Replication and Nef Function
Aniqa Shahid,aAlex Olvera,bGursev Anmole,cXiaomei T. Kuang,cLaura A. Cotton,aMontserrat Plana,dChristian Brander,b,e,f Mark A. Brockman,a,c,gZabrina L. Brummea,g
Faculty of Health Sciences, Simon Fraser University, Burnaby, Canadaa
; IrsiCaixa AIDS Research Institute, HIVACAT, UAB, Barcelona, Spainb
; Department of Molecular Biology and Biochemistry, Simon Fraser University, Burnaby, Canadac
; AIDS Research Group, IDIBAPS, HIVACAT, Hospital Clinic, UAB, Barcelona, Spaind
; Institució Catalana de Recerca i Estudis Avançats, Barcelona, Spaine
; University of Vic and Central University of Catalonia, Vic, Barcelona, Spainf
; British Columbia Centre for Excellence in HIV/ AIDS, Vancouver, Canadag
ABSTRACT
HLA-B*13 is associated with superior
in vivo
HIV-1 viremia control. Protection is thought to be mediated by sustained targeting
of key cytotoxic T lymphocyte (CTL) epitopes and viral fitness costs of CTL escape in Gag although additional factors may
con-tribute. We assessed the impact of 10 published B*13-associated polymorphisms in Gag, Pol, and Nef, in 23 biologically relevant
combinations, on HIV-1 replication capacity and Nef-mediated reduction of cell surface CD4 and HLA class I expression.
Muta-tions were engineered into HIV-1
NL4.3, and replication capacity was measured using a green fluorescent protein (GFP) reporter T
cell line. Nef-mediated CD4 and HLA-A*02 downregulation was assessed by flow cytometry, and T cell recognition of infected
target cells was measured via coculture with an HIV-specific luciferase reporter cell line. When tested individually, only
Gag-I147L and Gag-I437L incurred replicative costs (5% and 17%, respectively), consistent with prior reports. The
Gag-I437L-medi-ated replication defect was rescued to wild-type levels by the adjacent K436R mutation. A novel B*13 epitope, comprising 8
resi-dues and terminating at Gag
147, was identified in p24
Gag(GQMVHQAI
Gag140 –147). No other single or combination Gag, Pol, or
Nef mutant impaired viral replication. Single Nef mutations did not affect CD4 or HLA downregulation; however, the Nef
dou-ble mutant E24Q-Q107R showed 40% impairment in HLA downregulation with no evidence of Nef stability defects. Moreover,
target cells infected with HIV-1-Nef
E24Q-Q107Rwere recognized better by HIV-specific T cells than those infected with HIV-1
NL4.3or single Nef mutants. Our results indicate that CTL escape in Gag and Nef can be functionally costly and suggest that these
ef-fects may contribute to long-term HIV-1 control by HLA-B*13.
IMPORTANCE
Protective effects of HLA-B*13 on HIV-1 disease progression are mediated in part by fitness costs of CTL escape mutations in
conserved Gag epitopes, but other mechanisms remain incompletely known. We extend our knowledge of the impact of
B*13-driven escape on HIV-1 replication by identifying Gag-K436R as a compensatory mutation for the fitness-costly Gag-I437L. We
also identify Gag-I147L, the most rapidly and commonly selected B*13-driven substitution in HIV-1, as a putative C-terminal
anchor residue mutation in a novel B*13 epitope. Most notably, we identify a novel escape-driven fitness defect: B*13-driven
substitutions E24Q and Q107R in Nef, when present together, substantially impair this protein’s ability to downregulate HLA
class I. This, in turn, increases the visibility of infected cells to HIV-specific T cells. Our results suggest that B*13-associated
es-cape mutations impair HIV-1 replication by two distinct mechanisms, that is, by reducing Gag fitness and dampening Nef
im-mune evasion function.
T
he natural history of HIV-1 is influenced by human leukocyte
antigen (HLA) class I polymorphisms (
1–4
). Notably,
HLA-B*57 and B*27 are associated with viral load control and slower
disease progression (
5
,
6
), but other “protective” HLA class I
al-leles have also been identified (
4
,
7
,
8
). HLA-B*13 is one such
allele, but it remains understudied in part because it is relatively
rare in many populations (expressed in less than 4% of Caucasians
[
9
] and Africans [
10
,
11
] although its frequency exceeds 10% in
some Asian populations [
12–14
]). HLA-B*13, of which the
most common subtype is B*13:02 (
15
), is associated with lower
viral loads in chronic infection (
10
,
11
,
14
,
16
,
17
) and lower
hazard ratios of progression to AIDS in historic natural history
studies (
18
). HLA-B*13 is also enriched among HIV
control-lers (
6
,
19
,
20
).
B*13-mediated protective effects are attributed, at least
par-tially, to sustained targeting of conserved B*13-restricted
cyto-toxic T lymphocyte (CTL) epitopes, particularly in HIV-1 Gag
(
11
) and Nef (
21
). Optimally described B*13 epitopes have also
been identified in Pol (
22
) although these responses appear to
contribute less to control (
11
). Similar to the effects of B*57 (
23–
Received3 August 2015 Accepted31 August 2015
Accepted manuscript posted online9 September 2015
CitationShahid A, Olvera A, Anmole G, Kuang XT, Cotton LA, Plana M, Brander C, Brockman MA, Brumme ZL. 2015. Consequences of HLA-B*13-associated escape mutations on HIV-1 replication and Nef function. J Virol 89:11557–11571. doi:10.1128/JVI.01955-15.
