0022-538X/88/103545-06$02.00/0
Copyright t 1988, American Society for Microbiology
Multiple Endogenous Xenotropic and Mink Cell Focus-Forming
Murine
Leukemia
Virus-Related
Transcripts Are Induced
by
Polyclonal Immune Activators
ARTHUR M.
KRIEG,'*
ARIFA S. KHAN,2ANDALFRED D. STEINBERG'CellularImmunologySection, NationalInstitute of Arthritis and Musculoskeletal and SkinDiseases,'andLaboratory of Molecular
Microbiology,
NationalInstituteof Allergy
andInfectious
Diseases,2
Bethesda,
Maryland
20892Received 28March1988/Accepted 10 June 1988
Northern(RNA) analyseswereusedtostudythekinetics of inductionof endogenous minkcellfocus-forming
(MCF) and xenotropic murine leukemiavirus(MuLV)-related sequencesinNFS and C57BL/6mice ijected with thepolyclonalimmune activatorslipopolysaccharide (LPS), concanavalin A, and 8-bromoguanosine. All three mitogens induced 8.4-, 7.2-, 3.0-, and 1.8-kilobase (kb) MCF-related transcripts coordinately in the spleens of injected mice. Xenotropic MuLV-related expressionwasalsorapidlyinduced in spleens by the three
polyclonal immune activators, but in a noncoordinate manner: a distinct set of transcripts with different kinetics of expressionwasinduced by each mitogen. MCF-related induction after LPS
injection
wasbothrapidandsustained;it began within 30 minandpersisted foratleast8days in thespleensof bothNFSandC57BL/ 6mice. LPS also caused prolonged induction of xenotropictranscriptsinspleensofC57BL/6butnotNFSmice. Thegld mutation, whichcausespolyclonal immuneactivation, induced 8.4-, 10.0-, and 13-kbMCF-related
transcripts in C3H/HeJ mice without altering expression of 7.2-, 5.6-, 4.0-, 3.0-, or 1.8-kb MCF-related
transcripts. The data demonstrate that individual endogenous MuLV-related transcripts can be induced
coordinately or independently and suggest that expression of these transcripts is linked to early stages of lymphocyte activation.
Infectious murine leukemia viruses (MuLVs) can be
di-videdinto three classes with different host ranges. Ecotropic
retrovirusesinfectonly mouse cells;xenotropicretroviruses
infect only nonmouse cells; and polytropic, or mink cell
focus-forming (MCF), viruses infect both mouse and non-mouse cells. Genomic line DNA from all inbred mouse
strains contains multiple copies of MuLV-relatedsequences,
the majority of which are replication defective (29). While infectious ecotropic and xenotropic retroviruses are present
in the germ lines of many inbred mice, infectious MCF MuLVsdonotpreexist in thegenome.Instead, MCF viruses arise via recombination between endogenous ecotropic,
xe-notropiclike, and MCF-relatedsequences(4, 14, 35).
Endog-enous MCF-related transcripts can be distinguished from recombinant MCFviruses; only the formerhave a
190-base-pair
(bp) cellular DNA insert in their long terminal repeat(LTR) (16).
Expression of endogenous MuLV-related sequences can
be induced by activation ofthe immune system. In vitro stimulationofspleen cells withtheB-cell mitogen
lipopoly-saccharide(LPS) inducesexpression of
xenotropic
virus andgp70
protein in many mouse strains (6, 24). This inductionwas detected inBcells after 2 to 3 days of stimulation and was blocked by preventing immunoglobulin M expression withanti-immunoglobulinMorbyinhibitingDNA
synthesis
(1, 31, 34). These in vitro studies
suggested
thatLPS-inducedxenotropic
expression
requires
B-cellproliferation
and differentiation. It is unknown whether MCF-related expression is also induced
by
B-cellmitogens.
T-cellmito-gens, such as concanavalin A
(ConA),
generally
have notbeen foundtoinduce
significant
retroviralexpression (1, 31),
*
Corresponding
author.although increased T-cell membrane MuLV-relatedgp70has
beenreported(37).
We have found a novel endogenous 8.4-kilobase (kb)
MCF-relatedtranscript which is
highly expressed
inautoim-munebut notnonautoimmunemousestrains(19).Polyclonal B-cell activation is a characteristic of autoimmune mouse
strains(10, 17; Y. Ishigatsubo, A. D. Steinberg, and D. M.
Klinman, Eur. J. Immunol., in press). To investigate the
relationship between immune activation and endogenous MuLV-related expression, we examined endogenous
xeno-tropicandMCF-relatedexpression in nonautoimmune mice injected withthe T-cellmitogen ConA and theB-cell
mito-gens LPSand8-bromoguanosine (8-BG). While LPSappears tobindtoamembranereceptor,8-BGactsintracellularlyto
induce polyclonal B-lymphocyte
proliferation
and differen-tiation(39). We alsostudiedtheeffects of thegldmutation,
whichinduces polyclonal B-cell activation and autoimmun-ity in C3H/HeJ mice (28; Ishigatsubo et al., in press), on MuLV-related expression.
