0022-538X/90/094428-10$02.00/0
Copyright © 1990,American Society forMicrobiology
Function of
the Human
Immunodeficiency
Virus
Types
1and
2
Rev
Proteins
Is
Dependent
on
Their
Ability
To Interact with
a
Structured
Region
Present
in
env
Gene
mRNA
PATRICK J. DILLON, PETER NELBOCK, ANN PERKINS,AND CRAIG A. ROSEN* Department ofMolecularOncology andVirology, Roche Institute ofMolecularBiology,
RocheResearch Center,
Nutley,
NewJersey
07110-1199 Received 19April1990/Accepted 15 June 1990The interaction of the humanimmunodeficiency virustype 1(HIV-1)Revproteinwithastructured region inenvmRNA(the Rev-responsive element [RRE])mediatestheexportof structuralmRNAs from thenucleus tothecytoplasm. We demonstrated that unlikeHIV-1 Rev, which functions with both theHIV-1and HIV-2 RREs, HIV-2 Rev functions only with theHIV-2 RRE. Rev-RREbinding studiessuggested thatthe lackof nonreciprocal complementation stems from the inability of HIV-2 Rev to interact with HIV-1 RRE RNA. Maintenance ofRNAsecondarystructure, ratherthan the primarynucleotidesequence, appearedto be the major determinant for interaction of both HIV-1 and HIV-2 Rev with the HIV-2 RRE.Moreover,thebinding domain of the HIV-2 RRErecognized by HIV-1 Revwasdissimilar to thebinding domain of the HIV-1RRE, intermsof both secondarystructureandprimary nucleotidesequence. Our resultssupportthehypothesisthat function of HIV RevproteinsandpossiblythefunctionallysimilarRexproteinsencodedbythe human T-cell leukemia viruses (HTLVs) HTLV-I andHTLV-II is controlled by the presence ofRNA secondarystructure generated within the RRE RNA.
Regulation ofhuman immunodeficiency virus(HIV)gene
expressionisdependentonexpressionofnumerouscell- and
virus-encoded trans-acting regulatory proteins. Expression oftwoofthesevirus-encodedproteins, Tat(1, 37)and Rev (12, 36),isrequiredforreplication (7, 15, 38). The evolution-arily and structurally related HIV type 2 (HIV-2) virus displays many similarities with HIV-1, including the pres-enceoftatandrevgenes (18).
The cis-acting elements responsive to Tat and Rev are
known. Both proteins function through RNA secondary structures present in either the 5' long terminalrepeat (14, 31)or envmessage, respectively (8, 11, 13, 19, 20, 27, 32).
Although the HIV-1 and HIV-2 Tat proteins function in a
reciprocal manner through the trans-acting responsive
re-gions ofeach other(10), Rev function appears to be more
restricted. For example, whereas HIV-1 Rev can function
withastructured regionin theenvRNA(the Rev-responsive element [RRE])ofHIV-2,thereciprocalcasedoesnothold true (26). Recent studies have shown thatHIV-1 Rev inter-actswith HIV-1 RRERNA(3, 6, 9, 27-29, 39)and that this interaction is primarily determined by the presence of a
hairpin loop structure within the RRE (9, 22, 28, 29). Once bound tothe RRE, Rev protein, eitherby itselforthrough
interaction with additional cellular factors, is thought to
mediate the export of viral structural mRNAs from the nucleustothe cytoplasm.
Tounderstandthemechanism for HIV-2Revfunctionand thebasisfor its inabilitytofunction with theHIV-1 RRE,we
examined the nature of HIV-2 Rev interactions with the different RRE elements. Our results show that, like HIV-1 Rev, theHIV-2 Rev protein interacts with the HIV-2 RRE and that maintenance of secondary structure, rather than primary nucleotide sequence, is the major determinant for
thisinteraction.Inaddition,the HIV-2RRE RNAstructures
* Corresponding author.
required for HIV-1 Rev interaction closely coincide with those required for HIV-2 Rev binding yet are completely dissimilar, in terms of both structure and nucleotide
se-quence, to the HIV-1 Revbinding domainidentified within the HIV-1 RRE.
MATERIALS AND METHODS
Plasmid constructions. (i) Construction ofeucaryotic Rev expression vectors. The construction of the simian virus 40-basedvectorforexpressionofRev, BLpSVRev,hasbeen described previously (5). For expression of HIV-2 Rev, a
synthetic cDNA wasfirst derived by using the polymerase chain reaction (PCR) method (34, 35). Exons 1 and 2 of HIV-2 Revwereamplified froman
HIV-2ROD
bacteriophageplaque. The 5' primer used for amplification of exon 1 contained a BamHI site, and the 3' primer contained an
extension ofcodingsequenceforamino acids 25to 35,with
an engineered KpnI siteatamino acid 34. The 5' primerfor
exon 2contained anengineered KpnI site atamino acid 34, and the 3' primer contained a HindIII site. PCR-amplified exons were cut, ligated, and cloned into BL-KS (Strata-gene).Theresultantplasmid, BLHIV-2Rev,wassequenced
and contained a single nucleotide difference from the
pub-lished sequence. This difference changed the aspartic acid present atamino acid66toasparagine. TheHIV-2 Revgene was cloned into a simian virus 40 expression plasmid,
BLpSV, which contains the simian virus 40 promoter and polyadenylation signals. Briefly, the HindlIl site of BLHIV-2Revwas convertedtoBglII,andafragmentcontaining the
HIV-2RevcDNA wascloned atthe XbaI-BglII sitesin the polylinker of BLpSV. The resultant plasmid, pSVHIV-2Rev, was usedfortransfections ineucaryotic cells.
(ii)Constructionof Escherichia coli Revexpressionvectors.
The E. coli Rev expression plasmid, pRC-25 synart, has beendescribedpreviously (3). Expressionof HIV-2 Rev in E. coli wasachieved through the use of PCRtomodify the HIV-2Revsequencetocontaincodonsmostfrequently used
4428
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTION OF HIV REV PROTEINS 4429 in E. coli (P. Dillon and C. Rosen, BioTechniques, in press).
Briefly, four overlapping oligonucleotides, each 105 base pairs in length, spanning the entire Rev coding sequence were synthesized. These oligonucleotides contained codons which were derived from an E. coli high-usage codon table. Thefour oligonucleotides were amplified in a standard PCR mixture for 8 cycles. A second PCR mixture containing short flanking primers gave rise to a 320-base-pair PCR product which was cloned as a BamHI-HindIII fragment into the procaryotic expression plasmid pDS56 (4). This plasmid contains sequences upstream from the BamHI site which code for a tag sequence, Met-Arg-Gly-Ser-(His)6-Gly-Ser, which precedes in frame the amino terminus of HIV-2 Rev. The resultant plasmid, designated pDSHIV-2H6Rev, was usedfor expression of HIV-2 Rev in E. coli. In addition, an EcoRI-XbaI fragment containing the HIV-2 Rev sequence and the extra amino-terminal amino acids from pDSHIV-2H6Rev was cloned into BLpSV and the resultant plasmid, pSVHIV-2H6Rev, was assessed for function in eucaryotic cells.
