JOURNAL,OFVIROIOGY, Dec. 1993,P.7648-7653 0022-538X/93/127648-06$02.00/0
Copyright ©D 1993, American Societyfor Microbiology
Functional Interaction between Epstein-Barr Virus DNA
Polymerase Catalytic Subunit and Its Accessory
Subunit
In Vitro
TATSUYA TSURUMI,* TOHRU DAIKOKU, RYUTARO KURACHI, AND YUKIHIRO NISHIYAMA
Laboratory ofVirology, Research Institute for Disease Mechanism and Control, Nagoya University Schoolof Medicine, Tsurunta-cho, Showa-ku, Nagoya 466, Japan
Received 28 June 1993/Accepted 12 September 1993
The Epstein-Barr virus (EBV) DNA polymerasecatalytic subunit (BALF5 protein) and itsaccessory subunit (BMRF1 protein) have beenindependently overexpressed and purified (T.Tsurumi, A. Kobayashi, K. Tamai, T. Daikoku, R. Kurachi, and Y. Nishiyama, J. Virol. 67:4651-4658, 1993; T.Tsurumi, J. Virol. 67:1681-1687, 1993). In an investigation of the molecular basis of protein-protein interactions between the subunits of the
EBV DNApolymeraseholoenzyme,wecompared the DNA polymerase activity catalyzed by the BALF5 protein
inthepresence orabsence of the BMRF1 polymeraseaccessorysubunit in vitro. The DNApolymerase activity
ofthe BALF5 polymerase catalytic subunit alonewas sensitive to high ionic strength on an activated DNA template (80% inhibition at100 mM ammonium sulfate). Addition of the polymeraseaccessorysubunittothe reaction greatly enhanced DNA polymerase activity in the presence of high concentrations of ammonium
sulfate(10-fold stimulationat100 mM ammoniumsulfate). Optimal stimulationwasobtained when the molar ratio of BMRF1proteintoBALF5proteinwas2ormore.The DNA polymerase activity of the BALF5 protein
along with theBMRF1 proteinwasneutralizedbyamonoclonalantibodytothe BMRF1 protein,whereas that
of the BALF5 protein alone was not, suggesting a specific interaction between the BALF5 protein and the BMRF1 protein in the reaction. The processivity of nucleotide polymerization of the BALF5 polymerase catalytic subunitonsingly primed M13 single-stranded DNA circleswaslow(-50 nucleotides). Addition of the
BMRF1 polymerase accessory subunit resulted in a strikingly high processive mode of deoxynucleotide polymerization (>7,200 nucleotides). These findings stronglysuggestthat the BMRF1 polymeraseaccessory
subunit stabilizes interaction between the EBV DNA polymerase and primer template and functions as a sliding clampatthegrowing 3'-OH end of the primer terminustoincrease theprocessivityofpolymerization.
Epstein-Barr virus (EBV) is a human herpesvirus with a lineardouble-stranded DNA 172 kb long (1). EBV has botha latent state and a lytic replicative cycle in the nuclei of EBV-infected lymphoblastoid cells. During the latent phase of the EBV lifecycle, the EBVgenomeis maintainedasacircular plasmid molecule synthesized by host DNA polymerases. OriP,
onereplication origin of EBV, mediates thistypeofreplication (31). However, after induction of the lytic phase of viral replication, EBV replication proteins are induced and the EBVgenome is amplified 100- to 1,000-fold. The replication product isa head-to-tail concatemer, which issynthesized by the EBVDNApolymerase (Pol) viaarolling-circle mechanism initiated from the other replication origin, ori-Lyt (8). A number of features of EBV DNA replication make it an
attractive model system for the study of eukaryotic DNA replication. Our goal istounderstand the enzymaticprocesses
thatoccurduring the lytic phase of EBV DNA replication.
Ofthe approximately 84genesthat have been identified in
the genome, EBV encodes at least 7 viral genes that are
essential for ori-Lyt-dependent DNA replication (6). These
genes and their functions, which are known or have been predicted on the basis of sequence comparison with herpes simplex virus type 1, are BALF5, the DNA Pol catalytic subunit; BMRFI, the DNA Polaccessorysubunit; BALF2, the single-stranded DNA-binding protein; BBLF4 and BSLFI, the helicase and primase; BBLF2/3, a potential homolog of the
*Correspondingauthor.
third component of the helicase-primase complex; andgenes
encoded bythe EBVSall Ffragment, which hasanunknown function.
We have focusedoureffortsonthestudy of EBV DNA Pol.
The 110 kDa of the EBV DNA Pol catalytic polypeptide copurifies with the BMRF1 gene product (11, 15, 24). The
neutralization of EBV DNA Pol activity by a monoclonal antibodytothe BMRF1protein (2) and the low level of activity in the DNA Pol fraction lacking the BMRF1 protein (12) stronglysuggestthat the EBV DNA Polcatalytic subunit forms a tight complex with the BMRF1 protein in EBV-producing cells to function asthe Pol holoenzyme. The EBV DNA Pol
holoenzymepossessesbothDNAPol and 3'-to-5' exonuclease activities (24, 26). Furthermore, the EBV DNA Pol
holoen-zymeexhibits strikingly high processivity, which isadesirable
featureinthesynthesis of multiple copies of theEBVgenome
in rolling-circle DNA replication (25).