Editor:F. Kirchhoff
Address correspondence to Zabrina L. Brumme, [email protected].
G.A. and X.T.K. contributed equally to this article.
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /JVI.01955-15.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
on November 7, 2019 by guest
http://jvi.asm.org/
25
) and B*27 (
26
), the protective effects of B*13 may also be
me-diated to some extent by costs to viral replicative fitness incurred
as a result of immune escape within certain CTL epitopes, notably
in Gag (
27
). These replicative costs may limit the net viral
“bene-fit” gained via immune evasion, leading to relative viral control or
reduced pathogenesis. In particular, selection of a Gag-I437L or
-I437M escape mutation at the C-terminal anchor residue of the
B*13-restricted p1
GagRI9 epitope (the 9-residue Gag sequence at
position 429 to 437, RQANFLGKI
429 – 437) substantially impaired
in vitro
HIV-1 replicative capacity (
27
,
28
). Likewise, the
B*13-driven Gag-I147L polymorphism, located four codons
down-stream of the p24
GagVV9 epitope (VQNLQGQMV
135–143
) confers
minor replication defects (
28
).
Though fitness-reducing CTL escape mutations in HIV-1
commonly occur in Gag (
28
,
29
), escape in Pol can also incur
replicative costs (
30
,
31
). In addition, naturally occurring
se-quence variation in Nef (
32–34
), including HLA-driven escape
mutations (
35
,
36
), can modulate this protein’s major functions,
including cell surface CD4 (
37
) and HLA class I (
38
)
downregu-lation. However, the functional costs of B*13-driven immune
es-cape mutations outside Gag and their possible contributions to
B*13-mediated protective effects have not been investigated.
Moreover, the extent to which secondary (compensatory)
poly-morphisms can restore fitness costs of primary B*13-driven
es-cape mutations also remains incompletely known. Such effects,
however, are supported by observations of reduced replication
capacity of patient-derived B*13 Gag sequences in early, but not
chronic, infection (
25
).
To further elucidate the replicative and functional
conse-quences of B*13-driven HIV-1 evolution, we investigated the 10
polymorphisms in Gag, Pol, and Nef that are most commonly
selected in B*13-expressing individuals (
39
). Specifically, we
tested these polymorphisms alone and in biologically observed
combinations for effects on
in vitro
HIV-1 replication capacity
and Nef-mediated CD4 and HLA class I downregulation
activ-ity. Our results extend current knowledge of B*13-driven Gag
replicative costs (
27
,
28
) by identifying Gag-K436R as a
com-pensatory mutation for Gag-I437L. We also provide evidence
for a novel B*13-restricted CTL epitope terminating at Gag
codon 147, B*13’s most frequent escape site in HIV-1 (
39
).
Notably, although B*13-associated substitutions in Pol and
Nef did not impair viral replication, the B*13-associated Nef
poly-morphisms E24Q-Q107R, when expressed together, reduced
Nef’s ability to downregulate HLA class I from the infected cell
surface. This, in turn, enhanced the recognition of infected cells by
HIV-1-specific T cells
in vitro
. Though this mutation combination
is observed in less than 5% of B*13-expressing persons, it
never-theless represents a novel consequence of CTL escape in HIV-1.
Specifically, our results suggest that B*13-mediated immune
con-trol could also be attributable, at least in part, to CTL
escape-driven attenuation of Nef’s immune evasion function.
MATERIALS AND METHODS
Definition of HLA-B*13-associated polymorphisms. Ten published HLA-B*13 and/or B*13:02-associated polymorphisms in HIV-1 Gag (n⫽
4), Pol (n⫽4), and Nef (n⫽2), originally identified via statistical asso-ciation with phylogenetic correction in the International HIV Adaptation Collaborative (IHAC) cohort comprising ⬎1,800 antiretroviral-naive chronically HIV-1 subtype B-infected individuals (39), were selected for analysis (Table 1andFig. 1A; see also Fig. S1 in the supplemental
mate-rial). These polymorphisms represent all B*13/B*13:02-associated sites in Gag, Pol, and Nef for which a specific “adapted” (CTL escape form) amino acid was identified at aPvalue of⬍0.0001 andqvalue of⬍0.05 in the original study (39). At two of these codons (protease 63 and reverse trans-criptase [RT] 369), the original study identified two possible adapted forms; the one with the lowerPvalue was selected for the present analysis. The extent to which these polymorphisms are enriched in B*13:02-posi-tive (B*13:02⫹) versus B*13:02-negative (B*13:02⫺) individuals in the original study (39), along with their locations respective to published or bioinformatically predicted B*13-restricted CTL epitopes, is shown in Fig. S1 in the supplemental material.
