J
OURNAL OFV
IROLOGY,
0022-538X/00/$04.00⫹0
Apr. 2000, p. 3418–3422
Vol. 74, No. 7
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Assembly and Processing of Human Immunodeficiency Virus
Gag Mutants Containing a Partial Replacement of the Matrix
Domain by the Viral Protease Domain
CHIN-TIEN WANG,* YEN-CHIOU CHOU,
ANDCHIEN-CHENG CHIANG
Institute of Clinical Medicine, National Yang-Ming University School of Medicine, and Department of Medical
Research and Education, Taipei Veterans General Hospital, Taipei 112, Taiwan, Republic of China
Received 4 August 1999/Accepted 20 December 1999
We constructed human immunodeficiency virus (HIV) mutants by replacing the matrix domain with
se-quences encoding the viral protease or p6* and protease. The chimeras retaining matrix myristylation and
processing signals underwent efficient autoprocessing with severely defective particle budding. The budding
defects of the chimeras were rescued by suppressing the chimera protease activity either through addition of
an HIV protease inhibitor or through inactivating the chimera protease via a substitution mutation of the
catalytic aspartic acid residue. This resulted in the release of chimeric virus-like particles with the density of
a wild-type retrovirus particle. In addition, the assembly-competent but processing-defective chimeras
pro-duced proteolytically processed particles with significant reverse transcriptase activity when a downstream
native
pol
gene was present. These results suggest that HIV has the potential to adapt heterologous sequences
in place of the matrix sequence without major effects on virus-like particle budding. In addition, the positions
of the protease and substrate accessibility may contribute significantly toward avoiding a premature Gag or
Gag-Pol process, which leads to severe defects in both particle budding and incorporation.
The structural proteins of all retroviruses, including human
immunodeficiency virus (HIV), are encoded by the
gag
genes
(2, 3, 5, 23). During or shortly after virus budding, the HIV
Gag precursor Pr55 is cleaved by the
pol
-encoded protease
(PR) into four major products: the matrix (p17; MA), capsid
(p24; CA), nucleocapsid (p7; NC), and C-terminal p6 protein
(4, 7, 9, 11). The
pol
product is translated as a Pr160
gag-polfusion protein by a ribosomal frameshifting mechanism that
occurs at a frequency of about 5% during translation of Gag
(6). The relatively low level of Gag-Pol is thought to avoid
premature Gag processing so that Gag assembly can proceed.
Mechanisms of PR activation are unclear; it is proposed that
PR dimerization, a prerequisite for PR activation, is promoted
by the Gag domains (10, 20, 26). PR, once activated,
auto-cleaves from Gag-Pol and subsequently processes Gag and Pol
into mature products. Within Pr160
gag-pol, C-terminal p6 is
truncated and replaced by a domain referred to as p6* (13).
p6*, adjacent to PR, separates NC from Pol. A number of
studies suggest that p6* may be functionally involved in the
regulation of PR activity (14, 17, 27).
It has been demonstrated that chimeras derived from a
re-placement of the HIV or Rous sarcoma virus C-terminal
gag
sequences by foreign protein sequences can still direct
virus-like particle assembly and release (21, 22, 25). In this study, we
substituted the HIV-1 PR coding sequence for MA and
ana-lyzed the assembly and processing of the resultant chimeric
proteins. The p6*-PR and PR sequence fragments were
am-plified by PCR using primers containing a
Cla
I and
Sal
I
re-striction site in the 5
⬘
and 3
⬘
primers, respectively. The
PCR-generated fragments then were treated with
Cla
I and
Sal
I and
used to replace the fragment from
Cla
I (HIV nucleotide [nt]
[image:1.612.311.546.382.630.2]* Corresponding author. Mailing address: Department of Medical
Research and Education, Taipei Veterans General Hospital, No. 201,
Sec. 2, Shih-pai Rd., Shih-pai, Taipei 11217, Taiwan, Republic of
China. Phone: 886-2-2871-2121, ext. 2655. Fax: 886-2-2874-2279.
E-mail: [email protected].