Subcloning and mutagenesis of the HIV-2 and SIV RREs. The RREs for HIV-2 and simian immunodeficiency virus
(SIV) wereobtained by PCR. The HIV-2 RRE (nucleotides 7608to7973)wasamplified froman
HIV-2ROD
phage plaque and cloned as a ClaI-XbaI fragment into the Bluescript vector(Stratagene). The SIV RRE (nucleotides 8086 to 8454) wasamplified from plasmid pBK28-SIV and was also clonedinto Bluescript as a ClaI-XbaI fragment. Stem-loop
muta-tions(PD-1 through PD-21) were generated by site-directed mutagenesis (25) of the Bluescript vector containing the HIV-2 RRE, BLHIV-2RRE. 5' and 3' deletion mutations weregeneratedby PCR. A 5' PCR primer which contained a 5' T7promoter-ClaI site RRE coding sequence was
synthe-sized. The 3' primer contained an XbaI site to facilitate
cloning directly from the PCR products. Amplicons were
used forboth T7 transcription in vitro and subcloning into
the RevindicatorplasmidpIlIAR(seebelow).
Construction of Rev-responsive vectors. The plasmid
pIIIAR has been described previously (32). Briefly, this
plasmid contains the HIV-1 long terminal repeat and the gene for chloramphenicol acetyltransferase (CAT) fused to the env sequence from HIV-1. Theenvgene has aBamHI
frameshift which prevents expression of Rev. In addition,
the env sequence has been modified to contain a ClaI site
(nucleotide 7204) upstream and an XbaI site (nucleotide 7684) site downstream ofthe Rev response element. The wild-type andmutantRRE sequencesdescribed abovewere cloned into the unique ClaI-XbaI sites of the
pIlIAR
plas-mid.
Expression and purification of Rev protein from E. coli.
Expressionand purification ofHIV-1 Rev
protein
has beenpreviously described (3). Expression and
purification
of HIV-2 H6Rev was accomplished byusing
methods de-scribedpreviously (4, 16).Briefly,
expression
ofHIV-2Revproteinwasinduced bytheadditionof1mM
isopropyl-3-D-thiogalactopyranoside (IPTG) to a
logarithmically
growing
culture. The induced E. coli were
harvested,
lysed
withguanidine hydrochloride, and
passed
over a nickel chelateaffinity matrix; and HIV-2 H6Rev
protein
was eluted from the column. The eluantcontaining
HIV-2 H6Rev wasdia-lyzed into
phosphate-buffered
salinecontaining
0.5 M NaCl and then filterconcentrated
(Amicon
Corp.).
Thepurified
HIV-2 H6Rev protein was
analyzed by
sodiumdodecyl
sulfate-polyacrylamide gel
electrophoresis.
In vitro transcription ofthe
Rev-responsive
elements. BL-RREplasmidswerelinearizedattheirXbaIsite and usedastemplatesfor T7 RNA polymerasein vitro. PCR T7 ampli-cons wereuseddirectly forT7transcription in vitro.The T7
transcription reactions were performed by using the
sug-gested conditions of the manufacturer (Promega Biotec, Madison, Wis.). The T7 transcripts were labeled by
incor-poration of[32P]UTP during transcription. DNA templates were digestedwithDNase, and unincorporated nucleotides were removed by using Sephadex G-50 spun columns. The
integrity ofthe labeledtranscriptswasconfirmedbyanalysis
on6% denaturing polyacrylamidegels.
Rev-RRERNAbinding assays. The conditions for assess-ment ofRev-RRE RNA interactions have been previously
described(3, 29). Briefly, Revproteinand
32P-labeled
RREtranscripts
were incubated at 30°C for 10 min in bindingbuffer (5 mM HEPES
[N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid]
[pH 7.0], 25 mM KCl, 2 mMMgCI2,
3.8%glycerol)which contained 10 ,LgoftRNAas a
nonspe-cific
competitor. Following binding,
7 U ofRNase T1 was added and reactions were incubated at 37°C for 10 min.Binding
reactionswereterminated and thenanalyzed by gelelectrophoresis
undernondenaturing
conditions in 0.5x Tris borate-EDTA buffer. Polyacrylamide(4%)
gels were usedfor
analysis
ofHIV-1 RRE-RNAcomplex formation.HIV-2 Rev-RRE complexes were analyzed on 5% gels, which allowedfor betterresolution ofthe RNA-protein complex. Gels were fixed in 10% aceticacid, dried,
and exposed forautoradiography
at-70°C.
Cell culture, transfections, and CAT assays. CHO cells
constitutively expressing
HIV-1 Tatprotein
(CHOZiptatiii)
(31) were maintained in Iscove modified Dulbecco medium
containing
antibioticsand10%fetal bovineserum. Transfec-tions were carried outby
using
a modified DEAE-dextran and calciumphosphate procedure (33).
Twenty-four
hourspriortotransfection,cells wereplatedat a
density
of2.5 x105in 35-mm
(diameter)
dishes andtransfected with250 ngofeach
plasmid
DNA. Cellswereharvested 48 hposttrans-fection,
and CAT assays wereperformed
aspreviously
described
(17).
RESULTS
Expression and
purification
of HIV-2 Revprotein.
Forexpression
of HIV-2Rev,
asynthetic
cDNA wascon-structed
by
using
the PCR method withproviral HIV-2RoD
DNA (see Materials and
Methods).
Theamplified
insertwhich was tailed with the
appropriate
restriction siteswasthenclonedintoa
eucaryotic
expression
vector.Theability
of this artificial HIV-2 Rev cDNA to
produce
a functional HIV-2 Revprotein
ineucaryotic
cellswasassessedby using
a
previously
describedRev-dependent heterologous
geneexpression
assay(32)
whichwasmodifiedtomeasureHIV-2 Rev function.Briefly,
HIV-1 env sequencesharboring
theregions
thatmediatenuclear retentionofviral mRNAaswell asthe RRE arepositioned
3' tothe termination codon andincorporated
into the 3'untranslatedportion
ofthebacterial CAT reporter genetranscript.
Theresult of this modification is that CAT geneexpression
becomessubject
toregulation
by
Rev.Expression
ofBLpSV-Rev,
encoding
the HIV-1 Revprotein, fully
restored CAT geneexpression
directedby
plasmid
pIlIAR,
which contains the HIV-1Rev-responsive
sequences
(Fig.
1).
Cotransfection withplasmid
pSVHIV-2Rev,
capable
ofexpressing
the HIV-2Revprotein,
didnot restoregeneexpression
directedby
pIlAR,
consistent withprevious
findings
showing
that HIV-2 Rev cannot functionthrough
the HIV-1 RRE(26).
However,
plasmid pIIIH2,
amodification of
pIIIAR
in which the HIV-1 RRE was re-VOL.64, 1990on November 10, 2019 by guest
http://jvi.asm.org/
4430 DILLON ET AL.
aAT-LJ i.t.
lXL CAT
-L
'Venj-----A
B
Riev
_- - 1 2 - 2 - 2
*
**
**4
*--...._... _ _ pl H2 pil
plJ31-111
pill AR plilH2 plllSIY2 H6 Rev
/1
4_, i-_11
1
FIG. 1. Functionalanalysisof theHIV-1,HIV-2,and SIV RRE
elements in aRev-dependent heterologous geneexpressionassay.
