Copyright
©
1994, American Society for MicrobiologyIntegrase
Mutants of
Human Immunodeficiency
Virus
Type
1
with
a
Specific
Defect in
Integration
BRUNELLA
TADDEO,1t WILLIAM A.
HASELTINE,2
ANDCHRISM.FARNET'*
Divisionof
Human Retrovirology, Dana-Farber CancerInstitute, Boston, Massachusetts02115,1and Human Genome
Sciences, Inc.,Rockville,Maryland 208502
Received7 June 1994/Accepted 23 August 1994A
previous genetic
analysis of the humanimmunodeficiency virustype1integraseprotein failedtoidentifysingle amino acid
substitutions thatonly block the integration of viralDNA (C.-G. Shin,B. Taddeo,W. A.Haseltine, and C.
M. Farnet, J. Virol. 68:1633-1642, 1994). Additional substitutions of amino acidsthat arehighly
conserved amongretroviral integraseswereconstructed in human immunodeficiencyvirus type1 andanalyzed
fortheir
effects
onviral protein synthesis and processing,virion morphology, and viralDNA synthesisand
integration
inanattempt toidentifymutantswithaspecific defect in integration.Four single amino acidsubstitutions
resultedin replication defectiveviruses. Conservative, single aminoacid substitutions of thetwoinvariant
aspartic acid residues found inall retroviral integrasesprevented the integration of viral DNA andhad
no detectableeffect
on the other stages in the viral replication cycle, indicating that these mutantsexhibited
aspecific defect
in integration. Mutationsat twopositions, S-81 and P-109, blockedtheintegrationof
viral DNA
but also resulted in the productionof viral particles that exhibitedreducedreversetranscriptaseactivity, suggesting additional
defects in viral replication.Substitution
of the highly conserved amino acid T66had
noeffect
onviral
replication in aCD4+
human T-cell line. Thisanalysis
extends the range of possiblephenotypes
thatmaybe produced by single amino acid substitutions in conserved residues of the integraseprotein.
The
integration of
a DNA copyof the
retroviral
RNAgenome into
the host cell
genomeis essential for viral
repli-cation. The viral
integrase
protein, encoded by the 3' end of
the viralpol
gene, isrequired
forintegration (1, 3, 4, 17, 18,
20). Comparison of the amino acid
sequencesof
anumber of
retroviral integrases
hasdemonstrated the
presenceof highly
conserved residues and motifs that may be
important
forintegrase
function(7, 9).
Themosthighly conserved region
of retroviral integrases lies in the centralregion
oftheprotein
and is definedby
three acidic amino acid residues in a conserved spatial arrangement [the D,D(35)E motifl (9, 10).Mutations in any one of the three residues result in the
complete
lossof allintegrase enzymatic
activitiesinvitro(5, 6,
10, 12, 13, 24)
andcanblockviralreplication
in cell cultures(12, 22). Establishing
a correlation betweenspecific
in vitroenzymatic
defects andtheresulting
virusreplication phenotype
is
complicated by
thepleiotropic
effects of someintegrase
point
mutantsonviralreplication.
Aprevious analysis
ofsingle
amino acid substitutionsin the humanimmunodeficiency
virus type 1(HIV-1) integrase protein
failedtoidentify
mutations thatonly
blocked theintegration
ofviral DNA andestablishedthat
integrase
mutationsmaysuffer defectsin virionprecursorpolypeptide processing,
virionmorphology,
or viral DNAsynthesis.
Theintegrase protein
isinitially
synthesized
as apart
ofa
gag-pol
precursorpolyprotein, possibly accounting
for theeffect ofintegrasemutationsonthe other
enzymatic
functionsencoded
by
thepol
gene(19, 21, 23).
Inan attempt to
identify integrase changes
thatspecifically
prevent theintegration
ofviralDNA,
additionalpoint
muta-tions
affecting
highly
conserved amino acid residues of theHIV-1
integrase protein
were constructed andanalyzed
fortheir effects on viral
replication
in cell culture. Five HIV-1*Corresponding author.
tPermanent address: IstitutoSuperiorediSanita,Rome,Italy.
integrase mutants were
constructed by introducing single
amino acid substitutions in
the
highly conserved D,D(35)E
region
of theprotein. The
natureand location of each point
mutation
areindicated
in thelegend
toFig. 1. The integrase
mutantswereanalyzed
for their effectsonviral
replication by
monitoring multiple
stepsin the viralreplication cycle.