[image:2.585.297.544.86.391.2]Characterization of escape rates and frequencies in B*13ⴙ individ-uals.The frequencies and kinetics of selection of B*13-associated poly-morphisms in acute/early HIV-1 subtype B infection were defined using longitudinal plasma HIV-1 RNA sequences from nine B*13:02⫹ serocon-verters from cohorts in Berlin (Jessen-Praxis Medical Clinic, Berlin, Ger-many), New York (Aaron Diamond AIDS Research Center, New York, NY, USA), Boston (Massachusetts General Hospital, Boston, MA, USA), and Vancouver (British Columbia Centre for Excellence in HIV/AIDS; Vanguard and Vancouver Injection Drug Users Study [VIDUS], Canada) (40,41). All patients provided written informed consent, and the cohort studies were approved by their respective institutional review boards. For four patients, HIV-1 sequences were already published (41). For others, HIV-1 RNA was extracted from plasma, and Gag, Pol, and Nef were am-plified in independent nested reverse transcription-PCRs (RT-PCRs) us-ing HIV-1-specific primers. Amplicons were bidirectionally sequenced on a 3130xl automated DNA sequencer (Applied Biosystems, Inc.), and chro-matograms were analyzed using Sequencher, version 5.0 (Genecodes).
TABLE 1Single and combination mutations in HIV-1 Gag, Pol, and Nef evaluated in this study
Protein and
mutation type Mutation(s)
Mutant namea
Gag
Single A146S
S---I147L
-L--K436R
--R-I437L ---L
Combination I147L-I437L -L-L
A146S-I147L SL--I147L-K436R -LR-I147L-K436R-I437L -LRL A146S-I147L-K436R SLR-K436R-I437L --RL A146S-I147L-K436R-I437L SLRL A146S-I147L-I437L SL-L Pol
Single L63S (protease)
S---Q334N (RT)
-N--T369A (RT)
--A-K374R (RT) ---R
Combination L63S-T369A-K374R S-AR
T369A-K374R --AR
L63S-T369A
S-A-L63S-K374R S--R
Nef
Single E24Q
Q-Q107R -R
Combination E24Q-Q107R QR
a
Mutants are named according to the scheme shown at the bottom ofFig. 1BtoD. Combination mutations are listed in descending order based on their prevalencesin vivo.
on November 7, 2019 by guest
http://jvi.asm.org/
FIG 1HLA-B*13-associated polymorphisms in HIV-1 Gag, Pol, and Nef. (A) Locations of 10 published HLA-B*13-associated polymorphisms in HIV-1 subtype B Gag, Pol, and Nef (HXB2 codon numbering) (39). (B to D) Top panels show the rates of escape of these polymorphisms, calculated from longitudinal plasma HIV-1 RNA sequences from nine B*13:02 seroconverters using Kaplan-Meier methods. Note that curves for Gag-A146S-K436R and Pol-L63S-T369A are superimposed. Prevalences of these polymorphisms in B*13:02⫹individuals in early (1 year postinfection;n⫽9) and chronic (n⫽69) infection are shown in the middle panels. Frequencies of these polymorphisms, alone and in combination, in 69 B*13:02⫹individuals with chronic infection are shown in the bottom panels. Polymorphisms are listed in decreasing frequency, with hyphens denoting the wild-type (nonescaped) form. Single and combination mutants indicated with a dot were engineered into NL4.3 and tested forin vitroviral RC. PR, protease.
on November 7, 2019 by guest
http://jvi.asm.org/
Sequences were aligned to the HIV-1 subtype B reference strain HXB2 (GenBank accession numberK03455) using an algorithm based on the HyPhy platform (42). Time from estimated infection date to the first detection of the specific B*13-associated polymorphism was determined using Kaplan-Meier methods; these data were also used to calculate escape prevalence after 1 year of infection. The prevalence of these mutations in chronic infection was calculated using published plasma HIV-1 RNA se-quences from 69 B*13:02⫹individuals in the IHAC cohort (39). Note that cryopreserved peripheral blood mononuclear cells (PBMCs) were not available from these patients. Patient HIV and HLA data are available upon request in accordance with the Providence Health Care/University of British Columbia Institutional Review Board protocols.