The plasmid pIlIAR contains theHIV-1 RRE andis shown
sche-maticallyatthetop.TheHIV-1RRE present inplasmidpIlIARwas
replaced with the HIV-2 and SIV RRE elements to generate the plasmids, pIIIH2 and pIIISIV, respectively. (A) Transfection of CHOZip,ta,Ii wasperformed in the absence (lanes -)orpresenceof
HIV-1Rev,pSV-Rev (lanes 1),orHIV-2Rev,pSVHIV-2Rev (lanes
2).(B) Comparison of transactivation with HIV-2 H6 Rev, pSVHIV-2H6Rev(lane H6Rev)(all other lanesarethesame asin panel A).
placed with the HIV-2 RRE, wasresponsive toboth HIV-1 and HIV-2 Rev (Fig. 1). Furthermore, both HIV-1 and HIV-2 Revproteins werecapable of restoringgene
expres-siondirectedbypIIISIV in which the SIV RRE replaced the HIV-1 RRE. In addition to showing that the HIV-2 Rev proteinis functional, this result demonstrates that both the HIV-2 and SIV RREs can work within the context of the HIV-1 repressive sequences which mediate the nuclear retention of structuralmRNA thatoccurs in the absence of Revexpression.
We have previously described a strategy for expression and purification of large quantities of HIV-1 Tat and Rev protein fromE. coli(4, 16). Briefly, asynthetic Rev cDNA
was constructed by the PCR method (34, 35) to contain codonspreferentially utilized inE.coli.This cDNAwasthen cloned into an E. coli expression vector (pDS [2]), which placed anoligonucleotide encoding six histidine residues at
the 5' end of the cDNA insert (4). Following induction of HIV-2H6Revexpression by the addition of 1mMIPTGto
logarithmically growingcultures, Rev proteinwasextracted
by guanidine hydrochloride lysis of theE. coliisolatesand purified by a single-column chromatography step with a metal chelate affinity resin that has selectivity for proteins
M
1
23
-!4.3-
-4 HIV-1 R, vHIV-2 Rev
FIG. 2. Purification of HIV-1 and HIV-2 RevproteinsfromE.
coli. HIV-1 Rev and HIV-2 H6 Rev proteins were purified as
described inMaterials and Methods. Proteins wereanalyzed ona
15% sodiumdodecyl sulfate-polyacrylamide gelandvisualized by
stainingwithCoomassie blue. Lanes:M,molecularweight markers; 1,HIV-2 H6 Revproteinstoredat4°Cfor 6weeks; 2,HIV-2 H6Rev
protein storedat-20°Cfor 2months; 3,purifiedHIV-1 Revprotein. kDa,Kilodaltons.
containing neighboring histidine residues (23). A relatively
purefractionofHIV-2 H6 Revproteinwasobtained(Fig. 2).
The function of the HIV-2 H6 Rev protein was first assessed in mammalian cellsasdescribed aboveby usingthe
eucaryotic expressionplasmid, pSVHIV-2H6Rev.The HIV-2 H6 Rev protein retained function (Fig. 1B), although its activity was somewhat lower than that obtained with the
unmodified Rev, which is consistent with previous results obtained with a modified HIV-1 H6 Rev protein (4). It is
unclear ifthe loweractivity of HIV-2 H6 Rev is duetothe amino-terminal modification of theproteinorlower levels of H6 Rev expressionin transfection.
Interactionof HIV-2Rev with HIV-2 RREandHIV-1RRE RNAs. Toexamine Rev-RRE RNAinteractions, theHIV-2
envgeneregion containing theputative HIV-2RRE (nucle-otides 7608 to 7973) fromHIV-2RODwasamplified by PCR
and cloned downstream ofthe T7 promoter present in the Bluescript transcription vector. 32P-labeled transcriptswere
generated in vitro with T7 RNA polymerase. Aqualitative gelshiftassay wasused inwhich RNAswereincubated with
purifiedRevprotein in the presenceofnonspecific compet-itortRNA, digestedwith RNase T1, and then analyzed on
nondenaturingpolyacrylamide gels (see Materials and Meth-ods).RRE RNAwhich formscomplexes with Revproteinis protected from nuclease digestion, whereas those RNAs which fail to bind Rev protein are digested to small frag-ments and oligonucleotides. The HIV-2 H6 Rev protein boundtoboth theHIV-2 RRE RNAandRNAcorresponding to thenucleotides present in the putative SIV RRE (nucle-otides 8086 to 8454) (Fig. 3). The HIV-2 H6 Rev protein-RRE RNAcomplexwasvisualizedas aband migratingnear
the top of the gel. In some experiments, a second,
much-faster-migrating bandwas present; however, this bandwas
Ii'
a 4L lw
0..
I
0
.t
;d1
..
_....__..._...,,
,~~~~~~~~~~i
J. VIROL.
.--..:
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.323.557.79.315.2]FUNCTION OF HIV REV PROTEINS 4431
cG
UCG
C G
A A
A A
A C AAG
J U U
C A A ;:
GGG
G;CA A
JCGA C
;LC A
AGuAG AG
A
A c
GGU;CAAAA
U
CAA
A
LoIB
Ln XI-L
wF C: t:Oo
N:- Nv
tor z km nw) n J I 4 u e ir
St+-+-+.+.+-+.+
.+.+.+ -+-+-+
-+
FIG. 3. Complex formation of HIV-1 and HIV-2 Rev proteins
with the HIV-1, HIV-2, and SIV RRE elements and deletion
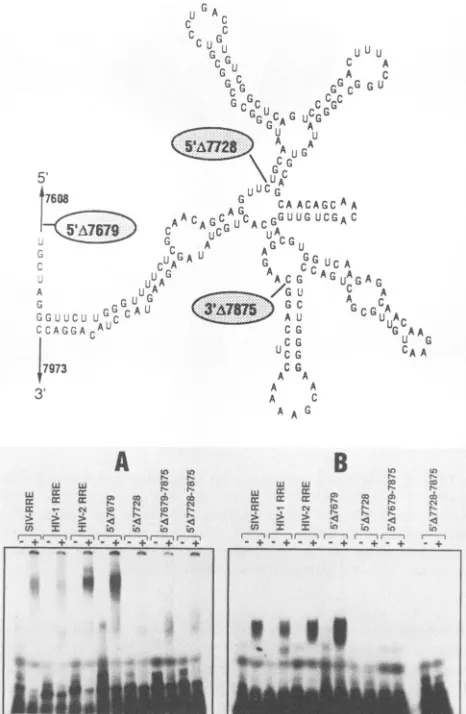
analysisof the HIV-2RRE.Thecomputer-predictedstructureofthe HIV-2RREis shown schematically atthe top, with the nucleotide positions ofthe deletions. 32P-labeled RRE RNAwas generated in
vitro by using T7 polymerase. RRE RNA was incubated with
purified HIV-2 H6 Rev or HIV-1 Rev proteins, and binding was
assessedby the qualitative nuclease resistance gelretardationassay.