Each of theintegrase
mutations wasintroduced into
anHXBc2
provirus,
andwild-type
and mutantproviral DNAs
were
transfected
intoCOS-1
cells.Forty-eight hours following
transfection,
thecellsweremetabolically labeled with
[35S]cys-teine,
and viralproteins
wereimmunoprecipitated
from celllysates
by using
anHIV-1-infected
patient
serum.As shown inFig. 1,
thepatternof viralproteins
produced by
theintegrase
mutants was identical to thatproduced by wild-type virus,
demonstrating
that none of the amino acid substitutions had detectable effects on thesynthesis
orprocessing
of viralproteins.
Virion release from COS-1 cells transfectedwith
wild-type
andintegrase
mutantproviruses
was measured 48 h aftertransfection
by assaying
culture supernatants for particle-associated viralproteins,
viralp24 antigen,
and reversetran-scriptase (RT) activity.
Awild-type
pattern of [35S]cysteine-labeled viralproteins
was found in the supernatants of cellstransfected with all of the
integrase
mutants(Fig. 1), indicating
that the mutations had nodetectableeffect ontheprocess ofvirion formation. Consistent with this observation was the
demonstration that each of the
integrase
mutants releasedwild-type
levels ofparticle-associated p24 antigen
into theculture supernatants, as measured
by
aradioimmunoassay
(data
notshown). Surprisingly,
supernatants
recovered fromCOS-1 cells transfectedwith mutants S811 and
P109S
consis-tently
showedareduced level ofparticle-associated
RTactivity
(Fig. 2),
eventhough essentially wild-type
levels of viralparticles
were detected in culturesupernatants by
sodiumdodecyl
sulfate-polyacrylamide gel electrophoresis
(SDS-8401
on November 9, 2019 by guest
http://jvi.asm.org/
8402 NOTES
c e
--" < -. r)
r-1 2 3 4 5 C)
V rIo rn
< - G i0
-cc; _
7( 8 9 10 i1 M
B
cel virion
V LU LU
12 3 4 5
E.. ..s
gp 120
p24
p17
..,:e 1 . ...!I
--- 7.:
tlillpml,
I.-I-.,",
M-'.11-14
gp160
gp120
-p55
lqmw~Molw.' p 24
p17
FIG. 1. SDS-PAGEanalysisof viralproteins presentincelllysatesand virions after transfectionofCOS-1 cells withwild-typeorintegrase
mutantproviralDNA. TheintegrasemutantswereconstructedasdescribedbyShin et al.(22).Briefly,a1,671-bp fragment spanningtheEcoRV and EcoRI sites of HXBc2(nucleotides2977to5122, accordingtothenumberingscheme ofMyersetal.[16])that encodesaportionofintegrase proteinwassubcloned intopSL1180(Pharmacia),andtheresulting plasmidwasusedas atargetfor site-directedmutagenesis using procedures already described (11). The mutagenicoligonucleotides used areD64E (5'GGAATATGGCAACTAGAgTGTACACATITA3'), T66A(5'TG
GCAACTAGATTGTTgCACATYTTAGAAGGA3'),
S81I (5'GCAGTTCATGTAGCCAtTGGATATATAGAA3'), P109S(5'TTAGCAGGAA
GATGGtCAGTAAAAACAATA3'), and D116E (5'AAAACAATACATACTGAaAATGGCAGCAAT3') (nucleotide changes introduced to createthemutationareindicatedbylowercaseletters).COS-1 cellsweretransfectedwith 10,ugofproviralDNAbyaDEAE-dextranprocedure (15). The procedures formetaboliclabelingofCOS-1 and viralproteinimmunoprecipitationwereperformedaspreviouslydescribed(22).COS-1werelabeledwith[35S]cysteine48 h aftertransfection,and the viralproteinswereimmunoprecipitatedfromcelllysates(A,lanes 1 to6; B,lanes 2and3)orfrom virionpelletedthroughasucrosecushion from culture supernatants(A,lanes 7to11; B,lanes4and5)andresolvedon a10to 15% (wt/vol) polyacrylamide gradient gel. (A)The cellsweremocktransfected(lane 1) ortransfected with T66A(lanes2and7),S811(lanes3 and8),P109S (lanes4 and9),orD116E(lanes5and 10)mutantproviralDNAorwild-type proviralDNA(WT; lanes6 and11).Therelative positions of HIV-1 proteins andprotein markers (lane M; GIBCO) are indicated, on the left and right, respectively. (B) Cellswere mock transfected(lane 1)ortransfected withwild-type proviral (WT;lanes 2 and4)orD64Emutantproviral (lanes3and5)DNA. Thepositionsof HIV-1proteinsarealso shown.