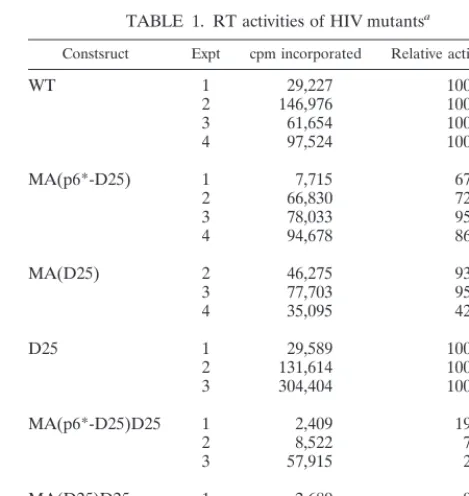
TABLE 1. RT activities of HIV mutants
aConstsruct Expt cpm incorporated Relative activity (%)b
WT
1
29,227
100
2
146,976
100
3
61,654
100
4
97,524
100
MA(p6*-D25)
1
7,715
67
2
66,830
72
3
78,033
95
4
94,678
86
MA(D25)
2
46,275
93
3
77,703
95
4
35,095
42
D25
1
29,589
100
2
131,614
100
3
304,404
100
MA(p6*-D25)D25
1
2,409
19
2
8,522
7
3
57,915
2
MA(D25)D25
1
2,689
9
2
5,892
5
3
21,414
2
aSupernatants were prepared and RT assays were performed as described in
Materials and Methods. For each sample, virus-associated Gag or chimeric protein levels were quantitated by scanning mutant and WT Pr55, p41, and 24 or p24-associated band densities from immunoblots. Results of four separate trans-fection experiments are given.
bRT activities of the processing-defective chimeric mutants
MA(p6*-D25)D25 and MA(MA(p6*-D25)D25 were compared with that of their parental PR-defective mutant D25. Relative activities were determined as percentages of WT or D25 activities (100%) by the equation 100⫻[(mutant counts per minute⫺ background)/mutant Gag protein⫻WT or D25 Gag protein densitometry units/ (WT or D25 counts per minute⫺background)]. All RT activities with counts per minute were at least threefold over the background level (730⫾239).
3418
on November 9, 2019 by guest
http://jvi.asm.org/
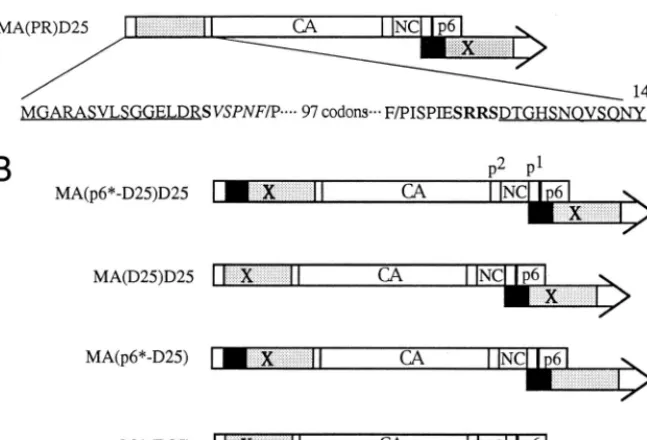
FIG. 1. Schematic presentation of the WT and mutant HIVgpt constructs. Mature WT processed Gag proteins and the p6ⴱ(black) and PR (stippled) domains of
polare indicated. The “X” indicates a PR-defective point mutation (D253Asn). (A) The D25 mutant, which contains a substitution of an Asn residue for the PR catalytic Asp residue, is defective in Gag processing. The MA(p6ⴱ-PR)D25 mutant contains a deletion of 105 codons and a replacement of the p6ⴱ-PR coding sequence in the MA protein. The myristylation signal residues and a few residues in the C terminus of MA remain intact (underlined). Changed or added codons (boldfaced) and residues in the N and C termini of the PR domain are indicated. Upstream of PR, there are 45 codons (italics) of the p6ⴱdomain starting from the N-terminal 12th codon, K. The MA(PR)D25 mutant is identical to the MA(p6ⴱ-PR)D25 mutant except that it contains only five C-terminal codons of p6ⴱ. Instead of having 132 codons as in the WT MA protein, the MA(p6ⴱ-PR) and the MA(PR) constructs contain a total of 182 and 142 codons in their MA regions, respectively. (B) Mutant constructs were derived from the constructs shown in panel A. MA(p6ⴱ-D25)D25 and MA(D25)D25 were identical to MA(p6ⴱ-PR)D25 and MA(PR)D25, respectively, except that the former two contain the PR-defective mutations (D25) in their chimera PR fragments. Recombination of the WT with MA(p6ⴱ-D25)D25 and MA(D25)D25 yielded MA(p6ⴱ-D25) and MA(D25), respectively.