MATERIALS ANDMETHODS
Mice.
C57BL/6,
BALB/c, C3H/HeJ,
andC3H/HeJ-gld/gld
micewereobtainedfromJacksonLaboratory (Bar
Harbor,
Maine),
and NFS mice were from the Frederick CancerResearch
Facility
(Frederick, Md.).
Polyclonalimmuneactivators.ConAtype
IV-S,
8-BG,
and Salmonella typhimurium LPS werepurchased
fromSigma
Chemical Co. (St. Louis, Mo.). Protein-free LPS was
gen-erously
provided by
D. Morrison(University
ofKansas,
Kansas City). All activators were diluted in sterilephos-phate-buffered saline and
injected
intraperitoneally
in a volume of0.1 ml.Probes and Northern (RNA) blot
hybridizations. MCFenv
and xenoenvoligonucleotide
probes
consistof 16 and 30bp
of the MCF andxenotropic
5' envregions,
respectively,
and 3545on November 10, 2019 by guest
http://jvi.asm.org/
hybridize specifically to endogenous MCF and xenotropic transcripts (15, 20). Transcriptsdetected with these probes are herein referred to as MCF-related and xenotropic tran-scripts, respectively. Mouse beta-2-microglobulin is a 0.6-kb cDNA generously provided by F. Mushinski (National
In-stitutes of Health). Glyceraldehyde-3-phosphate dehydroge-naseisarat1.3-kb cDNAobtained fromM. Piechaczyk(25).
LTR19J
(previously called B-34162 [15]) is an AluI-to-BglII fragment from the U3 region of the B-34 MuLV LTR andconsists of162bp ofthe 190-bpcellular DNA insert charac-teristicofendogenousMCF-relatedLTRs(16).Thepol gene
probe is a 1.4-kb XhoI-to-HpaII fragment of the AKR 623
ecotropic MuLVDNA, and the gag gene probe is a0.9-kb BglII-to-BamHI fragment ofAKR MCF 247 MuLV DNA (20). Ecotropic, xenotropic, and MCF MuLV total RNAs were includedwith each blot for molecularsize determina-tion andhybridization specificity (15).
Mouse tissues were homogenized with a polytron for 1 min athighspeedin 4 Mguanidinethiocyanate(5). RNAwas separated bycentrifugation througha5.7Mcesium chloride cushion, followed bysuspension and extractionwith phenol-chloroform (1:1) and chloroform. Poly(A)+ RNA was se-lected by passage over oligo(dT) columns (2). RNA was denatured in 50% formamide-1.3% formaldehyde-20 mM
MOPS
(morpholinepropanesulfonic
acid) (pH 7.0) at 55°Cfor15 min and thensize fractionatedina1%agarose-20mM
MOPS(pH7.0)-0.7% formaldehyde gel containingethidium
bromide. Custom-made longnarrowgel combswere
manu-factured by Bethesda Research Laboratories, Inc.
(Be-thesda, Md.). RNA was blotted onto nitrocellulose paper
(Schleicher & Schuell, Inc., Keene, N.H.), and complete
transfer was documented by UV photography before and
afterblotting. Blots probedwitholigonucleotide probeswere
prehybridized at 45°C in 50 mM Tris (pH
8)-10%
dextransulfate-1% sodium dodecyl sulfate (SDS)-1 mg of yeast RNA (type X-S; Sigma) per ml-1 M NaCl, hybridized at
45°C in the same mixture containing 3 x 106 cpm of
oligonucleotide end labeled with polynucleotide kinase (Pharmacia, Inc., Piscataway, N.J.) per ml and [-y-32P]ATP (>5,000Ci/mmol;AmershamCorp.,ArlingtonHeights, Ill.),
and washedat45°C for
MCFenv
or 42°C for xenoenv for 15 min in 6x SSC (lx is 0.15 M NaCl plus 0.015 M sodiumcitrate)-0.1%
SDS and for 30 min in lx SSC-1% SDS.Northern blothybridizationswithnonoligonucleotide probes
were performed by prehybridization at 42°C in 50% forma-mide-5x SSC-5x Denhardt solution-0.1%SDS,
hybridiza-tion at 42°C in 50% formamide-5x SSPE-1x Denhardt
solution-10% dextran sulfate-0.1%
SDS-106
cpm of[a-32P]dCTP
probe labeled with a random priming kit(Phar-macia) perml, and washing once at room temperature and once at55°Cin 2x SSC-0.1%SDSfor15min andfour times at55°C in0.2x SSC-0.1%SDSfor 15 min each. Stripping of
probes wasachieved by washing in 0.01x SSPE (lx is 0.15 MNaCl, 0.01 M sodium diphosphate, 0.001 M
EDTA)-0.1%
SDS at70°C for 10 to 30 min in an agitating water bath and wasconfirmed by autoradiography. Probes were purified by
centrifugation through G-25 Sephadex (oligonucleotides) or G-50Sephadex (nonoligonucleotides) columns from 5 Prime-3 Prime Inc. (Philadelphia, Paj. Autoradiography was per-formed at -70°C with Kodak XAR film in the presence of Cronex Lightning-Plus intensifying screens.
