Market
access,
population
density,
and
socioeconomic
development
explain
diversity
and
functional
group
biomass
of
coral
reef
fish
assemblages
Tom
D.
Brewer
a,*
,
Joshua
E.
Cinner
a,
Rebecca
Fisher
b,
Alison
Green
c,
Shaun
K.
Wilson
d,eaAustralianResearchCouncilCentreofExcellenceforCoralReefStudies,JamesCookUniversity,Townsville,QLD4811,Australia
b
AustralianInstituteofMarineScience,UWAOceansInstitute(M004),35StirlingHwy,Crawley6009,Australia
c
TheNatureConservancy,51EdmonstoneStreet,Brisbane4101,Australia
d
MarineScienceProgram,DepartmentofEnvironmentandConservation,17DickPerryAve,Kensington6151,Australia
e
OceansInstitute,UniversityofWesternAustralia,Crawley6009,Australia
1. Introduction
There is overwhelming evidence that human activities are profoundly altering marine ecosystems on a global scale (e.g.
Hughes,1994;Pandolfietal.,2003).Ofparticularconcernarethe poorly understood, yet potentially disastrous environmental changesthathumanactivityiscausingtothefunctioningofcoral reefecosystemsuponwhichmillionsofpeopledepend(Moraetal., 2011). Ecosystem function is conceptually and analytically complex, requiring a diverse array of metrics to understand ecosystemresponsetohumanactivity.Highbiologicaldiversityis thoughttocontribute tomaintainingecosystem resilience (e.g.
McCann,2000;Cardinaleetal.,2006;Tilmanetal.,2006)through increased response diversity to perturbations and functional redundancy (Naeem, 1998; Chapin et al., 2000; Hooper et al., 2005; Maestre et al., 2012; but see Ives and Carpenter, 2007), assumingthat species respond to threats uniquely. The useof diversitymetricsassurrogatesforecosystemfunctionhowever, does not come without criticism. There is, for example, some evidencethatparticularspecies(Bellwoodetal.,2003,2006;Hoey
and Bellwood, 2009), and functionalgroups (e.g.Hughes et al., 2007) perform disproportionately important functional roles, actingasenergyconduitsthroughtrophiclevelsandmaintaining broaderecosystemprocesses.Therefore,itisimportanttoconsider measuresofbothdiversityandfunctionalgroupstounderstand howecosystemsmayrespondtohumanactivities.
Coral reef fish are perhaps the most diverse of vertebrate communities, are vital to ecosystem function, and provide significantgoodsandservicestopeople.Arangeoffactorshave beenidentifiedasimportantdriversofthediversity,biomass,and abundanceofreeffishfunctionalgroups.Atlargebiogeographic scales,distributionsofdiversityandfunctioncanbeexplainedby environmental factors, including available habitat, latitude– longitudegradients,themid-domaineffect,gyreinfluence,history of environmental stress, and larval subsidy from species-rich regions(BellwoodandHughes,2001;Connollyetal.,2003;Mora et al., 2003; Bellwood et al.,2005; Mora and Robertson, 2005; McClanahan et al., 2011). At local and national socio-political scales,variousenvironmentalandsocialfactorshavebeenusedto explainfishdiversityandbiomassoffunctionalgroups. Environ-mental factors include depth, exposure to prevailing weather, season,reefzone,coralcover,substraterugosity,habitat complex-ity,andlarvaldispersal(LuckhurstandLuckhurst,1978;Molles, 1978; Bell and Galzin, 1984; Roberts and Ormond, 1987; ARTICLE INFO
Articlehistory:
Received23September2011
Receivedinrevisedform24January2012 Accepted26January2012
Availableonline23February2012
Keywords:
Distaldriver Proximatedriver Fishingpressure Coralcover
Structuralequationmodel Trade
ABSTRACT
Thereisoverwhelmingevidencethatmanylocal-scalehumanactivities(e.g.fishing)haveadeleterious effect on coral reef fish assemblages. Our understanding of how broad social phenomena (e.g. socioeconomicdevelopment)affectthediversityandfunctionofcoralreeffishassemblageshowever,is still poor. Here, we use structural equation models to reveal how human population density, socioeconomicdevelopment,andmarketaccessaffectfishingpressureandcoralcoverto,inturn,explain thediversityandbiomassofkeyfunctionalgroupsofreeffishassemblageswithinSolomonIslands. Fishingpressureispredominantlydrivenbybothmarketaccessandlocalpopulationdensity,andhasa clearnegativeeffectonthediversityandfunctionofcoralreeffishes.Thestrongpositiveeffectofmarket accessonfishingpressuremakescleartheimportanceofunderstandingsocial-ecologicallinkagesinthe contextof increasinglyconnected societies.This studyhighlightstheneed toaddressbroad social phenomenaratherthanfocusingonproximatethreatssuchasfishingpressure,toensurethecontinued flowofcoralreefgoodsandservicesinthistimeofrapidglobalsocialandenvironmentalchange.
ß 2012ElsevierLtd.Allrightsreserved.
*Correspondingauthor.Tel.:+61433976561.
E-mailaddress:[email protected](T.D.Brewer).
ContentslistsavailableatSciVerseScienceDirect
Global
Environmental
Change
j ou rna l hom e pa ge : w w w. e l s e v i e r. c om/ l o ca t e / gl oe n v cha
FriedlanderandParrish,1998;Friedlanderetal.,2003;Gratwicke andSpeight,2005;Jonesetal.,2005;Grahametal.,2006)(Table1). Incontrasttothedepthofworkassessingenvironmentaldriversof fish diversity and function, assessments of the potentially important role that human activity might have in shaping ecologicalassemblageshavefocusedlargelyonhumanpopulation density(Jenningsetal.,1995;JenningsandPolunin,1996,1997; Bellwoodetal.,2003;Dulvyetal.,2004a,b;Mora,2008;Williams etal.,2008,2011;Stallings,2009)andfishingpressure(DeMartini etal., 2008;Wilsonetal., 2008). However, recent researchhas highlighted the potentially important role of factors such as market access and socioeconomic development in explaining functionalgroupdistributions(AswaniandSabetian,2009;Brewer etal.,2009;Cinneretal.,2009a;Stallings,2009).Whatisnotclear, however,ishowmarketaccessandsocioeconomicdevelopment affect fish species diversity, and whether market access and socioeconomicdevelopmenthaveaneffecton fishdiversityand functionbeyondwhatisexplainedbyhumanpopulationdensity. Thispaperaimstocontributetothisresearchgapbyexamining relationshipsbetweensocialdriversandthediversityandfunction ofreeffishcommunities.
