Rochester Institute of Technology
RIT Scholar Works
Theses
Thesis/Dissertation Collections
11-21-1995
Non-steady time series analysis in the developing
embryo
Daniel Kettles
Follow this and additional works at:
http://scholarworks.rit.edu/theses
This Thesis is brought to you for free and open access by the Thesis/Dissertation Collections at RIT Scholar Works. It has been accepted for inclusion
in Theses by an authorized administrator of RIT Scholar Works. For more information, please contact
Recommended Citation
NON-STEADY TIME SERIES ANALYSIS IN
THE DEVELOPING 'EMBRYO
A Thesis Submitted in Partial Fulfillment of the Requirements for the
Masters of Science Degree in Mechanical Engineering
at Rochester Institute of Technology
on November 21, 1995
By
Daniel E. Kettles
Approvals:
Dr. Charlie Haines, Department Head
Dr. Mark H. Kempski, Thesis Advisor
Dr. Kevin Kochersberger, Thesis Committee
Release
I, Daniel
E.
Kettles, do hereby give permission to Wallace
Memorial Library to reproduce this Thesis, entitled
Non-Steady Time
Series Analysis in the Developing Embryo,
in
whole or
in
part. Any such
reproductions may not be for commercial use or profit.
Signed,
Daniel
E.
Kettles
Acknowl
edgments
To
recognize all ofthe
people whohelped,
influenced
or encouraged mein
someway
whileI
wasworking
onthis
thesis
wouldbe nearly
impossible.
However. I
wouldlike
to
thank the
following
people who cometo
mind.My
parents,
Frank
andCynthia
Kettles,
along
the
rest ofmy
family
at105
Green Clover Drive for
their
emotionalsupport;
My
graduateadvisor,
Dr. Mark
Kempski,
for his
support andencouragement;
Dr. Edward Salem
andDr. Kevin
Kochersberger,
for accepting my
invitation
to
be
onmy
thesis committee,
andfor
agreeing
to
reviewmy
thesis
so closeto
final
exams andthe
end-of-quarterrush;
Shri,
Mike,
Andrew
and everyone else atthe
Biosystems Lab
(including
the
students who usedthe
lab
as ahangout!)
for
their
encouragement andassistance;
Table
of
Contents
Table
ofContents
i
List
ofFigures
iii
Abstract
viii1.
Introduction
1
1
.1
Hemodynamics
ofthe
Embryonic
/
Fetal
Cardiovascular System
1
1.2
Justification
for Research
6
1.3
Research
Goals
7
2.
Methodology
10
2.1
Methods Employed
in
Previous Work
10
2.2
Relationship
to
Other Research
13
2.3
Removing
Spurious Data
16
2.4
Grouping
Embryo Spectra
24
2.5
Band Power
Determination
28
3.
Results
42
3. 1
Frequency
Domain Effects
ofData Removal
42
3.2
Grouped
Embryonic
Data
50
3.2.1
Algorithm Validation
withTest Data
50
3.2.2
Sample Results
withChick Data
52
3.3
Spectral
Band
Power
58
3.3.1
Algorithm
Validation
withTest
Data
58
3.3.2
Sample Results
withChick Embryo Data
61
3.4
Joint
Time-Frequency
Analysis
3.4.1
Algorithm Validation
withTest
Data
70
3.4.2
Spectrograms
withChick Embryo Data
72
4.
Discussion & Conclusions
79
4.1
Discussion
ofResults
79
4.2
Future Lines
ofResearch
82
References
86
Appendix
A
Discrete Spectral
Analysis
89
A.l
The Discrete
Fourier
Transform
(DFT)
89
A.2
The
Fast
Fourier
Transform
(FFT)
92
A.3
Spectral
Representations
andPower
93
A.4
Zero
Padding
98
Appendix B
Special Case in
Spectral
Analysis-Frequency
Domain
Analysis
ofSectioned Sinusoids
107
Appendix C Algorithm Usage &
Programming
Overview
114
C.l
Edit
and concatenate.vi114
C.2
Linear interpolation Il.vi
120
C.3
Band
power calculation(5 bands).vi
Band
powercalculation(10 bands).
vi127
C.4
Short
time FFTl.vi
130
Appendix D MATLAB Routines
D.l
Spectra.m
137
List
of
Figures
Figure
Description
Page
Section
1
1-1
Transformation
ofthe
heart
tube
into
the
maturefour
chambered
heart.
2
1-2
Schematic
ofthe
cardiovascular control mechanismsresponsible
for
heart
ratevariability
in
the
matureheart.
4
Section
2
2-1
Interrelationship
between
algorithms usedin
this
work andpreviously defined
algorithms.14
2-2
Derivation
of cardiovascularvariability
time
series.15
2-3
Corrupted human fetal blood
velocity
data.
17
2-4
(a)
Blood
velocity
time
series with corruptionfor 'tail
to
head'concatenation example.
(b)
Results
of'tail
to
head'concatenation.19
2-5
(a)
Schematic
ofthreshold
crossing
concatenation method.(b)
Resulting
velocity
time
series afterthreshold
crossing
concatenation.
21
2-6
Concatenated
velocity
data from Figure 2-5
with subsequentscaling.
23
2-7
Typical
result ofinterpolating
andaveraging
two
embryospectra.28
2-8
'Chirp'data
usedfor
JTFA
example.33
2-9
Discrete PSD
ofthe
chirp'
signal
from Figure
2-8.
34
2-10
Three
dimensional
time-frequency
analysisrepresentationof a'chirp'
Figure
Description
Page
2-11
Basic
procedurefor
shorttime
Fourier
transform.
37
2-12
Block
diagram
ofmoving
overlapped windows on adata
set.39
Section 3
3-1
Sinusoid
with section removed as an example ofsectioning
outdata from
asinusoidally varying data
set.44
3-2
Superimposed
amplitude spectrafor
the
original and alteredsinusoids
in
Figure 3-1.
45
3-3
Three
dimensional
amplitude spectrumrepresentationof asinusoid with an
incrementally increasing
section removedfrom
the
middle.47
3-4
(a)
Interpolated
peakvelocity variability
PSD
for
two
Stage
14
embryos.(b)
Average
PSD.
54
3-5
(a)
Interpolated
peakvelocity variability
PSD
for
two
Stage
27
embryos.
(b)
Average PSD.
55
3-6
(a)
Interpolated heart
ratevariability
PSD
for
two
Stage
14
embryos.(b)
Average PSD.
56
3-7
(a)
Interpolated
HRV PSD for
two
Stage
27
embryos.(b)
Average PSD.
