampicillin-resistant colonies were selected and the resulting plasmids, pLB210,
pLB211, andpLB212,wereisolated. To clone only the Elageneofeachvirus(B),one,ug ofeachplasmid described abovewascleavedwith
BamHIandHpaI andligatedto100ngofpDR33plasmid DNA cleaved with thesame enzymesand Sall. Ampicillin-resistant colonieswere
selected aftertransformations ofE. coliHB1Ol cells, and plasmidspLB213,pLB214, and pLB215wereisolated.
(Table 1). However, like hrl, ifdllOl-infected CREF cells were maintained at 37°C, the frequency of transformation was fivefold higher than for sub3O9 or Ad5wt-infected
cul-tures(Table 1). It should also be noted thatdllOlandinlO6
(describedbelow), like hrl, had the same minimal cytopathic
effects on CREF cells as sub3O9, and thus the increased
transformation frequency produced by these mutants must be attributeddirectly to the altered viral product.
Allfocigenerated by the mutant virus at 37°C exhibited a
fibroblasticmorphology; in contrast, sub3O9 and Ad5wt foci
contained a mixture of both fibroblastic and epithelioid
colonies. Shift-up and shift-down experiments with
dllOl-infected CREF cells were similar to the previous findings with hrl (1). When mutant-infected cell cultures were shifted
from 32 to37°C 2 or 3 weeks after infection, the frequency of
transformation approached thatof infectedcells maintained at 37°C throughout theexperimental period (Table 1).
Con-versely, upon shifting cultures from 37 to 32°C, the number
of foci decreased, the majority of the transformed foci
becameflattened, and the cells were not dissimilar from the
surrounding CREF cells.
It is important to note that the cold-sensitive nature of
thesevirusesis not absolute. CREF cells infected with hrl or
dllOl,andmaintainedat32°C for aperiodof 8 weeks, could
developa few small foci, which probably reflected a small
amountof"leakiness" associatedwith thediminished
func-tion of thetruncated protein.
From theseobservations itcan beinferred that hrl
proba-blydoes notcontain asecond-sitemutation. In addition, the
functional domain of the 51-kd proteincould now bemore
precisely positioned sincethemutationpresentin dllOl was
47bp upstream from the hrl deletion.
To determinewhether theadditional missense amino acids
present in the truncated protein encoded by dllOl were
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.164.479.72.460.2]Ad5 Wt
499 561 ACT ATG
+
" 1544
1112J,
"^1229 TAA 1632is %I t I
61lbp 1085 -,A
1055 TGA , '
-GGG6ACCJ-T
H5hrl
1000
a5bp
H5dllO
GGAGCACCMCAC-1000
H5dlI05
GGAG1000
669bp
Protein
M.W.
51K
28K
^ 1241
,' %. TGA
, v
ol%
J. %
(30K)
1544 TAA
(47
K)
1013 TAG v 16bp
[image:5.612.62.531.77.282.2]H5in
106
GGAGCACCCCCTCTAGAGCTCTAGAGG
'-(24K
)
FIG. 4. Nucleotidesequencesofdeletionsandinsertions constructed in the 13S mRNA of the ElaregionofAdSandproposedtranslation
product. A DNA fragment extending from 2.6 (ClaI)to4.5(HpaI)MU fromdllOl, d1105,andinlO6wasclonedintoplasmidvectorpDR33and
waspropagated inadam strain of E. coli(GM33). The DNAsequencefromtheClaIsitewasdeterminedbythechemicaldegradationmethod
ofMaxamandGilbert(28). ThesequencedatapresentedforAd5wt and hrlweredeterminedbyvanOrmondt et al.(44)andRicciardi etal.
(35),respectively. Dashed linesrepresentsequencesremovedduringsynthesisof the 13S mRNA. Hatchedboxesshow thedeleted(A)DNA
sequence ineach mutant, and the bold line above the sequence indicates insertions (V) of linkerDNA. Parentheses around the protein molecular weight (M.W.) values indicate predicted sizesfromDNAsequence data(44).
actingtostabilizeit inaconditionallyactiveconformationat
37°C, a mutant virus was constructed, inlO6, which
con-taineda16-bp insertionat2.8MUandgeneratedanin-frame
translational termination signal beginning at the newly
in-serted nucleotide 1,013 (Fig. 4). The position of this stop
codonenables the 13S mRNAtoencodeapredicted
truncat-ed protein of 24 kd, containing the same amount ofsense
aminoacidsasdilOl but onlyone missenseamino acid.
Transformation studiesperformed usinginlO6 withCREF
cellsrevealed that it hadaconditional, cold-sensitive
pheno-typesimilartothat of dllOl(Table 1). Thisindicated that the
missense amino acids present in the truncated protein
en-codeddllOl werenotacting tostabilize it in aconditionally
activeform.