RESULTS
Kinetics of induction of endogenous MCF-related and xeno-tropic transcripts in NFS mice by LPS. NFS mice were
Oh 4h Sh 24h 72h 192h
U = 7.28.4Kb
4 * g -1 .8Kb
MCFenv
- 8.4Kb
xeno
env~~~~~~~~~~~..Q.
.0.-28S -I2.6Kb
* -1.8S
E -- 1.18Kb
GAPDH
h"
- 1.3KbFIG. 1. Induction ofspleen retroviralexpression by LPS. Four-month-old male NFS micewereinjectedintraperitoneally with 50jig
ofS.typhimurium LPS.Attheindicatedtimes, spleen tissues from twotofour micewereprocessed intoRNA.Total RNA(20 ,ug)was used forNorthern blotanalyses. The same blot was stripped and reprobed serially with
MCFen,,, xenoen,,,
and glyceraldehyde-3-phosphatedehydrogenase(GAPDH).injected intraperitoneally withLPS; atvarious times
after-ward, RNA was extracted from tissues and Northern blot
analyses were performed. Within 4 h ofLPS injection, the
spleen contained increasedamountsof7.2-,3.0-,and 1.8-kb MCF-related transcripts aswell as newly detectable 8.4-kb MCF-related RNA(Fig. 1).Allfourtranscriptswereinduced
within 2 h in a subsequent experiment (data not shown).
These four transcripts
hybridized
with theLTR19J
probe (data not shown) and therefore contained the 190-bpDNA sequences characteristic ofendogenous MCF-related LTRs (16). The 8.4-kb transcripts were no longer detectable 8 hafter injection, but the shorter transcripts continued to be
highly expressedforat least 8days (Fig. 1).
Induction of faint 8.4-kb spleen xenotropic RNA was
observed at 4 h, but not at later time points. A 2.6-kb transcriptwasdetectable from 4to24hafterLPSinjection,
anda 1.8-kb xenotropic transcriptwaspresentatrelatively
constant levelsthroughoutthe experiment (Fig. 1).
LPS injection also increased MCF-related RNA in the NFSmousethymus (Fig. 2). At 4h, expressionof 7.2- and
3.0-kb MCF-related sequences wasgreatlyincreased; how-ever,by24hthisexpressionhad returnedtobase line. There wasno apparentinduction of 8.4-kb MCF-related RNA nor ofxenotropic expression (Fig.2). Wecannotrule out adirect
effect of LPS on thymocytes; however, mediators of the
acute-phase response induced by LPS may well be respon-sible for theobservedthymic induction.
Liver 7.2- and 3.0-kb MCF-related transcripts were in-creased by 4 h, peaked at 8 h, and returned to base line within 24 h after LPS injection (Fig. 2). No 8.4-kb MCF-relatedtranscriptsweredetected. A 2.6-kb xenotropic tran-scriptwasinducedintheliverat 4and 8h after LPSinjection
(Fig.2).
ConA and 8-BG induce MuLV-relatedexpression in NFS mice. Intraperitoneal injection of the T-cell mitogen ConA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.328.556.76.302.2]THYMUS
Oh 4h 24h
Oh 0.5h lh 2h 4h 26h 96h 264h
LIVER
Oh 4h 8h 24h
-7.2Kb -3.0Kb
*i466 0a
-28S
* .. 4:. ..
xenoenv V
#;
ij $I
47:*'
V*
4
1.2
-2.6Kb
-18S
-1.8Kb
[image:3.612.57.300.73.293.2]GAPDH
-1.3KbFIG. 2. Induction of thymus andliver retroviral expression by LPS.Total thymusandliverRNAs(20
jig)
from thesameNFSmicestudied in Fig. 1 wereprobed with MCFenV, xenoenv, and
glyceral-dehyde-3-phosphate dehydrogenase(GAPDH). Time in hours after LPSinjectionis indicated above each lane.