57
3-8
PSD
ofthe
sequence givenby
eq.(3.1)
in
arbitrary
units.60
3-9
Stage 14
peakvelocity
variability
spectralband
power.63
3-10
Stage 27
peakvelocity variability
spectralband
power.63
Figure
Description
Page
3-12
Stage
27 HRV
spectralband
power.64
3-13
Stage
14
peakvelocity variability
spectral powerdistribution.
66
3-14
Stage
27
peakvelocity variability
spectral powerdistribution.
66
3-15
Stage 14 HRV
spectral powerdistribution.
67
3-16
Stage
27 HRV
spectral powerdistribution.
67
3-17
Stage
14
HRV
spectral powerdistribution
with10
.25Hz
wide
frequency
bands.
69
3-18
Stage 27 HRV
spectral powerdistribution
with.25Hz
frequency
bands.
69
3-19
Spectrogram
oftest
data
seriesin
eq.(3.4).
72
3-20
(a)
Stage 14
chick embryoblood velocity data
segment.(b)
Spectrogram
ofStage
14
chick embryoblood
velocity
time
seriesdata.
74
3-21
(a)
Stage
14
chickembryo peakblood
velocity variability
time
seriesdata.
(b)
Spectrogram
ofStage
14
peakvelocity
variability
time
series.75
3-22
Peak
velocity
variability
spectrogram with1024
point windowlength
and256
pointincrement.
77
Appendix
A
A-l
The
frequency
domain
effects ofsampling
ananalog
signal.91
A-2
Sample
discrete
PSD
showing
the
development
ofintegration
Figure
Description
Page
A-3
(a)
Discrete
PSD
ofChirp
data
without zero padding.(b)
Discrete PSD
of zero padded 'chirp'data
withoutzeropadding
correction.(c)
Discrete PSD
of zero padded'chirp'
data
withzeropadding
correction.
101
A-4
Superimposed
Harming
and rectangular windows.104
A-5
Sinusoid from
eq.(A.15)
with aHanning
window applied.105
A-6
PSD
(log
magnitudeform)
of sinusoid with and withoutHanning
window applied.105
Appendix B
B-l
Sinusoid
withsection removed.108
B-2
Individual
sinusoidsthat
sumto
form
a sectioned sinusoidAbstract
Cardiovascular diseases
with originsin
early fetal
development
are aserious
health
concernin
the
United States.
Presently,
there
arefew,
if
any,
diagnostic
tools
availablefor doctors
to
determine
the
overallhealth
ofthe
early
fetal
cardiovascular systemin
a non-invasive manner.This
thesis
is
astep
toward
the
development
of a non-invasive systemfor
the
detection
ofcardiovascularmalformations
in
the
early human
fetus. The
computeralgorithms presentedin
this
workdovetail
with other algorithmsto
represent embryonic/fetal
cardiovascular output and
variability
data
in
the
frequency
domain. Algorithms
in
this thesis
allowthe
capability
to
editexternal artifactfrom
data,
determine
common
frequency
spectraforms
and calculate spectralband
powerto
aidin
study group
comparison.Additionally,
an algorithmis
presentedto
represent1.
Introduction
Congenital
cardiovascular malformations are a serioushealth
concernin
the
United
States
(Clark
andTakao 1990). At
presentthere
are no clinicaltools
to
asses
the
health
ofthe
early
embryonic/fetal
cardiovascularsystem.However,
it
has been hypothesized
that
beat
to
beat variability
in
embryonic/fetal heart
rateand
blood velocity
correlatedirectly
to
cardiovascular wellbeing
(van
Ravenswaaij-Arts
1993,
Breborowicz
1988,
Kempski 1995).
This
thesis
is
astep
toward the
development
of a non-invasivetechnique
for assessing
the
cardiovascularhealth
ofthe
10
to
18
weekhuman fetus.
Early
detection
of abnormalfetal
cardiovascular output canlead
to
more effectivetreatment
protocolsearly
in
gestation.This
sectionbegins
with an overview ofthe
development
andhemodynamics
ofthe
embryonic cardiovascular system.Later,
an overview ofcongenital cardiovascularmalformations
is
presented as ajustification
for
the
research.
This
section concludes with asummary
of previous work(Gallagher
1995)
whichdirectly
impacts
the
scope and objectives ofthis thesis.
1.1 Hemodynamics
ofthe
Fetal
Cardiovascular System
The
cardiovascularsystemis
the
first
functioning
organin
the
embryo/fetus.
Through
a complexprocessknown
asmorphogenisis, the
to
afour
chamberedheart.
In
the
human
fetus,
the
musclewrappedheart
tube
begins
beating during
the
third
gestational week.For
the
nextthree to
four
weeks,
mechanicalforces
relatedto
function,
morphogenisis and growthtransform
the
heart
tube
into
the
maturefour
chamberedheart (Clark
andVan
Mierop
1989,
Marieb
1992)
(Figure
1-1). Few
majorstructuralchangesoccurin
the
heart
during
midto
late
gestation.Superiorvenacava Right pulmonary artery Pulmonarytrunk Right Atrium Right pulmonaryveins Fossa ovalis Pectinate. muscles Tricuspid valve Bightventricle Chordae tendineae Trabecuiae carneae Inferior vena cava Aorta Leftpulmonary artery Lettatrium Lettpulmonary veins Pulmonary semilunarvalve Bicuspid (mitral)valve Aortic semilunarvalve Leftventricle Papillary muscle Interventricular septum Myocardium Visceral pericardium
Figure 1-1.
Transformation
ofthe
heart
tube
into
the
maturefour
chamberedThe early
embryo/fetal
cardiovascularsystem,
like
that
ofthe
maturesystem,
is
regulatedto
meetdemand
(Kempski 1993
and1995,
Clark
1990).
Unlike
the
maturesystem,
however,
variationsin
embryo/fetalcardiac outputearly
in
development
are not regulatedby
the
central nervoussystem.Autonomic
cardiovascular controlin
the
chick embryodoes
notbegin
until afterStage
271(Pappano
1977,
Sissman 1970).
Thus,
the
early
cardiovascularsystemdistributes
oxygen andnutrientsto
the
growing
embryo withoutthe
sophisticated regulation mechanisms present
in
the
mature system.The
mature cardiovascularsystem,
in
contrast,
is
regulatedby
a complexfeedback
control system(Figure
1-2). Beat
to
beat
pacing
is
controlledby
sympathetic and
parasympathetic2 nervous system
activity
andfeedback
occursthrough
respiratory
sinusarrhythmia3,
baroreflex"
and
thermoregulation
(Marieb
1992,
vanRavenswaaij-Arts
1993,
Kempski
1995,
Saul
1991).