Asa meansofmoreaccurately determining what domain
of the 51-kd protein was essential to retain some function, d1105 virus (Fig. 4) was used to infect CREF cells in a
transformationassayat 32 and37°C. Uponinfection withas
muchas30PFU/cell,nodiscernible focideveloped ateither
temperatureforincubation periods extendingup to8weeks
(Table 1). The 69-bp deletion asymetrically located around
2.8 MU distinguished dllO5 from dIlO1 in two ways. First,
the sense amino acids incorporated downstream from the
deletion present in dllO5 appeared unable to influence the
function of the truncated protein. Second, the coding
se-quences upstreamfromthe start of the deletions present in
both dllOl and dllO5 revealed a difference of only five
nucleotides. As a consequence, the truncated protein
en-coded by dllO5 contained two fewer sense amino acids
(codons GAG and CAC at positions 148 and 149; refer to
Table 2). Therefore, the nucleotide difference between the
function ofaprotein activeat37°C andaprotein thatcannot
produce transformationateithertemperaturecouldbe
local-ized to a 5-bp region extending from nucleotides 1,002 to
1,007.
[image:5.612.314.556.405.648.2]Biological properties and integration patterns of mutant
TABLE 1. Comparative transformation of CREF cellsbyAd5wt,
sub309,and Eladeletion and insertionmutants atvarious
temperatures
Incubation Transformation
Expttemp period Fociperplate
(wk) (mean+ SD)h Frequency
Ad5wt
32 6 26±2 2.6x 10-4
37 6 24 3 2.4 x 10-4
sub3O9
32 6 29 5 2.9 x 10-4
37 6 35 3 3.5 x 10-4
dIlO1
32 6 0 <1 x 10-5
37 6 179 ± 7 1.79 x 10-3
32-37
2+4 149±6 1.49x 10-332-37 3 + 3 85 ±2 8.5 x 10-4
37 32 2+4 7±2 7x 10-5
37-32 3+3 42±7 4.2x 10-4
d1105
32 8 0 <1x10-5
37 8 0 <1X10-5
in106
32 6 0 <1 x 10-5
37 6 163 ±5 1.6 x 10-3
aCREF cellswereinfected with 10 PFUof
Ad5wt,
sub309, dIll1,
orin106orwith 30 PFUofd1105percell; after2 hof viraladsorption
at32°C, cellswereresuspendedandreseededat105cells per 60-mm
plate at32or37°C. Cultures were either maintainedat32or37°C continuouslyfor 6to8weeksorshiftedtotheindicatedtemperature
at2or3weekspostinfection. Allcultureswerefixedandstainedat
thetimes indicated.
b Meannumberof transformed coloniesorfociper105 cells. Each groupconsisted ofeight culture plates.
.0
0
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Summary of DNA sequences, proposed amino acid positions, and transformation data of Ad5wt and Ela mutant viruses
Mutation Amino acid atposition': Transformation
Virus Type Site 148 -149-150
-151 -152
Foci'
Cold(bpY' sensitive
Ad5wt glu -his-pro-gly -his +
5hrl Al 1,055 glu -his-pro-gly -his + +
dIlOl A5 1,007-1,013 glu -his-pro-arg-leu + +
inlO6 V16 1,009-1,010 glu -his-pro-leu -stop + +
dllO5 A69 1,002-1,072 asp-ser- leu -cys -tyr
a A,Deletion; V, insertion. Amino acids and location were not determined experimentally, but were inferred from the nucleotide sequence of this gene by van Ormondt et al. (44) and the deletion and insertion mutants presented in Fig. 4.
bAbility to induce focus formation at 37°C.
virus-transformed CREF cellsat32 and37°C. Thepatternsof
viral DNA integration into chromosomal DNA oftwo setsof
cloned dllOl- and inlO6-transformed CREF cell lines were
examined by cleavage with XbaIorBglII followed by DNA
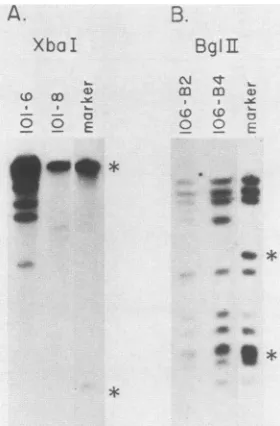
transfer-filter hybridization analysis (40). Cell line 101-6
appearedtocontain dllOlgenomeDNAintegratedatseveral
sites (Fig. 5A), whereas 101-8 appeared to contain a single
viral DNA integration site. BglII cleavage of cellular DNA
isolated from cell lines 106-B2 and106-B4revealedthatmost
of the viral genome was present in these cells, with the
terminal fragments being larger owing to integration (Fig.