Socialscientistsworkingonhuman–environmentinteractions oftendifferentiatebetweenproximate(e.g.fishingpressure)and distal(e.g.marketaccess,socioeconomicdevelopment,andhuman populationdensity)driversofenvironmentaldegradation( Forest-er and Machlis, 1996; Agrawal and Yadama, 1997; Geist and Lambin,2002;Krameretal.,2009;McKinneyetal.,2010).Inacoral reefcontext,thereisclearevidencethat,atthelocal-scale,people directly affect coral reef fish diversity and function through proximatedriverssuchasfishingpressureandhabitatdegradation (Wilsonetal.,2008).Whatisnotclear,however,istheroleofdistal drivers,inshapingtheseproximatedriversandultimatelycoral reeffishdiversityandfunction.Forexample,increased socioeco-nomic development does not directly affect fish diversity and function,butmightintensifylocalfishingpressurethroughgreater
accesstomoreefficientfishinggear,whichmight,inturn,decrease diversityand functionofcoralreeffish.Alternatively,increased socioeconomic development might reducedependence on local resources, or enable improved resource management practices, resulting inincreasedfish diversity andfunction (Cinneret al., 2009a).
Here,weexplorethelinkagesbetweenthreerecognizeddistal drivers (population density, socioeconomic development and marketaccess),twoproximatedrivers(fishingpressureandcoral cover),andarangeofmetricsoffishdiversityandfunction.Weuse structuralequationmodelstounderstandthesequentialeffectsof distal drivers on proximate drivers, and proximate drivers on diversityandfunctionmetricsacross25sitesinSolomonIslands. SolomonIslandsisalow-incomenationwithexceptionallyhigh fishspeciesdiversity(Greenetal.,2006a).Thevastmajorityofthe rapidlygrowingpopulationlivesalongthecoastlinethroughout themultitudeofislandswhichcomprisethenation.Reeffishand fishingaresignificanttoSolomonIslandslivelihoods,yetfuture demandforfishisexpectedtoexceedecologicalreplenishment (Bell et al., 2009). Increasing market access, and evidence of declining fish biomass withincreasing market proximity (ANU Enterprise, 2008), particularly for certain taxonomic groups (Breweretal.,2009),suggestthatfishingpressure,andassociated social and economic change, contribute to variability in the distributionofreeffishdiversityandfunction.Byassessingand comparingtheeffectofdistalandproximatedriversonreeffish assemblagesthisstudyquantifiestherelativeimportanceofthese socialdriversinadiverseecologicalsystem.
2. Methods
2.1. Siteselectionanddelineation
The25sitesinthisstudyspansevenofthenineprovincesin SolomonIslands(Fig.1).Wecollectedbothecologicalandsocial
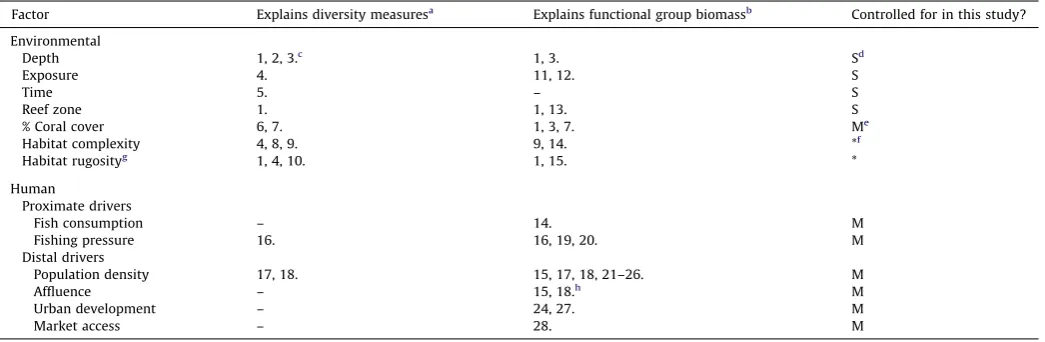
Table1
Keyenvironmentalandhumanfactorsthatexplaininsitucoralreeffishdiversityandfunctionalgroupdistributionsatbiogeographicscalesrelevanttothisstudy.
Factor Explainsdiversitymeasuresa
Explainsfunctionalgroupbiomassb
Controlledforinthisstudy?
Environmental
Depth 1,2,3.c
1,3. Sd
Exposure 4. 11,12. S
Time 5. – S
Reefzone 1. 1,13. S
%Coralcover 6,7. 1,3,7. Me
Habitatcomplexity 4,8,9. 9,14. *f
Habitatrugosityg
1,4,10. 1,15. *
Human
Proximatedrivers
Fishconsumption – 14. M
Fishingpressure 16. 16,19,20. M
Distaldrivers
Populationdensity 17,18. 15,17,18,21–26. M
Affluence – 15,18.h
M
Urbandevelopment – 24,27. M
Marketaccess – 28. M
aReferencesrelatetoanymeasureofdiversity(e.g.richness,evenness).
bReferencesrelatetoanyfunctionalgroupabundanceorbiomassmeasure(e.g.herbivorebiomass) c
Supportingreferences:1.FriedlanderandParrish(1998),(2).RobertsandOrmond(1987),(3).O¨ hmanandRajasuriya(1998),(4).Friedlanderetal.(2003),(5).Molles (1978),6.BellandGalzin(1984),(7).Wilsonetal.(2006),(8).GratwickeandSpeight(2005),(9).Grahametal.(2006),(10).LuckhurstandLuckhurst(1978),(11).Fultonand Bellwood(2005),(12).Floeteretal.(2007),(13).Russ(2003),(14).Wilsonetal.(2008),(15).Cinneretal.(2009a),(16).Jenningsetal.(1995),(17).JenningsandPolunin (1997),(18).Stallings(2009),(19).DeMartinietal.(2008),(20).JenningsandPolunin(1996),(21).Bellwoodetal.(2003),(22).Dulvyetal.(2004a),(23).Dulvyetal.(2004b), (24).Mora(2008),(25).Williamsetal.(2011),(26).Williamsetal.(2008),(27).AswaniandSabetian(2009),(28).Breweretal.(2009).Referencesincludeonlythosethat show,statistically,theeffectofthefactorslisted,onmeasuresofcoralreeffishdiversityorfunction.
dControlledduringsampling. e
Controlledinmodel. f
Correlatedwithcoralcover(Grahametal.,2008). g
Alsomeasuredasnumberof‘holes’andholevolumeofreefsubstrate(FriedlanderandParrish,1998). h
[image:2.595.34.557.461.632.2]dataforeachsite.Sitelocationswerepredeterminedbyamarine assessmentwhichsurveyedbothreeffishpopulationsandcoral substrate (Green et al., 2006a). Itwas necessary to define the spatialextentofeachsitetodeterminewhichhumanpopulations were associated with the coral reef at the location where the marineassessmentoccurred.Thespatialextentofeachsitewas elicitedfromindividualspossessinglocalknowledgeofresource usebypeopleresidinginadjacentvillages.Themarineboundary wasdefinedasthearea,withinthevicinityofwherethefishwere surveyed,which was likely tobe exploitedby people living in villagesontheadjacentcoastline(Breweretal.,2009).Alternative methodsexistforestimatingresourceuseboundaries,including frictionmappingusingthiessenpolygons(MullerandZeller,2002), ethnographic studies (Aswani, 1999), and participatory GIS mapping(Aswani,2011).However,thelarge-scalenatureofthis study inhibited the use of these more localized resource use mappingtechniques.Ourapproachmadeuseofthebestavailable information at the spatial scale of our study. The terrestrial boundaryofeachsitewassetat1kmfromthecoastline,extending alongthecoasttowherethemarineareaboundaryintersectedthe coast.Socialdataon villageswithintheaforementioned bound-aries wasprovidedby theSolomon Islandsstatisticsoffice and derived from a national census (1999), and a national village resourcesurvey(2007/2008).