1
'Stage27'
is from
the Hamburger-Hiltonscalefor
measuring
chick embryodevelopment. Stage
27 is approximately 51/2 days
ofgestation,and correlatestoabout37
days
ofhuman fetal
gestation(Sissman 1970). 2
The
sympatheticnervous system causesheart
ratetoincrease in
responsetocrisis situationsand exercise.The
parasympathetic nervous system counterstheaffectsofthesympatheticnervous system oncethecrisis situation or exercisehas
passed.3
Respiratory
sinusarrhythmiaisa phenomenathatrelatesthefrequency
ofheart
ratevariability
to thebreathing
rate.4
Vagus
SANode
ILVtt) respiration
K
Sjrnnparetics
LPF
0 Hz 05
LPF
0 Hz 05
t
hea-thtt)
A
ventriclesand vasculature
bar
preceptorsarterial pressure
ABPCt)
respiration ILVtt)
*dt
Figure
1-2.
Schematic
ofthe
cardiovascular control mechanisms responsiblefor
heart
ratevariability
in
the
matureheart.
Redrawn
from Saul
1991.
Less is known
aboutthe
exact mechanisms offetal
cardiac control andnormalcardiac output
early
in
gestationthan
in
the
mature system.However,
research on
White
Leghorn
chick embryos(Kempski
1993,
Hu
andClark
1989)
and
sheep
embryos(Adamson
1992)
has
indicated
that
four determinants
ofmyocardial contractility.
These four
determinants
are also presentin
the
maturecardiovascular system
(Clark 1990).
Preload is
determined
by
diastolic,
orventricularfilling
events.Changes
in
preload occurin
responseto
environmental and metabolicdemands
in
the
embryo.
The
Frank-Starling
Law
of
the
Heart,
whichdescribes
the
biomechanical
properties of
stretching heart
muscle,
is
abeat
to
beat
regulation phenomenon.According
to the
Frank-Starling
Law
ofthe
Heart,
stroke workis
controlledby
the
amount of preload onthe
cardiac muscle cellsjust
before
they
contract.More
details
onthis
process canbe
found
in
ref.(Marieb
1992).
Afterload
refersto the
impedance
ofthe
vascularbed. Vascular
impedance is
due
to
a combination of resistanceto
flow,
fluidic
capacitance,
blood
inertance,
varying
blood
vessel walltension
and vessel caliber.In
the
chickembryo,
vascularimpedance is
alsoinfluenced
by
temperature.
In
addition,
neurohumeral
agents5
present
in
the
blood
stream ofthe
early
chick embryo(2
to
6 days
of a21
day
gestation)
are speculatedto
provide somehemodynamic
regulationrelated
to
afterload(Hu
andClark
1989,
Clark
1990,
Nakazawa
1989).
Heart
ratein
the
chickembryois
temperature sensitive,
or poikilothermicin
nature.
It
has
been
shownthat
as embryonictemperature
increases,
the
heart
rate(and
cardiacoutput)
increases,
while asthe
embryocools,
the
heart
rate5
Neurohumeralagents are chemicals present at nerve endingsthatexcite adjacentstructures,
decreases.
Embryo
temperature
alsohas
aninverse
relationship
to
afterload(Nakazawa
1985). The
heart
rate ofthe
human
fetus, however,
is
regulatedby
hypoxia
andhyper
capnict\
Myocardial
contractility
canbe
shownthrough
the use ofthe
pressure-volumeloop,
which relatesinstantaneous
ventricular pressureto
volume(Clark
1990). The
total
area enclosed withinthe
pressure volumeloop
is
a measure ofthe
amount of myocardial work(Suga
1979).
1.2
Justification
for Research
Cardiovascular
malformations are a serioushealth
problemin
the
United
States.
Specifically,
for every
1000
births,
8
to
10 infants
have
somekind
of congenital cardiovasculardefect (Clark
1990).
In
many
cases, the
defect
canbe
corrected with post natal surgery.However,
20%
of allinfants
withcardiovascular malformations will
die before
their
first
birthday
regardless ofheroic
surgicalintervention.
Examples
of cardiovascularmalformationsinclude
myocardial(heart
muscle)
defects,
valvedefects
andtetrology
ofFallot7. Despite
advanced surgicaltechniques
to
correctmany fetal
cardiovasculardefects,
there
is
still alingering
morbidity
that
limits
the
life expectancy
ofmany
children and adults with congenitalheart
defects. In
addition,
some adult cardiovascular6
Hypoxia
referstolow blood
oxygenlevels. Hypercapnia
referstohigh blood
carbondioxide
levels.
7
Tetrology
ofFallot is
a seriousheart defect
involving
many
conditions,including
constrictionofdiseases,
such ascoronary artery
disease
andhypertension
may have
their
origins
early
in
cardiacdevelopment
(Clark
1990).
1.3
Research Goals
The
long
term
goals of research relatedto this thesis
areto
correlatebeat
to
beat
andlong
term
variability
in
embryonic/fetal
cardiovascular outputto
cardiovascular well
being.
To
accomplishthis,
a control system approach willbe
used
to
analyzevariability
in
fetal
cardiovascularoutput,
similarto that
shownin
Figure
1-2.
The
computer algorithms presentedin
this thesis
dovetail
withothers
(Gallagher
1995),
and willbe
usedto
quantify
'normal' and'abnormal'
fetal
heart
rate andblood
velocity
variability.Using
results generatedfrom
the
algorithms presentedin
this thesis
andothers
(Gallagher
1995),
temperature
regulationin
chickheart
ratevariability
canbe
investigated.
For
the
human
fetus,
cardiovascular control mechanisms whichaffect
heart
ratevariability
canbe
scrutinized.In
addition,
analysis of peak andmeanpulsatile
blood
velocity variability
canbe
usedto
identify
othercardiovascularcontrolmechanisms such as vascular
impedance.
Use
ofhuman
data. The
algorithms presentedin this thesis
willbe
usedto
analyze cardiovascular
data from both
chick embryos andhuman fetuses.
It is
very
important
to
notehere
that
human fetal
cardiovasculardata
is
obtainedgranted
before any fetal data
is
acquired and analyzed.In
addition,
data from
other
subjects,
such asadultsandneonatesmay be
analyzedin
the
future
using
these algorithms,
subjectto
full
consent.The
algorithmsdescribed herein
serveto
supplementsoftware algorithmswhichacquireandreconstruct
Doppler
velocity
waveforms.In
addition, the
currentalgorithmsseek
to
extendthe
data
analysis capabilities whenperforming
power spectral analysis on
blood velocity
andheart
ratevariability
waveforms.Due
to
suchfactors
asvariability
amongst embryonic/fetalstudy
groups,
spurious
inputs
due
to
movement artifact orthe
non-steady
nature ofcardiovascular
time series,
additionalpost-processing
is
implemented
using
the
algorithms presented
here.