Ininvestigations of protein-protein interactions between the subunits of the EBV DNA Polholoenzyme, the overexpression and purification systems of the individual components have beendeveloped byusingthe baculovirusexpressionsystem(27, 29). The BMRF1 gene products expressed in insect cellsare made upoftwo phosphoproteins of 52 and 50 kDa and one unphosphorylated proteinof 48 kDa. The BMRFI DNA Pol accessory subunits exhibit higher binding affinity for
double-stranded DNA but have neither DNA Pol nor exonuclease
activity (27). The BALF5geneproductisaproteinof 110 kDa.
The BALF5DNAPolcatalytic subunit has been demonstrated to catalyzeboth DNA Pol and 3'-to-5' exonuclease activities
7648
Vol. 67, No. 12
on November 9, 2019 by guest
http://jvi.asm.org/
(29).
However,
thecatalytic
properties
of theEBV Pol catalyticsubunit aredifferent from thoseof theEBV DNA Pol
holoen-zymefromEBV-infectedcells in severalrespects (29),
suggest-ing thatthe BMRFI Pol accessory
protein
doesinfluence theenzymatic
properties
of the BALF5 Pol catalytic subunit. Inthis
study,
wecompared
the Polactivity
catalyzedby the EBVDNA Pol
catalytic
subunit in the presence or absence of the Polaccessoryprotein
on a shortorlong
single-stranded
DNAtemplate
to assess the contribution of the BMRFI Polacces-sory
protein
to thecatalytic efficiency
of the EBV DNA Pol holoenzyme.The EBV DNA Pol
catalytic
subunit was purified fromcytosolic
extracts of recombinant baculovirusAcBALF5-in-fected Sf9 cells
essentially
aspreviously
described (29). The EBV DNA Pol accessory subunit wasprepared
from nuclear extracts of recombinant baculovirus AcBMRFl-infected Sf9 cells aspreviously
described(27),
through
theheparin agarosecolumn
step.
It waspurified
further as follows. The heparin agarose column fraction of the BMRFIprotein
was loadedontoa
single-stranded
DNAagarose column equilibratedwithbuffer A
containing
20 mM Tris-HCI(pH
7.6), 1 mM EDTA, 25%glycerol,
mMdithiothreitol,
4 jigof leupeptin per ml,4jig of
pepstatin
A,
and mMphenylmethylsulfonyl
fluoride.The
single-stranded
DNA agarose column was eluted with a16-ml linear
gradient
from 0 to 0.6 M NaCl inbuffer A. TheBMRFl
protein
eluted at0.2 MNaClwaspooled
andstoredat-
80°C.
Thispreparation
wasnearly
homogeneous
and hadnodetectable DNA Pol or nuclease activity.
DNAPol
activity
wasroutinely
assayed
inareaction mixture(25
pLI)
thatcontained50 mMTris-HCI(pH
8.0); 10%glycerol; 6mMMgCl,;
100 jgof bovine serum albumin per ml; 80 jigof activated calf
thymus
DNA perml;
mM dithiothreitol;dATP,
dCTP,
and TFP at,u
25 M each, and[3H]dGTP
at 10 jiM(900
cpm/pmol).
The reaction was started by addition ofthe enzyme fraction, and incubation was for 20
min
at35°C.
Reactions werestopped by
addition ofIll
30 of 2 mg ofsonicatedsalmon sperm DNA per ml containing0.1 Msodium
PPiand
quenched
with 0.6ml of cold 10% trichloroacetic acidcontaining
0.1 M sodium PPi.Incorporation
oflabelednudle-otides into acid-insoluble material was measured in an Aloka
liquid scintillation counter.
BMRF1
Pol accessoryprotein
drastically
changes the saltsensitivity
of the DNA Polactivity
of the BALF5 proteinonanactivated DNA
template.
The DNA Pol activity of the EBVDNA Pol
holoenzyme
purified
fromEBV-producing
lympho-blastoid cells was stimulated
by
ammonium sulfate whenactivated DNAwas usedas the
template
primer
(11,
25). Thisfeature iscommonto DNAPolsin the
herpesvirus
family(11,
13,
25).
Insharp
contrast, the Polactivity
of the EBV Polcatalytic
subunitpurified
fromAcBALF5-infectedSf9 cellswasinhibited
by
the salt in adose-dependent
manner (29). Todetermine whether
complex
formation between theBALF5
and BMRFI
proteins
results inaquisition
of thesalt-stimula-tory property,
the saltconcentrationdependency
of DNA Polactivity
was determined afterpreincubation
with the purifiedBALF5 and
BMRF1
proteins
at0°C
for 20min
(Fig. 1). TheBMRF1
Pol accessoryprotein
alone did not exhibit DNA Polactivity
at any concentration of ammonium sulfate. Asre-ported
previously
(29),
theactivity
of thePol
catalyticsubunitalone was sensitive to the salt
(50%
inhibition at 50 mMammonium
sulfate).
However,
addition of the BMRFI Pol accessoryprotein
tothereactionresulted in adrasticchangeinsalt
sensitivity.