2.2. Ecologicalresponsevariables
Fish species used in this study includedall fishessurveyed acrossthe25sites(ESM1).Fishweresurveyed,fromMaytoJune 2004,usingunderwatervisualcensusalongfive50mtransectsat eachsite,atadepthof8–10monreefslopeinlocationsexposedto prevailingweatherconditions(Greenetal.,2006b).Fourmetricsof speciesdiversity wereusedto testthe effectofproximate and distaldriversonfish diversity;(1) speciesrichness, (2) Pielou’s speciesevenness,(3)averagetaxonomicdistinctness(AvTD),and (4)variationintaxonomicdistinctness(VarTD).Speciesevenness warrantsinvestigationasitconsiders therelative abundanceof species and can have important ecosystem ramifications well beforespeciesbecomelocallyextinct(Chapinetal.,2000).AvTDis ameasureoftheaveragedistancebetweenallpairsofspeciesina taxonomic tree, which captures phenotypic differences and functional richness (Clarke and Warwick, 1999; Rogers et al., 1999).VarTDisthevarianceofthepathlengthsbetweeneverypair ofspeciesinataxonomictree,andrepresentstheunevennessof the taxonomic tree (Clarke and Warwick, 2001). Taxonomic hierarchylevels used tomeasure AvTD and VarTDwere Class,
Order,Family,GenusandSpecies.Pathlengthsbetweentaxonomic levelswereequallyweighted.
Totalbiomassestimateswerederivedfortwo keyfunctional groups: piscivores and herbivores.Piscivores wereclassified as fishesthat,basedongutcontentanalyses(FroeseandPauly,2011) predominantlyconsumefishes.Piscivorescaninhibitincreasein abundanceoflowertrophiclevelspeciesthroughpredatorprey interaction(Jenningsetal.,1995; Grahametal.,2003),and are particularlysensitivetofishingpressure(e.g.JenningsandPolunin, 1997;DeMartinietal.,2008;Sandinetal.,2008).Herbivoreswere classifiedasthosespeciesthatpredominatelyfeedonlargefleshy algae orthe epilithicalgal matrix (censu Wilsonand Bellwood, 1997).Thisincludesfishthatremovepartofthereefbyscrapingor excavating the substratum, and grazers that mainly ingest filamentous algae (censu Choat et al., 2002). Herbivores are thought to play a critical role in the resilience of coral reef ecosystems by preventing algal overgrowth that can smother corals(Mumby,2006;Hughesetal.,2007;GreenandBellwood, 2009).Piscivoreandherbivorespeciesweredividedintotargetand non-targetspeciestofurtherexploretheeffectoffishingonthese functional groups.Thelistoftargetand non-targetspecieswas constructedusingexpertopinionofSolomonIslandstargetspecies (Greenetal.,2006b),andcreelsurveydatafromadjacentPapua NewGuinea(Cinneretal.,2009b)(ESM1).
2.3. Proximatedrivers
Wemeasured twoproximatedriverspreviouslyshowntobe related to fish diversity and function: (1) coral cover, and (2) fishing pressure. Coral cover was measured along the same transectsusedinthefishsurvey.Substratetypewasclassifiedas coralornon-coralatthreepointsat2mintervalsalongeach50m transect totalling 375points ateach site (Hughes,2006). Coral cover,asapercentageoftotalsubstrate,rangedfrom13to68% (mean=44%)amongsites.Wemeasuredfishingpressureasthe number of people selling fishand numbers of fiberglassboats, eskies(insulatediceboxesusedforfishpreservation),spearguns and fishing nets per coral reef area at each site. We included numberofpeoplesellingfishasavariablebecauseofevidencethat themarket-basedfisheryadverselyaffectscoralreeffishstocksin SolomonIslands(AswaniandSabetian,2009;Breweretal.,2009). Coral reef area wasmeasured usingGoogle Earth Professional, followingthemethodsdiscussedinBreweretal.(2009).Fishing pressurevariableswereconsolidatedtoasinglevariable(
l
=3.3; varianceexplained=65.4%)usingPrincipalComponentsAnalysis. Allfishing pressurevariables loaded positivelyon theprincipal componentat0.65.2.4. Distaldrivers
We developed metrics for three key distal drivers that we hypothesizedcouldstronglyinfluencereeffishassemblages:(1) humanpopulationdensity,(2) socioeconomicdevelopment,and (3)marketaccess.Humanpopulationdensitywasmeasuredasthe totalnumberofinhabitantspercoralreefareaateachsite.Human populationdensityrangedfrom11to1035(mean=159)people perkm2ofcoralreef.Basedoncalculatedpopulationdensitiesand conservative per capitafish consumption estimates (Bell et al., 2009),eightofthe25sitesexceededestimatedsustainableharvest levelsof5metrictonneskm2yr1
(Newtonetal.,2007). Socioeco-nomic development was measured as the sumof a set of un-weightedinfrastructuresandamenitiessimilartothoseusedin previousstudies(Cinneretal.,2009a;Pollnacetal.,2010); pre-school, kindergarten,primary school,highschool,clinic,wharf, tradestore,supermarket,postalservice,fueldepot,creditfacility andairportateachsite.Theinfrastructureandamenityvariables
[image:3.595.47.289.53.216.2]displayedhighinternalconsistency(Cronbach’s
a
=0.823) allow-ingaggregationofvariablesintoasinglemeasureofsocioeconomic development.Theaggregatescoreforeachsitewasthendividedby the number of villages at each respective site, to control for infrastructureandamenityaccessibility(CinnerandMcClanahan, 2006).Marketaccesswasmeasuredastheun-weightedsumofthe shortestdistancefromeach ecologicalsamplinglocation tothe nearestlocalfishmarket,provincialcapital,andnationalcapital,all ofwhichhavefishmarkets,usingroadsandseaaspossibleroutes withinthesamedistancemeasure(Breweretal.,2009).2.5. Modelconstruction
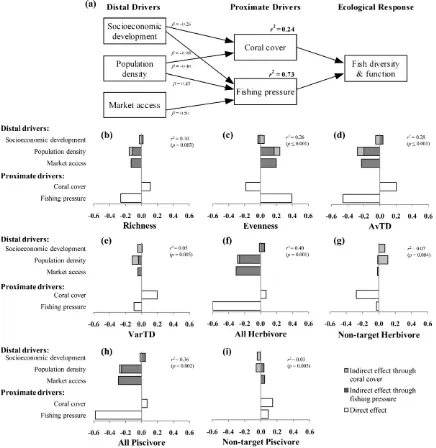
Weusedpartialleastsquaresregression,intheprogramWarp PLS,tobuild structural equationmodels (SEMs)of the general form: distal drivers!proximate drivers!ecological response.