Thus,
the
specific goals ofthis thesis
are:To
present an algorithmto
'edit'spuriousdata from
ablood
velocity
time
series.
The
spuriousdata
may be
presentdue
to
embryo/fetus
movement,
maternal movement or
instrumentation
noise.This
algorithmis
to
be
used onreconstructed
velocity
time
seriesbefore any
analysisis
implemented.
To
compensatefor
variability
among
embryos/fetusesin
a particularstudy
group
anddifferences
in
data
recordlengths,
an algorithmis
presentedto
interpolate
groups of spectraldata
setsto
a commonfrequency
axis.Quite
oftenin
spectralanalysis,
it is
desirable
to
calculatethe
amountofspectral
'power'
Dominant
frequency
bands
canbe
therefore
determined for heart
rate andblood velocity
variability
in
the
embryo/fetus.
Thus,
an algorithmis
presentedto
calculatethe
amount of'power'contained within specificfrequency
bands
ofthe
PSD
data.
Cardiovascular
outputis
non-steady
in
nature.In
otherwords, the
regulatory
mechanisms(afterload,
blood
oxygenation,
chick embryotemperature
etc.)
causethe
componentfrequencies
ofblood velocity variability
andheart
ratevariability
to
vary
overtime.
Spectral
analyses of adata
settaken
in
toto
cannotdescribe
how
orwhenthe
frequencies
change.Thus,
an algorithmis
presentedto
showthe time-dependent
spectral content of cardiovascularvariability
data.
The
methodologiesbehind
these
goals are presentedin
the next section.2.
Methodology
In
orderto
correlate variationsin
cardiovascular output withcardiovascular control
mechanisms,
mathematical methodsthat
accurately
quantify
these
variations are needed.Recall
from
the
previous sectionthat
variations
in
heart
rate andblood velocity may be linked
to
hemodynamic
control mechanisms
in
the
embryo/fetus. The
control mechanisms areinfluenced
by
afterload, thermoregulation
(in
the
chickembryo)
and oxygenregulation(in
the
human fetus).
The
analysis ofthe
cardiovascular outputvariations,
eitherin
the
time
domain
orthe
frequency
domain,
canhelp
explain andquantify
the
biological
mechanismsfor
embryonic/fetal
cardiovascular control.In
this section,
an overview ofthe
methodologiesthat
are utilizedin
this
thesis
is
presented.Theories behind
these methods,
along
withthe
applicablemathematical
derivations
arepresentedin
Appendices A
andB.
Programming
and usage
details
for
the
software algorithmspresentedin
this thesis
are givenin
Appendices
C
andD.
2.1
Methods
Employed
in Previous Work
Much
workhas
been
done
to
quantify
the
variability
of cardiovascularsignals,
particularly
withheart
ratevariability
in the
adult andinfant.
Two basic
time
domain
analysis(van
Ravenswaaij-Arts
1993,
Parer
1985,
Yana
1993)
and
frequency
domain
(spectral)
analysis(van
Ravenswaaij-Arts
1993,
Kempski
1993,
Saul
1991,
Akselrod
1985,
Breborowicz
1988,
Kuo
1993,
Adamson
1992,
Goldstein 1993
and1994)
Time
domain
methods.There
are a number ofdifferent
methodsavailable
to
analyze cardiovascular signalsin
the time
domain.
One
approachinvolves
the
calculation ofshort-and long-
term
indices
directly
from
the time
series representation of
the
cardiovascular signal(van
Ravenswaaij-Arts
1993,
Parer
1985).
Short
term
variability
involves
the
changes ofthe
cardiovascularsignal on a
beat
to
beat basis.
Long
term
variability
seemsto
have different
interpretations
depending
onthe study,
but
canbe
thought
of as alonger-duration
changein
heart
rate(i.e.,
over severalcardiovascular cycles).One form
ofthe
short- andlong-term
indices involves
the standarddeviation
from
the
meanheart
rate.Others involve
such methods as modificationof
the mean,
sorting
andselecting,
and calculations ofslope changes. Short- andlong-
term
index
methodshave
been
shownto
be computationally
inexpensive
ways
to
categorize shortterm
heart
rate variability.However,
only
one of nineindices
validly
measuredlong
term
variability
(Parer 1985).
Since
the
range offrequencies below 1
Hz1(Kempski
1993,
Akselrod
1985,
Breborowicz
1988,
1
Goldstein 1993
and1994)
is
ofprimary
interest
in
evaluating
cardiovascularfluctuations,
thedetermination
ofvariability
in
the
sub-heartrate rangemay be
problematic
using
the
above methods.There
are other methods availablefor analyzing
cardiovascularsignalsin
the
time
domain. Autocorrelation
is
essentially
the
time
domain
comparison of adelayed
version of a signalto
itself,
andis
equalto the
inverse
Fourier
transform
of a
Power
Spectral
Density
(PSD)
representation ofthe
signal(Stremler
1990).
The
subsequentFourier
transform
ofthe
autocorrelationfunction
yieldsthe
PSD
of
the
signal.Autocorrelation
is
usedin
applicationswherethe
signalsto
be
analyzed are corrupted
by
noise(Stremler
1990). A
possibledrawback
to
autocorrelation,
however,
is
that
it
adds a secondstep
(in
the time
domain)
to the
computation of the
frequency
domain
representation of a signal.Frequency
domain
(spectral)
analysis.Using
spectralanalysis, the
variations of
heart
rate and peakblood
velocity
canbe
representedby
their
frequency
components.The
Fourier
transform
is
thebasis
for
frequency
domain
analysis
(see
Appendix
A). In
essence,
frequency
domain
methodologiesinvolve
the
decomposition
of periodic or aperiodic signalsinto
a summation of sinewaves at
different frequencies. For digital
computerimplementation,
the
Fast
Fourier
Transform
(FFT)
algorithmis
primarily
used.Thanks
to the
advent ofbecome very
popularin
signalprocessing
applications,
including
cardiovascularsignals
(van Ravenswaaij-Arts
1993).
The
abovetime
andfrequency
domain
methods allowthe
quantificationof cardiovascularoutputas
it
varieswithtime.