Generation of mutant HIV-1.B*13-associated polymorphisms were engineered into the HIV-1 subtype B reference strain NL4.3 (NIH AIDS Research and Reference Reagent Program, catalog 114; contributed by Malcolm Martin [43]). First, pNL4.3gag,pol(covering protease and the first 400 codons of reverse transcriptase), andnefwere subcloned into pCR2.1-TOPO (Life Technologies). Individual mutations were intro-duced into these plasmids using a QuikChange II XL site-directed mu-tagenesis kit (Stratagene). Combination mutants were generated by suc-cessive rounds of mutagenesis. All mutants were confirmed by DNA sequencing. Mutant subcloned genes were then used to generate recom-binant NL4.3 viruses by homologous recombination (25). Briefly, mutant
gag,pol, andnefwere reamplified from subclones using 100-mer primers complementary to NL4.3. After verification on an agarose gel, each am-plicon was cotransfected, along with the relevant linearized pNL4.3 vector (⌬gag,⌬pol, or⌬nefvector), into a Tat-inducible CEM-GFP-reporter (where GFP is green fluorescent protein) T cell line (GXR25) (44) via electroporation. Cells were incubated for 10 days at 37°C in R20⫹ me-dium (RPMI meme-dium with phenol red, supplemented with 20% [vol/vol] fetal bovine serum [FBS] and 1% [vol/vol] penicillin-streptomycin) to allow the generation and propagation of infectious virions. GFP ex-pression (indicating HIV-1 infection) of these cultures was monitored daily by flow cytometry (Guava easyCyte 8HT; Millipore); once this reached ⬎15%, supernatants containing infectious mutant virions were harvested, aliquoted, and frozen at⫺80°C. The median time to harvest was 13 days (range, 11 to 14 days). HIV-1 RNA from each virus stock was reextracted (Invitrogen PureLink Genomic DNA/RNA kit; Life Technologies) and reamplified, and the relevant gene was rese-quenced to confirm the presence of the desired substitution(s) and the absence of any others.
In vitroHIV-1 replication capacity assays.Viral titers were assessed as described previously by infecting GXR25 reporter cells in order to de-termine the volume necessary to achieve a multiplicity of infection (MOI) of 0.003 (0.3% HIV-infected cells) at 2 days after infection (25,31,45,46). For subsequent viral replication assays, GXR25 cells were infected with control (wild-type NL4.3 [wtNL4.3]) and mutant viruses at an MOI of 0.003, after which the percentage of GFP-positive (GFP⫹; HIV-infected) cells was monitored daily by flow cytometry until day 6. For each virus, the natural log of the slope of virus spread (measured as percent infected cells) was calculated during the exponential phase. This value was then normal-ized by the mean rate of spread of wtNL4.3 such that replication capacity (RC) values of⬍1.0 or⬎1.0 indicate a rate of viral spread lower than or higher than that of NL4.3, respectively. Each virus was assayed in a mini-mum of two independent experiments with each containing three repli-cates.
Characterization of a novel putative HLA-B*13:02-restricted epitope in p24 Gag.The presence of a novel B*13:02-restricted epitope overlapping Gag146and Gag147was investigated bioinformatically and experimentally. First, the HIV-1 consensus B amino acid sequence at Gag135–155was scanned for the presence of B*13:02-restricted 8- to
11-mer epitopes using NetMHCpan, version 2.8 (47,48) (http://www.cbs .dtu.dk/services/NetMHCpan/). This algorithm identifies candidate epitopes based on their predicted half-maximal inhibitory concentrations (IC50), where values of⬍500 nM, 500 to 5,000 nM, and⬎5,000 nM are
predictive of strong binders, weak binders, and nonbinders, respectively (47,48). After candidate epitopes were experimentally verified, binding predictions were repeated for variants containing B*13-associated poly-morphisms at Gag146and/or Gag147. PBMCs from seven B*13:02⫹
chron-ically HIV-1 subtype B-infected individuals recruited at the Hospital Clinic Barcelona (n⫽4) and at IrsiCaixa AIDS Research Institute, Bad-alona (n⫽3), Spain, collected with written informed consent, were screened for responses against C- and N-terminally extended versions of the predicted optimal epitope using a gamma interferon (IFN-␥) enzyme-linked immunospot (ELISpot) assay as described previously (31). The threshold for positive responses was defined as the highest of the following three criteria: a minimum of five spots per well, the mean number of spot-forming cells (SFC) of the negative-control wells plus three times its standard deviation, or three times the mean of negative-control well val-ues (49). Plasma samples were also available from three of the patients; these were subjected to HIV-1 RNA Gag sequencing.
Assessment of Nef-mediated CD4 and HLA class I downregulation in infected cells.Nef-mediated CD4 and HLA class I downregulation was assessed in a GXR25 cell line stably transduced to express HLA-A*02:01 (GXR-A*02). Briefly, one million GXR-A*02 cells were infected with Nef control and mutant viruses at an MOI of 0.01. Control viruses included wtNL4.3, NL4.3 with a deletion of Nef (⌬Nef), and NL4.3 with an M20A mutation in Nef (NefM20A) (the latter two kindly provided by Takamasa
Ueno, Kumamoto University), as well as an HIV-uninfected culture. The NefM20Avirus was used as a control since it is impaired for HLA class I
downregulation (50). The NL4.3-⌬Nef virus was pseudotyped with vesic-ular stomatitis virus G protein (VSV-G) to enhance infection in GXR-A*02 cells (51). Once HIV-1 infection reached⬇10 to 15%, 500,000 cells were stained with allophycocyanin (APC)-labeled anti-CD4 and phyco-erythrin (PE)-labeled anti-HLA-A*02 antibodies (BD Biosciences), and cell surface expression of these molecules was measured by flow cy-tometry. The level of Nef-mediated CD4 downregulation of each virus was expressed as the ratio of the median fluorescence intensity (MFI) of CD4 expression in GFP⫹(HIV-infected) cells to the MFI of CD4 in the uninfected cultures (the latter was used because CD4 downregula-tion precedes GFP expression in GXR25 cells, leading to lower MFIs in the GFP⫺gate in infected cultures). These values were then normal-ized to the value of the wtNL4.3 control. The calculation was as fol-lows: [(1⫺(MFI of GFP⫹nef mutant/MFI of GFP⫺uninfected))/(1⫺(MFI of GFP⫹wtNL4.3/MFI of GFP⫺uninfected))]⫻100. As such, normalized
values of⬍100% and⬎100% indicate CD4 downregulation functions lower than or higher than values for wtNL4.3, respectively. The same calculation was used to quantify Nef-mediated HLA-A*02 downregula-tion. Viruses were assessed in a minimum of three independent experi-ments.