on November 9, 2019 by guest
http://jvi.asm.org/
831) to
Sal
I (nt 1147) of an HIV
gag
mutant that contained a
Sal
I linker at nt 1147 (18). To assess the proteolytic activities of
the inserted PR domains, the chimeric constructs were
sub-cloned into an HIV PR-defective mutant, D25, of which the
PR catalytic residue Asp was replaced with Asn. The resultant
construct was designated MA(p6*-PR)D25 or MA(PR)D25
(Fig. 1A). The backbone of all mutant constructs was HIV gpt,
which carries simian virus 40
ori
and
gpt
genes in the
env
region
(12). Wild-type (WT) and mutant HIVgpt plasmids were
trans-fected into 293T cells. Expression and release of HIV Gag
proteins were probed by immunoblotting using an anti-p24
gagmonoclonal antibody (1, 19). As shown in Fig. 2, the WT Pr55,
the p41, and the mature p24
gagproteins were detected in the
medium and in cell samples (lanes 3 and 9). A major band
representing Pr55
gagwas seen in the medium and cell samples
of D25 (lanes 2 and 8). In contrast, chimeric proteins derived
from processed MA(p6*-PR)D25 or MA(PR)D25 were
de-tected only in the cell samples (lane 10 or 11, respectively). A
faint band corresponding to Pr55
gagobserved in the
MA(p6*-PR)D25 medium sample (Fig. 2, lane 4) may have resulted
from a spillover from the adjacent WT sample because it was
not seen in any repeat experiments.
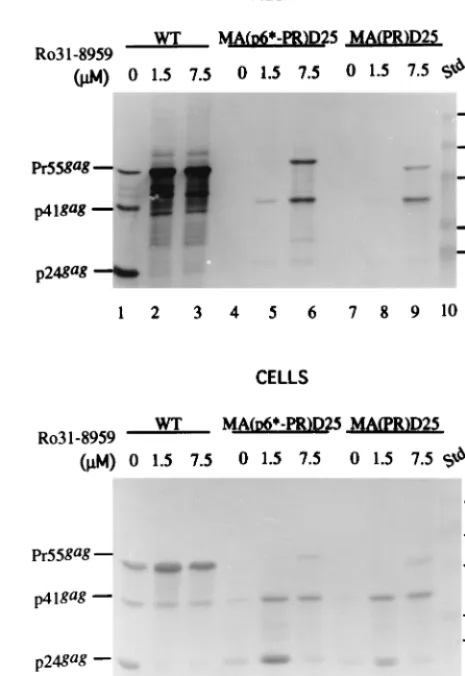
To test whether the inability of the chimeras to release from
cells is due to PR-mediated premature autoprocessing (8, 24),
we added an HIV-1 PR inhibitor, Ro31-8959 (15), to the WT
and chimera transfectants. Figure 3 shows that proteolytic
Pr55
gagprocessing was significantly suppressed in the presence
of the PR inhibitor (lanes 2 to 3 and 12 to 13) compared with
that of untreated samples (lanes 1 and 11). The levels of
re-leased chimeric proteins correlated with the degree of the PR
activity suppression (Fig. 3, lane 6 versus lane 5, and lane 9
versus lane 8). The expected chimera intermediates p6*, PR,
and CA are absent in Fig. 2 and 3; instead, a band migrating
with WT p41
gagis readily observed. This might result from
altered PR preferential cleavage sites. Alternatively, the p41
gagchimera was derived from the incompletely cleaved product
CA-NC-p6 (11). Further experiments are required to test this
proposition.
