JOURNALOFVIROLOGY,Aug.1975,p.366-387 Copyright01975 AmericanSocietyforMicrobiology
Vol. 16,No. 2 Printed inU.S.A.
Structure and
Composition
of the Adenovirus
Type
2
Core
DENNIS T. BROWN, MONIKA WESTPHAL, BYRON T. BURLINGHAM,I UTE WINTERHOFF,
AND WALTER DOERFLER*
InstituteofGenetics, University ofCologne, Cologne,Germany
Received forpublication 1 April 1975
Thestructure and composition ofthe core ofadenovirus type 2 wereanalyzed
by electron microscopy and biochemicaltechniquesafterdifferentialdegradation
ofthe virionbyheat,by pyridine, orby sarcosyltreatment. Innegativelystained
preparationspurified sarcosyl cores revealspherical subunits of21.6-nm
diame-terinthe electronmicroscope.It issuggestedthat these subunitsareorganizedas
anicosahedronwhich has its axesofsymmetrycoincident with thoseofthe viral
capsid. The subunits are connected by the viral DNA molecule. The sarcosyl
cores contain the viral DNA and predominantly the arginine/alanine-rich core
polypeptide VII. When sarcosyl cores arespread onaproteinfilm, tightlycoiled
particles are observed whichgradually unfold giving riseto arosette-like pattern
due to the uncoiling DNA molecule. Completely unf'olded DNA molecules are
circular.Pyridine coresconsist ofthe viral DNA and polypeptides Vand VII. In
negatively stained preparations of pyridine cores the subunit arrangement
apparent inthesarcosylcoresismaskedbyanadditionalshell whichisprobably
formed bypolypeptide V. In freeze-cleaved preparations ofthe adenoviriontwo
fracture planes can'be recognized. One fracture planeprobably passes between
the outer capsid of' the virion and polypeptide V exposing a subviral particle
whichcorrespondstothepyridinecore.The second fractureplane observed could be located between polypeptide Vand the polypeptide VII-DNA complex, thus
uncovering a subviral structure which corresponds to the sarcosyl core. In the
sarcosyl core polypeptide VII is tightly bound to the viral DNA which is
susceptible todigestion with DNase. The restrictionendonuclease EcoRI cleaves
the viral DNA in the sarcosyl cores into the six specific f'ragments. These
fragments can be resolvedon polyacrylamide-agarose gels provided the sarcosyl
cores aretreatedwith pronase afterincubationwith the restriction endonuclease.
When pronasedigestionisomitted,acomplexofthe terminalEcoRIfragments of
adenovirus DNA and protein can be isolated. From this complex the terminal
DNAfragmentscanbe liberatedafter pronasetreatment.The complex described
is presumably responsible for the circularization of the viral DNA inside the
virion. The nature of' the protein(s) involved in circle formation has not
yet been elucidated.
The chemical composition ofthe adenovirus
core has been studied intensively. The core
contains the viral DNA and two different
pro-teins, polypeptidesVand VII(17, 18, 22, 23, 32,
33, 34) accordingtothe nomenclatureofMaizel
et al. (26). Polypeptide VII is the
alanine/argi-nine-rich protein (17, 22, 32, 34, 38) which
contains 18 to
19%.0
of alanine and 21 to 23% ofarginine residues. The core proteins have been
isolated by several methods (17, 18, 22, 33, 34)
and have been characterized as to molecular
weight and number of polypeptide chains per
virion (17, 18). Polypeptides V and VII have
molecular weightsof48,500 and 18,500,
respec-'Present address: Division of Bioldgy, Kansas State
University,Manhattan,Kan. 66506.
tively, and itisestimated thatthere are 180 and
1,070molecules per virion, respectively (17, 18,
32, 33, 34). Polypeptide VII comprises
approxi-mately
14%7r
ofthe viral protein, and it has beencalculated that the arginine residues of this protein are sufficient to cover approximately
60% ofthe phosphate charges of the viral DNA
(32). Laver (22) has suggested that the alanine/
arginine-rich protein molecules are evenly dis-tributed along the DNA molecule.
As yet, the arrangement ofthe core insidethe
adenovirion and the relation between the viral
DNA and thecore proteins have received little
attention. Robinson et al. (36) and Doerfler et
al. (11) have provided evidence that the viral
DNAinthe virionis keptina circular
configu-366
on November 10, 2019 by guest
http://jvi.asm.org/
CORES FROM ADENOVIRUS TYPE 2 367
ration perhaps through an as yet unidentified
"linker protein" (36). Everitt et al. (18) and
Everitt and Philipson (17) have proposed that
the core polypeptide V is associated both with
the viral DNA and the penton bases, whereas
polypeptide VII is tightly bound to the viral
DNA.
Recently, Everittetal. (16) have developeda
refined model of the type 2 adenovirion. This
model is based on results obtained by
en-zymatic iodination of intact and disrupted
vi-rions, by immunoprecipitation of intact virions
with specific antisera, and by chemical
cross-linkage of the structural proteins. These results suggest that polypeptides V, VI, VII, and VIII
are located inside the virion. Polypeptide V is
found in close proximitytothe penton-bases,to
hexons, andto protein IIIa. Polypeptide V can
be chemically cross-linked also to polypeptide
VII, hence some molecules of polypeptide V
mustbe located closetopolypeptide VII.
In the present study, the ultra structure of
adenovirus cores was investigated by electron
microscopy offreeze-etched preparations of
vi-rions in infected cells and by electron
micros-copy of negatively stained cores isolated by
severalprocedures.The data obtainedsupporta
model oficosahedral arrangementofthe
DNA-polypeptideVIIcomplex inside the virion.
Sar-cosyl cores of adenovirus type 2 consist ofthe
viral DNA and polypeptide VII predominantly. In contradistinction, pyridine cores (32, 33)
contain viral DNA andpolypeptides Vand VII.
Acomparison of thecompositionofsarcosyland
pyridine cores therefore suggests that the type
ofassociation of thesetwopolypeptideswiththe
coremust befundamentally different. Sarcosyl
cores are tightly packed, gradually unfold on
storage,andcontain circular viral DNAwhich is
sensitive both to pancreatic DNase and the
EcoRI restriction endonuclease. Evidence is
presented that the terminal EcoRI
endonucle-asefragmentsofadenovirustype2DNA, A and
C,arelinkedbyprotein.Acomplexbetween the
DNA termini andprotein has been isolated.
MATERIALS ANDMETHODS
Media and solutions. Cells were grown in Eagle medium or Eagle medium modified for suspension cultures(15) supplementedwith 10% calfserum.Calf serumwaspurchasedfromFlowLaboratoriesorfrom Grand Island Biological Laboratories. The
composi-tion of phosphate-buffered saline (PBS) was de-scribedpreviously (14); TEis 0.01 M Tris-hydrochlo-ride, pH7.2to7.5, 0.001 M EDTA; CsClsolution A consisted of 15gofCsCl (Merck, Darmstadt)and 10 ml of TE. Buoyant CsCl solution contained ngrams
of CsCl and n/2 ml of 0.02 M Tris-hydrochloride, pH8.0. Forthedeterminationofradioactivity
toluene-methanol- and toluene-based scintillators were used which consisted of 5 g of 2,5-diphenyloxazole (Merck, Darmstadt) and 0.3 g of 2,2'-p-phenylene-bis-(5-phenyloxazole) (Merck, Darmstadt) per liter of a 1:1 toluene methanol mixture or of toluene. Electro-phoresis buffer (TEB buffer) consisted of 0.089 M Tris-hydrochloride, 0.089 M boric acid, 0.0025 M EDTA, 0.01 M propylamine, and 0.5% sarcosyl.
Cells and virus. KB cells(CCL17fromthe Ameri-can Type Culture Collection) were grown in
mono-layer cultures in Eagle medium (15) or in Eagle medium for suspension cultures both supplemented with 10% (vol/vol) calf serum. Human adenovirus type 2(Ad2) was propagated in KB cells in suspension cultures andwaspurified aspublishedpreviously (10, 11, 13). The production of Ad2 radioactivelylabeled with [6-_Hlthymidine, [14C formate-sodiumor a mix-ture of 3H-labeled amino acids described elsewhere (10, 11, 13). Allradioactivelylabeled compounds were purchased from Amersham-Buchler, Braunschweig, Germany.
Viral DNA. Adenovirus DNAwasextracted from CsCl-purified virionsbymethodsreported previously (5, 11, 13).