Partialleastsquareswaschosenovercovariance-basedapproaches primarily because it suited our small sample size (Chin and Newstead,1999;Reinartzetal.,2009).Thedistalandproximate driversremainedconsistentacrossmodelswithonlytheecological response changing between models (Fig. 2a). All distal drivers (market access, socioeconomic development and population density)werelinkedtothetwoproximatedrivers(fishingpressure andcoralcover),exceptmarketaccesstocoralcoverastherewas no theoreticaljustification for this link.Bothproximate drivers werelinkedtotheecologicalresponsevariableinallmodels.This resultedinuniquemodelsfor eachecologicalresponse; species richness,Pielou’sevenness,AvTD,VarTD,totalherbivorebiomass, non-targetherbivorebiomass,totalpiscivorebiomass,and non-targetpiscivorebiomass.Thepartialleastsquaresmethodpartials out each analysis (e.g. the effect of population density and socioeconomic development on coral cover) from the overall
[image:4.595.77.513.237.685.2]modelandistherefore,inthisstudy,theequivalenttosetsof non-linearregressions,exceptthatoverallmodelfitstatisticsarealso generated.Themodelswerebootstrapped,setat100 iterations. Weconstrainedallmodelstosecond-orderpolynomial relation-ships,therebyallowingsimple,non-linearrelationshipsbetween variables.WarpPLSsoftwarehasaninbuiltfunctionwherebythe relationshipbetweentwovariableswilldefaulttoasmallerorder polynomial if it is deemed linear (Kock, 2010). The output generatedincludedindividualstandardizedpathcoefficients(
b
), partialmodel fit scores (r2), overall model p values calculatedthrough resampling estimations coupled with Bonferroni-like corrections(Kock,2010),andindividualexplanatoryandresponse
x,yplots(ESM2).Thetotaleffectofdistaldrivers(marketaccess, socioeconomicdevelopmentandpopulationdensity)were calcu-latedbymultiplyingthestandardizedcoefficients(
b
)withineach pathwaythensummingthesevaluesforpathwaysassociatedwith eachdriver.3. Results
3.1. Effectsofproximatedriversonfishfunctionanddiversity
Fishing pressure had a clear negative effect on both fish diversity and the biomass of key functional groups of fish. Specifically, fishing pressure correlated negatively withspecies richness,andAvTD,andpositivelywithspeciesevenness(Fig.2b– d).Fishingpressurehowever,didnotnoticeablyaffectVarTD,with onlyasmalldecreaseinVarTDassociatedwithincreasedfishing pressure(Fig.2e).Also,fishingpressurehadaclearnegativeeffect on both allpiscivore and allherbivore biomass, yetnon-target biomassofthetwofunctionalgroupswasnegligiblyaffectedby fishingpressure (Fig. 2f–i).Coral covergenerally had a smaller effect on diversity and functional group metrics than fishing pressure(Fig.2b–i);Coralcoverwaspositivelyrelatedtorichness, AvTD,VarTDandnon-targetpiscivores.Coralcoverwasstrongly negatively correlated with species evenness and all herbivore biomass(Fig.2c,g).
3.2. Effectsofdistaldriversonfishfunctionanddiversity
Distal drivers explained much of the variance of fishing pressure (r2=0.73), particularly market access and population density(Fig.2a).Populationdensityandsocioeconomic develop-mentwere,however,comparativelypoordescriptorsofcoralcover (r2=0.24).Socioeconomicdevelopmenthadaweaknegativeeffect
onfishingpressureandoncoralcover,thushadbothpositiveand negative effects on fish diversity and function, except for all herbivorebiomasswhichwaspositivelyaffectedbysocioeconomic development through both decreased fishing pressure and decreased coralcover. Population densityhad a negative effect onallherbivorebiomassthroughincreasedfishingpressure,anda positive, but weaker, effect on all herbivore biomass through decreased coral cover. Increased market access and population density, more than socioeconomic development, explained de-creaseddiversity and functionofcoral reeffish(Fig.2b–i).The strongindirecteffectofmarketaccess,ondiversityandfunction, was particularly noteworthy because the model specified that market access indirectly affected function and diversity only throughfishingpressure,ratherthanthroughbothfishingpressure andcoralcover(Fig.2a).
4. Discussion
This study explored how habitat and social factors explain spatialvariabilityinthediversityandfunctionalgroupbiomassof coralreef fishesat25 sitesacrossSolomon Islands.Our results
demonstratethatpopulationdensityandmarketaccessincrease fishing pressure, which is a major driver of fish diversity and functional group biomass. These distal social drivers have a negative effect on the biomass of piscivores and herbivores targetedby fishers.Moreovertherelative abundanceof species becomesmoreeven,whilstspeciesrichnessandAvTDdeclineas populationdensityincreasesandmarketsbecomemoreaccessible.
4.1. Explainingtheeffectsofproximatedriversonfishfunctionand diversity
Adeclineintaxonomicdistinctnessisoftenassociatedwitha declineinfunctionaldiversity(Rogersetal.,1999;Chapinetal., 2000;Nystro¨metal.,2000).Thisissupportedherebythefinding thatfishingpressurehadanegativeeffectonaveragetaxonomic distinctnessandthebiomassoftwoimportantfunctionalgroups, herbivoresandpiscivores.Conversely,therewasnegligibleeffect offishingonnon-targetspeciesfromthesefunctionalgroups.Thus, thedirecteffectoffishingislikelytobeconfinedlargelytothose species and functional groups that are targeted by fishers. A decrease in diversity, and increased evenness with increased fishingpressure,mightrelatetoremovalofrelativelyrarelarge bodiedpredators,whichareoftentargetedbyfishers(Paulyetal., 1998).
Target specieson coral reefstendtobelarge bodied(Dulvy etal.,2004b;Grahametal.,2005)andmanyofthesmallspecies have closeaffiliation withthereefbenthos(Mundayand Jones, 1998).Thisissupportedbythefindingthatcoralcoverandfishing pressurehadasimilareffectonnon-targetherbivoreandpiscivore biomass, compared to all herbivore and all piscivore biomass whichwaslargelyexplainedbyfishingpressurealone.For non-targetherbivorestherelationshipwithcoralcoverwasnegative, possiblybecausemanyofthesefisharedamselfishesthatmaintain territories covered with algae (Ceccarelli, 2007) (ESM 1). Con-versely,biomassofpiscivores,particularlynon-targetedspecies, tendedtoincreasewithcoralcover.Thismaybebecausemany smallerbodied non-targetpredators and theirprey take refuge among corals. Indeed a loss of coral and associated structural complexitycanleadtodeclinesinsmallbodiedpreyfishandtheir mediumsizedpredators(Grahametal.,2007).Functionally, non-target speciesare likely toperforma verydifferentrole tothe larger bodied species targeted by fishers. Fishing and habitat degradationmightthereforehavedifferentconsequencesforboth herbivoreandpiscivoreassemblagesandthefunctionalservices associatedwiththesegroups.