Methods
have
alsobeen
developed
to
correlatecardiovascularoutput(in
adulthumans
anddogs)
to
control system
models,
including
systemimpulse
response(Yana
1993)
andtransfer
functions
(Saul 1991). Similar
methods canbe
usedin
future
researchto
develop
control parametersfor
the
fetal
cardiovascular system.2.2
Relationship
to
Other Research
The human
fetal
blood
velocity
data
used withinthis thesis
is
providedby
Dr.
Juriy
W. Wladimiroff (and
colleagues)
ofAcademic
University
Hospital
in
Rotterdam,
The
Netherlands.
The
chickembryodata,
which was used as 'pilot'data for
algorithmdevelopment,
wasprovidedby
Mr. Norman Hu
ofthe
University
ofRochester
andDr. Monique L. Broekhuizen
ofAcademic
University
Hospital,
Rotterdam.
The
algorithmsdeveloped for
this thesis
are usedin
concert with othersdeveloped
by
Francis J. Gallagher (1995).
All
custom analysis algorithmsfor
this
thesiswere
developed
using
Lab
VIEW2
graphical
programming
softwarefor
instrumentation
(see
the
end ofthis
sectionfor
alisting
ofthese
algorithms).A
flow
chartshowing
the
interrelationship
between
the algorithmsis
shownin
1
Figure
2-1
onthe
next page.The
algorithmsshownin
gray boxes
involve the
derivation,
spectralanalysisand normalization ofthe
cardiovasculartime
series.They
aredetailed further
in
(Gallagher,
1995).
Velocity
Time
Series
Reconstruction
From Video Tape
From
Data File
i
Edit Spurious
Information from
Velocity
Data
+
Generate
Variability
Waveforms
1 IPeak Amplitude
Variability
Heart
Rate
Variability
i
PSD
Analysis
onindividual
Embryos
Normalized
Non-Normalized
*
Interpolate
PSDs
to a
Common
Frequency
Axis
i
Band Power
Calculations
-?Joint
Time-Frequency
Analysis1
JTFA Spectragram
Time Variable Band Power
Leqend:
=Previously
generated algorithms(Gallag
her,
1995)
Figure 2-1.
Interrelationship
between
algorithms usedin
this
work andCardiovascular
variability
time
series(Gallagher,
1995).
Two
forms
ofcardiovascular
variability data
are ofinterest
for
spectral analysis(see
Figure
2-2). Instantaneous Heart Rate
andHeart Rate
Variability
are obtainedfrom
the
beat
to
beat periodicity
(AT;)
at a threshold crossing.The
threshold
crossing
is
equal
to
the
global meanblood
velocity.The beat
to
beat periodicity is
inverted
(1/AT;)
to
obtainthe
instantaneous heart
rate.Heart
ratevariability
is
obtainedby
subtracting
the
global meanheart
ratefrom
the
instantaneous
heart
rate.Peak
velocity variability
(AVpi)
representsthe
beat
to
beat difference between
adjacentamplitudes.
The derivations
ofinstantaneous heart
rate and peakvelocity
variability
arediagrammed
in
Figure
2-2.
o v (A
u o
>
Blood
Velocity
Profile (13 Week Human
Fetus)
Threshold
1
Av
T
AT,
Time
(sec.)
Figure
2-2. Derivation
of cardiovascularvariability
time
series.The FFT
ofthe time
seriesdata is
convertedto
the
Power
Spectral
Density
(mm/sec)2/Hz for
peakvelocity variability
spectra.The
frequency
axisis in Hz
(cycles/second).
Therefore,
the
PSD
spectrum canbe
thought
of as adistribution
of signal spectral power per unit
frequency.
Normalization
of spectra.Because different
embryos/fetuses
at a givengestational age
may have different
meanheart
rates and meanblood
velocities,
normalization of
the
spectral(PSD)
representationsis
performed.PSDs
of peakvelocity variability
data
are normalizedby
thesquare ofthe
cardiac cycle meanvelocity
ofthe
respectivedata
series.PSDs
ofthreshold
crossing
heart
ratevariability
are normalizedby
the square ofthe
meanheart
ratedetermined from
the
data
series.The
frequency
axes of all normalizedPSDs
aredivided
by
the
mean
heart
rate(in
Hz)
determined from
the
originalvelocity
time
seriesdata.
Normalized PSD
spectrahave
units of1/Hz,
with unitlessfrequency
axes.The
normalized
PSDs
are usedfor
relative comparisonsbetween different
embryos
/fetuses.
The
algorithms can also generate non-normalized spectra(i.e.,
absolute
PSD data).
2.3
Removing
Spurious Data
Fetal blood
velocity
data
can get corruptedby
fetal
or maternalmovement,
orby
instrumentation
noise.An
exampleof acorruptedblood
velocity
data
setis
shownin
Figure 2-3
onthe
next page.In
this example,
Human Fetal Blood
Velocity
Data
withCorruption
300-r
~ 250
Corrupted
Data20
Figure
2-3.
Corrupted human fetal
blood velocity
data. The
spuriousdata
occursfrom
about4
to
6
seconds and atthe
end ofthe
data
set.Using
the
custom algorithmEdit
andConcatenate.
vi, the
corrupteddata
in
the
blood velocity
time
seriesin
Figure 2-3
canbe
removed.The
algorithm canbe
used
to
remove small sections atthe
beginning,
middle or atthe
end of avelocity
time
series.Concatenation
refersto
joining
the
remaining
two
data
segmentsafter a corrupted section
in
ofthe
originaldata
seriesis
removed.This
algorithmhas
three
different built-in
optionsfor
concatenation.They
are:Concatenate
from
the
end of one signalto the
beginning
ofthe
other('tail
to
head'concatenation),
Concatenate
to
thresholdcrossing
points with subsequent relativescaling
Concatenate
to
thresholdcrossing
points withoutsubsequentrelativescaling.
The
first
type
of concatenationis
simply
the
connectionofthe
remaining
data
sets after a sectionis
removed('tail
to
head'concatenation).With
this type
of
concatenation,
the
last data
point ofthe
first
sectionis
linearly
connectedto the
first data
point ofthe
second section.This
type
ofconcatenationis
notrecommended
for
editing
in
the
middle of ablood
velocity
data
setbecause it
may
interrupt
the
pulsatile nature ofthe
data.
Depending
onthe
exact points ofconcatenation,
a'discontinuity'
may
be
introduced.
An
example ofthis type
ofconcatenation
is
shownin
Figure
2-4. In
Figure 2-4
(a),
the
portion ofthe
data
enclosed
in
the two
verticaldashed lines is
to
be
removed.Using
'tail
to
head'concatenation, the two
remaining
sectionsof the signalare connected at pointsjust
to the
outsideofthe
dashed lines. The
results ofthis type
of concatenationare shown
in
Figure
2-4
(b).