Rev protein-RRE RNA interaction resulted in anucleaseprotected
complexwhichmigratedslower than RNaseTl-digestedproductsin
the nondenaturing polyacrylamide gel. (A)Complexformation with
HIV-2Rev protein and RRE RNA analyzedon a5% polyacrylamide
gel. (B) Complex formation withHIV-1 Rev proteinas analyzedon
a4%polyacrylamide gel. RRE RNAwasincubated intheabsence
(lanes -)orpresence(lanes +)of Revproteins.
most likely a partial RNase T1 digest product, since it was
alsoseentoalesser extentin the absence of HIV-2 H6 Rev protein. To rule out the possibility that contaminating bac-terial proteins from the HIV-2 H6 Rev protein preparation boundtothe RRE RNA,acrudelysatefromnontransformed E.coliwasusedin the gelshiftassay, andno bindingtoRRE
RNA was observed (data not shown), indicating that the RREbinding shown in Fig. 3 was specific to the HIV-2 H6 Revprotein. Very weak, ifany, binding was observed with
the HIV-1 RRE RNA,although the HIV-1 Rev protein was
abletointeract with all three of the RRE elements (Fig. 3). This observation is consistent with the inability of HIV-2
Revto regulate HIV-1 gene expression in vivo.
Delineation of HIV-2 RRE sequences necessary for HIV-1
and -2 Rev interaction. Ina previous study with the HIV-1 RRE RNA, it was shown that formation of a large stem structure isrequiredfor HIV-1 Rev-RRE interactions(3). In
subsequent studies, the actual Rev binding sitewas mapped to oneof thesmallerhairpin loopstructures(22,28, 29). It is
likely that formation of the larger stem is necessary for generationof the smallerhairpinloop structures in the RRE,
includingthe one required for HIV-1 Rev interaction. Simi-lar to the HIV-1 RRE, the HIV-2 RRE contains a Simi-large stem structure,asindicatedby the computer-predicted secondary structure of the HIV-2 RRE RNA (26). The importance of thelarge stem found in the HIV-2 RRE was assessed by the PCR method to generate deletions from the 5' and 3' ends of the HIV-2 RRE. A 5' deletion (5'A7679) upstream from the
large stem allowed binding of both HIV-1 and HIV-2 Rev proteins (Fig. 3). Furthermore, a 247-nucleotide HIV-2 RRE containing nucleotides 7679 to 7926 (5'A7679-7926), which maintained an intact stem, was sufficient for Rev-RRE interaction (data not shown). However, both Rev proteins
failed tobind HIV-2 RRE transcripts containing deletions of
either the upstream (5'A7728) or the downstream (5'A7679-7875) sequences which comprise the stem structure. In addition, deletion of both parts of the stem (5'A7728-7875)
failed to support Rev binding. These results indicate the importance of the HIV-2 RRE large stem in Rev binding, which is consistent withobservations reported for the HIV-1 RRE(3). Introduction of minor alterations (Fig. 4) consisting of five or six nucleotide changes had no dramatic effect on either HIV-1 or HIV-2 Rev interaction, with the exception of mutant PD-10, which failed to associate with either Rev protein. However, computer analysis of this mutation pre-dicts formation of an RRE with a markedly different RNA
secondary structure (data not shown).
Whereas major deletions in the stem abolished Rev
inter-action, small changes in base pairing within the stem struc-ture had little effect on Rev-RRE interactions. These data
indicatethat stem formation may be required for the overall
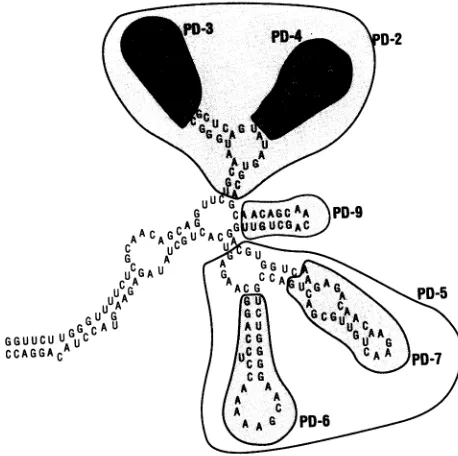
generation of the smaller secondary structures within the HIV-2 RRE shown in Fig. 5, of which one or more may be importantfor Rev interaction. This possibility was examined further by creating the mutations shown schematically in
Fig. 6and 7. The structures shown inFig. 6, 7, and 8 indicate the regions of the wild-type HIV-2 RRE affected by the mutations and are not intended to represent the actual RNA secondary structures formed by each mutated RRE.
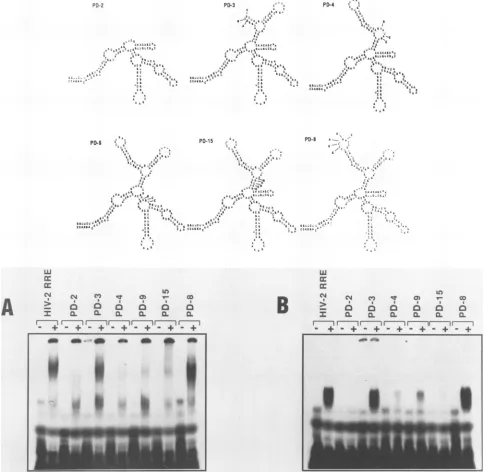
Dele-tion ofthe first two hairpin structures (as shown in PD-2) caused a loss of interaction of the HIV-2 RRE with either Rev protein. Deletion of thefirsthairpin alone (PD-3) did not affect RRE interaction with the Rev proteins. In addition, a changeof sequence in the loop of the first hairpin (PD-8)had noeffect on Rev-RRE interaction. However, deletion of the second hairpin (PD-4) resulted in complete loss of binding with HIV-2 Rev and a very weak interaction of the RRE with the HIV-1 Rev protein. Deletion of the third hairpin struc-ture (PD-9) gave substantially weaker interactions with both Rev proteins. A mutation (PD-15) which disrupted a stem structure between the second and third hairpins showed weak interaction with HIV-2 Rev and an extremely weak interaction with the HIV-1 Rev protein. Deletions of the fourth and fifth hairpins alone (PD-7 andPD-6, respectively) or together (PD-5) had no dramatic effect on interaction of the RRE with the HIV-2 Rev protein. However, deletion of both hairpins (PD-5) abolished HIV-1 Rev binding and deletionof the fourth hairpin alone(PD-7) resulted inamuch weaker interaction of the RRE RNA with the HIV-1 Rev protein. Inaddition, disruption ofa stemin the fourthhairpin
5,
*76O8
A L
3r
VOL.64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.72.305.77.434.2]u
A
A C
,GOG n U iUA_G, C
LUC A
A
:J aG
GG U 1 CLI
C G
A
A A
A C
A A G
a: D a:
X XcL a z X
a 7
XL :L
fl-.,. .- .i
0
I
FIG. 4. Analysis of the major stem involvement in Rev-RRE RNA complex formation. In vitro mutagenesis ofthe HIV-2 RRE
wasperformedas described in Materials and Methods. The
nucle-otidechangesareindicated in boldface in thediagramatthetop.(A) Complex formation of HIV-2 Rev protein with HIV-2 RRE
muta-tions.(B) Complex formation with HIV-1 Rev protein.