4.2. Explainingtheeffectsofdistaldriversonfishfunctionand diversity
Bydisaggregatingdistalandproximatedriversandmodeling fish assemblage response to different causal paths of human activity,wehaveshownthatsocialdriverscanhavebothpositive andnegativeeffectsonfishcommunitiesandtheirfunctionalrole inecosystems.Populationdensityhadbothpositiveandnegative effectson allherbivorebiomassthroughcoralcoverandfishing pressure,respectively.Thepositiveeffect,throughdecreasedcoral cover,mightbeexplainedbyincreasednutrientlevelsindirectly caused by high coastal population densities without access to sewagetreatmentfacilities.Resultantexcessnutrientshavebeen showntoincreasealgalgrowth(e.g.PastorokandBilyard,1985), andconsequentlyincreasefoodavailabilitytoherbivores.
increasedspeciesrichness, average taxonomicdistinctness, and functionalgroupbiomass.Ourresultsarebroadlyconsistentwith studiesconductedacrossfiveIndianOceancountriesthatfounda decreaseinfishingwithhigherlevelsofsocioeconomic develop-ment(Cinneretal.,2009a;CinnerandBodin,2010).Incomparison tothelargesocioeconomicdevelopmentspectruminthese multi-nationstudies,therelativelysmall effectsizeof socioeconomic developmenton fishingpressure presentedin this studymight reflectasmalldevelopmentgradientinSolomonIslands.
Themajorityofstudiesthathaveexploredtheeffectofhuman activityoncoralreeffishdiversityandfunctionhaveshownthat theseassemblage characteristicsareexplainedbyeitherfishing pressure (Jennings et al., 1995; Jennings and Polunin, 1996; DeMartinietal.,2008)orhumanpopulationdensity(Jenningsand Polunin,1997;Bellwoodetal.,2003;Dulvyetal.,2004a,b;Mora, 2008;Williamsetal.,2008,2011;Stallings,2009)(Table1).While itisclearthatlocalhumanpopulationdensityanddirectfishing effectsareimportantinexplainingecologicalgradients,wehave shownhere thattrade, measuredas distancetomarket,isalso important(Fig.2).IntheSolomonIslandstradelikelyaffectsfish diversityandfunctionthroughsmall-scalecommercialfishingto supplyurbanmarkets,whereaspopulation densitylikelyaffects diversityandfunctionthroughsemi-subsistencebasedfishingto supply local needs. Trade allows societies toacquire resources fromfurtherafield,externalizingenvironmentalfootprintsbeyond local human–environmentsystems (Arrow et al., 1995; Berkes et al., 2006; Shandra et al., 2009). Resource management and biodiversityconservationinitiativesmustrecognizethattradeand localpopulationpressurerepresentdifferentdriversofecological degradation, and should therefore consider applying different strategies to address their different effects on ecosystems. For example, strong governance of markets through sustainable harvesting certification, and market-specific gear and species restrictions,willbecomeincreasinglyimportantifcoralreeffish continuetobeareadilytradedcommodity(Berkesetal.,2006).
4.3. Futuremodelextensions
Expansionofthemodelsdevelopedinthispapertoother social-ecologicalcontextswouldhelptoprovideabetterunderstanding ofhowmarineecosystemswillrespondtokeysocialdynamics. However,threekeyadvancementsarenecessarytoimprovethe predictivecapacityofsuchmodels.First,analysisoftheindirect effects of distal drivers on theproportionate representation of multiplefunctionalgroups(includinghigherresolutionherbivore functional groups such as grazers, scrapers, and excavators) (Wilson and Bellwood, 1997) and species might lend further insightintotheroleofdistaldriversinshapingecosystemfunction (Wilsonetal.,2008).Second,coralcoverisonlyonemeasureof coral reef habitat and more detailed models including other environmental and habitat variables (e.g. Wilson et al., 2008), couldshedadditionallight ontherelativecontributionofdistal driversondiversityandfunction,particularlyforspeciesrichness and non-target assemblages of coral reef fish. Third, temporal assessmentswouldbevitaltounderstandthefeedbacksthatmight existinthissystem.
5. Conclusion
Managementmeasureswhichaddressproximatedrivers,such asfishingpressure,typicallyhavelocalizedeffectsondiversityand ecosystemfunction.Yet,theyarelimitedintheirabilitytoalleviate the effects of distal social drivers such as market access and socioeconomicdevelopment(Birkeland,2004).Therefore,whilst managing proximate threats represents an important (if not limited) management approach, and means of increasing local
resilience, governing reefs in a changing world will require becoming better acquainted withthe challenges, and potential solutions posed by broadersocial drivers suchas marketsand development.
Acknowledgments
WethanktheAustralianResearchCouncilCentreofExcellence forCoralReefStudiesforfundingthisresearch.ThankyoutoA. Hughes for providing substrate data. Thank you to theNature ConservancyandtheSolomonIslandsstatisticsoffice,particularly N.Gagaheforprovidingdata.ThankyoutoK.MoonandN.Graham forhelpfulcommentsonearlierversionsofthemanuscript.
AppendixA.Supplementarydata
Supplementarydataassociatedwiththisarticlecanbefound,in theonlineversion,atdoi:10.1016/j.gloenvcha.2012.01.006.
References
Agrawal,A.,Yadama,G.,1997.Howdolocalinstitutionsmediate marketand populationpressuresonresources?ForestPanchayatsinKumaon,India. De-velopmentandChange28,435–465.
ANUEnterprise,2008.People’sSurvey2008.AustralianNationalUniversity. Arrow,K.,Bolin,B.,Costanza,R.,Dasgupta,P.,Folke,C.,Holling,C.S.,Jansson,B.-O.,
Levin, S.,Ma¨ler, K.-G.,Perrings, C.,Pimentel, D., 1995.Economicgrowth, carryingcapacity,andtheenvironment.EcologicalEconomics15,91–95. Aswani,S.,1999.Commonpropertymodelsofseatenure:acasestudyfromthe
RovianaandVonavonaLagoons,NewGeorgia,SolomonIslands.HumanEcology 27,417–453.
Aswani,S.,Sabetian,A.,2009.Implicationsofurbanizationforartisanalparrotfish fisheriesintheWesternSolomonIslands.ConservationBiology24,520–530. Aswani,S.,2011.Socioecologicalapproachesforcombiningecosystem-basedand customarymanagementinOceania.JournalofMarineBiology,doi:10.1155/ 2011/845385.
Bell,J.D.,Galzin,R.,1984.Influenceoflivecoralcoveroncoral-reeffish communi-ties.MarineEcologyProgressSeries15,265–274.
Bell,J.D.,Kronen,M.,Vunisea,A.,Nash,W.J.,Keeble,G.,Demmke,A.,Pontifex,S., Andre´foue¨t,S.,2009.PlanningtheuseoffishforfoodsecurityinthePacific. MarinePolicy33,64–76.
Bellwood,D.R.,Hoey,A.S.,Choat,J.H.,2003.Limitedfunctionalredundancyinhigh diversitysystems:resilienceandecosystemfunctiononcoralreefs.Ecology Letters6,281–285.
Bellwood,D.R.,Hughes,T.P.,2001.Regional-scaleassemblyrulesandbiodiversityof coralreefs.Science292,1532–1535.
Bellwood,D.R.,Hughes,T.P.,Connolly,S.R.,Tanner,J.,2005.Environmentaland geometricconstraintsonIndo-Pacificcoralreefbiodiversity.EcologyLetters8, 643–651.