From
the
figure,
it
is
apparentthat the
timing
of theHuman Fetal Blood
Velocity
Data
withCorruption
Remove
10
Figure 2-4
(a). Blood
velocity
time
serieswith corruptionfor
'tail
to
head'concatenation example.
Concatenated Human Fetal Blood
Velocity
Data
250
0 2
Concatenation Point
10
Figure 2-4
(b). Results
of'tail
to
head' concatenation.Threshold
crossing
concatenation.A
'threshold
value'
for
the
velocity
data
is
usedto
determine
the
concatenation points.The
threshold
valueis
Y 4-Y
X= ' 2
(2.1)
2
where
\
and\
arethe
meansofthe
first
and seconddata
segments respectively.(The first
data
segmentis
the
portionbefore
the
removed section andthe
seconddata
segmentis
the
portion afterthe
removedsection.)
The
threshold
crossing
concatenationmethod
is
depicted
in
Figure 2-5. The
first
concatenation pointoccurs
just
before
the
thresholdcrossing
point onthe
rising
edge ofthe
last
pulse(before
the
sectionto
be
removed).The
second concatenation point occursjust
after
the threshold
crossing
point onthe
rising
edge ofthe
first
pulse(after
the
section
to
be
removed).The
threshold
concatenation optionmay, therefore,
resultin
aslightly
larger data
segmentbeing
removedthan
what wasoriginally
specified
to the
algorithm.The
actual size ofthe
removed sectiondepends
onthe
proximity
ofthe
appropriatethreshold
crossingsto
the
specified endpoints ofthe
removed
data
segment.The
threshold concatenation method preservesthe
pulsatile nature of
the
editeddata
by
locating
the
concatenation points relativeto
a
fixed
value(the
threshold).
The location
ofthe
concatenation pointsis
Human
Fetal Blood
Velocity
Data
withCorruption
-Threshold
Second Concatenation Point
FirstConcatenation Point
4 6
Time
(sec.)
10
Figure
2-5
(a).
Schematic
ofthreshold
crossing
concatenation method.Concatenated Human Fetal Blood
Velocity
Data
250
o <D VI E E
200
150
& 100 --o o a > 50
Concatenation Point
4 6
Time
(sec.)
10
Figure
2-5
(b).
Resulting
velocity
time
series afterthreshold
crossing
Comparison
ofFigures
2-4
and2-5
showsthat
the threshold
crossing
concatenation method
is
abetter
methodfor editing
in
the
middle of adata
seriesthan the
'tail
to
head'method.Scaling
afterconcatenation.When
removing
adata
segmentwithinalarger
velocity
time
series, the
mean values ofthe two
remaining
data
segmentsmay be different.
An
optionin Edit
andConcatenate
VI
is
availablefor
scaling
the
second
data
segmentby
the
ratioA2
where
X\
andXi
arethe
means ofthe
data
segments respectively.The
resultofthe
scaling
is
to
'normalize'the
seconddata
to the
first data
set.The
resultsofconcatenating
andscaling
the
same portion ofthe
velocity
data
in
Figure 2-5
areConcatenated
Velocity
Data
withScaling
250-r Concatenation Point
? Scaled Portion
-I 1 1
3 4
Time
(sec.)
Figure
2-6.
Concatenated velocity
data from Figure 2-5
with subsequentscaling.Care in data
removal.There
are some possible sources ofmisinterpretation when
editing
spuriousdata from larger data
sets andconcatenating
the
remaining
segments.These
pitfalls canbe
avoidedby
carefully
controlling
the
input
parametersto
Edit
andConcatenate.
vi.Specific instructions
and examples
for
using
this
algorithm aredetailed
in Appendix
C. Care
mustbe
exercised when
editing
data
from
the
center of atime
domain
series.Depending
on
how
muchdata
is
removed, the
frequency
domain
representations ofboth
the
velocity
data
andthe
derived
variability
data
may
be
altered significantly.The
biggest
impact
onthe
frequency
domain
representation of aconcatenatedtime
series
is
in
the
loss
offrequency
resolutiondue
to
shortening
the
data
recordlength. As
a generalguideline, there
shouldbe
an adequate number ofdata
frequency
domain.
In
addition,
it
shouldbe
notedthat
removing
data from
the
center of a series will
interrupt
the
continuity
(if
any)
in
the
rhythmic nature ofthe
variationsin
peakvelocity
andheart
rate.If
data is
to
be
removedonly
from
the
beginning
or end of adata
series, then the
only
concernis
having
anadequate amount of
data
afterwardfor
the
desired
spectral resolution.In
Appendix
B,
the
frequency
domain
effects of'sectioning'
sinusoidal
data
setsarediscussed
in
moredetail.
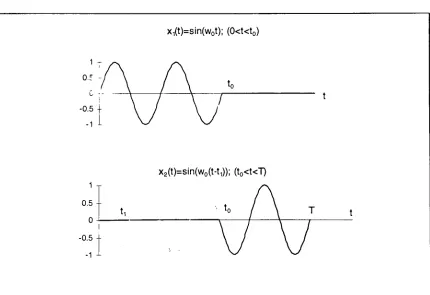
2.4
Grouping
Embryo/Fetal Spectra
The
primary step
in
post-processing
cardiovascular spectrafor
comparison purposes
is
to
'group'the
embryonic/fetal
spectrain
terms
ofage,
treatment protocols,
etc.Representative
group
averages canthen
be
obtainedfor
further processing
and comparison.Because
of variable recordlengths
ofthe
original
blood
velocity
data,
the
spectral representations ofindividual
embryonic
/fetal data
sets willhave different
frequency
axis resolutions.Additionally,
the
'effective
sampling
rates'of
the
time
domain
data
for
various
fetuses may
differ3. The
result willbe
differing
frequency
rangesin
the
individual
embryo spectra.Mathematically,
this is
shownby
Af
=(2.3)
N
3
The 'effective sampling
rate'is
equalto thenumber samplesdivided
by
the timeserieslength (in
where
fs
is
the
effectivesampling
rate andN is
the
numberof pointsin
the
data
series.
The
frequency
range ofthe
spectrum willbe
half
ofthe
effectivesampling
rate4.
An
exampleshowing
the
effects ofdiffering
data
setlengths
and effectivesampling
rates are shownin Table 2-1.