125
PAGE) analysis (Fig. 1A, lanes
8 and9)
andp24
radioimmu-noassay(data
notshown),
and levelsof
pl609a9-Po
precursorwere similar in
wild-type
and S81 andP109
mutant viruses(Fig.
1A, lanes 8, 9,
and11).
Virion
morphology
was assessedby
thin-section electron
microscopy
ofmonolayer preparations
ofCOS-1
cellstrans-fected with
wild-type and integrase
mutantproviruses. All of
themutants releasedvirionswith
amorphology
indistinguish-able from that of
wild-type virions (data
notshown), indicating
thatthe mutations
did
nothave
major effectsonthe latestagesofviral
replication, including virion assembly
and maturation. The effectsof
the integrase point mutations
onviralrepli-cation were
assessed
by
infecting
cells of theCD4+
humanSupTl
T-cellline
withsupernatantsprepared from
COS-1 cells 48 h aftertransfection
with mutant orwild-type
provirus.Parallel cultures of SupTl cells
wereinfected
with equivalentdoses
(RT units)
ofwild-type
or mutantvirus. The
courseof
virusreplication
wasmonitored by measuring
supernatant RT levels every3
days.
Only integrase
mutant T66Ashowed
aa:
E
x
100 T
75
50-0
[image:2.612.94.512.75.359.2]WT D64E T66A S811 P109S D116E
FIG. 2. Viral particle release from COS-1 cells transfected with wild-type (WT)orintegrasemutantproviralDNA. Virus production
wasmonitored 48 h aftertransfectionbydetermination of RT activity
onthe supernatants of transfected COS-1, using procedures already described (2). Barsindicate therange ofvaluesobtainedfrom four experiments.
A
J. VIROL.
1
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.332.525.530.667.2]A.
B.
3 --- D64E
T66A
-0- S811 -_- P109S -*- D116E
- WT
10 20
Days after infection
2
L--E
cm C
,IQ
0\4 0L
30
0
* S811
A P109S -*-- WT
30
1-0 20
Days After Infection
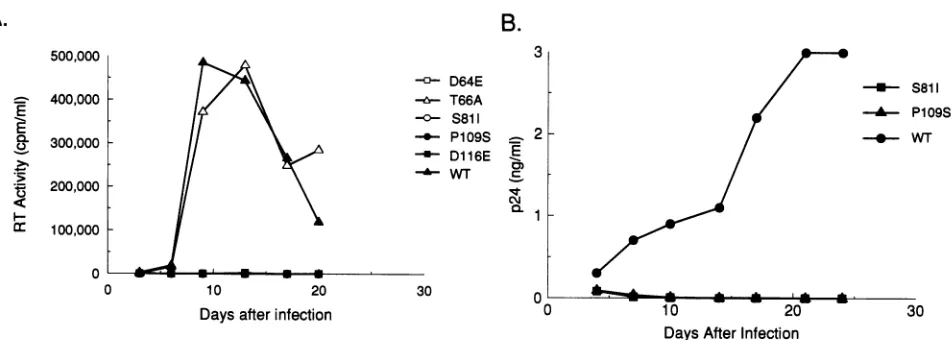
FIG. 3. Replication of wild-type (WT) and integrasemutantviruses in humanSupTlcells. HumanSupTlcells (106)wereinfected with an
aliquot of filteredsupematantrecovered from transfected COS-1 equivalentto
105
cpmofRTactivity. The infected cellswerefedwith freshmediumeverythird day, and thesupernatantswereassayed forRT activity (A) and p24 antigen production (B)by usingacommercially available
HIV-1 p24 enzyme-linked immunosorbent assay kit (NEN/DuPont, Boston, Mass.). The p24 antigen measured at day 4 includes antigen
contributedby residual input virus.