FIG. 4. Retroviral induction in C57BL/6 mice by LPS. Four-month-old maleC57BL/6 micewereinjected intraperitoneallywith
50 p.gof LPS.At thetimesshown, total RNAwaspreparedfrom the
spleensofindividualmice, and20 p.g was used forNorthern blot analysis. The same blot was probed serially with MCFenv and xenoenv. Crosshybridization ofxenoenvwith rRNAwasreduced by
washingat 45°C. Ethidium bromide staining was used to confirm thatcomparableamountsofRNAwerepresentin all lanes.
Con
A
C
4h
8h
26h
8.4 Kb
_-7.2 Kb- ^
3.0Kb-
6
1.8
8.4 7.2
Kb-28S
3.0 2.6
Kb-
18S-1.8
Kb-ao
a AkAim
ow
6,0,
6-114
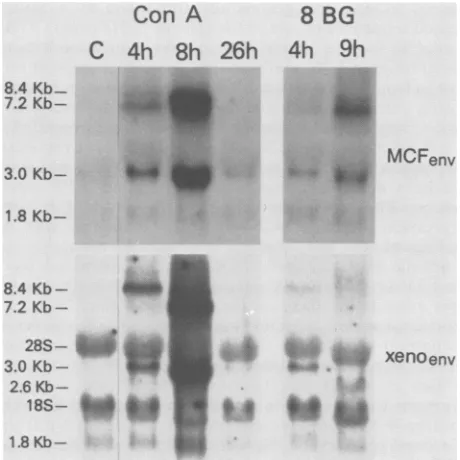
FIG. 3. Induction ofsplenicretroviral 8-BG. Five-month-old male NFS micewi
allywith100 p.gof ConAor75 p.gof8-BC ofhours after injection (the control, C, preparedfromspleentissues of individual forNorthern blotanalyses. All lanesareJ
wasprobedwithMCFenv,thenstrippeda
Ethidium bromidestainingwasusedtoc
ofRNAwereloadedineachlane.
ledtoarapid increase in splenic levels of 8.4-, 7.2-, 3.0-, and
1.8-kb MCF-related RNA (Fig. 3). In contrast to the
sus-8 BG tained increaseinsplenicMCF-relatedtranscriptsafter LPS
injection, levels ofMCF-related RNA returnedtobase line 4h 9h within 26 h after ConAinjection (Fig. 3).ConAinjectionalso caused marked induction of xenotropic RNA: 8.4- and 3.0-kb bands were seen at 4 and 8 h; large amounts ofa
7.2-kbtranscript werepresent at8 h (Fig. 3).
MCF-related induction after 8-BG injection of NFS mice MCFenv was similarto that seen afterConA or LPS: 8.4-, 7.2-, 3.0-,
and 1.8-kb transcripts appeared within 4 h (Fig. 3). At 4 h
after 8-BG injection, xenotropic 3.0-kb and faint 8.4-kb transcriptswereinduced;at9h, xenotropic 2.6-kb and faint 8.4-kbtranscriptswere detected(Fig. 3).
MCF-related andxenotropic induction by LPSinC57BL/6 mice. Within 30 min of LPS injection, C57BL/6 mice
ex-pressed increased amounts of 8.4-, 7.2-, 3.0-, and 1.8-kb MCF-relatedtranscripts (Fig. 4). Expressionof 8.4-kb MCF-related RNA peaked at 4 h and then decreased, while
xenoenv expression of the shorter transcripts peaked later and
con-ens tinuedata moreconstant level for the entire 11days ofthis experiment. Expression ofxenotropic 7.2- and 3.0-kb
tran-ak. M scripts was also increased by 30 min and persisted for 11
days (Fig. 4). Afaint 8.4-kbxenotropicbandwasdetectable
from 2 h to 11 days after LPS injection. There was no
apparent induction of1.8-kb xenotropicRNA. Inaseparate
expression byConA and experiment, 8.4-, 7.2-, 3.0-, and 1.8-kb MCF-related
tran-ereinjectedintraperitone- scripts and 2.6- and 8.4-kb xenotropic transcripts were
i.Attheindicated number increased in spleens of BALB/c mice within 2 h of LPS
imice,and 20,ugwasused injection(datanot shown).
fromthe same blot,which
gld
mutation induces several MCF-related transcripts.Thend
xenoeoo
reprobedwith . gld mutation arose spontaneously in C3H/HeJ mice; itonfirmthatequalamounts causes polyclonal B-cell activation with autoantibody pro-duction, splenomegaly, and massive lymphadenopathy (28;
MCFenv
MCFenv-8.4 Kb -7.2 Kb -3.0 Kb -1.8Kb
xenoenv
-8.4 Kb -7.2 Kb
-3.0 Kb -1.8 Kb
'.z
Ow* .0
W.
.k .... ..::'If I'o " r... go....w1w.
46
Ae.