Fetus
Time Series
Length
Number
ofData
Points
Effective
Sampling
Rate
Nyquist
Frequency
Af
004AUA
13.86
sec.512
36.94 Hz
18.47
Hz
.072Hz
004BUA
12.00
sec.512
42.67
Hz
21.33 Hz
.083Hz
022AUMA
27.00
sec.1024
37.93
Hz
18.96 Hz
.037Hz
Table 2-1.
Example
offrequency
axis resolutiondifferences for
agroup
ofhuman
fetuses.
Table 2-1
showsthe
differences
among
agroup
ofthree
11
week(gestational
age)
human
fetuses
in
terms
oftime
series recordlength
in
secondsand
discrete
points.If
aPSD
(either
heart
rate or peakblood
velocity
variability)
for
eachfetus
were plottedseparately, the
frequency
axes wouldhave
different
ranges and
increments.
To
compare eachPSD
across specificfrequencies,
aninterpolated
point wouldhave
to
be
determined
for
some ofthe
PSDs,
asthey
may
nothave discrete
points atthat
exactfrequency.
Likewise,
if it
wasdesired
to
determine
a'group
average'
PSD
for
the
threefetuses,
a'common
frequency
axis'
would
have
to
be determined
for
the
spectra.It is
necessary,
therefore,
to
interpolate
theindividual
embryo spectrato
a commonfrequency
axisbefore
any
spectralaveraging
and comparison can occur.The
forgoing
exampleis
4
In
accordance withtheNyquist
Sampling
Theorem,
a conditionby
whichthehighest
resolvabletypical
whendetermining
agroup
averagespectrumfor
severalfetuses
orembryos.
The
frequency
axisdetermination,
spectralinterpolation,
andgroup
averaging
are all performedin
the
custom algorithmLinear
interpolation Il.vi.
Common
frequency
axes.As
withany
graphicalrepresentation,
it is
desirable
to
maximizethe
resolution ofthe
data
being
presented.Using
the
scenario
in
table
2-1,
the
spectral representationfor
the
data from fetus
'022AUMA'
has
the
finest
resolutionalong
the
frequency
axis(lowest
resolvablefrequency).
Therefore,
allthree
fetal
spectra shouldbe
interpolated
to
a commonfrequency
axisthat
has
increments in
A/
equalto
orless
than
.037Hz
From
previousobservations,
heart
rate andblood
velocity variability
predominantly
occursin
the
sub2 Hz
range5.To
maximizethe
amount ofspectral
information
to
be displayed (and later
analyzed) it
is
sometimesrequired
to
'limit'the
frequency
range ofthe
spectra.The
custom algorithmLinear interpolation Il.vi derives
a commonfrequency
axisfrom
eitherthe
individual
spectralinformation
orfrom
a user-specifiedfrequency
resolution.If
the
frequency
axes ofindividual
spectra are normalized(i.e.,
divided
through
by
the
meanheart
ratedetermined
from
thevelocity
time series), then the
algorithmLinear
interpolation
Il.vi appropriately
interpolates
eitherto
the
finest
normalizedfrequency
resolution,
or a user-specified resolution.5
Spectral
interpolation.
After
a commonfrequency
axishas
been
determined,
the
individual
spectra arethen
interpolated to
it.
Linear
interpolation is
used.Linear interpolation is essentially
the
linear
determination
of a value
between
two
data
points.Mathematically
it
is
representedby
Vin
=y3
+m(Ax)
(2.4)
where
yt is
the
first
known
data
point,
mis
the
slope of aline connecting
the two
known
adjacentdata
points,
andAx
is
the
distance along
the
xaxisbetween
the
interpolated
point and thefirst
reference point.For
interpolating
PSDs,
yml is
the
PSD
valueto
be
interpolated,
mis
the
slope ofthe
line connecting
the two
known
adjacent
PSD
points,
andAx
is
the
frequency
axisincrement (A/).
Spectral
averaging.After
the
individual
spectrahave been
interpolated
to
a common
frequency
axis,
a'common'
spectrum
is
generatedby
averaging
the
individual
spectraacross eachdiscrete frequency. Figure
2-7
shows atypical
Week 18 Peak Amplitude
Variability
PSD
First Embryo
Second Embryo
1 1.5 2 2.5
Frequency (Hz.)
Figure
2-7.
Typical
result ofinterpolating
andaveraging
two
embryo spectra.Linear Interpolation
Il.vi
can writethe
individual interpolated
and'composite'
averaged
PSD
spectrato
a spreadsheet-readable 'text'file
for further
analysis.
2.5
Band
Power Determination
The
'grouped'PSD
representation of embryonic cardiovascularvariability
data
provides away
to
assessfluctuations
in
cardiovascular outputbased
ongestationalage.
However,
because
ofthe
non-stationary
variations ofembryonic
/fetal
cardiovascularoutput, the
PSDs
from
some subjectsmay have
spectral content spread over a
limited
frequency
range.In addition,
slightdifferences in
physiology
and cardiovascularactivity among
agroup
ofembryos
/fetuses
may
resultin
slightly different dominant frequencies
in
thegrouping
is
controlledby
gestational age andtreatment.
Therefore,
the
PSD
alone cannot
adequately
provide adirect
comparisonof representativeembryo/fetal groups
by
age andtreatment
protocols.One
way
to
comparethe
spectral
activity
of variousembryo/
fetal
groupsis
to
determine
the
amountof 'power'in
variousfrequency
bands
ofthe
spectrum.An
advantage ofthe
PSD
form
of afrequency
spectrumis
that
it
caneasily be
usedto
calculatethe
powerin
variousfrequency
bands.
The PSD is
afunction
that
showsthe
distribution
of spectral content of adata
series.It is
similarin
natureto
a populationdensity
map,
wherethe
population per square mile of geographic area
is
represented.Once
givena populationdensity
map, the total
populationof a certain region canbe found
by
integrating
the
populationdensity
data
overthe
region's geographical area.Likewise,
the
amountof'power'
in
afrequency
band
in
aPSD
canbe
found
by
integrating
the
PSD data
along
the
frequency
axisin
the
specifiedfrequency
band limits. Both
populationdensity
anddiscrete
PSDs
comefrom
observed orsampled
data.
In
this case,
numericalintegration
techniques
canbe
used.The
numerical
integration
for band
poweris
performedin
the
custom algorithmsBand
Power
Calculation (5
Bands).vi
andBand Power
Calculation (10 Bands).vi.
Since linear
interpolation
(a
first
orderpolynomial)
is
usedto
connectdata
points
in
the
frequency
spectra, the trapezoidal
ruleof numericalintegration
will1989). Refer
to
Appendix
A
for
mathematicaldetails
oftrapezoidal
ruleintegration
ofPSD
data. Because
the
trapezoidal
ruleis
the
lowest
order(two
point)
numericalintegration
method,
it is
computationally
efficientandis
usedin
the
band
power calculation algorithms.Frequency
bands.