In the absence of ammonium sulfate, the BMRFI Pol accessoryprotein
inhibited Pol activity 13-fold.However,
withincreasing
concentrations of ammoniumsul-fate,
the Polactivity
catalyzed
by
theBALF5
Pol catalytic30
10
30
20
10
0
0 50 100 15U
(NH4)2S04 (MM)
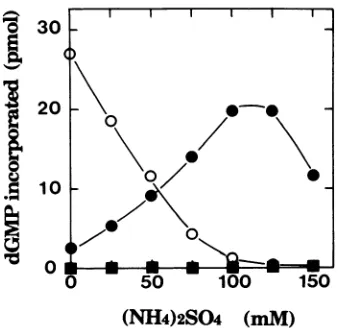
FIG. 1. Effect of ammonium sulfateonthe Pol activity catalyzedby
the BALF5 Pol
catalytic
subunit in the absence (0)andpresence(0)of the BMRFI Pol accessorysubunit. DNA Pol activitywasassayedby
using
an activated DNAtemplate
as described in Materials and Methods,exceptthatammoniumsulfate concentrationswerevariedasindicated. Prior to thestart of the reaction, the
BALF5
Pol catalytic subunit(50 ng)
and the BMRFI protein (50 ng) were mixed andpreincubated
O°C
at for 20min.
The same amount of the BALF5protein
alone(0)
or the BMRFIprotein
alone (U)wasalsopreincu-bated as described above. Datum
points
represent averages ofdupli-cate assays.
subunit
along
with the BMRFIprotein
was stimulatedgradu-ally
and maximum stimulation wasobserved at around 100 to 125 mM ammonium sulfate. At 100 mM ammonium sulfate,the BMRFI
protein
enhanced Polactivity
more than 10-foldcompared
with that of thePolcatalytic
subunit alone.Thus, thesalt-stimulatory
property
of EBV DNA Pol was recovered bypreincubation
of both subunits in vitro.Stoichiometric levels of
BMRF1
protein
with respect toBALF5
protein
can stimulate DNA Pol activity. Todeterminethe molar ratio of BMRFI
protein
toBALF5
protein which canenhance EBVDNAPolactivity
maximally,Polassayswereperformed
with an activated calfthymus
DNA template in thepresence of 100 mM ammonium sulfate
(Fig.
2). Ratedeter-mination was based on 20
min
of incubation under reactionconditions inwhich Pol
activity
was linearwith time foratleast40
min
(data
notshown).
A constant amount ofBALF5
protein
and the amounts ofBMRFI
Pol accessory proteinindicated in
Fig.
2 werepreincubated
for 20min
at0°C,
andthen the Pol assays were
performed.
Substoichiometric levels of BMRFIprotein
with respect toBALF5
protein caused adose-dependent
increase in DNAsynthesis.
Maximalstimula-tion occurred at molar
BMRF1/BALF5
protein
ratios of 2or more. Further addition of theBMRF1
protein
did notchangethe maximum
activity.
Thus,
the interaction of the Pol acces-soryprotein
with the Polcatalytic
subunit was apparently stoichiometric.Anti-BMRF1
protein MAb
neutralizes the Pol activitycat-alyzed by
the EBV DNA Polholoenzyme
reconstitutedinvitro. Monoclonalantibody (MAb)
9240(Dupont,
NEN) is ananti-BMRFl
protein-specific
MAb(19,
21) and is known toneutralize the DNA Pol
activity
of EBV DNA Pol purifiedfrom
EBV-producing
lymphoblastoid
cells(2).
TheEBVDNA Polcatalytic
subunit and the same amount of the BMRFI Pol accessory subunit were mixed andpreincubated
at0°C
for 10min.
Theamountsof MAb 9240 indicated inFig. 3were added to the mixture andfurther incubated at0°C
for20min.
The0
0
10
l
l '
po
E
_on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.352.523.73.237.2]7650 NOTES
100 4 < 80
360
0
<! 40
20
0
4
6
8
20
e
10
*0
0
mol
BMlRF1
protein
/ mol BALF5
protein
FIG. 2. Stimulation of the DNA Pol activity of the BALF5 Pol catalytic subunit by theBMRF1Polaccessorysubunit. Polassays were performedon anactivatedDNAtemplate inthepresenceof100 mM ammonium sulfate. The BALF5 protein (50 ng) and the indicated amountsof theBMRF1 proteinweremixed andpreincubatedat0°C
for 20 min, and then thereactionswerecarriedoutfor 20 minat35°C asdescribed in Materials and Methods. Acid-insoluble dGMP incor-poration was determined. Symbols: 0, reactions performed in the presenceof the BALF5 protein;0,reactionsperformed in the absence of the BALF5protein.Datum pointsrepresent averagesofduplicate assays.
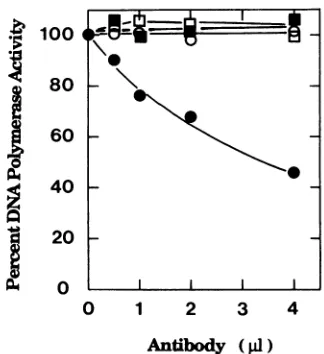
DNAPol reactionswerecarriedoutin the presenceof100 mM
ammonium sulfateat35°C for20 min.TheDNAPol assaysfor theEBV DNAPol catalytic subunit alonewere performed in the absence ofsalt. The results are
depicted
inFig.