.i la: :f .,XF
t 0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.316.560.75.293.2] [image:3.612.64.294.408.638.2]84b~
1/\:
8.4 Kb <^ 7.2 Kb -*
5.6
Kbb-gag.z
I 10.0 Kb --J 8.4 Kb
- 7.2 Kb
5.6 Kb r - 3.0 Kb \ - 1.8 Kb
pol
MCFenv
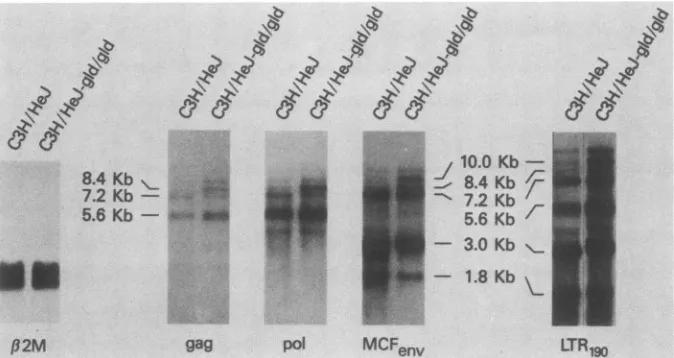
FIG. 5. EffectofgidmutationonMCF-relatedexpressionin C3H/HeJmice.Northernblotswereperformedon5p,gofspleen
poly(A)'
RNA from 5-month-old male mice. The filterwasprobed,stripped,andsequentially reprobedwithMCFenv,beta-2-microglobulin (,B2M), polgeneprobe,andgaggeneprobe. Aseparate blotwith thesameRNAwasprobedwithLTR190.
Ishigatsubo
etal., in press). To determine whetherconstitu-tive(asopposedtoexogenouslyinduced) polyclonal immune activation would alter endogenous MuLV-related
expres-sion, we studied
C3H/HeJ
andcongenic
C3H/HeJ-gld/gld
mice. Both strains had extremely low levels of
xenotropic
3.0- and 1.8-kb env-related transcripts and no
ecotropic
MuLV-related
expression (data
notshown).
Therefore,
anyotherbands detected with
MuLV-specific probes
should be transcribedfrom endogenous MCF-related sequences.Poly(A)+ RNA from
C3H/HeJ-gld/gld
spleens containedeight different
MuLV-relatedtranscripts,
five of whichwereexpressedatsimilar levels in C3H/HeJ
(Fig. 5;
datasumma-rized in Table 1). All
eight transcripts hybridized
withLTR19J,
aprobespecific
for the 190-bpsequence character-isticof endogenous MCF-related LTRs, thusconfirming
the MCF relatedness of thesetranscripts.
The8.4-,
10.0-,
and 13-kb MCF-related transcripts were induced inC3H/HeJ-gid/gid
and faintly detected in C3H/HeJ (Fig.5,
farright
panel). Many different subgenomic MuLV-related
tran-scripts have been reported previously
(15,
19, 20, 22), but supergenomic transcripts have not. Non-MuLVsuperge-nomictranscripts have been reported (8, 12).
Hybridization of 8.4-, 10.0-, and 13-kb MCF-related RNAs
to
MCFenv,
LTR190,
andpol andgagMuLV-specific probessuggested that these transcripts have complete retroviral
genomes
(Fig.
5;
Table 1). The 7.2-kbRNAdoesnotcontainpl5E
sequences and could therefore be distinguished fromthelarger transcripts which do (datanotshown). TABLE 1. Reactivityof MuLV DNAprobes with
C3H/HeJ-gld/gldRNA
Transcript Probe
size(kb) gag pol env
LTR1^
13 + + +
10 + + + +
8.4 + + + +
7.2 + + + +
5.6 + + - +
4.0 - + ? +
3.0 - - + +
1.8 - - + +
a?,
Possible faint band.C3H/HeJ mice haveamembrane defectresultingin
unre-sponsivenesstothemitogenic effects of LPS (11). To deter-mine whether this defect would prevent the LPS-induced increase in endogenous MuLV-related RNA, we injected C3H/HeJ mice withprotein-freeLPS. Therewas nochange inxenotropicorMCF-related RNA levels(datanotshown), demonstrating that the cellular pathways involved in the responsestoLPS andgidareatleastpartlyseparate.
DISCUSSION
The datapresented in thisreportdemonstratethat induc-tion ofmultiple endogenous xenotropic and MCF MuLV-relatedsequences occursinthespleens of NFSand C57BL/ 6 miceas soon as 30minafter invivostimulation with the B-cellmitogen LPS, implying thatthisinductionisanearly stepinlymphocyte activation. Previous in vitrostudies with spleen cells suggested that LPS induction of endogenous typeC retroviruses occurred after 2days and depended on B-cell differentiation(1, 34). Those studies, however, exam-inedproduction of gp7O proteinorinfectious virus and would not have detected induction of RNA alone. LPS rapidly induces RNA in spleen cells in vitro; within 1 h c-myc
expression increases markedly (13), compatible with our findings ofvery early changes ingeneexpression.