SB). Like hrl-transformed CREF cell lines (1), the
integra-tionpatternof the cloned cellswasnotalteredby shifting the
cultures to 32°C and maintaining them at that temperature
(datanot shown).
Whereas the transformation studies showed similarities
amonghrl, dllOl, and in106, itwas important to determine A.
XboI
a)
(0
-.
_ o *
B.
Bg II
._
*_ _
AP wh.
FIG. 5. AnalysisofdllOl and inlO6sequencesin CREF cells.A
10-,ug portion of cellular DNA isolated from the cloned cell lines indicatedwascleaved withXbaI (A)orBgIIl (B),and the fragments
were separated by electrophoresis in0.6% agarose, transferredto
nitrocellulose sheets by blotting, and hybridized to sub3O9 DNA
labeled with32P by nicktranslation (36, 40,45). The marker lane in
(A)representsfivegenomeequivalentsof sub3O9DNAcleaved with
XbaI,and the marker lane in(B)represents thesame quantity of
sub3O9 DNA cleaved withBglII.Thestarred DNA bandsrepresent terminal fragments which are not present in the transformed cell
linesduetoviral DNAintegration.
whether the cloned celllines derived from transformed cell
fociproduced by the latter twoviruses continuedtoexhibit
the cold-sensitivephenotype. Both cell lines 101-6 and 101-8
at37°C had cloning efficiencies inagarthatweresimilartoor
exceeded that of wt3A, an Ad5wt-transformed CREF cell
line (12) (Table 3). When assayed at 32°C, the cloning
efficiency of wt3A remained relatively unchanged, but cell
lines 101-6 and 101-8 were decreased by 73 and 95%,
respectively. Whereas the sizes of the growing colonies of
wt3A cellsat32 and 37°Cwere similar, therewas amarked
decrease in the colony sizes ofmutantvirus-transformed cell
coloniesgrownin32°C when compared with those incubated
at 37°C. In addition, the highagarcloning efficiency of cell
line 101-6at37°C incomparisonwithwt3A, 101-8, and
106-B4maybe duetoanenhanced level of viralgeneexpression,
perhapsas aresultof theintegration ofagreaternumberof
genomecopies (Fig. 5A).
Construction ofplasmidscontaining the ElaorEla andElb
genes from dllOl, hrl, and sub3O9. The studies described
above strongly indicated that the deletionpresentin the Ela
geneof hrlwasin fact thecauseof the conditional
transfor-mation phenotype and that a region around 2.8 MU was
critical for the function of the 51-kdprotein. It remained to
be determined whether the cold-sensitive nature of the
truncated proteins encoded by the mutantviruses required
functions of viral Elbgeneproductsorwaspossibly dueto
aninteraction of the Elaprotein withsomecellularfactor(s).
Totestthesepossibilities, plasmidswereconstructed which
TABLE 3. Cloning efficiencyin soft agarofuninfected, wt5-transformed, dllOl-transformed, and inlO6-transformed CREF
cells grownatvarious temperatures
Cloningefficiencyinagar(%)a
Cellline
370C 32°C
CREF <0.001 <0.001
Ad5wt-3A 45 ±2 38±2
dllOl-6 98 ±1 27±
2b
dllOl-8 47± 5 3.2± 1b
inlO6-B4 53±8 9± 2b
aApproximately 103,104,or105 cellswerepreparedinlow-Ca2+
medium containing0.4%Noble agar and seededat 37 or320Con 0.8% agar base layersprepared in thesamemedium. Plateswerefed
once aweek with2 to 3 mlof0.4% agaron
low-Ca2`
medium,and colonieswerecountedafter3weeksfor37°Cand after5to6weeks for320C cultures. Eachvalueisthemean ± standard deviation of fourplates.b The final sizes of the mutant virus-transformed cell colonies countedat320Cwereconsistently fourtofive times smallerthanthe colonies cultured at370C.