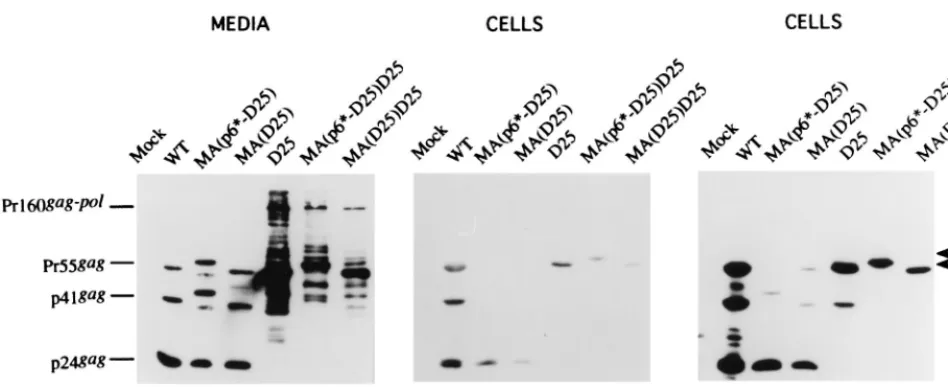
[image:3.612.56.288.72.230.2]To further confirm that suppression of the PR activity
pro-motes chimera release, the PR-inactivating mutation D25 was
introduced into the chimeras MA(p6*-PR)D25 and MA(PR)
D25, yielding constructs MA(p6*-D25)D25 and MA(D25)
D25, respectively (Fig. 1B). To test whether a normal PR
downstream of the chimeric mutations could functionally
com-pensate for the proximal, catalytically inactivated chimera PR,
chimeras MA(p6*-D25) and MA(D25) were constructed by
placing the native HIV
pol
gene downstream of the chimeras
(Fig. 1B). As shown in Fig. 4, chimeras MA(p6*-D25)D25 and
MA(D25)D25 were assembled and released efficiently, at a
level at least 1.7-fold higher than that of D25. Interestingly,
chimeras MA(p6*-D25) and MA(D25) exhibited an efficient
processing profile (Fig. 4, lanes 10, 11, 17, and 18) and had,
FIG. 2. Expression and processing of the chimeric proteins. 293T cells weretransfected with the designated constructs. At 48 h posttransfection, cells and supernatants were collected for protein analysis. Supernatant samples (lanes 1 to 5) corresponding to 50% of the total samples and cell samples (lanes 7 to 11) corresponding to 5% of the total samples were fractionated by sodium dodecyl sulfate–10% polyacrylamide gel electrophoresis and electroblotted onto a nitro-cellulose filter. HIV p24gagand p24gag-associated chimeric proteins were
de-tected with mouse anti-p24gagmonoclonal antibody at a 1:5,000 dilution,
[image:3.612.314.547.84.422.2]fol-lowed by a secondary alkaline phosphatase-conjugated sheep anti-mouse antibody at a 1:5,000 dilution, and alkaline phosphatase activity was determined. Positions of standard (Std.) molecular size markers (lanes 6 and 12) are indicated on the right, and those of HIV Gag proteins Pr55, p41, and p24 are shown on the left.
FIG. 3. Release of the chimeras into the medium in the presence of an HIV PR inhibitor. 293T cells grown on 10-cm-diameter dish plates were transfected with the WT, MA(p6ⴱ-PR)D25, and MA(PR)D25 HIVgpt constructs. At 18 h posttransfection, cells were split equally onto three 10-cm-diameter dishes and treated, respectively, with 0M (lanes 1, 4, 7, 11, 14, and 17), 1.5M (lanes 2, 5, 8, 12, 15, and 18), and 7.5M (lanes 3, 6, 9, 13, 16, and 19) concentrations of the HIV PR inhibitor Ro31-8959. Four hours later, the culture supernatants were removed and replaced with medium plus the designated concentration of the PR inhibitor. At 48 h after addition of the PR inhibitor, culture supernatants and cells were collected for protein analysis. Samples were fractionated by sodium dodecyl sulfate–10% polyacrylamide gel electrophoresis and subjected to immunoblot analysis with anti-p24gagantibody. Std., standards (lanes 10 and 20).
Positions of the molecular size markers are indicated on the right, and those of the HIV Gag proteins Pr55, p41, and p24 are shown on the left.
3420
NOTES
J. V
IROL.
on November 9, 2019 by guest
http://jvi.asm.org/
respectively, three- and sevenfold (lanes 3 to 4 and 10 to 11)
higher levels of release efficiency than the WT (lanes 2 and 9).