Bellwood,D.R.,Hughes,T.P.,Hoey,A.S.,2006.Sleepingfunctionalgroupdrives coral-reefrecovery.CurrentBiology16,2434–2439.
Berkes,F.,Hughes,T.P.,Steneck,R.S.,Wilson,J.A.,Bellwood,D.R.,Crona,B.,Folke,C., Gunderson,L.H.,Leslie,H.M.,Norberg,J.,Nystro¨m,M.,Olsson,P.,O¨ sterblom,H., Scheffer, M., Worm, B., 2006. Globalization, roving bandits, and marine resources.Science311,1557–1558.
Birkeland,C.,2004.Ratchetingdownthecoralreefs.BioScience54,1021–1027. Brewer,T.D.,Cinner,J.E.,Green,A.,Pandolfi,J.M.,2009.Thresholdsandmultiple
scaleinteractionofenvironment,resourceuse,andmarketproximityonreef fisheryresourcesintheSolomonIslands.BiologicalConservation142,1797– 1807.
Cardinale,B.J.,Srivastava,D.S.,EmmettDuffy,J.,Wright,J.P.,Downing,A.L., San-karan,M.,Jouseau,C.,2006.Effectsofbiodiversityonthefunctioningoftrophic groupsandecosystems.Nature443,989–992.
Ceccarelli,D.,2007.Modificationofbenthiccommunitiesbyterritorialdamselfish: amulti-speciescomparison.CoralReefs26,853–866.
ChapinIII,F.S.,Zavaleta,E.S.,Eviner,V.T.,Naylor,R.L.,Vitousek,P.M.,Reynolds,H.L., Hooper,D.U.,Lavorel,S.,Sala,O.E.,Hobbie,S.E.,Mack,M.C.,Diaz,S.,2000. Consequencesofchangingbiodiversity.Nature405,234–242.
Chin,W.W.,Newstead,P.R.,1999.Structuralequationmodelinganalysiswithsmall samplesusingpartialleastsquares.In:Hoyle,R.H.(Ed.),StatisticalStrategies forSmallSampleResearch.Sage,ThousandOaks,CA.
Choat,J.H.,Clements,K.D.,Robbins,W.D.,2002.Thetrophicstatusofherbivorous fishesoncoralreefs.1:Dietaryanalyses.MarineBiology140,613–623. Cinner,J.E.,McClanahan,T.R.,2006.Socioeconomicfactorsthatleadtooverfishing
Cinner,J.E.,McClanahan,T.R.,Daw,T.M.,Graham,N.A.J.,Maina,J.,Wilson,S.K., Hughes,T.P.,2009a.Linkingsocialandecologicalsystemstosustaincoralreef fisheries.CurrentBiology19,206–212.
Cinner,J.E.,McClanahan,T.R.,Graham,N.A.J.,Pratchett,M.S.,Wilson,S.K.,Raina, J.-B.,2009b.Gear-basedfisheriesmanagementasapotentialadaptiveresponseto climatechangeandcoralmortality.JournalofAppliedEcology46,724–732. Cinner,J.E.,Bodin,O¨ .,2010.Livelihooddiversificationintropicalcoastal
communi-ties:anetwork-basedapproachtoanalyzing‘LivelihoodLandscapes’.PLoSONE 5,e11999.
Clarke,K.R.,Warwick,R.M.,1999.Thetaxonomicdistinctnessmeasureof biodiver-sity:weightingofsteplengthsbetweenhierarchicallevels.MarineEcology ProgressSeries184,21–29.
Clarke,K.R.,Warwick,R.M.,2001.Afurtherbiodiversityindexapplicabletospecies lists:variationintaxonomicdistinctness.MarineEcologyProgressSeries216, 265–278.
Connolly,S.R.,Bellwood,D.R.,Hughes,T.P.,2003.Indo-pacificbiodiversityofcoral reefs:deviationsfromamid-domainmodel.Ecology84,2178–2190. DeMartini,E.E.,Friedlander,A.M.,Sandin,S.A.,Sala,E.,2008.Differencesin
fish-assemblagestructurebetweenfishedandunfishedatollsinthenorthernLine Islands,centralPacific.MarineEcologyProgressSeries365,199–215. Dulvy,N.K.,Freckleton,R.P.,Polunin,N.V.C.,2004a.Coralreefcascadesandthe
indirecteffectsofpredatorremovalbyexploitation.EcologyLetters7,410–416. Dulvy,N.K.,Polunin,N.V.C.,Mill,A.C.,Graham,N.A.J.,2004b.Sizestructuralchange inlightlyexploitedcoralreeffishcommunities:evidenceforweakindirect effects.CanadianJournalofFisheriesandAquaticSciences61,466–475. Floeter,S.R.,Krohling,W.,Gasparini,J.L.,Ferreira,C.E.,Zalmon,I.R.,2007.Reeffish
communitystructureoncoastalislandsofthesoutheasternBrazil:the influ-enceofexposureandbenthiccover.EnvironmentalBiologyofFishes78,147– 160.
Forester,D.J.,Machlis,G.E.,1996.Modelinghumanfactorsthataffectthelossof biodiversity.ConservationBiology10,1253–1263.
Friedlander,A.M.,Brown,E.K.,Jokiel,P.L.,Smith,W.R.,Rodgers,K.S.,2003.Effectsof habitat,waveexposure,andmarineprotectedareastatusoncoralreeffish assemblagesintheHawaiianarchipelago.CoralReefs22,291–305. Friedlander,A.M.,Parrish,J.D.,1998.Habitatcharacteristicsaffectingfish
assem-blagesonaHawaiiancoralreef.JournalofExperimentalMarineBiologyand Ecology224,1–30.
Froese,R., Pauly,D.,2011. Fishbase. World Wide WebElectronic Publication (accessed07.2011)www.fishbase.org.
Fulton,C.J.,Bellwood,D.R.,2005.Wave-inducedwatermotionandthefunctional implicationsforcoralreeffishassemblages.LimnologyandOceanography50, 255–264.
Geist,H.J.,Lambin,E.F.,2002.Proximatecausesandunderlyingdrivingforcesof tropicaldeforestation.BioScience52,143–150.
Graham,N.A.J., Dulvy,N.K.,Jennings, S.,Polunin, N.V.C.,2005. Size-spectraas indicatorsoftheeffectsoffishingoncoralreeffishassemblages.CoralReefs 24,118–124.
Graham,N.A.J.,Evans,R.D.,Russ,G.R.,2003.Theeffectsofmarinereserveprotection onthetrophicrelationshipsofreeffishesontheGreatBarrierReef. Environ-mentalConservation30,200–208.
Graham,N.A.J., McClanahan, T.R.,MacNeil, M.A.,Wilson, S.K.,Polunin, N.V.C., Jennings,S.,Chabanet,P.,Clark,S.,Spalding,M.D.,Letourneur,Y.,Bigot,L., Galzin, R.,o¨hman,M.C., Garpe,K.C., Edwards,A.J.,Sheppard,C.R.C.,2008. Climatewarming,marineprotectedareasandtheocean-scaleintegrityofcoral reefecosystems.PLoSONE3,e3039.