Infection of cells for electron microscopy. KB cells growinginmonolayercultureson 100-mm
diam-eter plastic dishes were infected with CsCl-purified
Ad2 at a multiplicity of 50 to 100 PFU/cell as describedpreviously (3).
Preparation of sarcosyl cores from adenovirions. (i) Equilibrium sedimentation in CsCl density gradients. Between 5 and 10optical density unitsat
260nm ofCsCl-purified Ad2("4C-labeled)in 0.3ml of buoyant CsCl solution were mixed with0.07mlof 10%
sarcosyl (N-lauroylsarkosin sodium salt)in waterand
0.63ml of TEandincubatedatambient temperature
for 30 min. In some experiments 5jigof 3H-labeled Ad2 DNA was added as density reference.
Subse-quently, this mixturewaslayeredontop of 3.5to4.0
ml ofCsCl solution A inanitrocellulose tube of the SW56 rotor and centrifuged to equilibrium in an
L2-65B Beckman ultracentrifuge at 37,000 rpm and
4C for at least 60 h. After centrifugation, 5-drop
fractions were collected. Refractive indices of every tenth fractionweredeterminedin aZeiss
refractome-ter.Radioactivitywasmeasuredin 5- to
10-Mgl
aliquotsofeach fraction.
(ii) Gel filtration on Sepharose 2B columns.
Sarcosyl cores were also obtained by adjusting the
CsClpurifiedAd2preparationto0.5%sarcosylinTE
and passing this mixtureover aSepharose2Bcolumn (1 cm indiameter by 10 to 15cm in length)
equili-brated with thesamebuffer. The columnwas devel-opedat 4 C with0.5%sarcosylinTEat aflowrateof2 ml/h.
Preparation of pyridine cores. Pyridine cores wereprepared accordingtotheprocedureofPrageet al. (33, 34). Ad2 ("IC-labeled), which had been
purified byequilibrium centrifugationinCsCldensity
gradients, wasdialyzed against0.005MTris, pH 8.1,
and disrupted in 10% pyridine and the cores were
subsequently isolatedbyzone sedimentationon10to
25% sucrosedensitygradientsin 0.002M
Tris-hydro-chloride, pH 7.5, and0.0002 MEDTA. Thesamples were centrifuged in the SW41 rotor of the Spinco
VOL.16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
368 BROWN ET AL.
L2-65B ultracentrifuge at30,000 rpm for 125min at 4 C.
Disintegration of adenovirus particles byheat. CsCl-purified preparations of Ad2weredialyzed into PBS or 0.02 M Tris-hydrochloride, pH 8.1, and were heated at 56 C for 5 min (39). These virions were examined in the electron microscope directly, or after rebanding in CsCl density gradients. In some cases, the polypeptide composition of the rebanded virus preparations was also analyzed by electrophoresis in sodium dodecyl sulfate (SDS)-polyacrylamide gels.
Enzymes and reaction conditions.DNase, electro-phoretically purified, wasbought from the
Worthing-tonBiochemicalCorp., andwasusedwithout further pretreatment. Adenovirus cores inavolume of0.1ml wereincubatedin 0.01MTris-hydrochloride,pH 7.5, and 0.005 M MgCl2 with 1 ug ofDNase per ml at
ambient temperature for 5, 10,or20min.Insomeof the experiments, DNase digestion was performed in
0.01MTris-hydrochloride, pH7.2,0.01MCaCl2, and
0.01 MMnCl2 (35). After DNase treatment, someof the samples were incubated with500ug of pronase per mland 1%SDS at 37 C for30minandsubsequently were extracted with 0.5 ml of redistilled phenol saturated with TE. The phenol and aqueousphases
were separated by centrifugation, and the aqueous phase was dialyzed against one-tenth strength elec-trophoresis buffer and analyzedby electrophoresison
polyacrylamide (1.5%)-agarose (0.8%) gels as de-scribed below.
Pronase B was obtained from CalBiochem, dis-solved in 0.01 M Tris-hydrochloride, pH 7.5. at a
concentration of5 mg/ml, and preincubated at37C for2h.Each lot of Pronase B purchased wasassayed afterautodigestion for the absence of endo-or
exonu-clease activity by incubating 3H-labeled Ad2 DNA with 250 Mg of preincubated pronase in 0.01 M
Tris-hydrochloride, pH 7.2, 0.1 MNaCl, and 0.002 M
MgCl2for 60 min at 37C. Subsequently, the size of
the DNA was determinedonneutral sucrosedensity gradients (6).
Restriction endonuclease RI from Escherichia coli (EcoRI). The purification of the restriction endonuclease EcoRI has been described previously (11;R. N. Yoshimori, Ph. D. thesis, Univ. of Califor-nia, San Francisco, 1971). For the experiments de-scribedinthis report, the DEAE-cellulose fraction of the endonucleasewasdialyzed against0.01M
potas-sium phosphate, pH 7.0, 0.35 M NaCl, 0.001 M EDTA, and 0.007 M
$l-mercaptoethanol
and was chromatographed on a phosphocellulose (Whatman, P-11) column (1 by 10 cm) previously equilibrated with 0.01 M potassium phosphate, pH 7.0, 0.35 M NaCl, 0.001 M EDTA, 0.007 M f,-mercaptoethanol, and 10% glycerol. Thephosphocellulose column was developedwithagradientofNaCl from 0.35 to 0.70 Min the same buffer. The EcoRI endonuclease was elutedatapproximately0.5M NaCl. Theenzyme was storedinthe same buffer it was eluted in, and proved stable at 0 to 4 C for several months. The reaction conditionsforthisenzyme as employed in this labora-tory werereported elsewhere (11).
Sarcosylcoresof Ad2tobedigested with the EcoRI
restriction endonuclease weredialyzed into TE and treated, in atotal volume of 100to 150 ul of 0.05 M Tris-hydrochloride (pH 7.5) and 0.01 M MgCl2, with
10 to 30 Ml of the phosphocellulose fraction of the enzymeat37Cfor 60 min.Subsequently, 5Ml ofa0.2 MEDTAsolution, and insomeofthe experiments 10
Mul
each ofa Pronase solution (5mg/ml) and a 10%SDS solution, was added and incubation was contin-uedfor 30 minat37C. In other experiments,Pronase addition was omitted (for details see legend to Fig. 13).The mixture was then extracted with phenol (TE saturated), the aqueous and phenol phases were
separated by centrifugation, and the aqueous phase wasdialyzed against one-tenth strength electrophore-sis buffer and analyzedby electrophoresis on polya-crylamide-agarose gels asdescribed below.
Gelelectrophoresis. (i) Analysis of polypeptides in SDS-polyacrylamide gels. The procedures of Maizel (25) were used with minor modifications. Samples of sarcosyl or pyridine cores or intact adeno-virions were dialyzed against 0.01 M ammonium-acetate, lyophilized in a Virtis lyophilizer,
resus-pended in 0.1 ml of sample buffer (0.0625 M Tris-hydrochloride, pH 6.8, 2% SDS, 1% mercaptoethanol, and 10% glycerol), and heated to 100 C for 2 min. Subsequently,5Ml of a 0.25%solution of bromophenol blue was added, and the samples were analyzed on 13%polyacrylamide gels. Cylindrical gels of 14 cm in length were used. The composition of the gels and of theelectrode buffer was described previously (13, 37). Electrophoresis was performed for 5.5 to 7 h at ambient temperature and 100 V. The gels were then fixed in 20% trichloroacetic acid, stained in 0.25% Coomassie blue in 20% methanol and 7% acetic acid for periods ranging from 30 min to 16 h and were destained in 20% methanol and 7% acetic acid for several days. In some of the experiments in which
radioactively labeled proteins were analyzed, the gels
were cut with a slicer and each slice (usually 0.8 to 1.0 mm thick) was processed as described below. For someexperiments slab gels were used of identical composition (21) andelectrophoresis was performed in
anapparatusasdescribedby Studier (41).
(ii) Analysis of DNA fragments in 1.5%
poly-acrylamide-0.8% agarose gels. The composition of
the gels and of the TEB electrophoresis buffer was described earlier (13). DNA samples (0.1 to 0.2 ml) in one-tenthstrength ofelectrophoresis buffer were lay-ered on cylindrical gels, and electrophoresis was
carriedout at40Vatambienttemperature for 8.5 to
12 h. In most experiments the EcoRI fragments of 14C-labeledAd2DNAwere added as molecular weight markers. Immediately after electrophoresis, gels were
cut into 0.8- to 1.0-mm slices and the slices were processed for counting. Individual gel slices were placed intoplastic tubes or directly into scintillation vials and extracted with 0.2 to 0.5 ml of 0.1% SDS with shaking at 37 C for 18 h. The extracts were adsorbed to GF/83 glass-fiber filters, dried at 80 C,
andcounted in5 ml ofthetoluene-based scintillator
inaPackardTriCarb liquid scintillation spectrome-ter, model 3385 or 3330. In someexperiments portions J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
CORES FROM ADENOVIRUSTYPE 2 369 ofthe extractswere counted directly in 5 ml ofthe
toluene/methanol-based scintillator.