(PD-14) had no effect on either HIV-1 or -2 Rev protein binding.
Toexamine furtherthe importance of secondarystructure
asopposedtotheprimary nucleotidesequencefor Rev-RRE
interaction, single and compensatory mutations were made
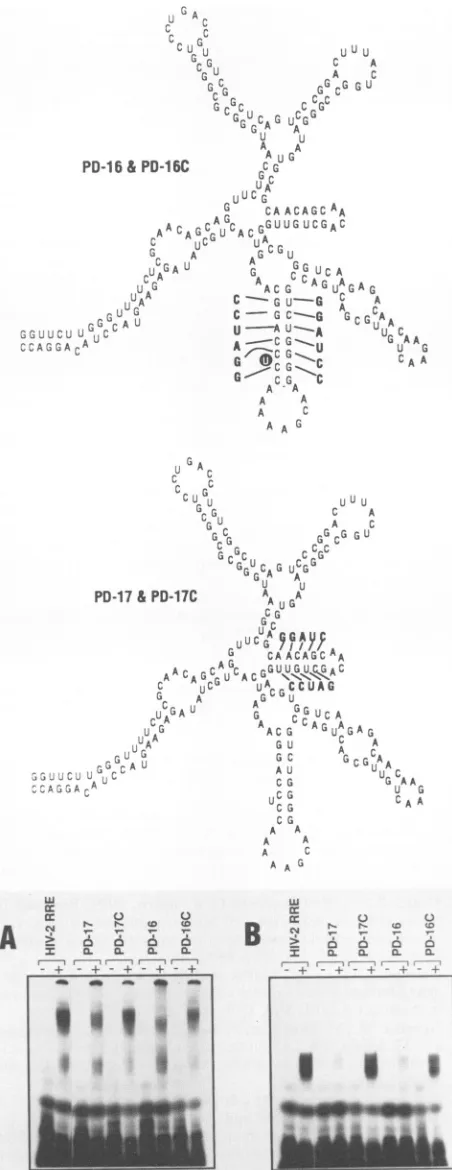
inthe third and fifthhairpin structures(Fig.7).Surprisingly, disruption of thestem structure in the third (PD-17) orfifth
(PD-16) hairpin causedasubstantial loss of interactionwith
HIV-1 but not HIV-2 Rev proteins. Restoration of HIV-1 Rev-RRE interaction was observed when compensatory
mutations were made which regenerated the base pairing necessary for stem formation in the hairpins (PD-17C and PD-16C).
Rev responseof the RRE mutations in vivo correlateswith in vitro Rev-RRE binding. The ability of the HIV-2 RRE mutants tofunction in vivo was examined by replacing the intact HIV-2 RRE present in plasmid pIIIH2 with the individualHIV-2 RRE mutations andmeasuring their ability
FIG. 5. Schematicrepresentationofhairpinloopdeletion
muta-tions in the HIV-2 RRE. Thecomputer-predicted structuresof the wild-type HIV-2 RRE RNAs affectedbyhairpin loopmutationsare
indicated by differential shading. The first through fifth hairpins referredtoin the textareclockwise fromPD-3(i.e.,thefirsthairpin
contains PD-3and fifthhairpin containsPD-6).
to respond to either HIV-1 orHIV-2 Rev incotransfection assays. The resultsobtained fromCAT assayswere scored aspositiveornegativefor Rev response andarepresentedin Table1. Ingeneral,RREmutantswhich didnotbindRev in thebindingassaysdidnotdisplayRevresponsesin vivo and thosemutationswhichinteractedweaklyfunctioned weakly
invivo.Examplesof thisweremutationsPD-15, PD-16,and PD-17, which bound weakly to HIV-1 Rev in vitro and showed weak responses in vivo. Interestingly, these same RREs interacted with HIV-2 Rev protein but failed to
function in vivo with HIV-2 Rev. Interestingly, mutations PD-3 and PD-5 showed weak or no response in vivo, although PD-3 bound well to both HIV-1 and HIV-2 Rev
proteins and PD-5 bound well to the HIV-2 Rev protein.
These observations are addressed further below. Signifi-cantly, the compensatory
mutations,
PD-16C and PD-17C,which restored secondary structure comparable to that of
the predicted wild-type HIV-2 RRE, responded to both HIV-1 and HIV-2 Rev proteins to the same extent as the
wild-typeRREitself.Therefore, as waspreviouslyobserved
with the HIV-1 RRE,maintenance of secondary structure is a major determinant for both Rev-RRE interaction and function.
DISCUSSION
The HIV Rev proteins, as well as the Rex proteins encoded by the human T-cell leukemia viruses (HTLVs), mediate the export of viral structural mRNAs from the nucleus tothe cytoplasm (11, 19-21, 24, 27). The ability of the HIV-1 Rev protein to interact directly with the RRE RNA(3, 6, 9, 27, 29, 39) isconsistent with the direct role of Revin the nuclear export of these mRNAs. Here we show that theHIV-2 Revproteininteracts with HIV-2 RRE RNA. Unlike the HIV-1 Rev protein, which can interact with the
-i~~~~~~~~~~~~~~~~~~
i.i9<w'* *
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.316.545.77.305.2] [image:5.612.54.294.79.487.2]FUNCTION OF HIV REV PROTEINS 4433
PD-2 PD-3
aai
C; r ;E t
_~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
A . C;AS_CfiCC£lS. a c
~~~~~~~~~~~~~~~~~~~~~~~A
r
A * t~~~~~~~~~~~~~~~~~~
-A S AffI,
A~~~~~~~~~~~~~~6 a a;
CC~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~I
m
w~~~~~~~~~~~~~~~~~w
C CrcC
~~~~~~~~~~~~~~~~~~~~~~~~AS BIL
A
a3
a a DmI
_,I_ w@,1 r ---CS.
, r
;
S Ca
ASC | eWt Ca
6,1 CCA
CF
PD-,
a6a
A
I:C
-cc
* r;LO
c co
a~~~~~~~-FIG. 6. Mutational analysis of the first three hairpins of HIV-2 RRE and Rev-RRE complex formation. Mutations are indicated
schematicallyonthe basis ofcomputer-predicted wild-type HIV-2 RRE RNA secondarystructure. These diagrams (Fig. 6-8) indicate the
structures affectedby the mutations andarenotintended torepresent actual RNA secondarystructuresformed by these mutations. Also
shown iscomplex formation of HIV-2 RREmutantswith HIV-2 H6 Revprotein (A) and HIV-1 Revprotein (B).
RRE of HIV-1 and HIV-2, significant binding of the HIV-2 Rev proteinwasonlyobserved with theHIV-2 RRE andto
a lesser extent with the SIV RRE. This observation is consistent withbiologicaldatashowingthat,whereasHIV-1 Rev can complement HIV-2 gene expression, HIV-2 Rev
onlyfunctions with its ownRRE and that of SIV.