Impact of Nef-mediated HLA class I downregulation on infected-cell recognition by HIV-1-specific effector T infected-cells.To determine the im-pact of Nef-mediated HLA-A*02 downregulation on the recognition of infected target cells by HIV-1-specific effector T cells, we employed a reporter T cell coculture assay. Briefly, 1 million GXR-A*02 target cells were infected with control (wtNL4.3-Nef,⌬Nef, or NefM20A) or Nef
mu-tant virus at an MOI of 0.01 in R20⫹medium without phenol red. Cul-tures were monitored daily by flow cytometry and used as “targets” when the proportion of GFP⫹(HIV-infected) cells reached⬇10 to 15%. HLA-A*02-restricted reporter “effector” T cells were generated using a modifi-cation of previously described methods (52,53). Briefly, Jurkat T cells were cotransfected with four expression plasmids: 3 g each of pSELECTGFPZeo(Invivogen) containing T cell receptor␣(TCR␣) and
TCRgenes isolated from a CTL clone specific for the A*02-restricted p24GagFK10Gagepitope (FLGKIWPSYK, Gag residues 433 to 442) (kindly
provided by Mario Ostrowski and Brad Jones, University of Toronto); 5 g of a CD8␣expression plasmid (Invivogen); and 10g of a luciferase reporter plasmid driven by nuclear factor of activated T cells (NFAT) (Panomics/Affymetrix). Reporter effector T cells were electroporated us-ing a Gene Pulser Xcell Electroporation System (Bio-Rad Laboratories)
on November 7, 2019 by guest
http://jvi.asm.org/
(square-wave protocol; 500 V, 2,000F, 3 ms, and 1 pulse), recovered at room temperature for 10 min, and incubated for 18 h in R10⫹ me-dium (RPMI 1640 meme-dium, 10% FBS, and 1% [vol/vol] penicillin-streptomycin) to allow the expression of TCR. A total of 50,000 HIV-1-infected target cells were cocultured with 50,000 effector cells (1:1 effector/target [E/T] ratio) for 6 h, and TCR-mediated signaling was detected as NFAT-driven luciferase expression using a Steady-Glo lu-ciferase system (Promega) and quantified using a Tecan Infinite M200 plate reader. Prior studies have indicated that this signal is dependent on endogenously processed HIV-1 Gag FK10Gagpeptide antigen
pre-sented on HLA-A*02.
Western blotting.Western blotting was performed as described pre-viously (36). GXR-A*02 cells were infected with control or mutant vi-ruses. When cultures reached⬇10 to 15% infection, 800,000 cells were pelleted and lysed. Due to the similar molecular masses (⬇27 kDa) of Nef and GFP (HIV-1 infection control), lysates were split in half, and two blots were processed in parallel (with Nef and the-actin housekeeping gene control on one and GFP on the other). Cell lysates were subjected to SDS-PAGE using Mini-Protean TGX 4% to 20% gels (Bio-Rad Laborato-ries), and proteins were electroblotted onto a polyvinylidene difluoride (PVDF) membrane (GE Healthcare). Nef was detected using a polyclonal rabbit antibody (NIH AIDS Research and Reference Reagent Program, catalog 2949; contributed by Ronald Swanstrom) (1:4,000) (54), followed by staining with secondary donkey anti-rabbit antibody (GE Healthcare) (1:30,000). Expression levels of-actin and GFP were assessed using pri-mary mouse anti-actin (Sigma) (1:20,000) and pripri-mary mouse anti-GFP (1:10,000), respectively, followed by secondary goat anti-mouse (Jackson ImmunoResearch) (1:20,000). Western blots were developed using Clar-ity Western ECL Substrate (Bio-Rad Laboratories), based on the chemi-luminescence detection method. Blots were developed with ImageQuant LAS 400 (GE Healthcare).
Statistical analyses.The one-samplettest was used to assess whether replication capacities and CD4/HLA-A*02 downregulation functions of mutant NL4.3 viruses differed significantly from those of the wtNL4.3 reference strain (whose function was set to 1.0 for RC or 100% for CD4/ HLA downregulation). A Bonferroni correction was used to adjust for multiple tests. Spearman’s correlation was used to assess the relationship between Nef-mediated HLA-A*02 downregulation in HIV-infected target cells and NFAT-mediated TCR signaling in HIV-specific effector T cells. Statistical analyses were performed in Prism, version 5.0 (GraphPad Soft-ware). All tests of significance were two tailed.