replication profile similar
tothat
of wild-type virus (Fig. 3A).The other integrase
mutantswere unableto establishapro-ductive infection of
SupTl cells, showing
noproduction of RTactivity for
upto25 days postinfection (Fig. 3A).
Because viralparticles produced by
mutantsS81I and
P109S showed
reducedlevels of
RTactivity, replication of these
two mutantswasalsomonitored
by measuring
supernatantp24 antigen levels.
Noviral p24 antigen
wasproduced by cells infected
with theS81I
or
P109S integrase
mutant(Fig. 3B).
Each of the replication-defective integrase
mutants wastested for the
ability
tosynthesize viral
DNAfollowing
infec-tionof
SupTl cells and
tointegrate the viral
DNAinto the
cellulargenome.Total DNAwasextracted from cells 24 h
after
infection with wild-type
orintegrase
mutantvirus. Viral DNA
was
readily
detectedby PCR amplification following infection
with
wild-type
orintegrase
mutantvirus(Fig. 4A), indicating
that viral
DNAsynthesis
was notdetectably affected by the
integrase
mutations. The same DNAsamples
weresubse-quently analyzed by using
amethod for the detection of viralDNA
integrated
into the cellular genome(14).
None of thereplication-defective integrase
mutantsgenerated
detectable levels ofintegrated
viral DNA(Fig. 4B,
lanes 3 to6),
incontrast to
wild-type
virus(Fig. 4B,
lane2), demonstrating
that thesemutantssufferadefect in viral DNAintegration.
The
replication phenotypes
of themutantviruses described here are consistent withprevious
biochemicalanalyses
of recombinant HIV-1integrase proteins carrying
similar amino acid substitutions. The T66A mutationhadnodetectable effect on the in vitroenzymatic
activities of recombinant HIV-1integrase (6, 8)
or onviralreplication
incell culture(Fig. 3A).
Thereplication
defectdisplayed by
mutantsD64E,
S81I,
P109S,
and D116E may result from the loss ofintegrase
enzymatic activity.
Conservative substitutions of D-64 andD-116 resulted in the loss of the
oligonucleotide cleavage,
DNA strandtransfer,
anddisintegration
activities of HIV-1integrase
invitro(6). Similarly,
theP109S
mutation reducedthe DNA
binding
andoligonucleotide cleavage
activities ofpurified
HIV-1integrase (5, 6, 10).
Mutation ofS81
has beenreported
tocompletely (8)
orpartially (24)
reduce thein vitroenzymatic
activitiesof HIV-1integrase,
but theseeffects mayresult from the reduced
solubility
of the mutantprotein (24).
The
integrase protein is synthesized in infected
cellsaspartof a
gag-pol polyprotein
precursorthat is subsequently
pro-cessed to
form the structural and enzymatic proteins
of themature
virion. Previous analyses of the replication
of integrasemutant
viruses demonstrated that single amino acid
substitu-tions
inthe
integrase protein
canproduce defects in virion
precursorpolypeptide processing, virion
morphology, and viral DNAsynthesis (22),
mostlikely by interfering with the
properfolding and proteolytic processing of the viral
precursorpolyprotein.
Incontrast, twoof the
replication-defective
inte-grase mutantsdescribed
here, D64E and D116E, specifically
blocked theintegration of
viral DNAand had
nodetectable
effect on other steps in the viral
replication cycle, providing
strong supportforthe idea
thatintegrase function is absolutely
required
for retroviralreplication
incell lines. The
natureof
the amino acid substitutionappeared
to be animportant
determinant ofthe observed viral
replication phenotype.