on November 10, 2019 by guest
http://jvi.asm.org/
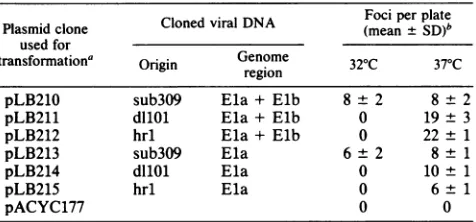
[image:6.612.116.256.410.623.2] [image:6.612.324.562.562.637.2]TABLE 4. Transforming activity in CREF cells ofhybrid plasmids containing ElaorElaand Elb gene sequences ofdIlOl,
hrl, and sub3O9
ClonedviralDNA Fociperplate
Plasmid clone Cloned viral DNA (mean ±SD)b used for
transformation' Origin Genome 32°C 37°C
region
pLB210 sub3O9 Ela + Elb 8± 2 8± 2
pLB211 dIlOl Ela +Elb 0 19± 3
pLB212 hrl Ela+ Elb 0 22 ± 1
pLB213 sub3O9 Ela 6 ±2 8± 1
pLB214 dIlOl Ela 0 10± 1
pLB215 hrl Ela 0 6± 1
pACYC177 0 0
aApproximately106CREF cellsweretransfected with7,ugof the desired plasmid DNA, using the calcium precipitation technique. After 3 h of incubationat37°C, 15%glycerol inphosphate-buffered saline was added, after which cellswereresuspended and reseeded
at
10'
cells per 60-mm plateat32°C for11weeksor at37°C. Cultures were maintainead at 32°Cfor 11 weeks or at 37°C for 8 weeks, at which time the monolayers were fixed, stained, and counted. A 7-,ug portion ofEla-containing plasmids equals 59-,ug genome equiva-lents; 7 ,ug ofEla+ Elb-containing plasmids equals31-p.ggenome equivalents.bMeannumberof transformed colonies per105 cells.
contained theDNA sequencesof eitherEla or Ela and Elb
from dllOl, hrl, and sub3O9, andtherecombinantplasmids wereusedtotransfect CREF cells to notetheir transforma-tionpotentials at32 and37°C.
Theconstruction of the varioushybrid plasmids
contain-ingEla or ElaandElbgene sequencesis describedinFig.
3. Mutant viral Ela and Elb DNA sequences (2.6 to 15.5
MU) were substituted for comparable wt sequences to
generate an intact viral sequence extending from 0 to 15.5
MU. With this approach, a series ofplasmidswere isolated containing the Ela and Elb regions of the desired viral
genomes. Thenewly isolatedplasmid strainswereusedas a
source for viral Ela sequences (O to 4.5 MU), which were
subcloned into plasmidvector pDR33 (Fig. 3B).
Plasmid DNAs were used to transfect CREF cells in a
transformation assay at 32 and 37°C. All of the plasmids
containing
viral DNA sequences werecapable
ofproducing transformed foci at 37°C (Table 4). However, no fociap-pearedat32°C whenanyof the
plasmids
derived fromhrl ordllOl were used. In contrast, sub3O9plasmids transformed CREFcellsatthesame
frequency
at32 or37°C. Therefore, the viraltransformation studies described above correlatedexactly with the results oftheplasmid transfection
experi-ments.
It should be noted thatthe morphology of Ela- and Ela
plus Elb-transformed CREF cells were different.
Trans-formed foci produced by Ela plasmids (pLB213-pLB215)
werefibroblastic and relatively flat. In contrast, sub3O9Ela
plus Elb plasmid-transformed CREFcells were amixture of fibroblastic and
epithelioid
cells, and the cell colonieswereraised.However, foci producedat37°C by plasmids
contain-ingboth Ela and Elb DNAfromhrlordllOl(pLB211 plus
pLB212) were solely fibroblastic but retained the ability to
produce raised colonies. DNA filter-hybridization studies
demonstratredthattransformed CREFcells,whichhad been
transfected with the various plasmid DNAs, contained the
expected viral DNA sequences (data not shown). In
addi-tion,theability ofthe clonedtransformedcell lines to grow
in agar correlated with their abilty to express the
cold-sensitivephenotype: the Ela- and ElaplusElb-transformed
cells derived from hrl or dllOl showed reductions in agar
cloning efficiencies when grown at 32°C as opposed to 37°C
(datanotshown).
DISCUSSION
The data presented in this paper demonstrate that when
frameshift mutations are introducedinto the 13S mRNA of
the Ela gene of Ad5 via deletion or insertion of genetic
material, some of the viral mutants obtained exhibit a
conditional, cold-sensitive transformation phenotype. This
phenotype can be attributed solely to the altered gene
product encoded in the 13S mRNA since the defects were
placed in theintrons ofthe 12S and 9S mRNA transcripts, thus leaving these mRNAs, and hence theirgene products,
unaltered. The initialviral mutantisolated,dllOl, was
engi-neered to contain a 5-bp deletion at 2.8 MU (nucleotides
1,007to1,012),and it wasutilized to investigate: (i) whether hrl might contain a missense mutation, in addition to the
single-base
pair deletion, to account for its conditionallylethal,
cold-sensitivephenotype; and (ii) how much of the51-kd Elaproteinwasrequired to transform cells.
Previous studies had shown that the truncated 28-kd
protein
encoded by the 13S mRNA ofhrl (35) effected theconditionally
functional phenotype for the maintenance of the transformed cell (1, 22). However, it was necessary todetermine whethera mutation other than the known
single-base
pair
deletion in hrl wasproducing
the conditionalphenotype
observed since it is uncommon for a nonsensemutationtodisplayaconditionallylethalphenotype.