RESULTS
Rates and frequencies of B*13-mediated escape in HIV-1 Gag,
Pol, and Nef.
Ten published HLA-B*13 and/or
B*13:02-associ-ated polymorphisms in HIV-1 were investigB*13:02-associ-ated: A146S, I147L,
K436R, and I437L in Gag; L63S in protease; Q334N, T369A, and
K374R in reverse transcriptase; and E24Q and Q107R in Nef (
Ta-ble 1
and
Fig. 1A
) (
39
). These polymorphisms are significantly
enriched in B*13:02
⫹individuals with chronic HIV-1 subtype B
infection, and nine of them lie within or near published or
pre-dicted B*13-restricted CTL epitopes (see Fig. S1 in the
supple-mental material). In addition, Gag-I437L and, to a lesser extent,
Gag-K436R are experimentally verified B*13-driven escape
muta-tions within the RI9
Gagepitope (RQANFLGKI
429 – 437
) (
11
) while
Nef-Q107R confers escape from CTL responses against the RI9
Nefepitope (RQDILDLWI
106 –114) (
21
). Together, these observations
support these 10 polymorphisms as commonly selected B*13
es-cape mutations
in vivo
.
To infer the extent and timing of HLA-B*13-driven escape
during HIV-1 infection, we analyzed longitudinal plasma HIV-1
RNA sequences from nine B*13:02
⫹seroconverters (
40
,
41
) and
cross-sectional sequences from 69 B*13:02
⫹individuals with
chronic infection (
39
). In HIV-1 Gag, 36% and 13% of B*13:02
⫹individuals harbored Gag-I147L and -I437L, respectively, at 1 year
postinfection; by the chronic stage, 69% and 26% of B*13:02
⫹individuals harbored these substitutions (
Fig. 1B
, top and
mid-dle). In contrast, Gag-A146S and Gag-K436R emerged later in
infection (reaching 17% and 26%, respectively, by the chronic
stage). For HIV-1 Pol, the protease-L63S, T369A, and
RT-K374R substitutions were observed in
⬇
12% of B*13:02
⫹individ-uals in early infection and in 25 to 37% of B*13:02
⫹individuals in
chronic infection (
Fig. 1C
, top and middle). RT-Q334N frequency
was higher in early (25%) than in chronic (9%) infection
se-quences, suggesting it as a possible transient escape form in some
individuals (
40
); however, our ability to conclude this is limited
since the data are derived from independent cross-sectional early
and chronic cohorts rather than from longitudinal samples from
the same individuals. For HIV-1 Nef, E24Q was observed in 0%
and 12% of early and chronic infection sequences whereas Q107R
was observed in 25% and 33% of sequences at these stages,
respec-tively (
Fig. 1D
, top and middle).
Chronic HIV-1 sequences were additionally analyzed for the
presence of naturally occurring mutant combinations (
Fig. 1B
,
C
,
and
D
, bottom). We began by engineering all combinations
ob-served at
⬎
5% frequency (plus the Nef double E24Q-Q107R
mu-tation, observed at 4% frequency) into an HIV-1
NL4.3backbone.
Later, we engineered two additional Gag combinations for our
analyses of compensatory mutation pathways. Together a total of
23 mutant viruses (10 single and 13 combination mutations) were
constructed (
Table 1
).
Impact of HLA-B*13-associated polymorphisms in Gag on
viral RC.
Mutant HIV-1
NL4.3viruses were evaluated using a
mul-ticycle
in vitro
replication capacity (RC) assay using the GXR25
GFP reporter cell line (
25
,
44
). Representative data are shown in
Fig. 2
, including flow cytometry results, viral growth curves, and
wtNL4.3-normalized replication capacity measurements for
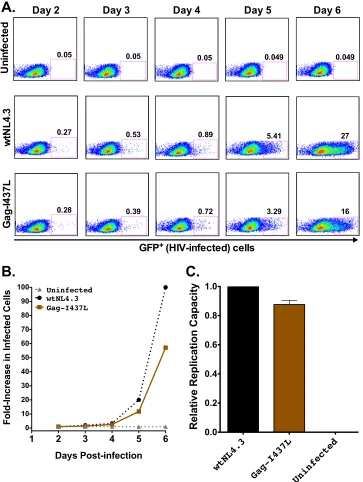
un-infected cultures and those un-infected with wtNL4.3 and Gag-I437L
virus (which is known to impair
in vitro
RC) (
27
,
28
).
When engineered alone into HIV-1
NL4.3, two of the four
B*13-associated Gag polymorphisms, Gag-I147L and Gag-I437L,
re-duced RC by 5% and 17%, respectively (
P
⬍
0.001), while
repli-cation capacities of Gag-A146S and Gag-K436R viruses were
comparable to the wtNL4.3 level (
Fig. 3A
and
B
). The RC
mea-surements for Gag-I147L, Gag-K436R, and Gag-I437L viruses are
consistent with those of previous studies conducted in Jurkat T
cells and CD8
⫹-depleted peripheral blood mononuclear cells
(PBMCs) (
27
,
28
). To our knowledge, no studies have previously
investigated A146S for RC costs (though A146P, which is selected
by HLA-B*57 and other alleles [
39
,
55
], does not carry a replicative
cost in PBMCs [
28
,
55
,
56
]).