Forexample,
thenonconservative substitution of alanine for aminoacid D-116
haddramatic effects
onthe late
eventsof the viralreplication cycle (22), resulting
in immature virionparticles
that containedno detectable RTactivity,
while the conserva-tive D116E mutation described here hadno detectable effect onthereplication cycle
outside ofablocktointegration.
The
analysis
of theS81I
andP109S
mutants extends the range ofpossible
effects ofintegrase
mutations on the viralreplication cycle.
Both of these mutants were similar towild-type
virus in thesynthesis
andprocessing
of viralproteins,
in theassembly
and maturation ofvirions,
andinthesynthesis
of viral DNAfollowing
infection ofSupTl
cells,
asmeasuredby
theassaysdescribed here.However,
viralparticles produced
inboth of thesemutantscontained lower levels ofRTactivity
thanwild-type virions, making
itimpossible
toassign
thereplication
defect ofthese mutantstoasole defectin integra-tion. The RTactivity expressed by
these mutantswasappar-ently
sufficienttodrive thesynthesis
of viralDNA in infectedcells
(Fig. 4A). However,
the reducedenzymeactivity
in theinvitro RTassayindicates that these mutationshave
pleiotropic
effects onpol
gene functions. TheS81I
mutation has beenreported
toreducethesolubility
andstability
of the HIV-1(8)
and HIV-2(24) integrase proteins
invitro, presumably by
interfering
with the properfolding
of thepolypeptide.
Im-500,000E
0
I-.t
:r
400,000
300,000 200,000 100,000 0 0
1
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.83.559.72.242.2]8404 NOTES
1 2 34 5 6
A.
-
595
bp
B.
561 bp
FIG. 4. PCR analysis of synthesisandintegrationof viral DNA in
SupTl cells infected withwild-typeorreplication-defective integrase
mutant virus. (A) The analysis of viral DNA synthesis in infected
SupTlcellswascarried outasdescribedbyShinetal. (22). Briefly, total DNAwasextracted from infected cells 24 h afterinfection, using
standardtechniques (15).DNAsampleswerepretreatedwith restric-tion endonucleaseDpnI (New England Biolabs, Inc., Beverly, Mass.) toremoveplasmidDNAspotentially contaminatingCOS-1 cell
super-natants. DNAs (1 ,ug of each) were subject to 35 cycles of PCR amplificationwithapairofprimers,env-1(nucleotides7950to7969of HXBc2) and env-2 (nucleotides 8545 to8526 ofHXBc2),whichare
abletoamplifya595-bp fragmentintheenvgeneof HIV-1. Eachcycle consisted of1 min15 sof denaturation (94°C), 1.5 minofannealing (57°C), and 2.5minof extension(72°C). Amplified products resulting from PCR were analyzed by electrophoresis on 1.5% agarose gel, transferredtoanylonfilter(Hybond-N; Amersham),andhybridizedat 42°C with a 32P-labeled specific oligonucleotide, env-3 (nucleotides
8285to8308 ofHXBc2),that iscomplementarytoaninternalregion ofamplified fragment. (B) The analysisofintegrated viralDNA in the infected SupTl cells wasperformed as described by Li et al. (14). Briefly, each high-molecular-weight DNAsamplewassubjectedto a
double digestionwith restriction endonucleases SacI and XbaI
over-nightat37°C. Digested DNAswereseparated by electrophoresison
1%low-melting-pointagarose,and sections of thegel containing DNA fragments between 1 and 5 kb in size were excised. DNAs were
extractedby usingtheRapid Gene Clean kit (Bio 101, Inc., La Jolla, Calif.)andweresubjectedtoligation in ordertoreconstructafull-size long terminalrepeat(LTR). The ligation mixtureswereused directly forPCRamplification. Primers used in this PCR assay wereLTR 1
(nucleotides 75 to98 of HXBc2) and AA55 (25), abletoamplify a
561-bp fragment between the U3 and R regions of the HIV-1 LTR. Theamplified products wereanalyzedasdescribed above, using the
oligonucleotide M669 (25) as a probe. The critical step of this procedure is the size exclusion of DNA fragments smaller than1kbp: molecules ofthis sizeareexpectedtocontain fragments of theHIV-1 LTR derived from Sacl digestion of linear and circularviral DNAs (note that XbaI does not cut HIV-1 DNA). LTR-containing DNA fragments longer than 1 kbp will be derived from viralLTRsthat have joined togenomic DNAasa result of integration. SupTl DNAwas
prepared after mock infection (lane 1), 24 h after infection with wild-type virus (lane 2), 24 h after infectionwithmutantD64E(lane 3), 24 hafterinfection withmutantS81I (lane 4),24h after infection with mutant P109S (lane 5), or24h after infection withmutant D116E (lane 6). The sizes of amplifiedproductsarealso indicated.