3. MAb 9240 neutralized the DNAsynthesis catalyzed by theBALF5 Polcatalyticsubunit in the presenceof the BMRFI protein buthad noeffect onthat ofthe EBV Polcatalytic subunit alone. Theeffect of an anti-BBLF4 protein-specific antibody
(29)
onEBV DNA Pol activity was also examined as a control. The anti-BBLF4 protein antibody did not affect the Pol
activity
catalyzed by the BALF5proteininthepresence orabsenceof the BMRF1 Pol accessoryprotein.
Specificinteraction between the Polcatalyticsubunit and its accessory subunit. To determine whether the effect of the
EBV BMRF1 protein on DNA Polactivity is
specific
for theEBV BALF5 Pol catalytic subunit or not, the
ability
of the BMRF1 proteintoinfluence theactivities of other DNA Pols was examined. With activated calf thymus DNA as thetem-plate,wetestedthe effectof the BMRF1 proteinon twoother DNAPols: theEscherichiacoliPollKlenowfragment and T4 DNAPol (datanot
shown).
The Pol assayswereperformedin the presence or absence of 100 mMammonium sulfateunderthe reaction condition in which Polactivitywaslinear with time
and dose. Neitherstimulation nor inhibition of the Polactivity ofthe E. coli Poll Klenowfragment and T4 DNA Pol by the BMRFI protein wasobserved in the presence or absence of ammonium sulfate. These findings indicate that the BMRFI protein specifically interacts with theEBV DNAPol catalytic
subunit and rule out thepossibilityof anonspecific stimulatory
orinhibitory effectonthetemplate or other componentsofthe reaction mixture.
EBV DNA Pol accessory protein increases the polymeriza-tion rate and processivity of the Pol catalytic subunit. An
oligodeoxynucleotide with the sequence CACAATTCCACA CAAC, complementary to nucleotides 6170 to 6185 of
M13mpl8
single-strandedDNA(30)waspurchased from NewEngland BioLabs, Inc. To form singly primed M13
single-J.VIROL.
0 1 2 3 4
[image:3.612.354.516.71.248.2]Antibody (
p)
FIG. 3. Effect ofanti-BMRFI protein-specific MAbon the DNA Polactivity catalyzed by the BALF5protein in thepresence orabsence of theBMRF1 protein. We mixed the BALF5protein (50 ng)and the BMRF1 protein (50ng), preincubated themat 0°Cfor 10 min,and then added MAb 9240(R3mouse immunoglobulinGI MAbtoEBV EA-D [1'9, 21]; Dupont, NEN)at 100 ,ug/ml (circles)oranti-BBLF4
protein-specific rabbit immunoglobulin at 1 mg/ml (squares) and further incubated the mixture for 20 min at 0°C. Pol assays were performedon anactivated DNAtemplatein thepresenceof 100mM ammonium sulfate for 20 min at35C. The 100% value of the EBV DNAPolactivity in thepresenceof the BMRF1proteinwas20pmol of dGMPincorporatedper20min. Polassaysdonein the absenceof theBMRF1proteinweredonewithoutammonium sulfate. The 100% value of EBV DNA Polactivity in the absence of theBMRF1 protein was 27 pmol of dGMP incorporated per 20 min. Datum points represent averages of duplicate assays. Closed symbols represent reactions performed in the presence of the BMRF1 protein. Open symbols are reactions performed in the absence of the BMRFI
protein.
stranded DNA,the synthetic 16-mer DNAwasannealed at a molar ratio of 20:1 to M13mpl8
single-stranded
DNA in a buffer (20mMTris-HCt [pH 8.0],
5 mMMgCl2, 0.3MNaCI).
The hybridization mixture was incubated at 90°C for 5 min, allowedtocool to room temperaturefor1h, and incubated for
afurther 1 h at 30°C.The primed M13
single-stranded
DNA wasseparated fromexcessprimer bycentrifugation througha spuncolumn ofChromaSpin-100
(Clontech
Laboratory,Inc.).
To determine the function of the BMRF1 Pol accessory
proteinindeterminingthecatalytic
efficiency
ofthe EBV DNA Pol holoenzyme on the long single-stranded DNAtemplate,
the holoenzyme was reconstituted onto
singly primed
M13mpl8 single-strandedDNAcirclesby using stoichiometric levels of BALF5 protein along with BMRF1
protein.
DNAsynthetic processivitywasmeasuredby
using
thesingly primed
M13 template in an excessmolar ratio of
primer-template
to Pol,sothat each Pol molecule wasboundtoaprimer terminus.In thisassay, each Polmolecule catalyzes multiple rounds of synthesis, but the
probability
that aparticular primer
will be extended more than once is very low because of the lowPol-to-primer ratio that is maintained throughout the experi-ment.dATP anddGTP areneeded to prevent removal of the
DNA
primer
by the 3'-to-5' exonucleaseactivity of the BALF5 protein. Synchronous DNA synthesis by the reconstituted forms ofthe holoenzyme was initiated upon addition oftheremaining deoxynucleoside triphosphates, the reaction was
sampledatvarioustimes,DNAsynthesiswasquantitated (Fig. 4A),and DNAproductswereanalyzed in anativeagarosegel
410
0 0 07.
0
I-<
-0~~~~
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.69.291.76.231.2]i
0 10 20 30 40
Time (min)
BALF5 BALF5+BMRFI
2 5 10203040 2 510203040
-RFII
I I ii hi
Ad1gj5
BALF5 BALF5+BNRFI
I _.