Electron microscopy. Monolayers of KB cells
infectedwithAd2weretransferredtoanicebath and
fixed with ice-cold 2% glutaraldehyde in PBS. After fixationonicefor 60min, the cellswerescraped from
theplates, washed with cold PBS, and storedat4C
until prepared forelectronmicroscopy.
Infected cells were prepared for thin sectioning after glutaraldehydefixation as previouslydescribed (3). Cellswerepostfixed for1hwitha1%solution of osmiumtetroxide in Millonig phosphate buffer (27).
The post-fixed cellswereembeddedinepon812by the
procedure of Luft (24). Silverto silver-gold sections werecuton aReichartOM U-2 ultra microtome.
Freeze etching of glutaraldehyde-fixed virus-infected cells was carried out by the procedure
de-scribed by Brown et al. (4). Some specimens were fractured and etchedafterwashingindistilledwater priortofreezing.
Negative stainingwascarriedoutby the procedure ofAnderson (2). Phosphotungstic acid was prepared
asa2%solutionindistilledwaterandwasadjustedto pH7.2by additionof NaOH.
Preparation of purified viral coresforthe electron microscope wascarriedout byamodification ofthe aqueousspreading procedure of Davisetal. (9).The spread monolayers of DNA and protein were trans-ferred to carbon-coated grids and were rotary shad-owedatan angle of 100 with platinum carbon.
Specimens were photographed in a Siemens 101
electron microscope utilizing either bright or dark fieldoptics. The magnificationofthe instrumentwas calibrated withacarbongratingreplica having2,160 linespermm (Ernest Fullam Co.,no. 1002).
RESULTS
Morphology of Ad2. It is generally accepted
that theadenovirions havea mean diameter of
about 80 nm (20, 43) and that the adenovirus
capsid is composed of hexons which are 7 to 8
nmindiameter andpentonswhichare8.0nmin
diameter. Morerecentstudieshave revealedthe
hexon to be a rod-like structure composed of
three proteins withanoverall diameter of9nm
and a lengthof 11 nm and the penton base to
representasphere of 8-nm diameter (30, 45; G.
Wadell, Ph.D. thesis, Karolinska Institutet,
Stockholm, 1970). We reinvestigated the
di-mensions of the type 2 adenovirion for the
purpose of relating the observations on the
internalmorphology of theparticletothe
struc-ture oftheoutersurface. We measured the size
of adenovirus hexons and pentons in both the
intact virion (Fig. la) and as free structures
after release from the virion by heating (Fig.
lb). We also determined the center-to-center
spacing ofthe hexons onthe face of the intact
virionandingroupsof hexons releasedfromthe
triangular facesofthe virus icosahedronduring
heating (Fig. lb). The dimensions of the various
structures are compiled in Table 1. The
pre-dicted size of the virion obtained by measuring
the center-to-center spacing of the hexons was
86.16 nm using the equations E = dT½ and D
= E/0.618 (40), where E is the edge of one
equi-lateral triangular face, D the diameter of the icosahedron, d the center-to-center spacing of
the hexons, and Tthe triangular number of the
icosahedron (for adenovirus T = 25). The
mea-sured value for the particle was 97.5 nm. This
suggeststhat in negatively stained preparations
the virions are distorted by compression as
described by Anderson (1) and by Moody (28), such that the observed cross-sectional diameter isincreasedby 13%. Byutilizing theequation V
= 2.1817 E3, where V is the volume of an
icosahedron and E the edge of one of the
equilateral triangular faces, we foundthe total volume of the virion to be 3.29 x 105 nm3.
Morphology
of thecoreof Ad2.Subtracting
the thicknessofthe hexon-penton shellfromthe complete virion left a subcapsomere space ofvolume1.35 x 105 nm3andaninternal diameter
of 64.15 nm. The measured diameter of cores
seen in thin-sectioned intact virions was 63.9
nm. The smaller size probably results from
shrinkage during the embedding process (29).
The volumeofthe space attributed to the core was therefore 41% of the total volume of the
virion.
The 63.9-nm electron dense core seen in
ultra-thin sections ofthe intact virion (Fig. 2) revealednoinformationregarding the
organiza-tionofthecomponents of the core. Weinitially
attemptedto study the interiorofthe adenovi-rionby freeze-cleaving ofpurifiedvirionsandof
virions attached to the surface of cells. Such
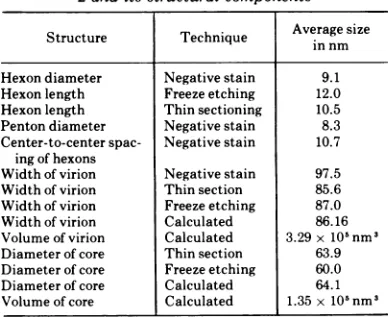
TABLE 1. Measured and calculated dimensionsof Ad
2and its structural components
Averagesize
Structure Technique in nm in nm Hexondiameter Negative stain 9.1
Hexonlength Freezeetching 12.0 Hexonlength Thinsectioning 10.5
Penton diameter Negativestain 8.3
Center-to-centerspac- Negativestain 10.7
ingofhexons
Widthof virion Negativestain 97.5
Widthof virion Thin section 85.6
Widthof virion Freezeetching 87.0
Widthof virion Calculated 86.16 Volume of virion Calculated 3.29x10'nm'
Diameter of core Thin section 63.9 Diameter of core Freezeetching 60.0
Diameter of core Calculated 64.1
Volume of core Calculated 1.35x10'nm9 VOL.16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.257.451.492.651.2]4 V4
FIG. 1. Negatively stained preparations of(a) intact type 2 adenovirions and (b) virus capsomeres released from the virion by heat treatment. The intact virions (a) show the classicalarrangement of six- and fivefold organized capsomeres. It is possible on a number of the virions to measure accurately the center-to-center spacing of the capsomeres and the diameters of the subunits. The capsomere in the disrupted viral capsid are also organized in sixfold arrays (b). The hexons have the same cross-sectional diameter as those in the intact virion and in addition have a recognizable substructure. Pentons with fibers can be seen in the background
(arrow). Magnification, x226,400.
370
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.58.453.49.606.2]* .. ' S .
4n '
*rr i
* 9. * .'.! ; '
**-''
.s-0'' *-.".
,*
S.,-_
* B-w@- S.;r'>
.* . t!. * *
*0
-. '. ,
* -S.
S.. * x*x,
'
*S .
-S....^:;
*:
,,-.tJ
k **;2'; a
**
*
V
|ebla. * / q . ~.J
r:
've:5<~~'t'v
;i
FIG. 2. Ultrathin sections ofintranuclearadenovirusparacrystallinestructures. (a) Lowmagnification of
cellat 36hpostinfection.Smallcrystallinestructuresare visible in the nucleus. (b) High magnification ofa
paracrystallineintranuclear inclusion. The cross-sectioned virions haveadistinctelectron densecenterandan
electrontran-sparent layer ofcapsomeres.Magnification: (a) x10,600; (b) x80,000. 371
'U
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.60.455.54.611.2]372 BROWNETAL.
experiments never produced fracture planes
whichpassedthrough the interior of thevirions,
but rather always exposed the outer surface of the particles (see ref. 3; Fig. 3). We interpret
this resultto indicate differences in the
hydra-tion oftheinterior ofthe isolatedvirion relative
to the surrounding medium creating a resist-ance to thedeveloping fractureplane. Altering
the concentrations of cryoprotective
(glycerol)
didnotproduce anychangeinthe imageofthe
virion afterfreeze-cleaving. In a further attempt
toproduce fracture planes within the virionwe
have freeze-cleaved intact KB cells late after
infectionwithAd 2. At late times after infection
(35 to 40 hpostinfection), the nucleiofinfected
cells containlocalized areasof veryhigh
concen-FIG. 3. A paracrystalline inclusion of the type shown in Fig. 2 after freeze-etching in 30% glycerol and
shadowing withplatinum carbon. Some of the virions are cross-fractured producing arelativelyflat structure in
which theouterring ofcapsomeres ispoorlyresolved(A).Othervirions have a more distinctly resolved outer ring
ofcapsomeres(doublearrow B). The internal corestructure (C) is resolved in some virions as either a smooth
sphericalstructure (C left) orcontaining a knob-like substructure (C right). Occasionally, virions have been
cleaved such that the inner content is removed showing the inner smooth surface of the capsid (D).
