0022-538X/92/095569-07$02.00/0
Copyright© 1992,AmericanSocietyfor
Microbiology
In
Vivo Binding
of
Wild-Type
and
Mutant
Human
Immunodeficiency
Virus
Type
1
Rev
Proteins:
Implications for Function
SALVATOREJ.ARRIGO,1* SHAUNHEAPHY,2 ANDJULIA K. HAINES'
Department of Microbiology and Immunology, MedicalUniversity ofSouth Carolina, Charleston,
South Carolina
29425-2230,1
andDepartment of Microbiology, University ofLeicesterSchoolofMedicine, Leicester, LEI9HNEngland2 Received 14 March 1992/Accepted 12 June 1992
The Rev transactivator protein of human immunodeficiency virus type 1 (HIV-1) is required for protein expression from theHIV-1 RNAswhich containabinding site forthe Rev protein,termed the Rev-responsive
element(RRE). This transactivatoractsbothatthe level of splicing/transportof nuclearRNAs andatthe level
oftranslation ofcytoplasmic RNAs. We useda monoclonal antibody specific fortheHIV-1 Rev protein to immunoprecipitate cellularextractsfromHIV-1-infectedand-transfectedcells. High levels of specific binding of
wild-type
Rev totheRRE-containing RNAswerefound incytoplasmic,
butnotnuclear,extractsfromthesecells. A Rev mutantwhichlacked both nuclear andcytoplasmic Rev function but retained RNA bindingin vivo wasgenerated.Thisbindingwasdetectablewithboth nuclear and cytoplasmicextracts.Theseresults verify the existence ofdirectbinding of RevtoHIV-1 RNAs in vivoand
conclusively
provethat binding of Rev is notsufficient fornuclearorcytoplasmic Rev function.The results alsosupportadirect role for Rev in thenuclear exportandtranslation ofHIV-1 RNAs.
Human immunodeficiency virus type (HIV-1) encodes a
regulatory transactivator, termed Rev, which is essential for viral replication (14, 31). The inability of the virus to replicate without Rev is in part due to a lack of viral
structural protein expression (14, 31). The Rev protein
appears to act at two levels in allowing the production of
structuralproteins.Itfunctionsatthe nuclearleveltoinhibit splicingorfacilitate the nuclearexport of HIV-1 RNAs (2, 13,16, 17, 24, 25),and itfunctionsatthecytoplasmic levelto permit the translation of HIV-1 RNAs (1, 4, 7). We have previously demonstrated that cytoplasmic accumulation of thesingly spliced Rev-responsiveelement(RRE)-containing HIV-1 RNAs encoding Vif, Vpr, Env, and Vpuwas
unaf-fectedby the presence orabsence ofRev; however, these RNAs were not translated owing to a defect in polysome
formation(1).Acis-actingelement found intheenvgeneof HIV-1,the RRE, has been shownto mediate both of these effects of Revonviral RNAs(2, 13, 16, 17, 24, 25).
The Rev protein localizes to the nucleus of transfected cells, andpurifiedRev proteinbinds with highaffinityto in vitro-synthesized RRE-containing RNA (6, 10, 15, 18, 27, 28, 30, 33). This binding is dependent on an extensive secondarystructurewithin the RRE. Site-directed mutagen-esis of the RRE has localized importantstructures involved in this binding and has shown that these structures are
importantin vivofor Rev function(6, 9-12, 19, 20, 22, 27, 29, 32). Site-directedmutagenesisof the Revprotein has
delin-eated domainsof Revimportant for RNAbinding,
multim-erization,and function(4, 21, 23, 26,28).These studies have
shown thatmutagenesisof thefunctional domain of Revcan
resultin mutants whichretain the in vitrobinding properties ofwild-typeRev butlack Rev function. These mutants have
*Corresponding author.
been shownto exerttransdominant repression ofwild-type Revprotein.
The binding of Rev to the RRE in vitro appears to
correlatewell with the function ofRev in vivo; therefore, the association of Rev withthe HIV-1RRE-containing RNAs in vivoisproposedtobeaprerequisite for Revfunction. Since Revlocalizes to and functions at the level of splicing and transport in the nucleus, Rev should initially interact with these RNAs in the nucleus. The binding of Rev to the RRE-containing RNAswould theneither inhibit the splicing
or facilitate the nuclearexportofthese RNAs. However, it is assumedthat binding of Rev isnotsufficient forfunction since transdominant mutants of Rev, which retain the in vitro binding and multimerization properties of wild-type Rev but donotretain invivofunction,have beengenerated (4, 21, 23, 27, 28). Since Rev has an effect on the transla-tional capacityof these RNAs in thecytoplasm, it seemed likely that Rev would continue its association with these RNAs afternuclear export.However,the associationof Rev with HIV-1 RNAs in vivo has notbeen established.
Using monoclonalantibodies raised against the Rev
pro-tein, we examined the in vivo association of Rev with the
HIV-1RRE-containingRNAs. We demonstratethat in HIV-1-infected and -transfected cells, Rev is found associated withhighlevels ofRRE-containingRNAs in thecytoplasm. This association isdependentonthepresenceofRev and the
RRE. The bulk of the wild-type Rev-RRE binding was
detected in the cytoplasmic, but not nuclear, fractions of
infected andtransfected cells. Resultswith anonfunctional Rev mutant, which retains RNA binding, indicate that in vivo binding of Rev is not sufficient for function. The
detection of nuclear bindingwith this mutant indicates that
thebindingof functionalRev toanRNA enhances itsexport from the nucleus. The continued association of Rev with
cytoplasmicRNAssuggeststhat Rev allowsthe association
of HIV-1RNAswith the translational machinery.