Splenic MuLV-related RNA remained increased for at least 11 days after LPS injection, whereas in thymus and liver tissues this increase lasted for less than 24 h. The protracted splenic inductionmayberelatedtothe increased proportion of spleen B cells in the S phase during the 2 weeks after LPS stimulation (26).
Hara et al. (7) reported that LPS induced liver gp7O synthesis within 4 h of injection inmousestrains with high basalgp7O levels, butnotinthose strains (suchasNFS) with low basalgp7O levels. We weretherefore surprised tofind NFS liver xenotropic and MCF-related RNA increased within 4 hafter LPSinjection.Presumably, these transcripts donotencodegp7O protein detectable by the radioimmuno-assayused in earlierstudies.
Full-length (8.4-kb) xenotropic and MCF-related tran-scriptswereinduced by mitogen in both NFS and C57BL/6 mice, but do not necessarily encode protein or infectious
virus. Indeed, there is no known endogenous infectious
MCFvirus,andtheinducible locusforinfectiousxenotropic
U.
P2M
LTR190
-b
4?
V
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.144.481.68.247.2]virus, Bxv-1, is present in C57BL/6 but not NFS mice (9, 18). While infectious virus is not induced by LPS in NFS mice, reverse transcriptase is, compatible with the expres-sion of endogenous defective retroviral sequences (33).
The regulation of expression of endogenous MuLV-re-lated sequences is largely unknown. The GV-1 locus is important in the coordinate regulation of several endogenous MuLV-related sequences (21). We observed coordinate in-duction of four MCF env-related transcripts (8.4, 7.2, 3.0, and 1.8 kb) by the mitogens LPS, ConA, and 8-BG. Contin-ued expression of these transcripts after LPS induction was somewhat noncoordinate: expression of the
8.4-kb
MCF-related transcript peaked early (within 4 h) and then de-creased, whereas the shorter transcripts continued to be highly expressed for more thana week in the spleens of both NFS and C57BL/6 mice. In contrast, induction of xenotropic expression was noncoordinate: in NFS mice, each mitogen induced a different combination of transcripts and had dif-ferent kinetics of expression. LPS induced xenotropic 8.4-and 2.6-kbtranscripts, ConA induced 8.4-, 7.2-, and 3.0-kb transcripts, and 8-BG induced 8.4-, 3.0-, and 2.6-kb xeno-tropic RNAs. These data suggest that multiple factors regu-late expression of endogenous MuLV-reregu-lated sequences. The various combinations of xenotropic transcripts induced by the three mitogens may reflect the different mechanisms ofmitogenesis.The mechanism by which mitogens induce MuLV-related transcripts is unclear and may differ for different transcripts or mouse strains. Some of the differences in expression between the different strains could be due to variations in their content of endogenous retroviruslike sequences. Al-though NFS mice have only one endogenous xenotropic env-related sequence (9), five different xenotropic env-re-lated transcripts were seen after mitogen induction (8.4, 7.2, 3.0, 2.6, and 1.8 kb). Possible explanations for this include mitogen-induced transcriptional readthrough, recombina-tional events, and changes in RNA splicing or processing. Variable RNA splicing has been reported in C3H/HeJ-gld/
gld
mice (30, 36) and could account for the additional high-molecular-weight MCF-related transcripts observed in those mice.Another possible mechanism of induction is suggested by the chromosomal location of MuLV-related sequences. Many xenotropic and MCF-related sequences are located near genes encoding lymphocyte antigens
(Thy-],
Ly-2, Lyb-2, Ly-6, Ly-7, Ly-10, Ly-22, and the immunoglobulin heavy chain and kappa light chain) (3, 27, 38). Mitogen-induced activation of these genes could secondarily result in induction of nearby MuLV-related sequences via a cis effect. Endogenous MuLV-related LTRs have promoter activity(23) and an enhancer region capable of binding to cellular trans-acting factors (20, 32). Therefore, activation of endog-enous LTRs by mitogen could cause transcription of nearby genes, possibly with important immunologiceffects.
The biologic significance of endogenous MuLV-related expression after polyclonal immune activation is unknown. We have recently reporteda novel 8.4-kb endogenous MCF-related transcript which is highly expressed in autoimmune but not nonautoimmune mouse strains (19). As discussed above, an 8.4-kb MCF-related transcript was induced in nonautoimmune mice by polyclonal immunestimulators, but its expression was briefer than that of the shorter transcripts, suggesting that the 8.4-kb RNA is subject todifferent regu-latory factors. Of course, it is unclear whetherthese different 8.4-kb MCF-related transcripts are transcribed from the same locus or are unrelated. It is possible that the
constitu-tive expression ofan 8.4-kb MCF-related transcript in
au-toimmune mouse strains contributes to the polyclonal
acti-vation which is a feature of their disease (10, 17).