Magnification: x162,000.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.90.426.165.593.2]CORES FROM ADENOVIRUS TYPE 2
373
rsa~~~~~V
FIG. 4. Anareaof a largeintranuclearparacrystalline inclusion afterfreeze-etching in distilledwaterand shadowing with platinum carbon. The hexagonal arrangement of the virions ispreserved inthefreeze-etched
paracrystals (large doublewhite-black arrows). Many of the virionsare brokenshowinginternal substructure
(singlewhite-edged arrow). Magnification: x140,000.
trations of virions
(paracrystals) (Fig.
2a,b).
within the nucleus could be cross-fractured.It was hoped that the virions in the cell
Cleavage
planespassed through
theouterlayer
nucleus would be similar in their cleaving ofcapsomeres and
exposed
inavaried mannercharacteristics tothe surroundingnucleoplasm the internal structure of the virions. In some
allowing fracture planes to pass through the instancesthe virions were cut sothatasmooth
outer layer of capsomeres and exposing the surface was
produced
whichbarely
revealed theinterior of the virion. The
paracrystals
were outerringof capsomeres(Fig. 3).
Suchparticles
readily revealed by freeze-cleaving (Fig. 3 and were fractured close to the center and had a
4), andin contrast toextracellularvirus, virions cross-sectional diameter
slightly
larger
thanVOL.16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.73.416.82.512.2]374 BROWN ETAL.
that found in thin-sectioned virions (Fig. 2b).
The difference in the size of the freeze-etched
virions compared to thin-sectioned structures
can probably be accounted for by shrinkage
occurring during dehydration and embedding
for thin sections. The diameters of the
freeze-etched virions of 87 nm agrees with the size of
the virion predicted in Table 1.
In other virus particles the fracture plane
passed through the outer layer of capsomeres
and then over the internal contents revealing
the inside of the virion as a smoothsphereoras
composed of closely packed spheres with a
diameter of 9to 10 nm(Figs. 3and 4). In virions
fractured in this fashion the thickness of the
outer hexon layer was 10.5 to 12 nm. Some
virions were broken such that the core was
removed and the inner surface of the capsids
was exposed. This inner surfacewas smooth in
appearance(Fig. 3).
Isolation of sarcosyl cores from purified
virions. When CsCl-purified preparations of
[I4C
]formate-labeledAd2 weretreated withsar-cosyl and subsequently centrifuged to
equilib-rium in CsCl density gradients as described
above, the bulkofthe "4C-label inAd2 banded in a density position
approximately
0.2 g x cm-3lighter than thatofthe3H-labeled marker DNA which was purified from Ad2 virionsby
the SDS-Pronase procedure (13; Fig. 5a). The"4C-label
ontopofthegradientwasduetoviralprotein. The apparent buoyant density of Ad2
DNA in this experiment was higher than the
value of p = 1.716 g x cm-3 reported
previ-ously(10), perhapsduetothe presence ofsmall
amounts of sarcosyl. For the analysis of the
sarcosyl cores (seebelow)fractions27to29 were
used. It will be shown by electron microscopy
(see Fig. 8) that these fractions contain viral
cores.
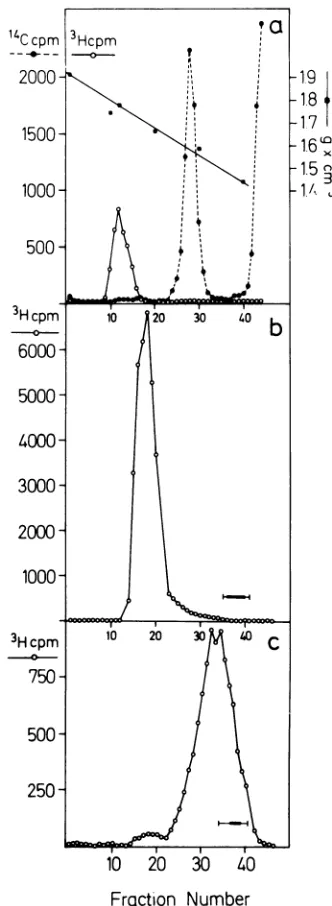
The data presented in Fig. 5b and c reveal
that sarcosyl cores can be readily separated
fromthebulkofthe soluble viral proteinsby gel
filtration on Sepharose 2B. When [3H
Ithymi-dine-labeled Ad2 wastreated with sarcosyl, the
cores eluted in a single sharp peak (Fig. 5b),
whereas Ad2 labeled with a mixture of
3H-labeled aminoacids yieldedtwopeaksof
[image:9.503.281.447.73.526.2]radio-activity upon treatment with sarcosyl and gel
FIG. 5. Isolation of sarcosylcores. (a) Equilibrium sedimentation in CsCl density gradient. Approxi-mately1.8optical densityunits at260nmof [14C]for-mate-labeled Ad2 in buoyant CsCl solution were
treated withsarcosyl andcentrifuged to equilibrium in a CsCl density gradient together with 3H-labeled Ad2DNA (0.3ug)asdescribed. The refractive indexes
2000 1,9
1.7
1500
16.1.5
1000- '
500
-3Hcpm 10 20 30 40b
-0-b
6000-
5000-
4000-
3000-
2000-1000,
3Hcpm 10 20 30
00
C
-0-
750-
500-
250-Frcaction
Number
were measured ineverytenthfractionandthe densi-tieswere calculated asdescribedelsewhere (44). The
3Hand 'IC activities were determined in 5- to J0-Mul
portions ofeachfraction as described. (b) Gel
filtra-tion on Sepharose 2B column. Approximately 2.3
optical density units at 260 nm of
[3H]thymidine-labeled Ad2 in buoyant CsCl solution were treated withsarcosylin TE andpassed overaSepharose2B columnasdescribed. The bar indicates theposition of
the dinitrophenyl-alanine dye which was used as a
marker.(c)Inasimilarexperiment2.0optical density units at 260 nm ofAd2 labeled with a mixture of
3H-labeled amino acids were used under conditions
identical,tothose described in (b).
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
CORES FROM ADENOVIRUS TYPE2 375
filtration (Fig. 5c). The minor peak represents
the sarcosyl cores, the major peak the viral
proteins (Fig. 5c). In this experiment, about
3.9% of the 3H-amino acid label was recovered
with the sarcosyl cores. This finding indicates
that only part of polypeptide VII is present in the sarcosyl core, since polypeptide VII
represents 14% of the viral protein (32). A part
ofpolypeptide VII wasapparently releasedand
could be found in the main peak (Fig. 5c). At
thepresent
time,
the mechanismofthisreleaseis notunderstood.
Sarcosylcores prepared by gelfiltration were
indistinguishable in the electron microscope
from sarcosyl cores obtained after equilibrium
centrifugation.
Athird way of preparing Ad2 cores wasby the
pyridine method (33) which yielded structures
different from sarcosylcores asjudged by
elec-tron microscopy and the polypeptide
composi-tion (see below).
Electron microscopy ofpurifiedcores. Sub-viral particles
produced
by heattreatment (39),pyridine treatment (33), or by treatment with
sarcosyl were examined in the electron
micro-scope. It was necessary to utilize a variety of
preparative procedures for the analysis of each
type ofsubviral particle, as aprocedure which
produced goodresultswith one type ofsubviral
a
structure frequently produced no information when applied to another.
Electron micrographs of structures produced
by heating adenovirions to 56C for 5 min are
shown in Fig. 6. Best preparative results were
obtained by negative staining of these
struc-tures with neutral phosphotungstic acid. The
subviral structures observed were similar to
those described by Russell et al. (39). Free
hexons, groups of hexons, pentons, and phous structures were seen (Fig. 6a). The
amor-phous structures were larger than the intact
virions from which they were derived and had
anaverage diameterof 167nm, suggesting that
they represent particles which have lost their
structural integrity and have relaxed into a
larger volume. The subviral particles produced by this procedurewerefoundtocontain most of
the viral structural proteins (see below). Some
substructure was recognized in the amorphous spheres (Fig. 6b). Sphericalstructures about20
nm indiameter with acenter-to-centerspacing
ofabout30nm canbedistinguished.Itwasnot
possible to determine if these substructures
reflect aninternalorganization ofthevirion or a
topological alteration oftheviral surface.