5569
on November 9, 2019 by guest
http://jvi.asm.org/
5570 ARRIGO ET AL.
MATERIALS AND
METHODS
RESULTSProviral constructs. The construction of
CSF/EBV-
andASal/EBV- has beenpreviouslydescribed(1).
TDBg/EBV-was generated by using polymerase chain reaction (PCR)
mutagenesis. The oligonucleotide primersTDBg andM112
were used first to amplify a PCR product containing the
desiredmutation.TheoligonucleotideprimersBamSandCR2
were used to amplify an overlapping PCR product. The
gel-purifiedPCRproductswerethenamplifiedwithBamSand M112oligonucleotideprimers.ThePCRproductwasdigested
with BamHIandXhoIand subclonedintopBluescript (Strat-agene).Thisfragmentwas thenisolated frompBluescriptand
cloned at these unique sites in the proviral construct. This
reconstructed a full-length provirus containing the desired
mutation.Thesequencesoftheseoligonucleotideprimersare
asfollows:TDBg,
5'-GCTACCACCGCTTGAGAGATCTAC
TCTFGAC-3'; M112,
5'-GCITACITTGTGATTGCTCCATG-3'; BamS,
5'-CACTCCTCAGGAGGGGATCC-3';
andCR2,5'-CTCTCAAGCGGTGGTAGCTG-3'.
Tissue culture, infections, and electroporation. Peripheral
blood lymphocytes were prepared by using lymphocyte separation medium (Organon Teknika). These were
main-tained for3daysinRPMI 1640 mediumsupplementedwith
20% fetal calfserum (GIBCO or Whittacre) and 0.5 mg of
phytohemagglutinin(Sigma) perml.Nonadherentcellswere
infectedwith HIV-1culture supematantsinthepresenceof
20 ,ugofPolybrenepermlfor 1 to 2h.Cells werepelletedby low-speed centrifugation and resuspended in RPMI 1640
mediumsupplementedwith20% fetalcalfserumand5 U of
humanrecombinantinterleukin-2(GIBCO) perml.
The 729 B-cell line was maintained in Iscove's medium
supplementedwith 10%fetalcalfserum. Cells were electro-porated as previously described (2). The wild-type,
Rev-mutant, and RRE-mutant DNAs refertoconstructs
previ-ously described as pYKJRCSF/EBV-, ASal/EBV-, and
ABX/EBV-,respectively (1).
Cellular fractionation and immunoprecipitation. Nuclear
and cytoplasmic fractionswere prepared as previously
de-scribed(2).Nuclearfractionswerepreparedfor
immunopre-cipitation by being treated in a mini-beadbeater (Biospec
Products)with0.1-mm glassbeadsfor 2min. The
superna-tantswerecentrifugedtoremovedebris.Then 100 to200 U
ofRNasin(PromegaBiotec)wasaddedtoeachextract,and
the extractswere aliquotedintothe appropriatenumber of
tubes. All tubes contained 40 to 50
,ul
ofa 50% slurry ofprotein A-Sepharose (Pharmacia) in Nonidet P-40 lysis
bufferand 5 p,gofrabbitanti-mouseimmunoglobulinG(18).
Controlimmunoprecipitationscontained 2 ,ugofanti-Act(a
cell surface antigen) or 1 ,ug of anti-albumin antibodies.
Specific immunoprecipitations of Rev-bound RNAs
con-tained 10 to 100 plof NR4/3C4.22 anti-Rev monoclonal antibodyculture supernatant (18). The sampleswere
incu-batedwithrockingat40Covernight.Theimmunoprecipitate
was preparedby low-speed centrifugation andfour washes ofthepelletwithNonidetP-40lysisbuffer.Transfectedcells
wereradiolabeled aspreviouslydescribed(1).Proteinswere
immunoprecipitated as above by using pooled serum from
patientswithAIDS (1).
RNApreparation andRNA PCRanalysis. RNAwas
pre-pared from the immunoprecipitates and
immunosuperna-tants aspreviouslydescribed (2). Yeast tRNA(40 p,g)was
added as carrier to each ofthe immunoprecipitates. RNA
PCR analysis of RNAswas performed as previously
de-scribed(3).
Rev
is bound in vivo tohigh
levels of HIV-1 RNA. Since Revhas
aneffect
at thecytoplasmic
levelon the translation ofRRE-containing
RNAs, weexpected
that Revbinding
tothese RNAs should
bedetectable in thecytoplasmic
fractionof HIV-1-infected
cells. All theRRE-containing
RNAsshould be
capable
ofbinding
Rev. These RNAs include thegag/pol, vif,
vpr, andenvlvpu2
RNAs. Incontrast,
thetat/rev RNA is
anHIV-1 RNA
that does not contain the RREand should not bind
Rev.Therefore,
weinitially
investigated
Rev-RRE
binding by using cytoplasmic
extracts fromphy-tohemagglutinin-stimulated
peripheral
bloodlymphocytes
infected with HIV-1.
Thecytoplasmic
extracts from twoseparate infections
ofperipheral
bloodlymphocytes
werealiquoted
andanalyzed
induplicate.