5 10 20 3040 s7'a
n 1W !H !lf
~~~~~~~-
~~~~~~~~-655Z7
4356123222'
2D227 2 5 10 20 3040 2
so. *4
FIG. 4. Time course of DNA replication on M13
single-stranded DNA by the BALF5 Pol catalytic
presence (0) or absence (0) of the BMRFI Pol
The BALF5andBMRFIproteins (115 ngofeach)
at
O)C
for 20min
and then added to a reactionRI)
containing 50 mM Tris-HCI (pH 8.0), 3 mM MgCll. reitol, 100 pLgof bovineserum albumin perml, 10%
singly primed M13single-strandedDNA(8.3pmol
NaCI,
0.5 mM ATP, and 50,uM each dATPandincubated
O°C
at for5min. Tostart thereaction,,uMp.M [t_32PIdCTP(3,t)00 Ci/mmol) were added, and
reactionwasincubated at35°C. Samples (25
[LI)
times indicated and quenched byaddition to anequal Cc
sodium dodecyl sulfate-SO mM EDTA.After quenching,
weredivided intothreeparts.One wasanalyzed
measurement of the radioactivity incorporated
material (A). The second was used for analysis
products by electrophoresis inneutral 0.7%agarose
wasused for analysisofthereplication products
an alkaline 1.2c agarose gel as previously described
arrows mark the positionsofreplicativeIIform DNAII;
duplex DNAwithasmallgapor nickinonestrand)
single-stranded DNA (ssDNA) standards. Heat-denatured
5'-termi-nally labeledHindIII DNA fragmentsA of DNA Haelll
fragmentsXX174of DNAwere runas size markers.
the
nucleotide
lengthstandards areindicated atRemoval ofATP fromthe reactionshad noeffect
The BMRFI protein alone exhibited noDNA
(Fig. 4B) or in an alkaline agarose gel (Fig. 4C). The reaction without added Polverified the lack of Pol contamination in the
BMRFI Pol accessory protein (data not shown).
With substoichiometric levels of the EBV Pol catalytic subunit
alone,
low levels ofDNAsynthesisweredetected(Fig. 4A). The EBV Pol catalytic subunit extended the primer slightly (-50 nucleotides), and no full-length product was observed (Fig. 4Band C). Analyses ofthe replication products in an alkaline agarose gel showed no increase in product sizethroughout the time course, indicating the distributive action of the Pol catalytic subunit in the absence of the BMRF1 Pol accessory protein. Addition of the BMRF1
protein,
however, resulted in accumulation of full-length replicative form II.Some part of the products consisted of specific bands ofpause sites on the template. We presume that these pause sites represent sites of a substantial helical region on t1 single-stranded DNA template. In the presence of thX3MRFl protein, the BALF5 Pol catalytic protein movedth .gh these barriers and completed synthesis of some of the 7.2-kb Ml3mpl8 template within 20 min. Most of the primed single-stranded DNA remained unchanged, as detected by
UV-induced ethidium bromide fluorescence during the time in
which full-length products (replicative form II) were formed
(data not shown). These findings supportthe highly processive mode of nucleotide polymerization (>7,200 nucleotides) by
the EBV DNA Pol holoenzyme reconstituted in vitro, consis-tent with previous findings on the EBV DNAPol holoenzyme
purified from EBV-producing lymphoblastoid cells (25). The 20 min required for complete replication of an M13
single-stranded DNA circle (7.2 kb) yields an average nucleotide turnover of six nucleotides per s per Pol molecule. In the
absence of the accessory protein, the template was replicated
at a rate of about 1.5 nucleotides pers (quantified by
measur-ing the kinetics of deoxyribonucleotide incorporation). Thus,
addition of the BMRF1 protein resulted in a highly processive
mode of polymerization and therebyat leastfourfold
stimula-tion of the rate of incorporation to six nucleotides per s.
Discussion and Conclusions. Several lines of evidence
sup-port the viewthatEBV DNAPol isa holoenzyme consisting of
the BALF5 and BMRF1 proteins (11, 15, 24). Despite exten-sive efforts, it has been difficult to isolate the BALF5 Pol catalytic subunit free from the BMRF1 Pol accessory subunit
(28). Only by cloning the genes for BALF5 and BMRF1 and overproducingthese products did it become possible tospecify their functions. Therefore, we separately overexpressed these proteins in a baculovirus expression system and purified them
to homogeneity. In this study, we compared the enzymatic
properties of theDNAPolactivity catalyzed by the BALF5 Pol
catalytic subunit in thepresence orabsence of the BMRF1 Pol
accessory proteinbyusingashortorlongsingle-strandedDNA
template to facilitate the study of the role of each of thesetwo
components in the EBV DNA Pol reaction.