Sucrose density gradient fractionation analysis indicated that
all the mutants had a WT retrovirus particle density of 1.16 to
1.18 g/ml (data not shown). To further assess the particle
incorporation of the chimera-Pol fusion proteins, the
particle-associated RT activity of the assembly-competent chimeras
was assayed using exogenous templates (19). Because particle
processing can affect the RT assay (16), RT activities of the
processing-defective chimeras MA(p6*-D25)D25 and MA
(D25)D25 were compared in parallel with those of D25. As
shown in Table 1, the chimeras MA(p6*-D25) and MA(D25)
possessed significant RT activity at a level just over 50% of that
of the WT. Surprisingly, the processing-defective chimeras,
MA(p6*-D25)D25 and MA(D25)D25, exhibited relatively low
RT activity; all levels were below 20% of the level shown by
D25 in three independent experiments. Because the results
shown in Fig. 4 indicate that the chimeric particles contained
significant levels of chimera-Pol (lanes 6 and 7), the low RT
activities of MA(p6*-D25)D25 and MA(D25)D25 were less
likely due to insufficient Pol incorporation. Inaccessibility of
substrates to the chimera-Pol construct and/or impaired
enzy-matic activity due to the chimeric mutations might account for
the low RT activity.
This work was supported by grant NSC88-2314-B010-075 from the
National Science Council and, in part, by grant DOH88-DC-1020 from
the Ministry of Health, Taipei, Taiwan.
The hybridoma clone 183 H12-5C was a gift provided by the AIDS
Research and Reference Reagent Program, Division of AIDS, NIAID,
from Bruce Chesebro. The HIV-1 protease inhibitor Ro31-8959 was
kindly provided by Hoffmann-LaRoche (Switzerland).
REFERENCES
1.Chen, Y.-L., P.-W. Ts’ai, C.-C. Yang, and C.-T. Wang.1997. Generation of infectious virus particles by transient co-expression of human immunodefi-ciency virus type 1gagmutants. J. Gen. Virol.78:2497–2501.
2.Freed, E. O.1998. HIV Gag proteins: diverse functions in the virus life cycle. Virology251:1–15.
3.Gelderblom, H. R.1991. Assembly and morphology of HIV: potential effect of structure on viral function. AIDS5:617–638.
4.Henderson, L. E., M. A. Bowers, R. C. Sowder II, S. A. Serabyn, D. G.
Johnson, J. W. Bess, Jr., L. O. Arthur, D. K. Bryant, and C. Fenselau.1992. Gag proteins of the highly replicative MN strain of human immunodeficiency virus type 1: posttranslational modifications, proteolytic processing, and complete amino acid sequences. J. Virol.66:1856–1865.
5.Hunter, E.1994. Macromolecular interactions in the assembly of HIV and other retroviruses. Semin. Virol.5:71–83.
6.Jacks, T., M. D. Power, F. R. Masiarz, P. A. Luciw, P. J. Barr, and H. E. Varmus.1988. Characterization of ribosomal frameshifting in HIV-1 gag-pol expression. Nature (London)331:280–283.
7.Kaplan, A. H., M. Manchester, and R. Swanstorm.1994. The activity of the protease of human immunodeficiency virus type 1 is initiated at the mem-brane of infected cells before the release of viral proteins and is required for release to occur with maximum efficiency. J. Virol.68:6782–6786. 8.Krausslich, H.-G.1991. Human immunodeficiency virus proteinase dimer as
component of the viral polyprotein prevents particle assembly and viral infectivity. Proc. Natl. Acad. Sci. USA88:3213–3217.
9.Leis, J., D. Baltimore, J. B. Bishop, J. Coffin, E. Fleissner, S. P. Goff, S. Oroszlan, H. Robinson, A. M. Skalka, H. M. Temin, and V. Vogt.1988. Standardized and simplified nomenclature for proteins common to all ret-roviruses. J. Virol.62:1808–1809.
10. Louis, J. M., N. T. Nashed, K. D. Parris, A. R. Kimmel, and D. M. Jerina.
1994. Kinetics and mechanism of autoprocessing of human immunodefi-ciency virus type 1 protease from an analog of the Gag-Pol polyprotein. Proc. Natl. Acad. Sci. USA91:7970–7974.
11. Mervis, R. J., N. Ahmad, E. P. Lillehoj, M. G. Raum, F. H. R. Salazar, H. W. Chan, and S. Venkatesan.1988. Thegaggene products of human immuno-deficiency virus type 1: alignment within thegagopen reading frame, iden-tification of posttranslation modifications, and evidence for alternativegag
precursors. J. Virol.62:3993–4002.
12. Page, K. A., N. R. Landau, and D. R. Littman.1990. Construction and use of a human immunodeficiency virus: vector for analysis of virus infectivity. J. Virol.64:5270–5276.