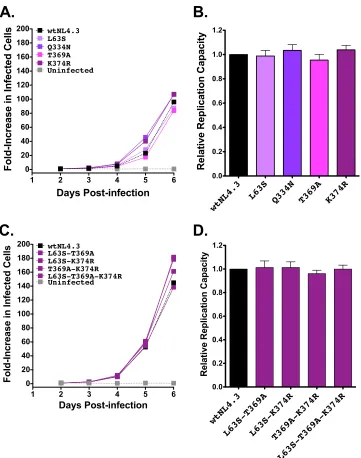
B*13-associated polymorphisms have not been evaluated
pre-viously in combination for their effects on RC; in particular, it is
unclear whether they all represent primary escape mutations or
may also include secondary (compensatory) changes. Eight
B*13-associated polymorphism combinations in Gag were therefore
as-sessed (
Fig. 3C
and
D
). The two combinations that contained
Gag-I437L in the absence of the adjacent K436R (i.e., I147L-I437L
and A146S-I147L-I437L) exhibited 9% and 17% reductions in
RCs, respectively. These values were significantly lower than the
wtNL4.3 value (
P
⬍
0.001) but not significantly different than the
value for Gag-I437L alone, indicating that neither A146S nor
I147L compensates for Gag-I437L-mediated replicative costs. In
contrast, mutant combinations that included Gag-I437L in the
on November 7, 2019 by guest
http://jvi.asm.org/
presence of K436R (K436R-I437L, I147L-K436R-I437L, and
A146S-I147L-K436R-I437L) exhibited RCs comparable to the
wtNL4.3 value, indicating that the replicative cost of Gag-I437L
was compensated by the adjacent K436R. Similarly, the three
combination mutants with Gag-I147L in the presence of the
adja-cent A146S and/or the downstream K436R also exhibited RCs that
were not significantly different from the wtNL4.3 value. This
sug-gests that the modest replicative cost of Gag-I147L in the absence
of I437L might be compensated by A146S or K436R although
further study will be required to elucidate these mechanisms,
par-ticularly in the case of K436R, which lies in a different Gag protein
subunit (p1).
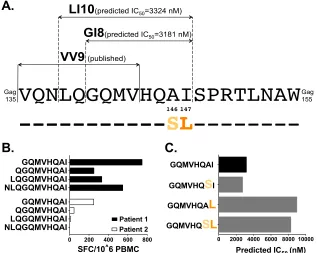
A novel putative B*13-restricted epitope in p24 Gag.
The
re-gion spanning Gag
135–155is dense in CTL epitopes (
22
). It has
previously been hypothesized that Gag-I147L confers escape from
CTL targeting the upstream B*13:02-restricted VV9 epitope (VQ
NLQGQMV
135–143) (
Fig. 4A
) via an antigen-processing
mecha-nism (
11
). Another possibility is that Gag-A146S and/or -I147L
confers escape from an undiscovered B*13:02-restricted epitope
at this site, a hypothesis that is consistent with the higher
preva-FIG 2HIV-1 replication capacity: control data. (A) Representative flow plots of virus spread in culture (measured as percent GFP⫹cells) for uninfected controls and cells infected with wtNL4.3 and mutant Gag-I437L viruses. (B) Growth curves for the data shown in panel A, expressed as the fold increase in infected cells from the day 2 value. (C) Mean RC value for Gag-I437L normalized to that of the wtNL4.3 control, from one representative experiment containing three replicates. Histograms and error bars denote the means and standard deviations, respectively.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.112.475.66.548.2]lence of escape within (versus outside) CTL epitopes (
39
). To
investigate this, we utilized NetMHCpan, version 2.8, to predict
B*13:02-restricted epitopes in the HIV-1 subtype B consensus
se-quence at Gag
135–155. We identified two overlapping candidate
epitopes, LI10 (Gag
138 –147) and GI8 (Gag
140 –147) (
Fig. 4A
), where
the latter is fully embedded in the former. Predicted half-maximal
(IC
50) HLA binding affinities were 3,324 nM (LI10) and 3,181 nM
(GI8), classifying them as weak binders of potentially low
immu-nogenicity (
57
) (
Fig. 4A
). However, for context, the published
adjacent VV9 epitope has a predicted IC
50of 9,174 nM, suggesting
that responses to both LI10 and GI8 are feasible. Of note,
Gag-A146S and -I147L lie at the penultimate and C-terminal residues
of both LI10 and GI8.
IFN-
␥
ELISpot assays were performed using PBMCs from two
chronically infected B*13:02
⫹individuals with responses to this
region. In both cases, GI8 elicited the strongest response to a series
of N-terminally extended peptide sequences that included LI10
(
Fig. 4B
). Removal of the C-terminal isoleucine abrogated
re-sponses, suggesting that this position served as the F-pocket
an-chor residue for HLA-B*13 (data not shown). Limited cell
num-bers precluded HLA restriction confirmation and experimental
validation of A146S and/or I147L as bona fide escape mutations.