Particles produced by treatment with
pyri-dine produced similar results whether stained
positively with uranyl acetate or negatively
b
FIG. 6. Negativelystainedpreparation ofAd2after heatingat56 Cfor5min.(a)Lowmagnificationshowing
viralcapsidcomponentsandlarge amorphouscores(arrows). (b)Higher magnificationofasubviralstructure showingasubunit-likeorganization.Inthebackground can beseenfree capsomeres. Negativelystained with
phosphotungsticacid.Magnification: (a) x80,OOO; (b) x,200,000.
VOL.16,1975
.1 ..
'... :1IC .i
,%.r
. A''.1-.1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.503.50.446.385.622.2]376 BROWN ET AL.
stained with
phosphotungstic
acid(Fig. 7).Thepyridine core structure was best preserved by dilutingthe preparationin 0.37%formaldehyde
inPBS aftergradientpurification.The pyridine
cores were tightly packed and had a mean
diameter of 66 nm (Fig. 7a). Occasionally, partially decomposed structures were observed
which seemed to contain avariable number of
smallerspheres approximately22 nm in
diame-ter (Fig. 7b). The internal spherical structures
seen in the pyridine cores had a haloof
associ-ated material which appeared to beofuniform
thickness. It is possible that the
sphere-associated material holds the 22-nm
substruc-tures together in the tightly packed structures
(Fig. 7a) and coats the entire core preventing
thevisualizationofsubstructureinthemajority
ofparticles observed.
Sarcosyl cores produced different images in
the electron microscope depending upon the
preparative procedure employed. The sarcosyl
cores were rather fragile when examined by
negative or positive staining, but could be preserved by treatment with formaldehyde as
described above. Positively stained sarcosyl
cores (Fig. 8b) revealed electron dense
struc-tures with asomewhat variable diameter which
measured 66 to 67 nm in the best preserved
structures. The structures tended toaggregate,
aN "I..L
a~~~~~~~~~~~~~~~~4
and
frequently
fibrousmaterial, possibly
DNA,
radiated from the
centrally
located electron dense structures. The size variation described aboveappeared
toresultfrom thereduction insize of the
centrally
located densebody
asmoreDNA wasreleased fromthe
particle.
Whentheparticles
werenegatively
stained(Fig. 8a),
itwas no
longer
possible
toidentify
the DNAfibersortofindsuch welldefined electron dense
centers as seen in
positively
stainedprepara-tions. The
negatively
stained structures,rather,
appeared
as a collectionofsmallersphereswith a constant diameter of 21.6 nm. Discreteclus-ters of
eight
to ten 21.6-nm diameterspheres
wereobserved
(Fig.
8c).
A
comparison
ofthemorphology
ofpositively
and
negatively
stainedsarcosyl
particlessug-gests that the structures of 67 nm diameter
described above are
composed
of a number ofspheres
with a diameter of 21.6 nm. Thesespheres appeared
very electron denseand were notdistinguished
in positivelystainedprepara-tions. In
negatively
stained preparations thedifferential penetration of staininto the 67-nm
particle
and the partial exclusion of stainfrom theinteriorofthe 21.6-nmsubstructuresfacili-tatesvisualizationoftheinternalmorphologyof thesarcosyl
particles.
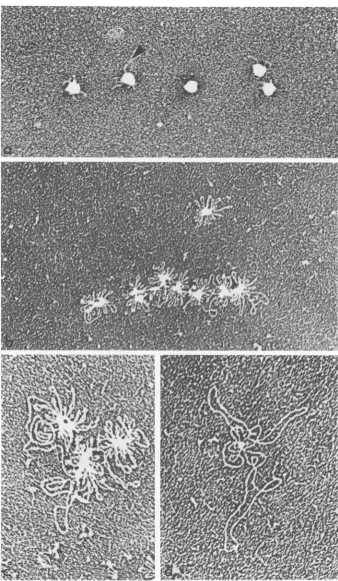
We attempted to verify that the fibrous
I. '-. 9 4 '
b
FIG. 7. Viralcores releasedfrom adenovirions bytreatment withpyridine. (a) Typical pyridine cores. The structures arehydrophobicandadhere to one another. The surface is smooth and bulges as though there were
spherical structures under the surface. (b) An unusual pyridine core. The particle appears to have opened somewhatrevealing thesphericalstructures within. The spheres are coated with some material which may have
been on the outer surface of the core (arrow). Negatively stained with uranyl acetate. Magnification: (a)
x370,000; (b) x300,OOO.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.503.65.460.384.605.2]CORES FROM ADENOVIRUS TYPE 2 377
'.
_.re*
tE'';;-
_' ;-;
.8w;417
s ' * g _ wW
'' *'.'s,. i;'''
,&, :
., o.'.
#.'.''.'..-;o
' s.FIG. 8. Cores released from Ad2 after treatment with sarcosyl. (a) Cores negatively stained with uranyl
acetate.Onecannotdetect the freeDNAor awell-defined central dense body. Rather the cores are made up ofa
numberof smaller spherical structures. Unlike the pyridine cores these sphereshave nohalo of material around them. Thecoresubunitssometimes appeartobe heldtogetherby fibrousmaterial(arrow). (b)Asarcosyl core
positively stained with uranyl acetate. The arrowpoints toDNAfibers protrudingfrom the electron dense
core. (c) High magnification ofa negatively stained sarcosyl core, showing 9orpossibly 10 subunits.
Mag-nification:(a) x116,000;(b) x156,000;(c)x420,000.
VOL16,1975
I-V
I1.I
..I
.S. 1..
'k.,
..'
-:
I.."A i
I :, 4'.. 1-Y
.4- ,*
1.
on November 10, 2019 by guest
http://jvi.asm.org/
378 BROWN ET AL.
structures seen radiating from the positively stained sarcosyl cores was DNA
by
spreading sarcosyl cores on a proteinmonolayer.
To oursurprise, freshly prepared particles did not
spread into the random DNA structure as
expected, butratherappearedas
tightly
packed particlesofapproximately thesizedescribedforthepositivelystained particles (Fig. 9a). Occa-sionally, these particles had small protrusions
ontheirsurface suggestingthe association with these structures of free DNA (Fig. 9a). If the particleswereallowedtostandfor afewdaysat 4 C, the amount of free DNA
protruding
in-creased, whereas the size of the central core
decreasedconcomitantly (Fig. 9b). Free endsof DNAwererarelyseen. Asthe DNAwasreleased
fromthe densecenter oftheparticle,itformeda
rosetteofclosed loops around the central
struc-ture. The
loops
of DNA werefrequently
highlytwisted
along
theirlength,
butopened
attheir distal ends like hair pins(Fig.
9b, c).
With increasing time ofincubation at4C,
the coresseemed torelease
progressively
moreDNAintolong random structures, some of which
rep-resented one genome
equivalent
of DNA (10to 11 ,um in
length).
Even in these highlydisorganized arrays free ends of DNAwereonly rarelyseen, and theDNA wassimply organized
inlarger loops which radiatedasbeforefromthe
surface of asmaller core structure (Fig. 9d).
Ifthe preparationswere allowedtostandfor a
period ofabout 1 week, thecore-like structures
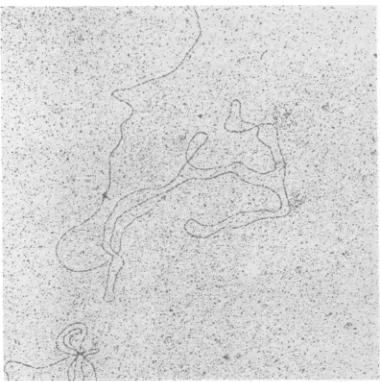
were only rarely seen. Instead randomly
ar-ranged circles of oneadenovirusgenomelength similar to those described
by
Robinson et al.(36) wereobserved (Fig. 10).Open moleculesof
one andtwovirallengthscould also be foundin
these preparations. Treatment ofthe purified sarcosyl cores with pronase at any stage after
preparation converted the dense bodies into
linear moleculesofone genome length. Polypeptide composition of sarcosyl cores
from Ad2. Since the
buoyant
densityofsarcosylcores was found to be
considerably
lower thanthat of Ad2 DNA (Fig. 5a), it was likely that
sarcosyl cores consisted of DNA andprotein. It
is not known whether sarcosyl can bind to the
cores and alter their
buoyant
density.There-fore, reliableestimates oftheamount ofprotein remaining associatedwithsarcosylcores cannot
be made from the buoyant density. The
poly-peptidecompositionofsarcosylcores was
deter-mined by electrophoresis on
SDS-polyacryl-amide gels using both 3H-amino acid-labeled
(Fig. 11a-c) and unlabeled cores (Fig. 12, slot
no.2). Acomparisonofthepolypeptidepatterns
presented in Fig. llb and c demonstrates that
digestion
of thesarcosyl particles
with DNaseprior
toelectrophoresis
does not affect theap-parent
polypeptide
composition
ofthesarcosyl
cores. In the
experiment
shown inFig.