Alternatively,
this expression may be a secondaryepiphe-nomenon. Studies of the regulation of expression of endog-enousretroviral sequences mayprovide insights into normal
cellular
activation mechanismsandabnormalities associatedwith autoimmune disease.
LITERATURECITED
1. Alberto, B. P., L. F. Callahan, and T. Pincus. 1982. Evidence that retrovirus expression in mouse spleen cells results from B-cell differentiation. J. Immunol. 129:2768-2772.
2. Aviv, H.,and P. Leder. 1972. Purification of biologically active globin messenger RNA by chromatography on oligothymidylic acid cellulose. Proc. Natl. Acad. Sci. USA 69:1408-1412. 3. Blatt,C., K.Mileham, M. Haas, M. N. Nesbitt, M. E. Harper,
and M.I.Simon. 1983. Chromosomal mapping of the mink cell focus-inducing and xenotropic env gene family in the mouse. Proc.Natl. Acad. Sci. USA 80:6298-6302.
4. Chattopadhyay, S. K., M. W. Cloyd, D. R. Linemeyer, M. R. Lander, E. Rands, and D. R. Lowy. 1982. Cellular origin and role of mink cell focus-forming viruses in murine thymic lympho-mas. Nature (London)295:25-31.
5. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J. Rutter. 1982. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
6. Greenberger, J. S., S. M. Phillips, J. R.Stephenson, andS. A. Aaronson. 1975. Induction of mouse type-C RNA virus by lipopolysaccharide. J. Immunol. 115:317-320.
7. Hara, I., S. Izui, P. J.McConahey, J. H. Elder, F. C. Jensen, and F. J. Dixon. 1981. Induction of high serum levels ofretroviral env geneproducts (gp7O) in mice by bacterial lipopolysaccha-ride. Proc. Natl. Acad. Sci. USA 78:4397-4401.
8. Herman, S. A., and J. M. Coffin. 1987. Efficient packaging of readthroughRNAinALV: implications for oncogene transduc-tion. Science 236:845-848.
9. Hoggan, M.D.,R. 0.O'Neill,andC.A.Kozak. 1986. Noneco-tropic murine leukemia viruses in BALB/c and NFS/N mice: characterization of the BALB/c Bxv-1 provirus and the single NFS endogenous xenotrope. J. Virol. 60:980-986.
10. Izui, S. P., P. J. McConahey, and F. J. Dixon. 1978. Increased spontaneous polyclonal activation of B lymphocytes in mice with spontaneous autoimmune disease. J. Immunol. 121:2213-2219.
11. Jakobovits, A., N. Sharon, and I. Zanbar. 1982. Acquisitionof mitogenic responsiveness by nonrespondinglymphocytes upon insertion of appropriate membrane components. J. Exp. Med. 156:1274-1279.
12. Kato, N., S.Pfeifer-Ohlson, M. Kato, E. Larsson, J. Rydnert, R. Ohlsson, and M. Cohen. 1987. Tissue-specific expression of human provirus ERV3 mRNA in human placenta: two of the three ERV3 mRNAs contain human cellular sequences. J. Virol. 61:2182-2191.
13. Kely, K., B. H. Cochran, C. D. Stiles, and P. Leder. 1983. Cell-specificregulation of the c-myc gene by lymphocyte mito-gens and platelet-derived growth factor. Cell 35:603-610. 14. Khan, A.S. 1984. Nucleotide sequence analysis establishes the
role of endogenous murine leukemia virus DNA segments in formation of recombinant mink cell focus-forming murine leu-kemia viruses. J. Virol. 50:864-871.
15. Khan, A. S., F. Laigret, and C. P. Rodi. 1987. Mink cell focus-forming (MCF)murine leukemiavirus-relatedtranscripts expressed in AKRmice. J. Virol. 61:876-882.
16. Khan, A. S., and M. A. Martin. 1983. Endogenous murine leukemia proviral long terminal repeats contain aunique 190-base-pair insert. Proc. Natl. Acad. Sci. USA 80:2699-2703. 17. Klinman, D. M., and A. D. Steinberg. 1987. Systemic
autoim-mune disease arises from polyclonal B cell activation. J. Exp. Med. 165:1755-1760.
18. Kozak, C. A., and W. P. Rowe. 1980. Genetic mapping of
on November 10, 2019 by guest
http://jvi.asm.org/
xenotropic murine leukemia virus-inducing loci in five mouse strains. J. Exp. Med. 152:219-228.