Monoclonal antibodiesgenerated
against the carboxyl terminus
of Rev were used inthe
immunoprecipitation
analysis
of these extracts. In this assay,the antibodies
are used toimmunoprecipitate
Revprotein,
afraction of which
might be bound
to RNAsthrough
the RRE. If the
carboxyl terminus
of the bound Revprotein
were
accessible to
theantibody
andbinding
of theantibody
did not
perturb the binding
of Rev to theRNA,
specific
immunoprecipitation
of Rev
and the bound RNA should bedetected. RNA
prepared from the immunoprecipitate
wasanalyzed
by using
aquantitative RNA
PCRprocedure
(1-3,
5)
to determine the
specific
immunoprecipitation
of
RRE-containing
RNAs
(Fig.
1A).
High
levels of theRRE-contain-ing vif
and
vprHIV-1 RNAs
werefound in theimmunopre-cipitate
only in the
presence ofanti-Rev antibodies and notwith the control antibodies.
Therefore,
it
appeared that
theanti-Rev monoclonal antibodies
were able torecognize
Revbound to
RRE-containing
RNAs in the
cytoplasm
of infectedcells. The
binding of the antibodies
did notperturb
theassociation of Rev with the
RRE-containing
RNAs.To determine the
specificity
of theimmunoprecipitation
and to determine the level of HIV-1
RNAs associated withRev,
weused anti-Rev antibodies in another
experiment
toimmunoprecipitate
acytoplasmic
extractof
anHIV-1-in-fected
peripheral
blood
lymphocyte culture.
In thisexperi-ment, RNA
wasprepared
from both
immunoprecipitate
andimmunosupernatant
fractions. The
amountof total
RNAprepared
from each fraction
wasanalyzed by
agarosegel
electrophoresis
(Fig.
1B). Although
high levels of
28S and18S rRNAs could be detected in the
immunosupernatant,
norRNAs
weredetected in the
immunoprecipitate.
An
HIV-1RNA which does
notcontain the
RRE,
tat/rev,
wasexam-ined
by
quantitative
RNA PCR
(Fig.
1B). This RNA
wasalmost undetectable in the
immunoprecipitate
and waspresent
atextremely
high
levels in the
immunosupernatant.
We estimate that less than 1% of this
RNA
wasnonspecifi-cally
immunoprecipitated
in this
experiment.
Incontrast,
analysis
of
RRE-containing
RNAs
demonstrated
thathigh
levels
(10
to50%)
of
vpr,env/vpu2, and full-length gag/pol
RNAs
werespecifically immunoprecipitated
in
thisexperi-ment.
Thus, binding
of Rev
toRNAs
appeared to bespecific
for RNAs which contained the RRE. These
resultsdemon-strated that Rev remains associated with
ahigh proportion ofthe
RRE-containing
RNAs after
transport of theseRNAs
from the nucleus
tothe
cytoplasm.
Thus theseresults
indicated that the
binding
of
Rev tothese
RNAs was aprolonged
association
and notmerely
a transientevent,
restricted
tothe nucleus.
If the antibodies
werebinding
toRevwhich
was bound tothe
RRE,
the
immunoprecipitation
of RNAs shouldbe
specific
for
RRE-containing
RNAs
andshould occur onlyin J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
IN VIVO Rev BINDING 5571
A INF 1 INF2 NF 1 INF2
-- vpr
-_N-- vif
B >
E-.i-- 28SrRNA
-..-W--18S
_tRNA
rRNAi
-**2- ---tat/rev
v.1
"O ....-* p
u.REV (.ACT
C: C: C
C: c: a:< a:CC
a ft- - - -w --*-
tat/rev
- - - - *
4o.
* _-m-a
gag/pol
*0S
-* ienvlvpu2-.s-gagpol
CLUn
a
FIG. 1. Detection of Rev bound to HIV-1 RNAs in vivo. (A) A total of 3 x
i07
peripheral blood lymphocytes stimulated for 3 days with phytohemagglutinin were infected with HIV-lJRcsF (7 ,ug of p24). Twoidentical infections (INF 1 and INF 2) were performed. At day 2 postinfection, the cells were harvested and cytoplasmic extracts were prepared. Immunoprecipitations were performed in duplicate with either anti-Rev or control (anti-ACT) antibodies. RNA was prepared from the immunoprecipitates, and RNA PCR analysis was performed with oligonucleotide primers specific for HIV-1 vif and vpr RNAs as previously described (3). (B) Infections, harvests, and immunoprecipitations were performed essentially as inpanel A. RNA was prepared from both the immunosupernatant (IMMSUP) and the immunoprecipitate (IMMPPT). Of the recovered RNA,15%o
was analyzed on a 1%agarose gel stained with ethidium bromide. RNA PCR analysis of HIV-1 RNAs was performed as in panel A. RNA standards (STDS) were made by sequential dilutions ofRNA from transfected or infected cells.thepresence of both an intact RRE and Rev. To determine whether these criteria were met, wetransfecteda B-lympho-blastoid cell line with a wild-type proviral construct, an RRE mutant, and a Rev mutant. The RRE mutant has been previously characterized in terms of its ability to still pro-duce a functional Revprotein andtruncated RNA species, deletedfor theRRE(1,2).The Rev mutant is a truncation of Rev whichremoves the activation domain and the antibody recognition site. Ourprevious resultshaveshownthat in this lymphoid cell type, the level of cytoplasmic HIV-1 full-length
gag/pol
RNA was decreased with both of these mutants (1, 2). Inaddition, thelevelof tat/rev RNA, which doesnotcontainthe RRE, was concomitantly increasedwith themutants; however, thelevels of theRRE-containing vif, vpr, and env/vpu2 RNAs were notaffectedbytheabsence of Rev or the RRE (1). Therefore we expected that, in this experiment, theseprevious results should be reflected inthe amount of nonspecific immunoprecipitation, since nonspe-cific immunoprecipitation should be independent of the presence of Rev or the RRE and constant within a given experiment. Cytoplasmic extracts from these cells were prepared and immunoprecipitated with anti-Rev antibodies or control antibodies. RNAextracted from the immunopre-cipitates was subjected to RNA PCR analysis (Fig. 2). Analysis of tat/rev RNA, which doesnot contain the RRE,- - - w * -- < vif
- - -*
-- I---
--*-env/vpu2a: Z cnco
i: < 06
I
FIG. 2. Detection of specificRevbinding requires Rev, the RRE, and anti-Revmonoclonal antibodies. Atotal of2 x 107cellsof a B-lymphoblastoid cell line (line 729)wereelectroporated with 100 ,gof the indicated DNA. At 2daysposttransfection, cytoplasmic extractswereprepared andimmunoprecipitationswereperformed essentiallyasinFig.1.Anti-human albumin antibodieswereusedas anegative control.RNA waspreparedfromtheimmunoprecipitates andanalyzedbyRNAPCRasinFig. 1.