Graham,N.A.J.,Wilson,S.K.,Jennings,S.,Polunin,N.V.C.,Bijoux,J.P.,Robinson,J., 2006.Dynamicfragilityofoceaniccoralreefecosystems.Proceedingsofthe NationalAcademyofSciences103,8425–8429.
Graham,N.A.J.,Wilson,S.K.,Jennings,S.,Polunin,N.V.C.,Robinson,J.A.N.,Bijoux,J.P., Daw,T.M.,2007.Lageffectsintheimpactsofmasscoralbleachingoncoralreef fish,fisheries,andecosystems.ConservationBiology21,1291–1300. Gratwicke,B.,Speight,M.R.,2005.Therelationshipbetweenfishspeciesrichness,
abundanceand habitatcomplexity ina rangeof shallowtropical marine habitats.JournalofFishBiology66,650–667.
Green,A.,Bellwood,D.R.,2009.MonitoringFunctionalGroupsofHerbivorousReef FishesasIndicatorsofCoralReefResilience–APracticalGuideforCoralReef ManagersintheAsiaPacificRegion.IUCN,Gland,Switzerland.
Green,A.,Lokani,P.,Atu,W.,Ramohia,P.,Thomas,P.,Almany,J.,2006a.Solomon IslandsMarineAssessment:TechnicalReportofSurveyConductedMay13to June17,2004.TNCPacificIslandCountriesReportNo.1/06..
Green,A.,Ramohia,P.,Ginigele,M.,Leve,T.,2006b.Fisheriesresources:coralreef fishes.In:Green,A.,Lokani,P.,Atu,W.,Ramohia,P.,Thomas,P.,Almany,J. (Eds.),SolomonIslandsMarineAssessment:TechnicalReportofSurvey Con-ductedMay13toJune17,2004.TNCPacificIslandCountriesReportNo.1/06. Hoey,A.S.,Bellwood,D.R.,2009.Limitedfunctionalredundancyinahighdiversity system:singlespeciesdominateskeyecologicalprocessoncoralreefs. Eco-systems12,1316–1328.
Hooper,D.U.,Chapin,F.S.,Ewel,J.J.,Hector,A.,Inchausti,P.,Lavorel,S.,Lawton,J.H., Lodge,D.M.,Loreau,M.,Naeem,S.,Schmid,B.,Seta¨la¨,H.,Symstad,A.J., Van-dermeer,J.,Wardle,D.A.,2005.Effectsofbiodiversityonecosystem function-ing:aconsensusofcurrentknowledge.EcologicalMonographs75,3–35. Hughes,A.,2006. BenthicCommunities,Solomon IslandsMarineAssessment:
TechnicalReportofSurveyConductedMay13toJune17,2004.TNCPacific IslandCountriesReportNo.1/06..
Hughes,T.P.,1994.Catastrophes,phaseshifts,andlarge-scaledegradationofa Caribbeancoralreef.Science265,1547–1551.
Hughes,T.P.,Rodrigues,M.J.,Bellwood,D.R.,Ceccarelli,D.,Hoegh-Guldberg,O., McCook,L.,Moltschaniwskyj,N.,Pratchett,M.S.,Steneck,R.S.,Willis,B.,2007. Phaseshifts,herbivory,andtheresilienceofcoralreefstoclimatechange. CurrentBiology17,360–365.
Ives,A.R.,Carpenter,S.R.,2007.Stabilityanddiversityofecosystems.Science317, 58–62.
Jennings,S.,Grandcourt,E.M.,Polunin,N.V.C.,1995.Theeffectsoffishingonthe diversity,biomassandtrophicstructureofSeychelles’reeffishcommunities. CoralReefs14,225–235.
Jennings,S.,Polunin,N.V.C.,1997.Impactsofpredatordepletionbyfishingonthe biomassanddiversityofnon-targetreeffishcommunities.CoralReefs16,71–82. Jennings,S.,Polunin,N.V.C.,1996.Effectsoffishingeffortandcatchrateuponthe structure andbiomassofFijian reeffishcommunities.JournalofApplied Ecology33,400–412.
Jones,G.P.,Planes,S.,Thorrold,S.R.,2005.Coralreeffishlarvaesettleclosetohome. CurrentBiology15,1314–1318.
Kock,N.,2010.WarpPLSß1.0UserManual.ScriptWarpSystems,Laredo,TX. Kramer,D.B.,Urquhart,G.,Schmitt,K.,2009.Globalizationandtheconnectionof
remotecommunities:areviewofhouseholdeffectsandtheirbiodiversity implications.EcologicalEconomics68,2897–2909.
Luckhurst,B.E.,Luckhurst,K.,1978.Analysisoftheinfluenceofsubstratevariables oncoralreeffishcommunities.MarineBiology49,317–323.
Maestre,F.T.,Quero,J.L.,Gotelli,N.J.,Escudero,A.,Ochoa,V.,Delgado-Baquerizo,M., Garcı´a-Go´mez,M.,Bowker,M.A.,Soliveres,S.,Escolar,C.,Garcı´a-Palacios,P., Berdugo,M.,Valencia,E.,Gozalo,B.,Gallardo,A.,Aguilera,L.,Arredondo,T., Blones,J.,Boeken,B.,Bran,D.,Conceic¸a˜o,A.A.,Cabrera,O.,Chaieb,M.,Derak,M., Eldridge,D.J.,Espinosa,C.I.,Florentino,A.,Gaita´n,J.,Gatica,M.G.,Ghiloufi,W., Go´mez-Gonza´lez,S.,Gutie´rrez,J.R.,Herna´ndez,R.M.,Huang,X., Huber-Sann-wald,E.,Jankju,M.,Miriti,M.,Monerris,J.,Mau,R.L.,Morici,E.,Naseri,K., Ospina,A.,Polo,V.,Prina,A.,Pucheta,E.,Ramı´rez-Collantes,D.A.,Roma˜o,R., Tighe,M.,Torres-Dı´az,C.,Val,J.,Veiga,J.P.,Wang,D.,Zaady,E.,2012.Plant speciesrichnessandecosystemmultifunctionalityinglobaldrylands.Science 335,214–218.
McCann,K.S.,2000.Thediversity-stabilitydebate.Nature405,228–233. McClanahan,T.R.,Maina,J.M.,Muthiga,N.A.,2011.Associationsbetweenclimate
stressandcoralreefdiversityinthewesternIndianOcean.GlobalChange Biology17,2023–2032.
McKinney,L.A.,Kick,E.L.,Fulkerson,G.M.,2010.Worldsystem,anthropogenic,and ecologicalthreatstobirdandmammalspecies:astructuralequationanalysisof biodiversityloss.OrganizationandEnvironment23,3–31.
MollesJr.,M.C.,1978.Fishspeciesdiversityonmodelandnaturalreefpatches: experimentalinsularbiogeography.EcologicalMonographs48,289–305. Mora, C.,2008. Aclear human footprintinthe coralreefs oftheCaribbean.