Under conditions of an excess of a long single-stranded
DNA template over BALF5 protein molecules, the BALF5 protein initiates DNA synthesis from the same number of
primers in the absence as in the presence of the BMRF1
protein. In theabsence of the BMRFI protein, the extensions
are short, around 50 nucleotides long. Even when given sufficient time for DNA synthesis, the product size was
un-changed although nucleotide incorporation increased with
time. The BALF5 protein dissociates from the primer after
incorporation ofaround 50 nucleotides. Thus, the BALF5 Pol catalytic subunit by itself is a quasiprocessive enzyme and acts
distributively. Furthermore, in the gap-filling reaction on an
activated calf thymus DNA template, increasing
concentra-tions of ammoniumsulfateprogressively reduced the DNAPol
A
-T -l
I
-ei~~~~~~~~~~~1
3
0 2
-0-i
1
iS
0 10 20 30
40--~
TI-2
1Iim mI IIB
c
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.65.297.83.448.2]7652 NOTES
activity catalyzed by theEBV DNA Pol catalytic subunit. On the basis of thesefindings,wespeculatethat thebindingaffinity of the Polcatalytic subunit for the primer terminus isweakand that high ionic strength destabilizes the catalytic protein-primer terminus interaction. With the BMRF1 Pol accessory
protein present in the reaction, the salt dependency ofEBV DNA Pol activity changed drastically. Although there is no
direct evidence that the BALF5 and BMRF1 proteins form a
complexinvitro, theBMRF1 Pol accessoryproteinappearsto interact with theBALF5Polcatalytic subunit and stabilize the binding of the BALF5 Pol catalytic subunit to the primer terminus. The complex formation appears to be
specific
and tight, because maximal stimulation ofEBV DNA Polactivity
wasobtained withstoichiometric levels ofBMRFIprotein with
respect to the BALF5 proteinpresent. In thepresence of the BMRF1 protein, theBALF5protein polymerizes thousands of nucleotideson thesameprimer template without
dissociation,
while the Pol catalytic subunit alone normally falls off the growing 3' end of the DNA chain. The double-stranded DNA-binding activity of the BMRF1 protein may play a
significant
role in theaffinity between the holoenzyme and the growing 3'-OH end of the primer template. To clarify these speculations, an in vitro study of the interactionbetween the BALF5 and BMRF1 proteins withregardtobinding affinity fortheprimer terminus is in progress.
Unexpectedly, in the absence of ammonium
sulfate,
the BMRF1 protein greatly inhibited the DNA Pol activity of theBALF5 proteinon an activatedDNAtemplate. However, the
BMRF1 protein didnotinhibit theDNAPolactivity catalyzed by theE. coli Poll large fragment orbacteriophage T4 DNA
Polon anactivatedDNAtemplate
(data
notshown). Although
themechanism forPolinhibition
by
the BMRF1protein
atlow ionicstrength isnotclear, the above-described results indicate that the inhibition is also dueto aspecific
interaction between the BALF5 and BMRF1 proteins.Kiehl and Dorsky
(12)
havereported
that the EBV Pol catalytic subunit alone exhibited noDNAPolactivity
in aninvitro translation system.However,
expression
of both theEBVBALF5 and BMRF1 proteins generated DNAPol
activity
inthereticulocyte lysate. Onthe other
hand,
Linetal.(16)
havefoundthat the BALF5
protein
exhibited DNAPolactivity
in an in vitro translation system. However, addition of theBMRF1 protein resulted in a decrease in Pol
activity.
Ourstudy
has solved thediscrepancy
betweenthem;
the saltconcentration assayed affectsEBV DNAPol
activity strongly.
That isto say, at low salt concentrations the BALF5
protein
exhibitsDNAPol
activity
but addition of theBMRF1protein
decreases Pol
activity.
Athigh
saltconcentrations the BALF5 protein exhibits little Polactivity
but addition ofthe BMRF1protein increases Pol
activity greatly.
The BMRF1Pol accessoryproteinincreased the
processivity
and thereby the
polymerization
rateof theBALF5protein
to six nucleotides pers.Thevalue obtainedwith the EBVDNA Polholoenzyme reconstitutedinvitro is closeto12nucleotides per s,thevalue obtainedwiththe EBV DNAPolholoenzyme purified from EBV-producing lymphoblastoid cells (25).Ad-dition of the EBV single-stranded DNA-binding protein (BALF2 protein) to the reaction mixture may increase the
polymerization rate more.Furtherstudyis needed.
The herpes simplex virus type 1 UL42 protein is a herpes
simplex
virus DNA Pol accessory protein that increases theprocessivity of polymerization (4, 7, 9).TheUL42proteinalso has double-stranded DNA-binding activity and specifically interacts with the UL30 Pol catalytic protein. Gottliebetal.(7) havereported that theUL42protein hadnoeffectonthe DNA
Pol activity ofthe UL30 Pol catalytic subunitwhen activated
DNAwasusedas a
primer template,
incontrasttothe resultsobtained here with EBV DNA Pol. The UL30 Pol
catalytic
subunit
by
itself isrelatively
resistant tohigh
salt concentra-tions(17). Thus, binding
of theherpes
simplex
virus UL30protein
totheprimer
terminusmaybetighter
than that oftheEBVBALF5
protein.
Weview the BMRF1
protein
as anauxiliary protein
totheEBV DNAPol
catalytic
subunit(BALF5 protein).
The roleofthe BMRFI
protein
with respect to the BALF5protein
isanalogoustothatof theaccessorysubunits of
phage
T4 DNAPol,
DNAPol8,
and theE. coliDNAPollllholoenzyme,
i.e.,
toincrease the
processivity
andthereby
themacroscopic
rateof DNA
synthesis
of theenzymatically
active core Pol. Inphage T4,
the gene 43protein by
itself is a DNA Pol and a3'-to-5' exonuclease
(14).
The three accessoryproteins
en-codedby
genes44/62
and 45 ofphage
T4 increase theprocessivity
and hence therateof Polactivity (10, 32).