revealed little or no difference between immunoprecipita-tions with anti-Rev or control antibodies of any of the constructswhichwere tested. Thisanalysis requiredalong exposure timetoenhance the detection of the low levels of nonspecific immunoprecipitation. As expected, the level of this RNA waselevatedcompared with thewild-type levelin all samples from the mutanttransfections. Analysisof full-lengthgag/pol RNA revealed that specific immunoprecipita-tion ofthis
RRE-containing
RNAwasdetectableonly in the transfections with the wild-type proviral construct. Asex-pected,
the level offull-length
gag/pol RNAwasdecreased in the mutantsamples compared withwild-type levels.
The vifandvprRNAsbothshowedspecific immunoprecipitation only when an intact RRE and Rev were present in the transfected construct. The level of these RNAswas similar inthewild-type controlsample,
themutantcontrol samples, andthemutantanti-Revsamples, indicating
thatthe nonspe-cificimmunoprecipitation
wasindependent
ofthe
presence of Rev or the RRE and the antibody used. These dataindicate
that,
invivo,
Rev isspecifically
bound to RNAs which containan intact RRE. The RREmutantproduces
a functionalRevprotein,
butnoincrease innonspecific
immu-noprecipitation
wasseenwith thismutantinthe presence of anti-Rev antibodies, eliminating the possibility of nonspe-cific interaction of Rev with RNAs.Consequently,
thespecific
immunoprecipitation
ofRRE-containing
HIV-1 RNAs from infected-cell extracts represents a biological association of Rev with HIV-1 RNAs in vivo which isabsolutely dependent
onthe presence of Rev and the RRE. Rev is bound to RNAs in thecytoplasm
but not in the nucleus. Since Rev exhibits aneffect
at the nuclear level(splicing
andtransport)
and has been showntolocalize
tothenucleus
(2, 8, 13, 15, 16, 17, 24, 25,
30), it is generally
accepted
that Rev mustinitially interact
withthe
RRE-containing
RNAs in thenucleus.Therefore,
weattempted
to VOL. 66, 1992on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.201.415.116.238.2] [image:3.612.204.411.494.618.2]5572 ARRIGO ET AL.
IMMPPT IMMSUP
C:
e
CE E EE
>. D )> > n >
<.:Ec: <: >.Er <: a:r£2 Er
Wild-Type
Rev
74 75 76 77 78 79 80 81 82 83 84
...GlnLeuPrProLeuGluArgLeuThrLeuAsp... ...CAGCUACCACCGCUUGAGAGACUUACUCUUGAC...
i. v .e
a#
w4
_. envvpu2-* vif
nuci cyto nucl cyto
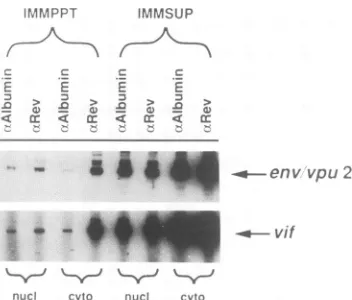
FIG. 3. Rev-RREbindingiscytoplasmicand not nuclear.
HIV-1-infected cells (asinFig. 1)wereharvested,and nuclear(nucl)and cytoplasmic(cyto)extractswereprepared.The fractionswerethen
immunoprecipitated, and RNAwaspreparedfrom the immunopre-cipitates (IMMPPT) and immunosupernatants (IMMSUP). This
RNAwassubjectedtoRNAPCRanalysisasinFig. 1.
detect Rev binding to RNAs in the nuclear extracts of infected cells. HIV-1 infected cells were separated into nuclear and cytoplasmic fractions and immunoprecipitated with anti-Revorcontrol antibodies. RNAwaspreparedfrom the immunoprecipitate and immunosupematant fractions andsubjectedtoRNA PCRanalysis (Fig. 3). High levelsof theRRE-containing env/vpu2andvifRNAswerespecifically immunoprecipitated from the cytoplasmic fraction of these cells.However, littleor noRevbindingwasdetectedinthe nuclear fraction. Thiswasreproduciblyobserved withcells infectedwith wild-type virus or transfected with the wild-typeproviralconstruct.These results indicated thatthebulk of the Rev-RREbinding,incells infectedortransfected with wild-type HIV-1, is in thecytoplasm.