ProceedingsoftheRoyalSocietyB:BiologicalSciences275,767–773. Mora,C.,Aburto-Oropeza,O.,AyalaBocos,A.,Ayotte,P.M.,Banks,S.,Bauman,A.G.,
Beger,M.,Bessudo,S.,Booth,D.J.,Brokovich,E.,Brooks,A.,Chabanet,P.,Cinner, J.E.,Corte´s,J.,Cruz-Motta,J.J.,CupulMagan˜ a,A.,DeMartini,E.E.,Edgar,G.J., Feary,D.A.,Ferse,S.C.A.,Friedlander,A.M.,Gaston,K.J.,Gough,C.,Graham,N.A.J., Green,A.,Guzman,H.,Hardt,M.,Kulbicki,M.,Letourneur,Y.,Lo´pezPe´rez,A., Loreau,M.,Loya,Y.,Martinez,C.,Mascaren˜ as-Osorio,I.,Morove,T.,Nadon, M.-O.,Nakamura,Y.,Paredes,G.,Polunin,N.V.C.,Pratchett,M.S.,ReyesBonilla,H.c., Rivera,F.,Sala,E.,Sandin,S.A.,Soler,G.,Stuart-Smith,R.,Tessier,E.,Tittensor, D.P.,Tupper,M.,Usseglio,P.,Vigliola,L.,Wantiez,L.,Williams,I.,Wilson,S.K., Zapata,F.A.,2011.Globalhumanfootprintonthelinkagebetweenbiodiversity andecosystemfunctioninginreeffishes.PLoSBiology9,e1000606. Mora, C.,Chittaro,P.M.,Sale,P.F.,Kritzer,J.P.,Ludsin,S.A.,2003.Patternsand
processesinreeffishdiversity.Nature421,933–936.
Mora,C.,Robertson,D.R.,2005.Causesoflatitudinalgradientsinspeciesrichness:a testwithfishesofthetropicalEasternPacific.Ecology86,1771–1782. Muller,D.,Zeller,M.,2002.LandusedynamicsinthecentralhighlandsofVietnam:
aspatialmodelcombiningvillagesurveydatawithsatelliteimagery interpre-tation.AgriculturalEconomics27,333–354.
Mumby,P.J.,2006.Theimpactofexploitinggrazers(Scaridae)onthedynamicsof Caribbeancoralreefs.EcologicalApplications16,747–769.
Munday,P.L.,Jones,G.P.,1998.Theecologicalimplicationsofsmallbodysizeamong coral-reeffishes.OceanographyandMarineBiologyAnnualReview36,373–411. Naeem, S., 1998. Speciesredundancy and ecosystem reliability. Conservation
Biology12,39–45.
Newton,K.,Cote,I.M.,Pilling,G.M.,Jennings,S.,Dulvy,N.K.,2007.Currentand futuresustainabilityofislandcoralreeffisheries.CurrentBiology17,655–658. Nystro¨m,M.,Folke,C.,Moberg,F.,2000.Coralreefdisturbanceandresilienceina human-dominatedenvironment.TrendsinEcologyandEvolution15,413–417. O¨ hman,M.C.,Rajasuriya,A.,1998.Relationshipsbetweenhabitatstructureandfish communitiesoncoralandsandstonereefs.EnvironmentalBiologyofFishes53, 19–31.
Pandolfi,J.M.,Bradbury,R.H.,Sala,E., Hughes,T.P., Bjorndal,K.A.,Cooke,R.G., McArdle,D.,McClenachan,L.,Newman,M.J.H.,Paredes,G.,Warner,R.R., Jack-son, J.B.C.,2003. Globaltrajectoriesofthelong-termdeclineofcoralreef ecosystems.Science301,955–958.
Pastorok,R.A.,Bilyard,G.R.,1985.Effectsofsewagepollutiononcoral-reef com-munities.MarineEcologyProgressSeries21,175–189.
Pauly,D.,Christensen,V.,Dalsgaard,J.,Froese,R.,Torres,F.,1998.Fishingdown marinefoodwebs.Science279,860–863.
Reinartz,W.,Haenlein,M.,Henseler,J.,2009.Anempirical comparisonofthe efficacyofcovariance-basedandvariance-basedSEM. InternationalJournal ofResearchinMarketing26,332–344.
Roberts,C.M.,Ormond,R.F.G.,1987.Habitatcomplexityandcoralreeffishdiversity andabundanceonRedSeafringingreefs.MarineEcologyProgressSeries41,1–8. Rogers,S.I.,Clarke,K.R.,Reynolds,J.D.,1999.Thetaxonomicdistinctnessofcoastal bottom-dwellingfishcommunitiesoftheNorth-eastAtlantic.JournalofAnimal Ecology68,769–782.
Russ,G.R.,2003.Grazerbiomasscorrelatesmorestronglywithproductionthan withbiomassofalgalturfsonacoralreef.CoralReefs22,63–67.
Sandin,S.A.,Smith,J.E.,DeMartini,E.E.,Dinsdale,E.A.,Donner,S.D.,Friedlander, A.M.,Konotchick,T.,Malay,M.,Maragos,J.E.,Obura,D.,Pantos,O.,Paulay,G., Richie,M.,Rohwer,F.,Schroeder,R.E.,Walsh,S.,Jackson,J.B.C.,Knowlton,N., Sala,E.,2008.Baselinesanddegradationofcoralreefsinthenorthernline islands.PLoSONE3,e1548.
Shandra,J.M.,Leckband,C.,McKinney,L.A.,London,B.,2009.Ecologicallyunequal exchange,worldpolity,andbiodiversityloss.InternationalJournalof Compar-ativeSociology50,285–310.
Stallings,C.D.,2009.Fishery-independentdatarevealnegativeeffectofhuman populationdensityonCaribbeanpredatoryfishcommunities.PLoSONE4, e5333.
Tilman,D.,Reich,P.B.,Knops,J.M.H.,2006.Biodiversityandecosystemstabilityina decade-longgrasslandexperiment.Nature441,629–632.
Williams,I.D.,Richards,B.L.,Sandin,S.A.,Baum,J.K.,Schroeder,R.E.,Nadon,M.O., Zgliczynski, B.,Craig, P.,McIlwain,J.L.,Brainard,R.E.,2011. Differencesin reef fish assemblages between populated and remote reefs spanning multiplearchipelagosacrossthecentralandWesternPacific.JournalofMarine Biology14.
Williams,I.D.,Walsh,W.J.,Schroeder,R.E.,Friedlander,A.M.,Richards,B.L., Sta-moulis,K.A.,2008.AssessingtheimportanceoffishingimpactsonHawaiian coralreeffishassemblagesalongregional-scalehumanpopulationgradients. EnvironmentalConservation35,261–272.
Wilson,S.,Bellwood,D.R.,1997.Crypticdietarycomponentsofterritorial damsel-fishes(Pomacentridae,Labroidei).MarineEcologyProgressSeries153,299– 310.
Wilson,S.K.,Fisher,R.,Pratchett,M.S.,Graham,N.A.J.,Dulvy,N.K.,Turner,R.A., Cakacaka,A.,Polunin, N.V.C., Rushton,S.P.,2008. Exploitationandhabitat degradationasagentsofchangewithincoralreeffishcommunities.Global ChangeBiology14,2796–2809.