Where-as marker rescue
(14)
of the known mutation or backcrossing
of the mutation into a known wt genome is thetraditional
approach
to theproblem,
dllOl answered thisquestion by revealingthataframeshift mutation introduced
47bpupstream from the lesion present in hrl could generate
the same
phenotypic
characteristics. Unlikehrl,
dllOl'smutation was introduced via in vitro
manipulation
andtherefore decreasedthelikelihood ofthere
being
an addition-almutation upstreamfrom theengineered
deletion.It is
important
to note that the cold-sensitive mutantsdescribed in this communication did not exhibit a normal
transformedphenotypeevenwhenmaintainedatthe
permis-sivetemperatureof37°C. Thus, allofthe cells transformed
by hrl, dllOl,
and inlO6 at 37°C werefibroblastic,
asopposed
toepithelioid,
and they weregenerated at afive-fold-higher frequency
thanwas wtvirus. These dataindicate thatatleastonedomain of the truncatedprotein
produced by these virusesmaybeconditionally
functional andthat,whenthis
domain(s)
was reducedby asfewasfivenucleotides, it hadlost itsabilitytofunctionnormally at either temperature. Itwaspreviously
shownthat hrl-transformedcells grown at 32or37°C
maintainthe sameviralDNAintegrationpattern(1)
and that hrl- ordllOl-infected CREF cells shifted afterprolonged periods
at32 to37°Cstill producetransformed cellfoci.These
findings
suggestthatthose eventsthat lead to theintegration
of the viral genome into the host cellchromo-some, and
possibly
the initialexpressionof Ela geneprod-ucts,areunaffectedbythemutationpresent or even enhance
the
ability
of the mutant virus to initiate these events.Studies with these viruses are presently under way to
determine whether an increase in the overall
frequency
ofintegration
is the event responsible forthe highertransfor-mation
frequencies
observed.The
position
of the deletion present in dllOl furthersuggested
thatonlyalimitedregionof the 51-kdproteinwason November 10, 2019 by guest
http://jvi.asm.org/
required to retain partialfunctionality. However, since this
mutant did not encode an in-frame stop codon(TGA) until
nucleotides1,241 to 1,243, it was possible that the additional
missense amino acids incorporated into the truncated
pro-tein werefunctioning to stabilize it in a conditionally active
form. This problem was approached by constructing a
mutant virus, inlO6, so that it contained a 16-bp insertion
betweennucleotides 1,009 and1,010,whichgeneratedan
in-frame stopcodon(TAG) at thenewlycreatedposition1,113
to 1,116 (Fig. 4). Transformation studies with this virus
revealed that it behaved like hrl and dllOl and thus
demon-strated thatthe missense amino acids present in the
truncat-ed protein encodtruncat-ed by dllOl and absent in the protein
encoded byin106were notrequired to retain the conditional
transformation functions observed for either virus.
DNAsequence analysis ofd1105revealed that itcontains a
69-bp deletion extending from nucleotides 1,002 to 1,072.
Unlike all ofthe previous viral mutants tested, this virus was
completely incapable of generating transformed CREF cell
foci atmultiplicities as high as 30PFU/cellregardlessof the
temperature of incubation. Since the deletion present in
d1105 was not symmetrical around the SmaI restriction
endonuclease cleavage site at1,009 bp, this mutant anddllOl
only differed by 5 bp upstream from the start of the deletion.
The mutants' characteristics (Table 2) note that wt5, hrl,
dlO1l, andinlO6retain the sameamino acid sequenceaswt
virus at positions 148 to 150, at which site missense amino
acids are introduced. WhereasdlO1lwas shown tocontaina
5-bp deletion, the first amino acid produced owing to the
frameshift remained a proline at position 150 due to
degen-eracy of theamino acid code. Whetherthishistidine-proline
amino acidsequence is absolutely required for thetruncated
proteins produced by these mutant viruses to function in
transformation, or possibly to maintain its integrity, is
presently being investigated by constructing an additional
viral mutant with a 4-bp deletion at this site. This type of
lesion would result in an amino acid sequence of histidine
and arginine at positions 149 and 150, respectively. It is
important to note that, although H5in500 (7) retains the
histidine-proline amino acids, itis transformation defective
just as are allof the previously isolated viralmutantsthat do
not retain this amino acid sequence (25, 39). However, the
lowfrequencies oftransformation reported bythese
investi-gators,perhapsowingtothedifferentratcells used,maynot
have allowed them to detect the partial transformation
phenotypethat is obtained with the Ela mutantsdescribedin
this communication.
Ho et al. (22)described the isolation of Ad5 mutantsthat
are cold sensitive for replication in HeLa cells. Both dllOl
and
inlO6
also display this type of conditionally host rangelethalphenotype for replication (manuscriptinpreparation).