A mutant which lacks Rev function. Owingto thenuclear localization of the Revprotein, itwasinitially expectedthat
Revbindingto RRE-containingRNAs should be detectable in the nucleus. Sincehighlevels ofcytoplasmicbindingand littleor nonuclearbindingwereobserved withthewild-type Revprotein, it seemed likelythatthis subcellular localiza-tionmightbe correlated with theinvolvement of Rev in the nuclear export of HIV-1 RNAs. If this were the case, a
nonfunctional Rev which retained RNA-binding capacity should bereadilydetectedboundto RRE-containing RNAs in the nucleus. Such a mutant would also provide direct evidence thatnuclearRevbinding exists andthatthebinding ofRevto anRNAinvivoisnotsufficient for Revfunction. Dissection ofthe Rev protein haselucidated RNA binding, proteinmultimerization, nuclearlocalization, and activation domains. Although the binding, multimerization, and local-izationdomains overlap, theactivation domainappearstobe separate. Mutants have been generated with mutations in thisdomain and showntoretainnuclearlocalizationandin vitrobindingandmultimerizationbuttobenonfunctional (4, 21, 23, 26, 28).Apoint mutant, TDBg/EBV-, in whichthe conserved leucine at amino acid81 of Rev is changedto a
serine,wasgenerated(Fig. 4).This mutation didnotalterthe aminoacidsequenceof the overlappingEnv reading frame. This leucine had been previously mutated to an alanine, abolishingRev function in fibroblast cells (26). The TDBg/ EBV- mutant preservedthe RNA-binding, protein multim-erization, nuclear localization, and antibody-binding do-mains ofthewild-type Rev protein.
TDBg Mutant
Rev
74 75 76 77 78 79 80 81 82 83 84
[image:4.612.98.274.78.228.2]...GlnLeuProP
roLeuGluArgSerThrLeuAsp...
...CAGCUACCACCGCUUGAGAGAUCUACUCUUGAC... FIG. 4. RNAandproteinsequencesofwild-type andmutant Rev constructs. Theaminoacidsequenceandcorrespondingnucleotide sequenceofRevbetweenresidues 74 and 84 isshown. Changesin nucleotide and amino acid sequencesbetween the wild type and mutant areunderlined.To determine whether the
TDBg/EBV-
mutant was de-fective for Revfunction,
we transfected it intolymphoid
cells in
parallel
withpreviously characterized
wild-type
and Rev mutant constructs(1). The cells
wereradiolabeled,
and HIV-1proteins
wereimmunoprecipitated and analyzed by
polyacrylamide gel
electrophoresis. The results
areshown
inFig. 5A.
Thewild-type
construct,CSF/EBV-, produced
high
levels ofp24 Gag,
p55 Gag
precursor,and
gpl20/160
Env. The
ASal/EBV-
Rev mutant did notproduce
detect-ablelevels
ofthese
proteins
but,
rather, produced
elevated levels of Nefcompared with the wild
type.These results
are consistent with ourpreviously published results with
these constructs(1). The
TDBg/EBV-
mutantproduced
aprotein
profile
indistinguishable from that of
ASal/EBV-, indicating
that the
TDBg/EBV-
mutantwasexhibiting
alack of
struc-tural
protein expression
and anincrease in Nef
production
characteristic
of aRev-
mutant.The
TDBg/EBV-
mutant wasalso
analyzed for
cytoplas-mic
RNAproduction.
Mutant andwild-type
constructsweretransfected into
lymphoid
cells, and
cytoplasmic
RNA wasprepared
48 hposttransfection.
This RNA
wassubjected
toquantitative RNA PCR
analysis
todetermine the level of
specific HIV-1 RNAs (Fig.
SB). The levels of unspliced
gag/pol
RNA
produced by
both Rev mutants werereduced
and the levels of
spliced
tat/revRNAproduced
by both Rev
mutantswereconcomitantly increased
comparedwith
wild-type
levels.
Thelevels
ofenv/vpu2
RNA weresimilar in
all constructs.These results
werein
agreementwith ourprevi-ous
results
(1) and
demonstrated that the nuclear
exportandsplicing function of Rev
wasabolished
inthe
TDBg/EBV-mutant. Although the level of cytoplasmicenv/vpu2
RNA wassimilar
tothat produced
by wild type, no Env protein wasdetectable
withthis
mutant.These results demonstratedthat this
mutant wasdeficient
in both the nuclear export/splicing
andtranslation functions of Rev.Rev binding is not sufficient for function. Although the
TDBg/EBV-
mutant produced a nonfunctional Rev, the mutantproteinretained
the RNA-binding, protein multimer-ization, nuclear localmultimer-ization, and antibody-binding domains of thewild-type
Revprotein. Functionally impaired mutants similar to theTDBg/EBV-
mutant have been shown to retain the binding properties of wild-type Rev in in vitrobinding
assays; however, it has not been demonstrated whetherthis binding
occursin vivo. To determine whether the mutant Rev protein was capable of binding in vivo to J.VIROL.ON IP
--W
on November 9, 2019 by guest
http://jvi.asm.org/
a)
CL
A
2a1
co > > tL -1
m
cc w w >- m0
0u
200i-- g
97.4
69
c c
E E
_ > : >
*-s---gaglpol
m u-0 vif
_- gpl20160
-a 0 0
-_-
env/vpu
2_ p55
46 _ ^
so_
- -.*-tat/rev30
--.