The T4gene
44/62 complex
has bothDNA-dependent
ATPase andprimer
terminus-binding
activities that are stimulatedby
thegene 45-encoded
protein.
The ATPaseactivity
isthought
toreflect an energy
requirement
to maintaina stablePol-acces-sory
protein complex
at theprimer
terminus. The gene 45-encodedprotein, by itself,
haslittleDNA-binding activity
but functions to increaseprocessivity.
As a secondexample,
pro-cessive DNA
synthesis by
DNA Pol6requires
eukaryotic
replication
factors RF-C and PCNA(14,
23).
RF-C exhibitsDNA-dependent
ATPaseactivity
andprimer
terminus-binding
activity,
as does the T4 gene44/62
complex
(22).
PCNAcooperateswith RF-Ctostimulate
processive
DNAsynthesis
by
DNAPoWI
like the T4 gene 45-encodedprotein. PCNA,
by itself,
has littleDNA-binding activity.
The DNA Pollllholoenzyme
ofE. coli shows the same functionalhomology
(14).
E. coli Pollll accessory subunits y/8, and1B
cooperate toincrease
processivity
(5).
The'y/8
complex
has DNA-depen-dentATPase.ATPhydrolysis
isrequired
for thesesubunitstoform a
preinitiation complex
on aprimed
DNAtemplate.
The( subunit ofE. coliPollllhasnointrinsic
affinity
forDNAbut dimerizes to form a torus that issterically
retained on the nucleic acid strand(18).
Alarge
excess of the 1 subunit isrequired,
probably
because of the weak interaction of the 1subunit withDNAPolIII*
(3).
Inall threesystems,theaction ofthe accessoryproteins
can bevisualized asclamping
of the core Pol to theprimer template,
enabling
processive
DNAsynthesis
to occur. In contrast to thesecomplex
cases, theBMRF1
protein
appears to besimple
and bifunctional.Al-though
theBMRF1protein
doesnot exhibitDNA-dependent
ATPase
activity
(27),
itpossessesdouble-strandedDNA-bind-ing
activity
forprimer
recognition
andactsas asliding clamp
to increaseprocessivity.
TheDNA-binding
propertyof the EBV BMRF1protein
isdifferentfrom that of the accessoryproteins
of the three systems. Forexample,
the BMRF1protein
binds double-stranded DNA withoutATPhydrolysis
and does notnecessarily require
aprimer terminus,
whereas RF-C bindsspecifically
totheprimer
terminusjunction
butnot tosingle-
ordouble-stranded DNA
(22). Thus,
the BMRF1protein
effi-ciently
stabilizes the BALF5protein
at theprimer
terminuswithout an energy
requirement
and slides on the single-stranded DNAtemplate. However,
itremains to beclarifiedhow theBMRF1 Pol accessory
protein
can slidetofollow theelongating primer
withoutacting
as abrake.Wethank K. Maeno andI. R.Lehmanfor encouragement. This work was supported by a grant-in-aid from the Ministry of Education, Science,and Culture ofJapan, bythe Uehara Memorial Research Foundation, by the Ishida Foundation, and by the Nitto Foundation.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
REFERENCES
1. Baer, R., A. T.Bankier, M. D. Biggin, P. L.Deininger,P. J. Farrell, T. J. Gibson, G. Hatfull, G.S.Hudson,S. C. Satchwell, C. Seguin, P. S. Tuffnell, and B. G. Barrell. 1984. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature (Lon-don) 310:207-21 1.
2. Chiou, J. F., J.K. K.Li,and Y.-C.Cheng. 1985.Demonstration of a stimulatory protein for virus-specified DNA polymerase in phorbol-ester treatedEpstein-Barr viruscarrying cells. Proc. Natl. Acad. Sci. USA 82:5728-5731.
3. Crute,J. J., R. J. LaDuca, K. 0.Johanson, C. S.McHenry,and R. A. Bambara. 1983. Excess 1B subunit can bypass the ATP requirementfor highly processive synthesis by theEscherichia coli DNA polymerase III holoenzyme. J. Biol. Chem. 258:11344-11349.
4. Crute, J. J., and I. R. Lehman. 1989. Herpes simplex-I DNA polymerase.Identification ofanintrinsic 5' to3' exonuclease with ribonucleaseHactivity. J. Biol. Chem. 264:19266-19270. 5. Fay, P. J., K. 0.Johanson, C. S. McHenry,and R. A.Bambara.
1982. Size classes of products synthesized processively by two subassemblies ofEscherichia coli DNA polymerase III holoen-zyme. J.Biol. Chem. 257:5692-5699.
6. Fixman, E. D., G. S. Hayward, and S. D. Hayward. 1992. trans-acting requirementsforreplicationofEpstein-Barrvirus ori-Lyt. J.Virol. 66:5030-5039.
7. Gottlieb,J.,A.I.Marcy,D. M.Coen, andM.D.Challberg. 1990. Theherpes simplex virus type 1 UL42 gene product:asubunit of DNApolymerase that functions to increaseprocessivity. J. Virol. 64:5976-5987.
8. Hammerschmidt, W., and B. Sugden. 1988. Identification and characterization oforiLyt, a lytic origin of DNA replication of Epstein-Barr virus. Cell 55:427-433.