However, although the final viral yield obtained in HeLa
cells witheither virus at 37°C approaches that of
Ad5wt,
theeclipse period is extended,suggesting that inreplicationasin
transformation only a partial function of the truncated pro-tein is attained at this temperature.
The lesionthat produces a conditional phenotype is
classi-callyamissense mutation at asingle basepair. Althoughour
observations that frameshift mutations brought about by
deletion or insertion of genetic material can yield a
cold-sensitivephenotype are unusual, they are notuniquetothis
system.Pintel et al. (34) isolated adeletion mutantofsimian
virus 40which lacks 81 bp at the C-terminal end of thelarge
T antigen and demonstrated that transformation of rat cells
by this viruswas cold sensitive. These findings were further
extended (33) to show that this virus was heatsensitivefor
viralreplication, whereas, in contrast, the transformed cells
derivedfrom this virus were cold sensitive for maintenance
ofthe transformed phenotype. Bryant and Parsons (6) have
constructed deletion mutants of Rous sarcomaviruses and
haveshown that one mutant, tsCH119, is heat sensitivefor
the maintenance of the transformed phenotype in chicken
cells.
It haspreviously been shown that the Ela geneofAd5(23)
or Adl2 (38) can partially transform rodent cells, whereas
studies thatused a DNAfragment fromAd5 extendingfrom
0 to 8.0 MU revealed that Ela and partial Elb geneproducts
(the 19K and truncated 58K proteins) can effect complete
transformation (16). Those findings suggest that a Elagene
product(s) may act independently or interact with cellular
factors to initiate and maintain the partial transformed phenotype,which alsohas been a proposedfunction forthe
Ela gene product(s)in viralreplication(31,32). The
expres-sion of the Elb-encoded 19K and truncated 58K proteins
mayproduce the completely transformed cells observed by
theirinteracting with Ela proteins or cellular factors or both,
or they may function independently.
Todetermine whether thecold-sensitive phenotype could
beexpressed in the absenceofElb geneproducts,
recombi-nantplasmids wereconstructed which contained the Ela or
Ela and Elb genes ofsub390, hrl, and dllOl, and these were
used to transfect CREF cells in a transformation assay.
Although it has been demonstrated that the processes lead-ing to cell transformation by infection or transfection of susceptible cells can vary (29), our transfection studies
correlated completely with the results of virion infections
(Table 4). The transformation frequencies with plasmids
containingElaor Ela and Elb gene sequences from hrl and
dIlOl werehigher at 37°C thanthose with plasmids derived
from sub3O9, in keeping with the increased transformation
frequency noted when the mutant virions were used. In
addition, the morphology ofthetransformed cells obtained
with the mutant viruses or sub3O9 Ela gene-containing
plasmid was fibroblastic in contrast to the epithelioid cells
produced by the sub3O9 plasmids containing Ela andElb.
Ela gene sequences transfected into CREF cells derived
from hrl or dllOl were unable to induce foci at 32°C, indicatingthat they hadretained theirconditional phenotype in the absence of viral Elbgene expression.
The datadescribed using viruses with specific mutations
intheportion ofthe genomeencodingthe Ela 51-kd
protein
imply that it is this viral gene product that is necessary to
maintaintherecognizedproperties of transformed cells.The
conditionally lethal, cold-sensitive phenotype
expressed by
the mutated genomes, whetherintroduced intoCREF cells
by infection with virions or by transfection with intact
genomes or plasmids, implies that the Ela 51-kd
protein
interacts with a viral (Ela 12S gene product) or a host macromolecule(s) or both. The cold-sensitive phenotype
probably reflects a decreased binding constant at 32°C
betweenthemutants' truncatedproteins and astill
unidenti-fied macromolecule(s). Identification of the components
involved and the characteristics of the viral protein
macro-molecularinteractions should revealcritical clues to
under-standingthe mechanisms of viral transformation.
ACKNOWLEDGMENTS
This workwassupported byPublicHealthServicegrant AI 12052
(H.S.G.) from the National Institute of Allergy and Infectious
on November 10, 2019 by guest
http://jvi.asm.org/
Diseases and grant CTR-1532 (P.B.F.) from the Council for Tobacco Research.
We thank Thomas Shenk forHSsub3O9 and Donald Mills, Lisa Brunet, and David Bechhofer for theirhelp inDNAsequencing.
LITERATURECITED
1. Babiss, L. E., H. S. Ginsberg, and P. B. Fisher. 1983. Cold-sensitiveexpression of transformationbyahostrangemutantof type5adenovirus. Proc. Natl. Acad.Sci. U.S.A. 80:1352-1356. 2. Babiss, L. E., C. S. H. Young, P.B.Fisher,and H.S.Ginsberg. 1983.Expression ofadenovirus Ela and Elb geneproducts and theEscherichiacoli XGPRT gene inKBcells. J. Virol. 46:454-465.