_4- Net
_-. p24
CL a
B -a
:
co > >D m
>- 0
*-gag/poi
so -4 tat/rev
o w_ _ onvlvpu2
FIG. 5. Effect of the Rev mutationon proteinandcytoplasmic RNA expression. (A) Lymphoid cells were transfected with the indicatedconstructs. At 48 hposttransfection, cellswere radiola-beled with35Stranslabel. Cytoplasmiclysateswere immunoprecip-itated with pooled serum from AIDS patients and analyzed by polyacrylamide gelelectrophoresis. (B) Lymphoidcellswere trans-fected with the indicated constructs. At 48 h posttransfection, cytoplasmicRNAwaspreparedandsubjectedtoRNA PCRanalysis for the indicated RNAs.
RRE-containing RNAs, wetransfected lymphoidcells with theTDBg/EBV- mutant. Nuclear andcytoplasmic extracts
were prepared and immunoprecipitated with anti-Rev or
control antibodies.RNAwaspreparedfrom the immunopre-cipitates and analyzed by RNA PCR for specific HIV-1 RNAs (Fig. 6). No specific immunoprecipitation oftat/rev
RNA was seen. In contrast,
gag/pol,
env/vpu2, and vifRNAswere all specifically immunoprecipitated bythe
anti-Rev antibodies, indicating high levels of binding of the
mutant Revprotein to the RRE-containing RNAs. As
op-nuci cyto
FIG. 6. Mutant Rev is bound to HIV-1 RNAs in the nucleus. Lymphoid cells were transfected with TDBg/EBV-. At 48 h post-transfection, cellswere harvested and nuclear (nucl) and cytoplas-mic(cyto) extracts were prepared. The fractions were immunopre-cipitated, and RNA was prepared from the immunoprecipitates. The RNAwassubjected to RNA PCR analysis as in Fig. 1.
posed
to results with the wild-type protein, binding wasreadily
detected in the nucleus as well as in the cytoplasm. This nuclearbinding was reproducibly seen in nuclear frac-tions from all experiments performed with this mutant.These
results show that the binding of the mutant Rev toRRE-containing
RNAsis both nuclear and cytoplasmic andthat this
binding is
notsufficient
for either nuclear orcytoplasmic
Revfunction.
DISCUSSION
Using
monoclonalantibodies
generated against the Revprotein,
wehave analyzed the association of Rev with HIV-1 RNAs invivo. Wehave demonstrated that these antibodies canbeusedtospecifically immunoprecipitate
HIV-1 RNAs which contain the RRE,depending on the presence of Rev, theRRE,
and anti-Rev antibodies. These RNAs include thoseencoding Gag, Pol, Vif,
Vpr, Vpu, and Env. These resultsindicate
that ahigh
level of theRRE-containing
RNAs
is associated
with Rev incytoplasmic
extractsfrominfected and
transfected cells and that Revis
capable
ofaprolonged association
with theRRE-containing
RNAs. Weconclude
from theseobservations
that Rev iscapable
ofmaintaining
itsassociation
with theRRE-containing
RNAs after their transport from the nuclear to thecytoplasmic
fraction. Since
thesecytoplasmic
RNAs areincapable
of translationin the absenceofRev(1),
theseresults suggesta direct rolefor Revin this process.The detection of both nuclear and
cytoplasmic
binding
with theTDBg/EBV-
mutanthasseveralimportant
implica-tions.First,
thebinding
ofRevtoRRE-containing
RNAsin vivoisnotsufficient
foreither the nuclearexport/splicing
ortranslational role of Rev.
Second,
the presence of nuclearbinding
with theTDBg
mutant and its absence with wild-type Revindicatesadirect role for Rev in thenuclearexport
of HIV-1 RNAs.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.385.494.77.293.2] [image:5.612.101.259.79.529.2]5574 ARRIGO ET AL.
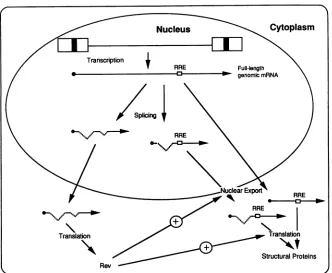
FIG. 7. Model forbimodal Rev function. Revwouldbe translated fromanRNA which is notdependenton Rev for itsexpression.Rev
would thenmigratetothenucleus,where it would bind toRRE-containingRNAs andpromotetheirexportfrom the nucleus. Revwould remain bound to the RNA during this process. In the cytoplasm, Rev would allow the association of the RNA with the translational
machinery.