12(slot
no.
2)
thesarcosyl
cores wereanalyzed
afterdigestion
with DNase which alsoregistered
inthe
gel
pattern. Theidentity
ofpolypeptide
VII was demonstrated
by
comparing
slot no. 2with slotno. 3which containedDNase
by
itself(Fig. 12).
The data indicate that it ispredom-inantly
thealanine/arginine-rich
polypeptide
VIIwhichwasassociatedwith the
sarcosyl
cores.The
identity
of thispolypeptide
was further documentedby
co-electrophoresis with ["C1-formate-labeled
polypeptides
fromadenovirions(Fig.
lla),
andby
thehigh
proportion ofarginine
and alanine in the amino acidcom-position
ofsarcosyl
cores. The sarcosyl corescontained also a small amount of the hexon
polypeptide (II).
From the data presented inFig.
llaandb,
itcanbe estimated that <3%ofthe total hexon
polypeptides
were stillassoci-ated with sarcosyl cores. Small amounts of
polypeptides
VIII and IX were also present in thesarcosyl
cores.Thisfindingisnotsurprising, since polypeptides VIII and IX are associatedwith the hexons (16, 17,
18).
Asa
control, pyridine
coreswerepreparedby
the method of
Prage
et al. (33) and wereanalyzed by electrophoresis
onSDS-polyacryl-amide gels (Fig. 12, slot no. 5). It has been
shown previously (17, 18, 32, 33) that
pyridine
cores contain
exclusively
polypeptides V andVII,
and thisfinding
is also documentedby
the results shown in Fig. 12, slot no. 5. This
control supports the conclusion that
sarcosyl
cores arefreeofpolypeptide V, since this
poly-peptide
canreadily
be detected inelectro-phoretograms
of corepreparations.Whenpurified virionswereheatedto56Cfor 5 min
(39),
repurified by equilibriumcentrifu-gation in CsCl density gradients, and then analyzed by electron microscopy and electro-phoresis on
SDS-polyacrylamide
gels, there-sulting
cores had a typical structure (Fig.6b)
and contained most of the viral polypeptides
(not shown).
The organization oftheviral DNA in
sar-cosyl cores. The sarcosyl cores contain intact
viralDNA. When [3H]thymidine-labeled
sarco-syl
cores were extracted by the SDS-pronase-phenol method (10, 13), Ad2 DNAwasobtainedwhich co-sedimented at 31S with "4C-labeled,
intact Ad2 marker DNA in neutral sucrose
density gradients. After incubation of
[3H ]thymidine-labeled sarcosyl cores with
pan-creatic DNase for 5 to 20 min as described
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
-
ia~~~~~W
A; 'I i. ~ ~ ~ ~ ~ ~ ~ ~ ~ ~
.--V 4b~~~~~~~lw~'r
FIG. 9. Electronmicrographs of sarcosylcoresprepared by spreadingon aprotein monolayeratvarioustimes after isolation. (a) First day of purification: thecores are tightlypackedstructuresfrom whichoccasionallya small amountof DNA protrudes. (b) Second day after isolation:moreDNAprotrudes inaradialfashion from the cores. The loops of DNAare closedstructures. Free endsare not seen. (c) Thirdday after isolation: the centralcorebecomes smaller and the freeDNAloops larger. Theloops remain closed. (d) Fourtofive days after isolation: thecentraldensebody has almost disappearedandthefreeDNAloops represent nearly the complete genomelength. Magnification: (a) x40,OOO; (b) x38,000; (c) x60,OOO; (d) x60,OOO.
379
on November 10, 2019 by guest
http://jvi.asm.org/
[image:14.503.78.416.36.617.2]380 BROWN ET AL.
FIG. 10. DNA of i mately 1 weekafter is viral genomeunitlend
Fig. 9 is no longer appi
illustrated in Fig. 13a this protein was removed by pronase digestion.
This interpretation was confirmed bv the
datapresented in Fig. 13c. Upon extraction and
Pronasedigestion of the 3H-labeled material on
top ofthe gel showninFig. 13b, the 3H-labeled
- --^ EcoRI A and C fragments were released from
this complex and could be identified by
co-electrophoresis with the appropriate "4C-labeled
-,SsFlar marker fragments (Fig. 13c). Fragments A
and C are located at either terminus of the
Ad2 DNA molecule (31) (insert to Fig. 13a) and
are presumably involved in the circularization
oftheDNAmoleculeinsidethevirion(36).The
major polypeptide found associatedwith
sarco-syl cores waspolypeptide VII. It is not yet clear
whethersomeofthe polypeptide VII chains or, perhaps more likely, another as yet unidentified polypeptide present in minute quantities is
4d2. Sarcosyl core at approxi- responsible for complex formation with
frag-olation. Thegenomeis acircleof ments A and C. This complex is interesting
gth. The central core as seen in because it might stabilize the viral DNA in a
rarent. Magnification: x32,710. circular configuration.
above, all the 3H-label was converted to low-molecular-weight material which migrated with
the bromophenol blue dye front upon electro-phoresis in polyacrylamide-agarose gels.
Whenthe sarcosyl cores were incubated with
EcoRI restriction endonuclease and when the
incubation mixture was subsequently treated with Pronase, the [3H ]thymidine-labeled Ad2
DNA inthe sarcosyl cores was cleaved into the
six specific fragments (31) which
co-electro-phoresed withthe six specific EcoRI fragments from
"4C-labeled
marker Ad2 DNA (Fig. 13a).These results were obtained independently of
thetime afterisolation ofthe cores and indicate
that, although theviral DNA was closely
associ-ated withpolypeptideVII in the core, the DNA
remained fully susceptible to digestion with
pancreaticDNase and that the six palindromic
sites recognizedbythe restriction endonuclease
EcoRI were accessibleto and canbe cleaved by the enzyme.
Strikingly different results were obtained
when sarcosyl cores were treated with theEcoRI
restriction enzyme and were subsequently not
digested with pronase (Fig. 13b). In this case the amount of the 3H-labeled EcoRI A fragment
was reduced in amount relative to the
l4C-labeled marker DNA, and the 3H-labeledEcoRI
C fragment was missing. Some of the 3H-label
was retained on top of the gel (Fig. 13b); this
could be due to the missing DNA fragments
which are complexed with protein and thus
cannot enterthe gel, whereas in the experiment
DISCUSSION
(i)Morphology ofthe adenoviruscore.The
electron microscopical observations presented
here suggest that the interior of the
adeno-virion is structurally highly organized and not
asimple amorphous electron dense mass.Upon
treating the intact adenovirion with sarcosyl,
spherical structures 66 to 67 nm in diameter
arise. This value is slightly larger than the
size of the core in the intact virion as
meas-ured in electron micrographs of thin sections of freeze-etch preparations. The slightly larger
size of the free core mayresult from distortion
due to compression of the particles which
generally occurs in air-dried preparations.
Al-ternatively, the increase in the size of the
free core may result from an expansion of
the structure once it is freed from the
con-fines of thecapsidshell.
The DNA ofthe 67-nm core ispacked into a
number of smaller spherical structures each
havinga diameterof 21.6 nm. There are8 to 10
such spheresdetectable foreach core, although
this number isprobablytoolow as thosespheres
which arecoveredby others cannot be counted.
Given the cross-sectional diameter ofthe
sub-structures it is possible to calculate their
vol-ume, assuming they are spherical, by the
for-mula V = 4.189 r3 (40), where Vis the volume
and r the radius of the sphere. The volume of
thisstructurewould be5.2 x 103nm3,and 25of
these spheres would be required to occupy the
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:15.503.65.255.75.266.2]CORES FROM ADENOVIRUSTYPE 2 381
It.sSIDsX,
j}HcpVIm300
X:O,
se '*~
CV U2000 02 04 0608 1OR,
01 In IVv VI 1 O , \s0V
.~~~~~~~~~~~~~J
'
_
10 0 30 0 50 60 20 80 90 10 110
b
V0 VUIx
0 0 30 4 5 60
s ~~~~~~~~~d
nmIv v VI vn v'll Ix
L
X=7080o YU 1010l 10 20 30 40 Fraction Number
050 60 70 80 90 100 110
FIG. 11. Analysis of polypeptide composition of sarcosyl cores. (a) Sarcosyl cores were prepared from 3H-labeled amino acid Ad2 by gel filtrationon Sepharose 2B, dialyzed against 0.01 Mammonium-acetate, lyophilized, and resuspended in 0.1 ml of sample buffer. A small amount of "4C-labeled Ad2 marker virus wasadded and the samplewaselectrophoresedona13%polyacrylamide gel(21)for7.5hat100V,asdescribed.