3. Berk, A. J., F. Lee,T.Harrison, J. Williams,and P. A.Sharp. 1979. Pre-earlyadenovirus5genomeproductregulates synthe-sis of early viral messengerRNAs.Cell 17:935-944.
4. Berk, A. J., andP. A.Sharp. 1978. Structure ofadenovirus 2
early mRNAs. Cell 14:695-711.
5. Bolivar, F.,R. L.Rodriquez,P.J. Greene,M.C.Betlash,H. L.
Heynecker, and H.W.Boyer. 1977.Construction and character-ization of new cloning vehicles. II. A multipurpose cloning system. Gene 2:95-113.
6. Bryant, D., andJ. T.Parsons. 1982. Site-directed mutagenesis of the src gene of Rous sarcoma virus: construction and characterization ofadeletionmutanttemperature sensitive for transformation. J. Virol. 44:683-691.
7. Carlock,L.R.,andN. C. Jones. 1981.Transformation-defective
mutant of adenovirus type 5 containing a single altered Ela mRNAspecies. J. Virol. 40:657-664.
8. Chang, A. C. Y., and S. N. Cohen. 1978. Construction and characterization ofamplifiable multicopyDNAcloning vehicles derived from the P1SA cryptic miniplasmid. J. Bacteriol.
134:1141-1156.
9. Chinnadurai, G.,S.Chinnadurai,andJ.Brusca. 1979.Physical mapping ofa large plaque mutation of adenovirus type 2. J.
Virol. 32:623-628.
10. Chow, L. T., T. R. Broker, andJ. B. Lewis. 1979. Complex splicingpatternsofRNAfrom theearlyregions of adenovirus2. J. Mol. Biol. 134:265-303.
11. Clewell, D. B., and D. R. Helinski. 1972. Effect of growth conditionsontheformation of therelaxationcomplexof super-coiledColElDNAandprotein in Escherichia coli.J.Bacteriol. 110:2235-2246.
12. Fisher,P.B.,L. E.Babiss,I. B.Weinstein,and H.S. Ginsberg.
1982. Analysis of type 5 adenovirus transformation with a
cloned rat embryo cell line (CREF). Proc. Natl. Acad. Sci. U.S.A.79:3527-3531.
13. Fisher, P. B., N. I. Goldstein, and I. B. Weinstein. 1979.
Phenotypic properties andtumorpromotor inducedalterations in rat embryo cells transformed by adenovirus. Cancer Res.
39:3051-3057.
14. Frost, E.,andJ. Williams. 1978.Mapping temperature sensitive and host-range mutations of adenovirus type 5 by marker rescue.Virology 91:39-50.
15. Gallimore, P. H., P. A. Sharp, andJ. Sambrook. 1974. Viral
DNA in transformed cells. II. A study of the sequences of adenovirus 2DNAin nine lines of transformed ratcellsusing
specific fragmentsoftheviral genome. J. Mol. Biol. 89:49-72.
16. Graham,F. L.,P. J. Abrahams, C. Mulder, H. L. Heijneker, S.0.Warnaar,F. A.J. DeVries,W.Fiers, andA.J.vanderEb.
1974. Studies on in vitro transformation by DNA and DNA
fragments of human adenovirus andSV40.ColdSpring Harbor Symp. Quant.Biol. 39:637-650.
17. Graham, F. L., T.Harrison, and J. Williams. 1978. Defective transforming capacity of adenovirus type5host-rangemutants.
Virology 86:10-21.
18. Graham,F. L., J. Smiley, W.C. Russell, and R. Nairn. 1977. Characteristics ofahumancell line transformed byDNAfrom human adenovirus type 5. J.Gen. Virol.36:59-72.
19. Graham,F.L., andA.J.van der Eb.1973.Anewtechnique for
the assay of infectivity of human adenovirus type 5 DNA.
Virology52:456-467.
20. Graham, F. L., A. J. van der Eb, and H. L. Heijneker. 1974.Size and location of the transforming region in human adenovirus type5DNA. Nature (London) 251:687-691.
21. Harrison, T., F. Graham, and J. Williams. 1977. Host range mutants ofadenovirus type5defective for growth in Hela cells. Virology77:319-329.
22. Ho, Y.-S., R.Galos, and J. Williams. 1982. Isolation of type S adenovirus mutants with acold-sensitive host-range phenotype: genetic evidence of an edenovirus transformation maintenance function. Virology 122:109-124.
23. Houweling, A., P.J. van der Elsen, and A. J. van der Eb. 1980. Partialtransformation of primary rat cells by the leftmost 4.5% fragment of adenovirus5DNA. Virology 105:537-550. 24. Jones, N., and T. Shenk. 1979. Anadenovirus type5early gene
function regulates expression of other early viral genes. Proc.