The detectionofwild-type Revbindingin the cytoplasm, but not in the nucleus (although a large amount of RRE-containingRNAs ispresumablyavailable assubstrateinthe nucleus),of infected andtransfected cellscannotcompletely exclude a role for Rev in the inhibition of splicing of RRE-containingRNAs.However,the resultsarenotstrictly in agreement with what might be expected from a simple
blockto splicing byRev. If the roleof Revwere simplyto inhibitsplicing,onemightexpecttofindahighlevel of Rev bindingwiththe wild type in the nuclear fraction. TheRev mutantwould be expectedto have alower level ofnuclear Revbinding asaresult ofanincrease in the removal of the
RREand thebound Revproteinby splicing.It isformallya
possibilitythat Rev inhibits thesplicingofHIV-1RNAs and that these RNAs are concomitantly exported from the
nu-cleus;however, thesimplest interpretation ofthedata leads tomodel inwhich thebindingof Revto anRRE-containing
RNA in the nucleus enhances the nuclear export of that RNA (Fig. 7). In the cytoplasm, Rev would allow the interactionofthat RNA with thetranslationalmachinery. By interacting with the RRE-containing RNAs in the nucleus and continuing its associationin thecytoplasm, Rev could provide both nuclear and cytoplasmic functions. In the absence of Rev and its associated enhancement of nuclear export, complete splicing ofprecursorRNA would be
aug-mented. Intheabsence of Rev, unspliced (atareduced level)
andsingly spliced HIV-1 RNAs would still accumulate inthe cytoplasmthroughan alternate (slower)transportpathway. WithoutRev, these RNAs would be incapable ofassociating withthetranslational machinery. Thus Rev wouldbe
capa-bleofabimodal effecton nuclearexportandtranslation of RRE-containing RNAs. More research is requiredtoanalyze
the precise logistics of Rev-RRE binding and the cellular
processesinvolved.
ACKNOWLEDGMENTS
We thank A. Lowe and S. Green, Laboratory for Molecular Biology, Cambridge, England,forproductionof theantibodyNR4/ 3C4.22. Wealso thank Irvin S. Y.Chen,in whoselaboratorysome
of thebasic concepts for this studyevolved, and M. Schmidt, P. Arnaud,andK.Arrigoforhelpfuldiscussions.
S.H. is a Medical Research Council Senior AIDS Research Fellow. This workwas supported inpart by grant 001524-11-RG fromtheAmericanFoundationforAIDSResearch(toS.J.A.).
REFERENCES
1. Arrigo, S. J., andI. S.Y. Chen. 1991. Rev is necessary for
translation butnotcytoplasmicaccumulation of HIV-1vif,vpr, andenv/vpu-2RNAs. Genes Dev.5:808-819.
2. Arrgo, S.J., S.Weitsman, J.D. Rosenblatt, andI. S.Chen.
1989.Analysisofrevgenefunctiononhumanimmunodeficiency virustype1replicationinlymphoidcellsby usingaquantitative
polymerasechainreactionmethod. J.Virol. 63:4875-4881. 3. Arrigo, S.J., S.Weitsman,J. A.Zack, andI. S.Chen. 1990.
Characterization and expression of novel singly spliced RNA species of human immunodeficiency virus type 1. J. Virol. 64:4585-4588.
4. Benko, D.M., S. Schwartz, G.N.Paviakis,and B. K. Felber.
1990. A novel humanimmunodeficiency virustype 1 protein, tev,sharessequenceswithtat,env,andrevproteins. J. Virol. 64:2505-2518.
5. Cann,A.J., J.A. Zack,A. S.Go, S.J.Arrigo, Y. Koyanagi, P.L.Green,Y.Koyanagi, S. Pang, and I. S. Chen. 1990. Human immunodeficiency virustype 1T-celltropismis determinedby
eventspriortoprovirusformation. J.Virol. 64:4735-4742.
6. Cochrane,A.W., C. H.Chen,andC. A. Rosen. 1990. Specific
NuclearExport' RRE
RRE
0~~~~~~~
-.%.Translation
/
~ ~~~~
Structural ProteinsRev
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.149.481.72.345.2]interaction of the human immunodeficiency virus Rev protein with a structured region in the env mRNA. Proc. Natl. Acad. Sci. USA 87:1198-1202.
7. Cochrane, A. W., K. S.Jones, S. Beidas, P. J. Dillon, A. M. Skalka, and C. A. Rosen. 1991.Identification and characteriza-tion ofintragenic sequences which repress human immunodefi-ciency virus structural gene expression. J. Virol. 65:5305-5313. 8. Cullen, B. R., J. Hauber, K. Campbell, J. G. Sodroski, W. A. Haseltine, and C. A. Rosen.1988. Subcellular localization of the human immunodeficiency virus trans-acting art gene product. J. Virol. 62:2498-2501.
9. Daefler, S., M. E. Klotman, and S. F. Wong. 1990. Trans-activating rev protein of the humanimmunodeficiency virus 1 interacts directly and specifically with its target RNA. Proc. Natl. Acad. Sci. USA 87:4571-4575.
10. Daly, T. J., K. S. Cook, G. S. Gray, T. E.Maione, and J. R. Rusche. 1989. Specific binding of HIV-1 recombinant Rev proteintothe Rev-responsive elementinvitro.Nature (London) 342:816-819.
11. Dayton,E.T., D. M.Powell, and A.I. Dayton. 1989. Functional analysis of CAR, the target sequence for the Rev protein of HIV-1.Science 246:1625-1629.
12. Dillon, P. J., P. Nelbock, A. Perkins, and C. A. Rosen. 1990. Function of the human immunodeficiency virus types 1and 2 Revproteins is dependent on their ability to interact with a structuredregion present inenvgenemRNA.J. Virol. 64:4428-4437.
13. Emerman, M., R. Vazeux, and K. Peden. 1989. The rev gene product of the human immunodeficiency virus affects envelope-specificRNAlocalization. Cell 57:1155-1165.