proper
folding
of the integrase polypeptide in thegag-pol
precursormayproduce subtle defects inprecursorprocessingor
incorporation into virions, resulting
in reducedparticle-associated RT
activity.
We thankJoseph Sodroski for critical review of the manuscript, Cha-Gyun Shin for the gift of plasmids, Stefano Fiore for helpful
discussion, and Virginia M. Nixon for manuscript preparation and
artwork.
This workwas supported by NIH grants P30 CA06516 (Cancer Center), P30 A128691 (Center for AIDS Research), U01
A124845
(NCDDG), and 5R01 AI31388 andbyagift from the G. Harold and Leila Y. Mathers Charitable Foundation. W. A. Haseltinewas therecipient of an award from Bristol Myers Squibb. B. Taddeo was
supported bya fellowship from Istituto Superiore di Sanita, Rome, Italy.
REFERENCES
1. Clavel,F.,M.D.Hoggan,R. L.Willey, K.
Strebel,
M. A.Martin,
and R Repuske. 1989. Genetic recombination of human immu-nodeficiencyvirus. J.Virol.63:1455-1459.2. Dayton, A.I., J.G.Sodroski,C. A.Rosen,W.C.
Goh,
and W.A.Haseltine. 1986. The trans-activator gene of the human T cell lymphotropicvirus type IIIis
required
forreplication.
Cell44:941-957.
3. Donehower, L. A. 1988. Analysis of mutant
Moloney
murine leukemia virusescontaining linker insertionmutationsin the 3' region of pol.J.Virol. 62:3958-3964.4. Donehower, L. A., and H. E. Varmus. 1984. A mutant murine leukemia virus withasingle missense codonin
pol
isdefective ina function affecting
integration.
Proc. Natl. Acad. Sci. USA 81:6461-6465.5. Drelich, M., R. Wilhelm, and J. Mous. 1992. Identification of amino acid residues critical for endonuclease and
integration
activities ofHIV-1INprotein
in vitro.Virology
188:459-468.6. Engelman, A., and R Craigie. 1992.Identification of amino acid residues critical forhuman
immunodeficiency
virus type 1inte-grasefunctionin vitro. J. Virol.66:6361-6369.
7. Johnson, M. S., M. A.McClure, D. F. Feng, J.Gray,and R. F. Doolittle.1986.Computer analysis of retroviral polgenes:
assign-ment ofeukaryotic functions to specific sequences and
homo-logueswith nonviral enzymes. Proc. Natl. Acad. Sci.USA 83:7648-7652.
8. Katz,RA., J.P.G.Mack,G.Merkel, J. Kulkowsky,Z.Ge, J. Leis,
and A. M. Skalka.1992. Requirement fora conserved serinein
bothprocessingandjoining activities of retroviral integrase.Proc.
Natl. Acad. Sci.USA 88:6741-6745.
9. Khan, E., J. P. G. Mack, R A. Katz, J. Kulkosky, and A. M. Skalka. 1991.Retroviralintegrase domains:DNAbinding and the recognition ofLTR sequences.Nucleic AcidsRes. 19:851-860.
10. Kulkosky,J.,K. S.Jones, R A. Katz, J. P.G. Mack, and A. M. Skalka. 1992. Residues critical forretroviralintegrative
recombi-nation in a region that is highly conserved among retroviral/
retrotransposonintegrasesand bacterialinsertionsequence trans-posases.Mol. Cell. Biol. 12:2331-2338.