The Romannumeralsdesignate the viral polypeptides. In the insert the logarithmsofthemolecular weightsof eachof the viralpolypeptides areplottedversusthe
Rf
values. (b and c) Sarcosylcores werepreparedfrom3H-labeled amino acid Ad2 by equilibrium centrifugationinCsCIdensity gradients andweredialyzed against 0.01MNH4-acetate buffer. One portion (b)wasincubated with50 ,ugofDNase under the conditionsdescribed andthereactionwasstoppedbytheadditionofEDTA.Anothersample(c)wastreatedidenticallyexceptthat DNase wasnotadded tothe reaction mixture.After lyophilization and resolubilization in samplebuffer, the samples wereanalyzed by electrophoresis on13%polyacrylamide gelsat100Vfor 7.3 h. (d) Asacontrol the polypeptides of "4C-labeledAd2wereanalyzed byelectrophoresison13%polyacrylamide gels.
entire internal volume (1.35 x 105 nm3) ofthe
virionwhichwehave allottedtothe-core (Table
1). By actualcountthe number of spheres in the
core is lower, as would be expected, since the
21.6-nm-diameter spherescontainonlythe viral
DNAandprotein VII.
It is possible to estimate more precisely the
number of spheres making up the core of the
virion by calculating the volume occupied by
the viralDNA and proteinVII. Ifoneassumes
the viral DNAtoapproximateacylinder2.0nm
wide by 109.4 nm long (12), its volumecan be
calculatedby the equation V = r2 irh(40) tobe
3.4 x 104 nm3 (where V is the volume, r the
radius, and h the height of the cylinder).Everitt
et al. (18) have shown that each virion contains
1,070copies of protein VIIwhichhasa
molecu-larweightof18,500. Assumingthatthisprotein
isglobular, similarto serum albumin, it would
have a volume of about 40nm3 (42). The total
volume of protein VII in the core would
there-fore be approximately 4.3 x 104 nm3, and the
totalvolume of theDNA and protein VII would
be about 7.7 x 104 nm3.Thus, 14ofthe 21.6-nm
diameter spheres would be required to contain
all of the DNA and protein VII.Thisnumber is
probably toohighasthe above calculation does
not take into accountthe fact thatsome ofthe
DNA must run between the 21.6-nm diameter
spheres connecting them together.
Another consideration in estimating the
num-ber ofDNA-proteinVII spheres which the core
contains is that of the symmetry relationship
between the icosahedral outer surface of the
virionand the innercore.Ifthecoreiscomposed
of subunits, as is implied here, and is to be
packed in an orderly manner into the T = 25
icosahedron, that is the adenovirus capsid, a
structuralcompatibilitymustexistbetween the
morphology of the core and thecapsid. This is
particularly desirable if the core isto be
pack-aged into the capsid by a mechanism of
self-assembly. The conditions ofcompatibility are
best met if the core, like the capsid, has
1500-
100-500
IHD 39HD l,-VOL.16,1975
lo
500
on November 10, 2019 by guest
http://jvi.asm.org/
[image:16.503.108.395.73.314.2]382 BROWNET AL.
1
23
dil
[image:17.503.97.230.75.412.2]4
5
FIG. 12. Analysis ofAd2andsubviral particlesby
electrophoresison13%polyacrylamide slab gels. Slab gel electrophoresis was performed at 40 Vfor 12 h under conditions as described. All samples were lyophilizedandredissolved insample buffer priorto electrophoresis. The individual slots contained the
following samples: (1) Ad2; (2) sarcosyl particles preparedandtreated withDNaseasdescribed inthe
legendtoFig. llb; (3) DNase; (4) Ad2; (5) pyridine corespreparedaccordingtothemethods described by
Prageetal. (33)andby Everittetal. (18).
icosahedralgeometry (20, 43). Percore, 8to 10
spheres of 21.6 nm diameter were observed in
electronmicrographs, and about14sphereswere
expected by the above calculations. We predict
that in reality each core contains 12 of these
21.6-nm-diameter substructures. This number
takesintoaccountthepossibility that becauseof
superposition ofthe 21.6-nm-diameter spheres
within thecoretheycannotall be distinguished
in electron micrographs, and thatnotall ofthe
viralDNAactuallycontributesitsvolumetothe
spherical structures. The number 12 would
imply that about 19%ofthe viral DNA isnotin
the 21.6-nm-diameter
spheres,
but may in fact link the 12 spheres to each other within thecore. Proposing that the core contains 12 of
the spheres has the added
advantage
that it allows an exactsymmetrical
relationship
be-tween the viral capsidP = 1, T = 25, and the
core P= 1, T= 1,using the notation of
Caspar
and Klug (8). The 12coresubunits would be so
oriented that the 5, 3, 2 axes of symmetry of
the inner core coincide with the same axes of
symmetryoftheouter
icosahedral
capsid.
That 12 coresubunits isnotanunreasonable
estimate for thenumber
of21.6-nm-diameter
spheres in the virus core is alsosuggested
by
examination of an icosahedroncontaining
12structural units. When such a structure is
viewedfrom one direction, aswould be thecase
inthe electron microscope, a maximum of nine
subunits can be
distinguished.
If theparticle
isslightly distorted, amaximum of 9 to 10
sub-unitswould still beseen,the remainingsubunits
being obscured.
Utilizing the above calculations the
DNA-protein VII complex would account for about
57% of the total volume allotted tothe core of
theintact virion
(Table
1).Everittetal. (16,18)
have suggested that the interior of the virus
directly below the outer
layer
of subunitscon-tains, in addition to protein VII and the viral
DNA viral proteins V, VI, and VIII. Everitt et
al. (18) have estimated the numberofcopiesof
each ofthe proteins per virion with the
excep-tion of protein VIII. Using the
procedure
de-scribed abovewecalculated the total volumeof
theseother proteins, exceptVIII, tobe 4 x 104 nm3. When this volume is combined with that calculated for the DNAandprotein VIIcore, it
amounts to a total ofabout 1.2 x 105 nm3 and
accounts for allbut 11%oftheinternal volume
of the virion which should be occupied by
protein VIII andwater.
(ii)
The protein components of the viralcore. Selective proceduresforthe stepwise
deg-radation of the adenovirion proved useful to
elucidate its structure (16, 18,
22, 33,
39). Wehave used heat (56
C) (39),
pyridine (33),
and sarcosyl(11)treatmenttodisrupt the virion andget access to the core. Each of these agents
produces a specific type of subviral particle.