14. Feinberg,M. B., R. F. Jarrett,A. Aidovini, R. C. Gallo,and S.F.Wong. 1986.HTLV-IIIexpression andproductioninvolve complexregulation at thelevels ofsplicingand translation of viral RNA. Cell 46:807-817.
15. Felber, B.K., C. M.Hadzopoulou, C. Cladaras,T. Copeland, and G. N. Pavlakis. 1989. rev protein of human immunodefi-ciency virus type1affects thestabilityandtransport of the viral mRNA.Proc.Natl.Acad. Sci. USA86:1495-1499.
16. Hadzopoulou, C. M.,B. K.Felber, C.Cladaras,A. Athanasso-poulos,A.Tse,andG.N.Pavlakis. 1989.Therev(trs/art)protein of human immunodeficiency virus type 1 affects viral mRNA and protein expression via a cis-acting sequence in the env
region. J. Virol. 63:1265-1274.
17. Hammarskjold, M. L., J. Heimer, B. Hammarskjold, I. Sang-wan, L. Albert, and D. Rekosh. 1989. Regulation of human immunodeficiency virusenvexpression bytherevgeneproduct. J. Virol.63:1959-1966.
18. Heaphy,S., C. Dingwall, I.Ernberg,M.J.Gait,S.M.Green, J. Karn, A.D.Lowe,M.Singh, andM.A.Skinner. 1990. HIV-1 regulator of virion expression (Rev) proteinbinds to an RNA stem-loop structure located within the Rev response element region. Cell 60:685-693.
19. Heaphy,S.,J.T.Finch,M.J.Gait, J.Karn,and M.
Singh.
1991. Human immunodeficiency virus type 1 regulator of virion expression,rev, formsnucleoproteinfilaments afterbindingtoapurine-rich "bubble" located within therev-responsive
region
of viral mRNAs. Proc.Natl. Acad. Sci. USA 88:7366-7370. 20. Holland, S. M., N. Ahmad, R. K. Maitra, P.
Wingfield,
and S.Venkatesan.1990. Human immunodeficiency virus Rev protein recognizes a targetsequence in Rev-responsive element RNA within the context of RNA secondary structure. J. Virol. 64:5966-5975.
21. Hope, T. J., D. McDonald, X. J. Huang, J. Low, and T. G. Parslow. 1990. Mutational analysis of the human immunodefi-ciency virus type 1 Revtransactivator: essential residues near the amino terminus. J. Virol.64:5360-5366.
22.
Kjems,
J., M. Brown, D. D. Chang, and P. A. Sharp. 1991. Structural analysis of the interaction between the human immu-nodeficiency virus Rev protein and the Rev response element. Proc.Natl. Acad. Sci. USA88:683-687.23. Malim, M. H., S.Bohnlein, J. Hauber, and B. R. Cullen. 1989. Functional dissection of the HIV-1 Rev trans-activator-deri-vation of a trans-dominant repressor of Rev function. Cell 58:205-214.
24. Malim, M. H., J. Hauber, R.Fenrick, and B. R. Cullen. 1988. Immunodeficiency virus rev trans-activator modulates the expression of the viral regulatory genes. Nature (London) 335:181-183.
25. Malim, M. H., J. Hauber, S. Y. Le, J. V. Maizel, and B. R. Cullen. 1989. The HIV-1 rev trans-activator acts through a structured target sequence to activate nuclear export of un-spliced viralmRNA.Nature(London) 338:254-257.
26. Malim, M. H., D. F. McCam,L. S. Tiley, and B. R. Cullen. 1991. Mutational definition of the human immunodeficiency virustype-1revactivationdomain.J.Virol. 65:4248-4254. 27. Malim, M. H., L. S. Tiley, D. F. McCarn, J. R. Rusche, J.
Hauber,and B. R.Cullen. 1990.HIV-1 structural gene expres-sion requires binding of the Rev trans-activator to its RNA targetsequence. Cell 60:675-683.
28. Olsen,H. S.,A. W. Cochrane, P. J. Dillon, C. M. Nalin, and C.A. Rosen.1990. Interaction of the humanimmunodeficiency virustype1Revprotein withastructuredregionin env mRNA isdependent onmultimer formation mediated throughabasic stretchof amino acids. Genes Dev. 4:1357-1364.
29. Olsen, H. S.,P. Nelbock, A. W. Cochrane, and C. A. Rosen. 1990.Secondarystructureis the major determinant for interac-tion ofHIV revproteinwith RNA. Science247:845-848. 30. Perkins, A.,A.W.Cochrane, S. M. Ruben, and C. A. Rosen.
1989. Structural and functional characterization of the human immunodeficiencyvirusrevprotein.J.AcquiredImmuneDefic. Syndr.2:256-263.
31. Sodroski, J.,W. C.Goh,C.Rosen,A. Dayton,E. Terwilliger, and W. Haseltine. 1986. A second
post-transcriptional
trans-activator generequiredforHTLV-IIIreplication.Nature
(Lon-don)321:412-417.
32. Solomin, L.,B. K. Felber,and G. N. Pavlakis. 1990. Different sites of interaction forRev, Tev, andRex
proteins
within the Rev-responsiveelement of humanimmunodeficiencyvirustype 1. J.Virol. 64:6010-6017.33. Zapp, M. L.,and M. R.Green. 1989.