9. Hernandez,T.R.,andI.R. Lehman. 1990.Functional interaction between the herpes simplex-I DNA polymerase and UL42 pro-tein. J. Biol. Chem. 265:11227-11232.
10. Javis,T.C.,L.S.Paul, J.W.Hockensmith,and P. H. vonHippel. 1989. Structure andenzymatic studies of theT4 DNAreplication system.II. ATPaseproperties of the polymerase accessory protein complex. J. Biol. Chem. 264:12717-12729.
11. Kallin, B.,L.Sternas,A. K.Saemundssen, J. Luka,H.Jornvall,B. Eriksson, P.-Z. Tao, M. T. Nilsson,andG. Klein. 1985. Purifica-tion ofEpstein-BarrvirusDNApolymerase from P3HR-1 cells. J. Virol. 54:561-568.
12. Kiehl, A., and D. I. Dorsky. 1991. Cooperation of EBV DNA polymerase and EA-D (BMRF1) in vitro and colocalization in nuclei of infected cells.Virology 184:330-340.
13. Knopf,K.-W.1979.PropertiesofherpessimplexDNApolymerase and characterization ofits associated exonucleaseactivity.Eur.J. Biochem. 98:231-244.
14. Kornberg, A.,and T. Baker. 1992.DNAreplication, 2nd ed. W. H. Freeman &Co., New York.
15. Li, J.-S., B.-S. Zhou,G.E.Dutschman, S. P.Grill, R.-S.Tan, and Y.-C.Cheng. 1987.Association of Epstein-Barr virus early antigen diffuse component andvirus-specifiedDNApolymerase activity. J. Virol. 61:2947-2949.
16. Lin, J.-C.,N. D.Sista, F.
Besenqon,
J.Kamine,andJ.S.Pagano. 1991. Identification and functional characterization ofEpstein-Barrvirus DNA polymerase by in vitro transcription-translation of acloned gene. J. Virol. 65:2728-2731.
17. Marcy, A. I.,P. D. Olivo, M. D.Challberg,and D. M.Coen.1990. Enzymatic activities of overexpressed herpes simplex virusDNA polymerase purified from recombinant baculovirus-infected insect cells. Nucleic Acids Res. 18:1207-1215.
18. O'Donnell, M., and P. S. Studwell. 1990. Total reconstitutionof DNApolymerase III holoenzyme revealsdual accessory protein clamps. J. Biol. Chem. 265:1179-1187.
19. Pearson,G.R., B. Vroman, B.Chase,T.Sculley,M.Hummel,and E. Kieff. 1983. Identification of polypeptide components of the Epstein-Barr virus early antigen complex with monoclonal anti-bodies. J. Virol. 47:193-201.
20. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed. Cold Spring Harbor Labo-ratory,ColdSpring Harbor, N.Y.
21. Takagi, S., K. Takada, and T. Sairenji. 1991. Formation of intranuclear replication compartments of Epstein-Barr virus with redistribution of BZLF1 and BMRFI gene products. Virology 185:309-315.
22. Tsurimoto, T., and B. Stillman. 1990. Functions of replication factor C andproliferating cell nuclear antigen: functional similar-ity of DNA polymerase accessory proteins from human cells and bacteriophage T4. Proc. Natl. Acad. Sci. USA 87:1023-1027. 23. Tsurimoto, T., and B. Stillman. 1991.Replication factors required
for SV40 DNA replication in vitro. I. DNA structure-specific
recognition of a primer-template junction by eukaryotic DNA polymerase and their accessoryproteins. J. Biol. Chem. 266:1950-1960.
24. Tsurumi, T. 1991.Characterization of 3'-to-5' exonucleaseactivity associated with Epstein-Barr virus DNA polymerase. Virology
182:376-381.
25. Tsurumi,T. 1991.Primer terminus recognition and highly proces-sivereplicationbyEpstein-Barrvirus DNApolymerase. Biochem. J.280:708-713.
26. Tsurumi,T. 1992. Selective inhibition of the3'-to-5' exonuclease
activity associated with Epstein-Barr virus DNA polymerase by
ribonucleoside monophosphates. Virology 189:803-807. 27. Tsurumi,T. 1993.Purification and characterization of the
DNA-bindingactivity of the Epstein-Barr virusDNApolymerase acces-soryproteinBMRF1geneproducts,asexpressedin insect cellsby usingthe baculovirus system. J. Virol. 67:1681-1687.
28. Tsurumi, T.Unpublishedresult.
29. Tsurumi, T.,A.Kobayashi,K.Tamai,T.Daikoku, R. Kurachi,and Y.Nishiyama.1993. Functional expression and characterization of theEpstein-Barrvirus DNApolymerase catalyticsubunit. J. Virol. 67:4651-4658.
30. Yanisch-Perron, C., J. Vieira, and J. Messing. 1985. Improved M13phage cloningvectorsand host strains: nucleotide sequences of the Ml3mpl8andpUC19vectors.Gene33:103-119. 31. Yates, J. L., N. Warren, D. Reisman, and B. Sugden. 1984. A
cis-actingelement from the Epstein-Barrviral genome that per-mits stablereplication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci. USA 81:3806-3810.
32. Young,M.C.,M. K.Reddy,and P. H. vonHippel. 1992. Structure and function of thebacteriophage T4 DNApolymerase holoen-zyme.Biochemistry 31:8675-8690.