Natl. Acad. Sci. U.S.A. 76:3665-3669.
25. Jones, N., and T. Shenk. 1979. Isolation of AdS host-range delection mutants defective for transformation of rat embryo cells. Cell 15:205-214.
26. Kitchingman, G. R., and H. Westphal. 1980. The structure of adenovirus 2 early nuclear andcytoplasmicRNAs.J. Mol. Biol. 137:23-48.
27. Marinus, M. G. 1973. Location of DNA methylation genes on the Escherichia coli K-12 genetic map. Mol. Gen. Genet. 127:47-55.
28. Maxam, A., and W. Gilbert. 1980.Sequencing end-labeled DNA with base-specific chemical cleavages. Methods Enzymol. 65:499-560.
29. McKinnon, R. D., S. Bacchetti, and F. L. Graham. 1982. TnS mutagenesis of the transforming genes of human adenovirus type 5. Gene 19:33-42.
30. Montell, C. E.,E. Fisher, M. Caruthers,and A.J.Berk. 1982.
Resolving the functions of overlapping viral genes by
site-specific mutagenesis at a mRNA splice site. Nature (London) 295:380-384.
31. Nevins, J. R. 1981. Mechanism of activation of early viral
transcription bytheadenovirus Ela gene product. Cell.
26:213-220.
32. Persson, H., H.-J. Monstein, G. Akusjarvi, and L. Phillipson. 1981. Adenovirus early gene products may control viral mRNA accumulationandtranslation in vivo. Cell 23:485-496.
33. Pintel, D., N.Bouck, and G. di Mayorca. 1981. Separation of
lytic and transforming functions of the simian virus 40 A region: two mutantswhich are temperature sensitive for lytic functions
haveoppositeeffects on transformation. J. Virol. 38:518-528. 34. Pintel, D., N. Bouck, G. di Mayorca, B. Thimmuppaya, B.
Swerdlow, and T. Shenk. 1979. SV40 mutant tsA1499 is heat-sensitive for lytic growth but generates cold-sensitive rat cell transformants. Cold Spring Harbor Symp. Quant. Biol.
44:305-309.
35. Ricciardi, R. L., R. L. Jones, C. L. Cepko, P. A. Sharp, and
B. E. Roberts. 1981. Expression of early adenovirus genes requires a viral encoded acidic polypeptide. Proc. Natl. Acad. Sci. U.S.A. 78:6121-6125.
36. Rigby,P.W., J. M. Dieckmann, C. Rhodes, andP.Berg. 1977. Labellingdeoxyribonucleic acid to high specific activity in vitro by nick-translation with DNA polymerase I. J. Mol. Biol. 113:237-251.
37. Russell,D.R., and G. N. Bennett. 1981. Cloningof small DNA fragments containing Escherichia coli tryptophan operon
pro-moterandoperator. Gene 17:9-18.
38. Shiroki, K.,H.Handa, H. Shimojo, S. Yano, S. Ojima, and K. Fujinaga. 1977. Establishment and characterization of rat cell lines transformed by restriction endonuclease fragments of adenovirus 12 DNA. Virology 82:462-471.
39. Solnick, D., M. A. Anderson. 1982. Transformation-deficient
adenovirusmutantdefective in expression of region 1A but not region 1B. J. Virol. 42:106-113.
40. Southern, E. M. 1975. Detection of specific sequences among DNAfragments by gel electrophoresis. J. Mol. Biol.
38:503-517.
41. Spector,D.J., M. McGrogan,and H.J. Raskas. 1978.
on November 10, 2019 by guest
http://jvi.asm.org/
tion oftheappearanceofcytoplasmicRNAsfrom region1of the
adenovirus-2genome. J. Mol. Biol. 126:395-414.
42. Stow, N. 1981. Cloning ofaDNAfragment fromthe left-hand
terminus oftheadenovirus type 2genome and itsuse in
site-directed mutagenesis. J. Virol.37:171-180.
43. vander Eb,A. J., C. Molder, F.L. Graham,and A.Houweling.
1977. Transformation with specific fragments of adenovirus
DNAs. I. Isolation of specific fragments with transforming
activity of adenovirus2and 5 DNA.Gene2:115-132. 44. vanOrmondt, H., J. Maat, and C. P. vanBeveren. 1980.The
nucleotide sequence of the transforming early region El of adenovirustype5 DNA.Gene 11:299-309.
45. Wahl, G. M., M. Stern, and G. R. Stark. 1979.Efficient transfer
oflargeDNAfragments fromagarosegelsto diazobenzyloxy-methyl paper and rapid hybridization using dextran sulfate.
Proc.Natl. Acad. Sci. U.S.A.76:3683-3687.