Althoughsubviralparticles preparedbyheating
purifiedvirions to 56C (39), by pyridine
(33),
orby sarcosyltreatment have some similarities in
appearance in electron micrographs of
nega-tively stained preparations (Fig. 6, 7, and 8),
theyarestrikingly different in theirpolypeptide
composition(Fig. 12). Virions heated to 56 C for
5min andreisolatedbyequilibrium
centrifuga-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
CORES FROMADENOVIRUSTYPE 2
383
10 20 30 40 50 60 70 80 90 100 110 120 130
[image:18.503.105.392.74.479.2]Fraction Number
FIG. 13. Cleavage of viralDNA insarcosyl cores with the EcoRI restriction endonuclease. Sarcosyl cores were prepared from [3H]thymidine-labeled Ad2, dialyzed against TE, and incubated with EcoRI restriction endonuclease. Allpertinent techniques have been described. Immediatelyprior to analysis of the cleavage
products byelectrophoresis on 1.5%polyacrylamide-0.8%agarosegels, 1.4to2.8ggof "4C-labeledAd2 DNA
previously cleaved withthe EcoRI endonuclease inaseparateincubationwasaddedasamarker. Subsequent
to the incubation with EcoRIendonuclease, themarkerwas extractedfrom the incubation mixtureby
SDS-Pronase treament andphenolization. Beforeelectrophoresis allsamples weredialyzedagainst 1/10 strength
ofTEBelectrophoresis buffer. (a)Incubation ofsarcosylcoreswithEcoRIendonuclease, followed byPronase
treatment.An amount of sarcosyl particles equivalent toapproximately 1.5Ag ofAd2 DNA was incubated under standard conditions with EcoRI endonuclease for 60 minat 37 C. The reaction was stopped by the additionof5M1of0.2MEDTA, 10
Al
ofPronase(5mg/mI),and10Al
of10%SDS and incubationat37Ccon-tinuedfor30min. (b) Omissionof Pronase incubation. Experimentalconditions wereessentially identicalto
thosedescribed under(a), except thatan amountofsarcosylparticles equivalent to0.8Ag of Ad2 DNA was
used and that Pronase was notused in the incubation subsequent tocleavage by EcoRI endonuclease. The
gelslice infraction1waseluted in0.2mlof0.1%SDS andfurther analyzedasdescribed under(c). (c)Pronase
treatment of theDNA-protein complex. The materialremaining on top ofthegel described under(b) and elutedfrom thegelslice(fraction 1) in0.2 ml of0.1% SDS wasincubatedat 37C with 20M'ofPronase(5
mg/ml)and20
Al
of10%SDSfor45min. Thereaction mixturewaschilled,and"4C-labeled Ad2 DNAfragments(EcoRI)
wereaddedpriortoelectrophoresis. VOL-16,1975on November 10, 2019 by guest
http://jvi.asm.org/
384 BROWN ET AL.
tion in CsCl density gradients contain most of
theviralpolypeptides (results not shown),
pyri-dine cores (18, 33) contain polypeptides V and
VII (Fig. 12, slot no. 5), and sarcosyl cores
contain predominantly polypeptide VII and
onlyminor amounts ofpolypeptidesVIIIand IX
and hexons (Fig. 11).
Inthe model oftheadenovirion suggestedby
Everitt et al. (16), theviralpolypeptideVis in
close neighborhood to polypeptide VII and the
penton base, and polypeptide VII is tightly
bound to the viral DNA. In the sarcosyl cores
described in thepresent report, polypeptideVII
isthe predominantproteincomponent, whereas
polypeptide Vcan be removedfrom thecore
by
sarcosyl treatment. Obviously, polypeptides Vand VII must be anchored in different ways in
the viral core. The electron microscope data
presented (Fig. 3, 4, 7, 8) are also consistent withthisinterpretation (see belowandFig. 14).
In fact, the arginine/alanine-rich polypeptide
VII can be quantitatively separated from the
viral DNA
by
proteolytic
enzymes, or alkalitreatment, or
by
boiling
in SDS. Ourdata,
however, do not
permit
us to draw definite conclusions about the type oflinkage betweenn
IV
FeP
V
VI
VIII
Ix
FIG. 14. Model ofthe type 2 adenovirion. The Roman numerals refer to the viralpolypeptides (26). The
locationof the fractureplanes(FP1andFP2)isderived from electronmicroscopedata (seetext and Fig. 3, 4, 7,
and8). The particlesgenerated by cleavage inFP1isprobably similar to pyridine cores, the particle that arises
by cleavageinFP2isprobably similar to sarcosyl cores. The arrangement of the viral DNA molecule as shown in
the model was chosen not to presuppose any definite organization. The loops extending into each of the
substructures in the model in reality represent tightly packed coils of viral DNA (see Fig. 9). The model represents a modification of the scheme proposed by Everitt et al. (16).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:19.503.98.425.220.591.2]CORES FROM ADENOVIRUS TYPE 2
385
polypeptide VII and the viral DNA. The poly-peptide analysis of sarcosyl cores by electro-phoresis on SDS-polyacrylamide gels gave very
similar results, regardless ofwhether the
sarco-syl cores were pretreated with DNase or were electrophoresed without predigestion of the DNA (Fig. 11).
Our ultrastructural studies revealed that the
sarcosyl core was atightlypacked sphere which
gradually unfoldedupon storage at 4 C(Fig. 9).
It is not understoodwhat typesof interactions,
DNA-proteinorprotein-protein, areresponsible
for the packing of the DNA or whether other
factors such as polyamines may play a role in
the organization ofthe viral DNA in the core.
The gradual unfolding of the sarcosyl core is
perhaps a consequence of the removal of all otherviralpolypeptideswhich serve tostabilize
the core structure. It is conceivable that
poly-peptide V is at least partly responsible for this
stabilization. Differences in ionic strength and
pH valuesintherangebetween 7.2and8.5did
notalterthemorphologyofthe sarcosylcores or.
noticeably affect the unwinding process.
(iii) The viral DNA. The viral DNA in the sarcosyl core can be
completely
digested
with DNase and is susceptible to cleavage by the EcoRI restriction endonuclease. The datapre-sented in Fig. 13 demonstrate that the termini ofthe linear Ad2 DNA molecule are linked to
each other probably by a protein, since this linkage can be disrupted by treatment with proteolytic enzymes (Fig. 13c). The finding that, uponincubationofsarcosylcoreswiththe EcoRI restriction endonuclease, a complex
be-tween protein and the terminal EcoRI
frag-ments A andC canbe isolatedontop ofthegel
(Fig. 13b) furtherargues forthe involvementof a
polypeptide
inthislinkage.
Thenatureofthepolypeptide(s)
in thiscomplex hasnotyetbeen determined.TheviralDNA inthesarcosyl coreshasbeen showntobea
circular
molecule(11;
Fig. 10)
and circular adenovirus DNA has also been foundafter treatment ofpurifiedvirionswith
guanidi-nium hydrochloride (36). These circular DNA
molecules are converted to a linear form
by
proteolytic enzymes and the linear Ad2 DNAhas a unique melting pattern (12), hence the
circular moleculemustbe
opened
by
proteolysis
at one specific site.
We concludethat the complex between viral
EcoRIDNAfragmentAandC and the
polypep-tide(s)responsible
for circularization of theviral DNA has been isolated in theexperiment
described in Fig. 13b. It is conceivable that
polypeptide VII is involved in the
circulariza-tion oftheviral DNA,
however,
anadditional,
morespecialized polypeptide may be present in
minuteamounts and may hithertohaveescaped
detection.
We also consider the possibility that the adenovirus DNA isa covalently closedcircular
molecule and that an activated form of the
virion-associated endonuclease (7) cuts the
cir-cle at the site of thespecificpalindrome dueto
the terminal repetition ofthe adenovirus DNA
(19, 46). This endonuclease could be activated upon disruptionofthestructural integrity ofthe
virion.
(iv) Correlation between morphological
and biochemical findings. The presence of
protein V inthe pyridine cores and itsabsence
inthe sarcosylcorescorrelates withthe presence ofthe haloofmaterialsurrounding the21.6-nm
core subunits in the particles produced by
pyridine, and the absence of this halo in the
particles prepared by sarcosyl treatment.
Pro-tein V seems to coat the protein VII-DNA
complex and togive the pyridinecores a
stain-ing characteristic which generally makes the internal structure non-distinguishable. In the freeze-cleaved virions, twofractureplanes were found inthe interiorofthevirion. One revealed thecoreas asmoothstructure,the other showed
thepresence ofsubunits. It isconceivable that
in the former case the fracture plane passed
between the outercapsid and protein
V,
and in the latter case between protein V and theproteinVIIDNAsubunits (Fig. 14).The smaller
size ofthecore subunits seen infreeze-cleaved
virions (Fig. 3, 4) is probably the result ofthe
fracture plane passing over the subunits at a
pointsomewhat distantfromthetrue
cross-sec-tion of a sphere. The most difficult aspect of
fittingthe viralcoreinto thecapsidis todefine
what types of interaction occur
between
theDNA-protein VII complex and the hexons and
pentons of the outer shell. Everitt et al. (16)
have elegantly described a number of
protein-protein interactions within the virion which
may explain the
anchoring
ofthe core in the viralcapsid. ProteinV seems tobindtoboth thepenton and theprotein VII-DNA
complex
(17)and could therefore attach each ofthe 12 core
subunits to one ofthe 12 penton structures in
the outer
capsid.
The strongbinding
ofhexonsto the oversize relaxed subviralstructures
pro-ducedby heating (Fig. 6b) suggests an
attach-ment ofthe hexons to the corestructure
under-neath. Such an interaction could be mediated byoneorbothofthe
internally
locatedproteins
VI and VIII. Our model of the adenovirion
correlating ultrastructural and biochemical findings (Fig. 14) is a modification of the model
proposed
by
Everittetal.(16).
VOL. 16,1975