Although manystudies have addressed the role of Rev in
the export of structural mRNAs from the nucleus to the cytoplasm, the mechanism for nuclear entrapment of these mRNAs, whichoccurs inthe absence of Rev expression, is
notunderstood.Similarly, althoughweknow thatsequences
present in both the env and gag-pol regions of HIV can
repress expression of heterologous genes when present in the 3'untranslatedregion,it isnotknownhow thisoccurs(8,
11, 13, 32). The studiespresented here do notaddress the mechanism for repression. However, we observed that replacement of the HIV-1 RRE with that of HIV-2 or SIV rendered the indicatorplasmid responsivetoboth HIV-1 and HIV-2 Rev proteins. Therefore, although the HIV-2 Rev proteincannotfunction with the HIV-1 RRE,inpartdue to its inability to bind, the interaction ofHIV-2 Rev with its
own RRE can overcome the repressive effects exerted by HIV-1 repressiveelements.
Inanefforttounderstand thenonreciprocal relationshipof
a
S
a aU'
I
-.5.
. )i..--.
VOL.64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.72.557.82.554.2]4434 DILLON ET AL.
PO-5
A
A
- A'C G
-A-P
CAC-"A ' C G
A~~~~~~~iA
PD-6
A
PO-7
CbricGC A
;A
CAG
CCL,G
C.AdACs C
CAAACCGA CA ACA' C
LCCCCA
G
CC r AC
C CCS,A CA G
.CACAA A)ACCC. CCAA
CI5
G~~~~~~u
CG.AA,ACCCACAAC
C A
A C C'
A G AA
G C w
cCAAC-CC AG C --G A Cl C ; AAwG> GAD A A
A C~ A
A,C~~~~~~~~~~~~~~~~~~~~~~~~~~~~~A
w
c:
cc
S *
-+o- +
ID r
S I I
- + _+ - +
_____
|.'K..>">-
-.s
i
.,X,,j,,.R
|
w
w...
w+.,, .x<+.
w .'
:_ 2{
a:
Ba:
C1 Ln rl (0
I I IS a
-
.r
+ M + + +c 6
CL CL
r r
-+ -+
FIG. 7. Mutationalanalysis of fourth and fifth hairpins of the HIV-2 RRE and Rev-RRE RNA complex formation. Mutations of HIV-2
RREareindicated schematically (the diagramsarenotintendedtorepresentactualRNAsecondarystructuresformed by these mutations). Alsoshown is complex formation of HIV-2 RREmutantswithHIV-2 H6 Revprotein(A)andHIV-1 Revprotein (B).
HIV-1 and HIV-2 Rev in binding the HIV-1 and HIV-2 RREs, mutational analysis and functionalassays were used
to identify and characterize the HIV-1 and HIV-2 Rev-binding sites within the HIV-2 RRE.Inarecentstudy, itwas
shown that binding of HIV-1 Rev to its own RRE is
determinedtoalargeextentbyformation ofasingle hairpin
loopstructurepresentwithin the RRE(22, 28, 29). Notably, altering the primary nucleotide sequence of the structure, whilemaintaining theproperRNAsecondarystructure,had littletonoeffectoneitherHIV-1Revinteractionorfunction
A
J. VIROL.
J- A_ MA
6A
-. L.
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTION OF HIV REV PROTEINS 4435
U;
A4
JG
ce
1- uUuu
G C A
u A C
GC GGCG GU
a-r
Gc cCG
GUC G CGG
GGA A`
U A A
PD-16&PD-16C C G
U A bU' G
G CAACAGCAA
~AG,.
GA ACAG uGU ACG UUGUCGAC
r, A G G U
C,~ A U
A A rUC
luA C-- Gu-G I A
CC,CGA~.,.,,,E E 6UALJ.-GACAAU CI GC---G GCGCAAU A
u
s sA U,a A
G--
CG
CA AG c AG
CG
~
CAA A A A
A C AA G
TABLE 1. Rev proteinbinding andfunctionalactivityofRREsa
Binding and function with:
RRE HIV-1 Rev HIV-2 Rev
Binding Function Binding Function
HIV-1 +++ +++ +/-
+/-HIV-2 +++ +++ +++ +++
SIV +++ +++ ++ ++
5'A7679 +++ +++ +++ +++
5'A7728 - - -
-5'A7679-7875 - - -
-5'A7728-7875 - NT - NT
PD-1 +++ +++ +++ +++
PD-10 - - +/-
-PD-11 + +/_ +
+/-PD-12 +++ +++ ++ +++
PD-2 - - - _
PD-3 +++ + +++
-PD-4 +/- +
-PD-9 +/- +/- +/- _
PD-15 +/- + +/-
-PD-8 +++ +++ +++ +++
PD-5 - - +++
PD-7 + ++ + +++
PD-6 ++ ++ +++ +++
PD-18 +++ +++ +++ +++
PD-14 ++ ++ ++ ++
PD-17 +/- + ++
-PD-17C +++ +++ +++ +++
PD-16 +/- + ++
-PD-16C +++ +++ +++ +++
PD-17& PD-17C
A'U
CG
UCG
CG
A A A A
A -A A G
C.)
CL N
r6.
Q.+
{D {D
6 6
0.0.
I
49I,P
9
w
cc cc
60.
rw
-
.t-aRev-RREbindingwasassayedbygelretardation analysis,andfunction
wasassessedby aRev-dependent heterologous gene expression assay (see text). Resultsareaveragesfrom several separate experiments: +++, full;
++, intermediate; +, weak; +/-, very weak;and -, no Revresponseor
interaction. NT,Nottested.
AG
through
theHIV-1
RRE(29).
The data obtained for theA HIV-2 RRE are consistentwith those observed for HIV-1.
cG AA Forexample,while nosingle stem-loopstructure was found
uGcA
U AG to be solely responsible for HIV-2 Rev-RRE RNAinterac-CAA tions,ourdata suggest that a groupofstructures mayconfer
the ability to bind. Consistent with observations for the HIV-1 RRE,alteration ofoneside of a stem structure shown to be required for Rev interaction prevented Rev binding.
However, generation of compensatory mutations in the same structure restored Revbinding and responsiveness in the functional assay. Thus, as was observed with HIV-1, D
6)
C: RNAsecondary
structureappearstobeamajor
determinantfor HIV-2 Rev-RRE interaction. Significantly, the region of --I the HIV-2 RRE RNA identified as being important for
interaction with HIV-1 Rev bears no resemblance with
regard to primary nucleotide sequence or secondary struc-ture tothe elementrequiredfor binding ofHIV-1 Rev in the g I, HIV-1 RRE. Moreover, asobserved with HIV-2 Rev bind-ing,
generation
ofalternate
secondary structures throughcompensatory
mutation restoredHIV-1
Revbinding.