19. Krieg, A. M., A. D. Steinberg, and A. S. Khan. 1988. Increased expression of novel full-length endogenous MCF-related tran-script in autoimmune mouse strains. Virology 162:274-276. 20. Laigret, F., R. Repaske, K.Boulukos, A. B. Rabson, andA.S.
Khan. 1988. Potentialprogenitor sequences of mink cell focus-forming (MCF) murine leukemia viruses: ecotropic-, xenotro-pic-, and MCF-related viral RNAs are detected concurrently in thymus tissue of AKR mice. J. Virol. 62:376-386.
21. Levy, D. E., R. A. Lerner,and M. C. Wilson. 1985. The GV-1 locuscoordinately regulates the expression of multiple endoge-nousmurineretroviruses. Cell 41:289-299.
22. Levy, D. E., R. A. Lerner, and M. C. Wilson. 1985. Normal expression of polymorphic endogenous retroviral RNA contain-ing segments identical to mink cell focus-formcontain-ing virus. J. Virol. 56:691-700.
23. Levy,P.E., R. D. McKinnon, M. N. Brolaskis, J. W. Gautsch, and M. C. Wilson. 1987. The 3' long terminal repeat of a transcribed yet defective endogenous retroviral sequence is a competent promoterof transcription. J. Virol. 61:1261-1265. 24. Moroni, C., and G. Schumann. 1975. Lipopolysaccharide
in-ducesC-type virus in short term culture of BALB/c spleen cells. Nature (London) 254:60-61.
25. Piechaczyk, M., J. M. Blanchard, L. Marty, C. Dani, F. Pana-bieres, S. El Sabouty, P. Fort, and P. Jeanteur. 1984. Post-transcriptional regulation of glyceraldehyde-3-phosphate-dehy-drogenase gene expression in rat tissues. Nucleic AcidsRes. 12: 6951-6963.
26. Raveche, E. S., and A. D. Steinberg. 1983. Flow cytometric analysis ofin vivoactivation of murine spleen cells. J. Immu-nopharmacol. 4:163-181.
27. Rossomando, A., and D. Meruelo. 1986. Viral sequences are associated with many histocompatibility genes. Immunoge-netics 23:233-245.
28. Roths, J. B., E. D. Murphy, and E. M. Eicher. 1984. A new mutation, gid, that produces lymphoproliferation and
autoim-munity in C3H/HeJ mice.J.Exp. Med. 159:1-20.
29. Rowe, W. P. 1978. Leukemia virusgenomes in thechromosomal DNA of the mouse. Harvey Lect. 71:173-192.
30. Saga, Y., J.Tung,F. Shen,and E.A. Boyse. 1987. Alternative use of 5' exons inthe specification of Ly-5 isoforms distinguish-ing hematopoietic cell lineages.Proc.Natl. Acad. Sci. USA 84: 5364-5368.
31. Schumann, G., and C. Moroni. 1976. Mitogen induction of murine C-type viruses. I.Analysis of lymphoid cell subpopula-tions. J. Immunol. 116:1145-1150.
32. Speck, N. A., and D. Baltimore. 1987. Six distinct nuclear factors interact with the 75-base-pair repeat of the Moloney murine leukemia virus enhancer. Mol. Cell. Biol. 7:1101-1110. 33. Stoye, J. P.,andC.Moroni. 1984. Phenotypicmixing of retro-viruses inmitogen-stimulatedlymphocytes: analysis of xenotro-pic anddefective endogenous mouse viruses. J. Gen. Virol.65: 317-326.
34. Stoye, J. P.,and C. Moroni. 1985.Proliferation and differentia-tionrequirements for the induction oftworetroviral lociduring B-cell activation.J. Gen. Virol. 66:109-120.
35. Thomas, C. Y., andJ. M. Coffin. 1982. Genetic alterations of RNA leukemia viruses associated with the development of spontaneousthymic leukemia in AKR/J mice.J.Virol. 43:416-426.
36. Tung, J.,Y.Saga,and E. A.Boyse. 1987. Theincongruous Ly-5 phenotype oflprllpr and gidigid T cells. Immunogenetics 25: 126-129.
37. Wecker, E. 1977. Expression of endogenous viral antigens during immune response. Med. Microbiol. Immunol. 164:231-238.
38. Wejman, J.C.,B. A.Taylor,N. A.Jenkins,and N. G.Copeland. 1984. Endogenous xenotropic murine leukemia virus-related sequences maptochromosomal regions encodingmouse lym-phocyteantigens. J. Virol. 50:237-247.
39. Wolfert, R. L., M. G. Goodman, and W. G. Weigle. 1984. Demonstration ofactivation-specific interactions among B-lym-phocyte membrane proteins by a photoreactive cross-linking agent. J.Immunol. 136:2703-2708.