11. Kunkel, T. A., J. D. Roberts, and R. A. Zakour. 1989.Rapid and
efficient site-specific mutagenesis without phenotypic selection.
MethodsEnzymol. 154:367-382.
12. LaFemina, R. L., C. L. Schneider, H. L. Robbins, P. L. Callahan, K. LeGrow, E. Roth, W. A. Schlief, and E. A. Emini. 1992.
Requirement of active human immunodeficiency virus type 1
integraseenzymeforproductiveinfection of human T-lymphoid
cells.J.Virol.66:7414-7419.
13. Leavitt, A. D., L. Shiue, and H. E. Varmus. 1993. Site-directed
mutagenesisof HIV-1 integrasedemonstrates differential effects onintegrasefunctions in vitro. J.Biol. Chem. 268:2113-2119. 14. Li, G., M. Simm, M. J. Potash, and D. J.Volsky. 1993. Human
immunodeficiency virustype 1 DNA synthesis, integration, and
efficient viral replication in growth-arrested T cells. J. Virol.
67:3969-3977.
15. Maniatis, T., E. F. Fritsch, and J. Sambrook 1982. Molecular
cloning: a laboratory manual. Cold Spring Harbor Laboratory,
ColdSpring Harbor, N.Y.
16. Myers, G., J. A. Bergofsky, B. Kober, R F. Smith, and G. N. Paolakis (ed.).Human retroviruses and AIDS 1991. A
compila-tionandanalysisof nucleic acid and amino acid sequences, May
1991, vol. 2. Theoretical Biology and Biophysics, Los Alamos NationalLaboratory, LosAlamos,N.Mex.
17. Panganiban,A.T., and H. M. Temin. 1984. Theretroviruspol gene
encodesaproduct requiredfor DNA integration: identification of J. VIROL.
M
W.,R-W
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.99.248.74.237.2]aretrovirus int locus. Proc.Natl.Acad. Sci. USA 81:7885-7889.
18. Quinn, T. P., and D. P. Grandgenett. 1988. Genetic evidence that the avian retrovirus DNA endonuclease domain ofpolisnecessary
for viralintegration. J. Virol. 62:2307-2312.
19. Roth, M. J., P. Schwartzberg,N. Tanese, and S. P. Goff. 1990. Analysis of mutations in the integration function of Moloney murine leukemia virus: effectson DNA binding and cutting. J. Virol. 64:4709-4717.
20. Sakai, H., M. Kawamura, J.-I. Sakuragi, S. Sakuragi, R. Shibata, A.Ishimoto, N. Ono, S. Veda, andA.Adachi.1993.Integration is
essential for efficientgeneexpression of human immunodeficiency
virustype1.J. Virol. 67:1169-1174.
21. Schwartzberg, P., J. Colicelli, and S. P. Goff. 1984. Construction and analysis of deletion mutations in thepolgene ofMoloney murine leukemia virus: a newviral functionrequired for
produc-tive infection. Cell 37:1043-1052.
22. Shin, C.-G., B. Taddeo, W.A.Haseltine,andC.M. Farnet.1994.
Genetic analysis of the human immunodeficiency virus type 1 integrase protein. J. Virol. 68:1633-1642.
23. Stevenson, M., S. Haggerty, C.A.Lamonia, C. M. Meier, S. K. Welch, and A. J. Wasia. 1990. Integration is notnecessary for expression of human immunodeficiency virus type 1 protein products. J. Virol. 64:2421-2425.
24. vanGent, D.C., A. A. M. Oude Groeneger, andR.H. A. Plasterk 1992. Mutational analysis of the integrase protein of human immunodeficiency virus type 2. Proc. Natl. Acad. Sci. USA 89: 9598-9602.
25. Zack, J. A., S. J. Arrigo, S. R. Weitsman, A. S. Go, A. Haislip, and
L.S. Y. Chen. 1990. HIV-1entryintoquiescent primary lympho-cytes:molecularanalysis revealsalabile, latent viralstructure.Cell 61:213-222.