Takentogether,
theseobservations offercompelling
evidencethat RNA secondary structure is a major determinant for both_I
HIV-1 and HIV-2 Rev binding and further suggest that FIG. 8. Analysisofcompensatory mutations ofHIV-2 RRE incomplex formation. Mutations ofHIV-2 RREare indicated sche-matically (the diagramsare notintended to represent actualRNA secondary structures formed by these mutations). Mutant PD-16 containsbasechangesindicatedontherightsideofthe fifthhairpin. The compensatorymutantPD-16C contains basechanges indicated
onboth sidesofthefifthhairpin.InPD-16C,theuridineresidue with
ablack background wasdeleted. MutantPD-17 contains the indi-catedbasechangesontheupper sideofthe thirdhairpin,whilethe compensatory mutant PD-17 contains changes on the upper and lower sides. Also shown is complex formation of HIV-2 RRE mutantswithHIV-2H6 Revprotein(A) and HIV-1Revprotein(B).
w cc rr
A
>I
_
_k-
Mdoll_
. 4P
VOL. 64, 1990
kG
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.68.294.78.663.2] [image:8.612.315.560.92.375.2]different secondary structures can be recognized by the HIV-1 Rev protein.
Since HIV-1 Rev can interact with the HIV-2 RRE, the remaining question concerns the inability of HIV-2 Rev to interact with the HIV-1 RRE.Althoughthe samesecondary
structuresarerequiredfor HIV-1 and HIV-2 Rev interaction
with the HIV-2 RRE, it appears that a smaller, more
well-defined structure is utilized for HIV-1 interaction with itsownRRE(22, 28, 29)which is insufficient forHIV-2 Rev protein binding.
Our mutational analysis of the secondary structures presentwithin the HIV-2 RRE indicates thatthe second and third hairpin structures are important for both HIV-1 and HIV-2 Revprotein interaction. Furthermore, itappearsthat the overall structure of the RRE RNA may be necessary
either forgeneration of the Rev protein-RRE RNA binding siteorforstabilityofacorrectcomplexformation necessary
for function. It could be postulated that binding of Rev protein tothe HIV-2 RREprovidesacomplex whichcanbe recognized byacellularfactor which facilitates transportof viral structuralmRNAs toacytoplasmicsite where transla-tionoccurs.Alternatively,Rev-RREinteractionmaycause a
conformationalchangein the mRNA which createsasite for interaction with cellular factor(s) that mediates nuclear
ex-port of structural mRNA. Either of thesepossibilities could explain the observation that mutations PD-3 and PD-5 are
capable ofbinding Rev but fail tofunction, presumably by formation of an inactive complex which is nonfunctional. Furthermore, the inabilityof HIV-2 Rev tointeract with the HIV-1 RRE may reflect the extent or type of secondary structuresthataregeneratedwithin the HIV-1 RRE. Indeed, only one stemstructure wasfound to berequiredforHIV-1
interaction with the HIV-1 RRE, while the data presented here demonstrate that interaction with the HIV-2 RRE is probably more complex and involves more than a single hairpin.
Our resultsmayalsoprovide insight relatingto theability of HTLV-I Rex protein tocomplement HIV-1gene
expres-sion versus the inability of HIV-1 Rev to complement HTLV-I(21, 30). Thepredictionis that HTLV-I Rex
recog-nizesanalternatesecondarystructurein the HIV-1 RRE and that a suitable binding site for HIV-1 Rev is absent in the HTLV-I RRE.
We propose that HIV Rev proteins and HTLV Rex proteins belong to a class of regulatory molecules that function through recognition of RNA secondary structure. Ourfindings would also be consistent withagroupofRNA secondarystructuresformingabindingdomain in which Rev
orRex would have severalcontact points.
ACKNOWLEDGMENTS
We thank Chein-Hwa Chen for providing purified HIV-1 Rev
protein. The following reagents were obtained through the AIDS
Research and Reference Reagent Program, Division of AIDS, NationalInstitute of Allergy and Infectious Diseases: reagent HIV-2ROD phage from contributor Ronald Desrosiers andreagent
pBK28-SIVfromcontributorJamesI. Mullins.
This work was supported in part with funds from a National
Cooperative Drug Discoverygrant.
LITERATURE CITED
1. Arya, S. K., C. Guo, S. F. Josephs, andF. Wong-Staal. 1985.. Trans-activatorgene of humanT-lymphotrophic virus typeIII (HTLV-III). Science 229:69-73.
2. Bujard, H., R.Gentz, M. Lanzer, D. Stueber, M. Mueller, I.
Imbrahimi, M. T. Haeuptley, and B. Dobberstein. 1987. A T5 promoter-based transcription-translationsystemfortheanalysis
ofproteinsin vitroand in vivo.Methods
Enzymol.
155:416-433. 3. Cochrane, A., C. H. Chen, and C. A. Rosen. 1990.Specific
interaction ofthe HIV revtransactivatorprotein withastruc-tured regionin the env mRNA. Proc. Natl. Acad. Sci. USA 87:1198-1201.
4. Cochrane, A. W., C. H. Chen, R. Kramer, L. Tomchak, and C.A. Rosen. 1989. Purification of
biologically
active human immunodeficiency virus rev protein from E. coli.Virology
173:335-337.
5. Cochrane,A. W.,A. Perkins, andC. A. Rosen. 1990. Identifi-cation ofsequences important in the nucleolar localization of human immunodeficiency virus Rev: relevance of nucleolar localizationtofunction.J. Virol. 64:881-885.
6. Daly, T., K. Cook, G. Gray,T. Maione, andJ. Rusche. 1989.
Specific binding of HIV-1 recombinant rev
protein
to the rev-responsiveelement in vitro. Nature (London)342:816-819. 7. Dayton, A. I., J. G. Sodroski, C. A. Rosen, W. C. Goh, and W. A. Haseltine. 1986. The transactivatorgene ofthe human T-celllymphotrophic virus type III isrequired
forreplication.
Cell 44:941-947.
8. Dayton, A. I., E. F. Terwilliger, J. Potz, M. Kowalski, J.G. Sodroski, and W. A. Haseltine. 1988.
Cis-acting
sequencesre-sponsive to the rev gene product of the human
immunodefi-ciencyvirus. J. AcquiredImmune Defic.
Syndr.
1:441-452. 9. Dayton,E.,D.Powell,and A.Dayton.1989. Functionalanalysis
of CAR, the target sequence for the rev
protein
of HIV-1. Science 246:1625-1629.10. Emerman, M.,M. Guyader,L. Montagnier,D. Baltimore,and M. Muesing. 1987. The
specificity
of the humanimmunodefi-ciencyvirustype2transactivator isdifferentfromthatofhuman
immunodeficiencyvirus type1. EMBO J. 6:3755-3760. 11. Emerman, M., R. Vazeux, and K. Peden. 1989. The revgene
productof thehuman
immunodeficiency
virusaffectsenvelope-specific RNAlocalization. Cell 57:1155-1165.
12. Feinberg,M.B.,R. F.Jarrett,A.Aldovini,R.C.Gallo,and F. Wong-Staal.1986.HTLV-III
expression
andproduction
involvecomplex regulation atthe levels of
splicing
and translation of viral RNA. Cell 46:807-817.13. Felber,B.K.,M.Hadzopoulou-Cladaras,C.Cladaras,T.