13. Partin, K., H. G. Krausslich, L. Ehrlich, E. Wimmer, and C. Carter.1990. Mutational analysis of a native substrate of the human immunodeficiency virus type 1 proteinase. J. Virol.64:3938–3947.
14. Partin, K., G. Zybarth, L. Ehrlich, M. DeCrombrugghe, E. Wimmer, and C. Carter.1991. Deletion of sequences upstream of the proteinase improve the proteolytic processing of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA88:4776–4780.
15. Roberts, N. A., J. A. Martin, D. Kinchington, A. V. Broadhurst, J. C. Craig, J. B. Duncan, S. A. Galpin, B. K. Handa, J. Kay, A. Krohn, R. W. Lambert, J. H. Merrett, J. S. Mills, K. E. B. Parkes, S. Redshaw, A. J. Ritchie, D. L. Tayor, G. J. Thomas, and P. J. Machin.1990. Rational design of peptide-based HIV proteinase inhibitors. Science248:358–361.
16. Srinivasakumar, N., M.-L. Hammarskjold, and D. Rekosh.1995. Charac-terization of deletion mutations in the capsid region of human immunode-ficiency virus type 1 that affect particle formation and Gag-Pol precursor incorporation. J. Virol.69:6106–6114.
17. Tessmer, U., and H.-G. Krausslich.1998. Cleavage of human immunodefi-FIG. 4. Assembly and processing of chimeric particles. 293T cells were transfected with the designated plasmid. At 48 to 72 h posttransfection, supernatants and cells were prepared for Western immunoblotting. HIV-1 CA-associated proteins were detected by an enhanced-chemiluminescence detection system (Amersham). The primary antibody was an anti-p24gagmonoclonal antibody used at a 1:5,000 dilution. The secondary antibody was a sheep anti-mouse horseradish peroxidase-conjugated
antibody used at a 1:5,000 dilution. Positions of chimeric protein precursors are indicated on the right (arrows), and those of the HIV Gag proteins Pr55, p41, and p24 are shown on the left.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.65.539.73.266.2]ciency virus type 1 proteinase from the N-terminally adjacent p6* protein is essential for efficient Gag polyprotein processing and viral infectivity. J. Vi-rol.72:3459–3463.
18.Wang, C.-T., and E. Barklis.1993. Assembly, processing, and infectivity of human immunodeficiency virus type 1 Gag mutants. J. Virol.67:4264–4273. 19.Wang, C.-T., H.-Y. Lai, and J.-J. Li. 1998. Analysis of minimal human immunodeficiency virus type 1gagcoding sequences capable of virus-like particle assembly and release. J. Virol.72:7950–7959.
20.Weber, I. T.1990. Comparison of the crystal structures and inter subunit interactions of human immunodeficiency virus and Rous sarcoma virus pro-tease. J. Biol. Chem.265:10492–10496.
21.Weldon, R. A., Jr., C. R. Erdie, M. G. Oliver, and J. W. Wills.1990. Incor-poration of chimeric Gag protein into retroviral particles. J. Virol.64:4169– 4179.
22.Weldon, R. A., and J. W. Wills.1993. Characterization of a small (25-kilodalton) derivative of the Rous sarcoma virus Gag protein competent for
particle release. J. Virol.67:5550–5561.
23. Wills, J. W., and R. C. Craven.1991. Form, function, and use of retroviralgag
proteins. AIDS5:639–654.
24. Xiang, Y., T. W. Ridky, N. K. Krishna, and J. Leis.1997. Altered Rous sarcoma virus Gag polyprotein processing and its effects on particle forma-tion. J. Virol.71:2083–2091.
25. Zhang, Y., H. Qian, Z. Love, and E. Barklis.1998. Analysis of the assembly functions of the human immunodeficiency virus type 1 Gag protein nucleo-capsid domain. J. Virol.72:1782–1789.
26. Zybarth, G., and C. Carter.1995. Domains upstream of the protease (PR) in human immunodeficiency virus type 1 Gag-Pol influence PR autoprocessing. J. Virol.69:3878–3884.
27. Zybarth, G., H.-G. Krausslich, K. Partin, and C. Carter.1994. Proteolytic activity of novel human immunodeficiency virus type 1 proteinase proteins from a precursor with a blocking mutation at the N terminus of the PR domain. J. Virol.68:240–250.
3422
NOTES
J. V
IROL.
on November 9, 2019 by guest
http://jvi.asm.org/