The
custom algorithmsBand Power Calculation (5
Bands).vi
andBand Power
Calculation
(10 Bands).vi
calculatethe
spectral powerin
each of
5
or10
user-selectablefrequency
bands
respectively.The
frequency
bands have been arbitrarily
setin
0.5 Hz increments
and0.25
Hz
increments for
the two
algorithmsby
default.
The
clinically
important
frequency
for
embryo
/fetal heart
rate and peakvelocity variability
data
series are yetto
be
defined,
as no suchdata
appearin
the
openliterature.
An
alternativeway
to
display
spectral powerdistribution is
to
use a'band
power ratio'.
Essentially
the
band
power ratiois
the
spectralband
power contentnormalized
by
the
powerin
the
entire spectrum.The
band
power ratiois
expressed as a percentage of
the
total
spectral power.This
computationis
alsoperformed
by
both band
power algorithms above.Band
powerof normalizedPSDs. Recall
that the
normalizedPSDs
have
units of
1/Hz,
withfrequency
axesthat
are expressed relativeto the
baseline
heart
rate.The
integration
ofthese
spectraalong
the
unitlessfrequency
axis(as
in
the
band
powercalculations)
will produceband
'power'values with units ofto
providea spectralpowerdistribution,
relativeto
the
power presentin
the
time
average
heart
rate(for heart
ratevariability
PSDs)
orthe time
average meanvelocity
(for
peakvelocity variability
PSDs).
Alternative
spectralgrouping
method.The
calculation ofband
powerfor
a
PSD
allows analternativeway
to
group
the
spectraldata from
different
embryos
/fetuses according
to
gestationalage,
treatment
protocols,
etc.Recall
from Section
2.4
that
individual PSDs
from
different
embryos/fetuses
canbe
grouped
by
averaging
linearly
interpolated
spectraalong
a commonfrequency
axis.
Using
the
band
powercalculations,
grouping
ofindividual
PSDs
canbe
performed as well.
Using
the
samefrequency
bands
eachtime,
the
band
powercan
be separately
calculatedfor individual
embryos/fetuses
in
the
group.The
individual
band
power calculationsfor
the
embryos/fetuses
canthen
be
averaged across each
frequency
band
to
obtain agroup
averageband
powerdistribution.
Spectral
leakage
effects onband
power.Care
mustbe
exercised wheninterpreting
spectralband
power results.Due
to
spectralleakage6,
spectralcontent associated with a particular
frequency
band
may be
present outside ofthe
frequency
band. In
otherwords,
the powercalculatedfor
a certainfrequency
band
may
include
spectralpowerfrom
adjacentfrequency
bands. This
point6
Spectral leakage is
thetendency
for
frequency
contentto'spread
out'overa
frequency
rangeillustrates
the needfor
anadequaterecordlength
whenperforming
any
discrete
spectral analysis.
2.6
Joint
Time-Frequency
Analysis
(JTFA)
Variations
in
fetal
cardiovascularoutputmay be
non-stationary.In
otherwords, the
componentfrequencies
offetal heart
rate andblood velocity
variability
canchangewithtime.
These
hemodynamic
changes arelikely
due
to
growth,
morphogenisis anddevelopment,
and occur as variationsin heart
rateand mean
velocity
about reference set pointsfor
gestational age.(This
is
anaspect
to
be
studiedthrough
future
application ofthe
algorithms presentedin
this
thesis.)
The
frequency
variationsin
fetal
cardiac output are akinto
pitch changesin
music and structuresundergoing
variablefrequency
vibrations.The PSD
spectradiscussed
thus
far
showheart
rate andblood velocity
variability
spectralpowerdistribution for the
entiredata
recordtaken
in
toto.
However,
the
spectrado
not show when orif
spectral changes occur.Such
spectral changes with
time
canbe
illustrated
using
the
'chirp' signal.The
discrete
form
ofthe signal,
with261
samplestaken
at asampling
rate of849
Hz,
is
shownin
Figure 2-8.
By
inspection
ofFigure
2-8,
the
frequency
ofthe
signalstarts at-1 X
0.35
Time
(sec.)
Figure 2-8.
'Chirp'data
usedfor JTFA
example.Note
frequency
changein
time.
A PSD
ofthe
'chirp'time
series(Figure
2-9)
showsthe
spectraldensities
ofthis
signal overthe
entire sampledtime
period.The PSD
alonedoes
not showwhen or
how
thefrequencies
are changing.However,
the
PSD
does
show aspread of
frequency
componentsbetween 20
and200 Hz Neither
the time
seriesnor
the
PSD
showhow
the
frequency
increase
takes
place.In
fact,
looking
atthe
PSD
alone would maskwhetherthe
seriesis
changing
at all.This
situation canbe
addressed
using
joint
time-frequency
analysis(JTFA).
Using
JTFA,
the
spectralcontent of
the
'chirp'time
series canbe
viewedin
both
the
frequency
andthe
Discrete PSD
ofChirp
Data
Q to Q.
H i
r-100 200 300
Frequency
(Hz.)
400 500
Figure
2-9.
Discrete PSD
ofthe
'chirp'signalfrom
Figure 2-8.
Figure
2-10
showsthe time-dependent
power spectraldensities,
orspectrogram,
of the'chirp'
signal
in
threedimensions. The Cartesian
x- andy-axes are
frequency
andtime,
respectively.The
z-axisis
the
PSD
value.It
canbe
seen
from
the
plotthat the
dominant
frequency
increases
linearly
withtime.
With
fetal
cardiovascularsignals,
JTFA
canbe
usedin
a similarfashion
to
show
how
cardiac outputvariability
is
mediated.In
addition,
the mediationin
output
variability
canbe
examinedfor different
stages of gestationalspectrogram
of
Chirp
signal
600
200
freq.
(Hz.)
Figure 2-10.
Three
dimensional
time-frequency
analysis representation of a 'chirp'signal.
Note
the
linear increase
in
the
dominant
frequency
withtime.
The
shorttime
Fourier
transform
(STFT). The
forgoing
example wasgenerated
using
the
short-timeFourier
transform
(STFT). The STFT is
one ofthe
most
commonly
used methodsfor JTFA
andis
adoptedfor
this
work.Other
methods
include
those
developed
by
Gabor, Wigner,
Ville
and others(Cohen
1989).
In
the
broadest
sense,
many
JTFA
algorithms arebased
onthe
calculat