Cope-land,andG. N.Pavlakis.1989.Therev
protein
of HIV-1affects thestabilityandtransport ofthe viral mRNA. Proc.Natl.Acad. Sci.USA86:1495-1499.14. Feng, S.,and E. C. Holland. 1988. HIV-1 tat trans-activation
requires the loop sequence within tar. Nature (London) 334: 165-167.
15. Fisher, A. G., M. B. Feinberg, S.F. Josephs, M. E. Harper,
L. M. Marselle, G. Reyes, M. A. Gonda, A. Aldovini, C. De-bouk,R. C.Gallo,and F.Wong-Staal. 1986.The transactivator gene of HTLV-III is essential for virus
replication.
Nature(London)320:367-371.
16. Gentz, R., C. H. Chen, and C. A. Rosen. 1989. Bioassay for trans-activation using purified human immunodeficiency virus tat-encodedprotein: transactivationrequiresmRNA
synthesis.
Proc.Natl. Acad. Sci. USA86:821-824.17. Gorman,C. M.,L. F.
Moffat,
and B.Howard. 1982.Recombi-nantgenomeswhichexpresschloramphenicolacetyltransferase
in mammaliancells. Mol. Cell. Biol. 2:1044-1051.
18. Guyader,M.,M.Emerman,P.Sonigo,F.Clavel,L.Montagnier, and M. Alizon. 1987.Genomeorganizationandtransactivation of the humanimmunodeficiencyvirustype2.Nature(London)
326:662-669.
19. Hadzopoulou-Cladaras, M., B. K. Felber, C. Cladaras, A.A.
Athanassopoulos, A. Tse, and G. N. Pavlakis. 1989. The rev
(trslart) proteinof humanimmunodeficiencyvirustype1affects viral mRNA andprotein expressionviaacis-actingsequencein theenv region.J.Virol. 63:1265-1274.
20. Hammarskjold, M.-L., J. Heimer, B. Hammarskjold, I. Sang-wan, L. Albert, and D. Rekosh. 1989. Regulation of human
immunodeficiencyvirusenvexpression bytherevgeneproduct. J. Virol. 63:1959-1966.
21. Hanly,S.M.,L. T.Rimsky,M. H.Malim,J. J.Kim,J.Hauber, M. DucDodon,S.-Y. Le,J. V.Maizel, B. R.Cullen,and W. C. Greene. 1989. Comparative analysis of the HTLV-1 rex and
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTION OF HIV REV PROTEINS 4437 HIV-1 rev trans-regulatory proteins and their RNA response
elements. GenesDev. 3:1534-1544.
22. Heaphy, S., C. Dingwall,I.Ernberg, M. J. Gait, S. M. Green, J. Karn, A. D. Lowe, M. Singh, and M. A. Skinner. 1990. HIV-1 regulator of virion expression (Rev) protein binds to an RNA stem-loop structure located within the Rev response element. Cell60:685-693.
23. Hochuli, E., W. Bannwarth, H. Dobeli, R. Gentz, and D. Stuber. 1988.Geneticapproach tofacilitatepurification ofrecombinant proteins withanovel metal chelate adsorbent. Bio/Technology 6:1321-1325.
24. Itoh, M., J. Inoue, H. Toyoshima, T. Akizawa, M. Higashi, and M. Yoshida. 1989. HTLV-1 rex and HIV-1 rev act through similar mechanisms to relieve suppression of unspliced RNA expression. Oncogene4:1275-1279.
25. Kunkel, T. A. 1985. Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
26. Malim, M. H., S. Bohnlein, R. Fenrick, S.-Y. Le, J. V. Maizel, and B. R. Cullen. 1989. Functional comparison of the Rev transactivators encoded by different primate species immuno-deficiency virus species. Proc. Natl. Acad. Sci. USA 86:8222-8226.
27. Malim, M. H., J. Hauber, S.-Y. Le, J. V. Maizel, and B. R. Cullen. 1989. The HIV-1 rev transactivator acts through a structured target sequence to activate nuclear export of un-spliced viralmRNA. Nature(London)338:254-257.
28. Malim, M. H., L. S. Tiley, D. F. McCarn, J. R. Rusche, J. Hauber, and B. R. Cullen. 1990. HIV-1 structural gene expres-sionrequires bindingof the Rev transactivator toitsRNA target sequence.Cell60:675-683.
29. Olsen, H., P. Nelbock, A. Cochrane, and C. Rosen. 1990. Secondarystructureisthemajor determinant for interactionof HIV revprotein withRNA. Science247:845-848.
30. Rimsky, L., J. Hauber, M. Dukovich, M. H. Malim, A. Langlois, B.R.Cullen, and W. C. Greene. 1988. Functional replacement
ofthe HIV-1 rev protein by the HTLV-1 rex protein. Nature (London) 335:738-740.
31. Rosen, C. A., J. G. Sodroski, and W. A. Haseltine. 1985. The location ofcis-acting regulatory sequences in the human T cell lymphotropic virus type III (HTLV-III/LAV) long terminal repeat.Cell41:813-823.
32. Rosen, C. A., E. Terwilliger, A. Dayton, J. G. Sodroski, and W.A.Haseltine. 1988.Intrageniccis-actingartgene-responsive sequencesofthe human immunodeficiency virus. Proc. Natl. Acad. Sci. USA85:2071-2075.
33. Ruben, S., A. Perkins, R.Purcell,K.Joung, R. Sia, R.Burghoff, W. A. Haseltine, and C. A. Rosen. 1989. Structural and func-tional characterization ofhuman immunodeficiency virus tat protein.J. Virol. 63:1-8.
34. Saiki, R. K., D. H. Gelfand, S.Stofel,S. J.Scharf,R.Higuchi, G. T. Horn, K. B. Mullis, and H. A. Ehrlich. 1988. Primer-directedenzymaticamplification ofDNA withathermostabile DNApolymerase. Science239:487-491.
35. Saiki, R. K., S.Scharf, F. Faloona, K. B. Mullis, G. T. Horn, H.A. Ehrlich, and N. Arnheim. 1985. Enzymatic amplification of,-globin genomicsequencesand restriction site analysisfor diagnosisof sicklecell anemia. Science230:1350-1354. 36. Sodroski, J., W. C. Goh, C. Rosen, A. Dayton, E. Terwilliger,
andW. A.Haseltine.1986. Asecondpost-transcriptional trans-activatorgene required for the HTLV-III replication. Nature (London) 321:412-417.
37. Sodroski, J., R. Patarca, C. Rosen, F. Wong-Staal, and W. A. Haseltine. 1985. Location of thetrans-activating region onthe genomeofhumanT-cell lymphotrophic virustype III. Science 229:74-77.
38. Terwilliger,E.,R.Burghoff, R.Sia,J. Sodroski, W. Haseltine, and C. Rosen. 1988. The art gene product of the human immunodeficiency virus is required for replication. J. Virol. 62:655-658.
39. Zapp, M., and M. Green. 1989.Sequence-specificRNAbinding bythe HIV-1 revprotein. Nature(London) 342:714-716. VOL.64, 1990