Still, the predicted IC
50values of GI8 variants harboring
Gag-I147L were nearly 3-fold higher than the wild-type value,
support-ing I147L as a potential escape mutation (
Fig. 4C
). This
conclu-sion is reinforced by the observation that responding patient 1
FIG 3Replication capacities of NL4.3 viruses encoding B*13-associated polymorphisms in Gag. (A) Representative growth curves of single Gag mutants, expressed as the fold increase in infected cells from the day 2 value. (B) NL4.3-normalized RC values of each single Gag mutant, derived from two independent experiments, each containing triplicate measurements. Histograms and error bars denote the means and standard deviations, respectively. ***,P⬍0.001. (C and D) Experiments are the same as those described for panels A and B, respectively, but with data for combination Gag mutants.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.113.474.60.517.2]harbored wild-type I147 whereas patient 2 harbored I147L in
plasma (data not shown): the latter’s lower GI8 response is thus
consistent with waning of CTL responses to the wild-type epitope
following
in vivo
escape.
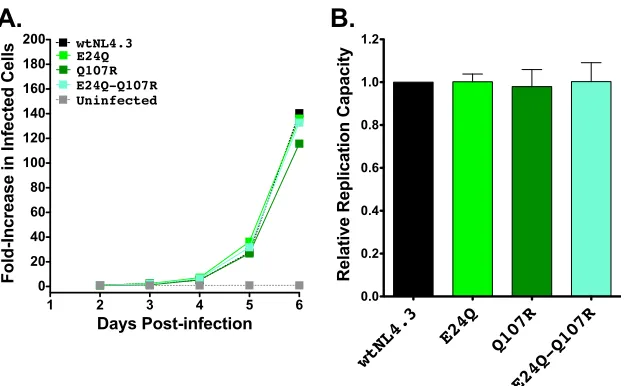
HLA-B*13-associated polymorphisms in Pol and Nef do not
impair viral RC.
We next examined the effects of B*13-associated
polymorphisms in Pol (L63S in protease and Q334N, T369A, and
K374R in RT) and in Nef (E24Q and Q107R) on viral replication.
None of the Pol substitutions significantly altered
in vitro
RCs
when engineered alone into NL4.3 (
Fig. 5A
and
B
), and the four
most common Pol mutant combinations observed in natural
se-quences also did not significantly affect RC (
Fig. 5C
and
D
).
Sim-ilarly, the two B*13-associated Nef polymorphisms did not impair
the RC, either alone or in combination (
Fig. 6A
and
B
). Note that
the Nef observations are not simply attributable to the lack of
requirement of this protein for viral replication
in vitro
as
⌬
Nef
virus replicated poorly in GXR25 cells (in experiments initiated at
an MOI of 0.01, HIV-1
⌬Nefand VSV-G-pseudotyped HIV-1
⌬Nefexhibited 45% and 32% reductions in RCs, respectively,
com-pared to the HIV-1
NL4.3level).
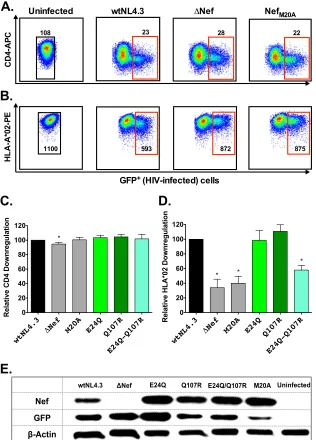
Impact of HLA-B*13-associated Nef polymorphisms on CD4
and HLA class I downregulation.
Naturally occurring sequence
variation in Nef can modulate its CD4 and HLA class I
downregu-lation activities (
33–36
,
58
). We therefore investigated the impact
of B*13-associated polymorphisms on Nef function. To this end,
the GXR25 cell line was stably transduced to express HLA-A*02:01
(GXR-A*02) as a representative HLA class I molecule. GXR-A*02
cells were infected with control viruses (wtNL4.3,
⌬
Nef, and
Nef
M20A; the last is impaired for HLA class I downregulation [
50
]
but replicates comparably to wtNL4.3 [data not shown]) or
mu-tant Nef viruses (E24Q, Q107R, and E24Q-Q107R) and stained
for surface CD4 and A*02 expression once infection reached
⬇
10
to 15%. Representative data for uninfected and control-infected
cultures are shown in
Fig. 7A
and
B
. Compared to the activity of
wtNL4.3,
⌬
Nef virus displayed a modest (5%) reduction in CD4
downregulation that was nevertheless statistically significant and a
substantial (66%) defect in HLA class I downregulation activity
(
P
ⱕ
0.01) (
Fig. 7C
and
D
). Efficient downregulation of CD4 by
⌬
Nef virus is likely due to the actions of Vpu and Env in our
culture system (
59
). The ability of Nef
M20Ato downregulate CD4
was comparable to that of wtNL4.3, but the HLA downregulation
function of Nef
M20Awas impaired by 60% (
P
⬍
0.01). Neither
Nef-E24Q nor -Q107R significantly affected CD4 or HLA class I
downregulation alone; moreover, the CD4 downregulation
func-tion of the E24Q-Q107R double mutant was comparable to that of
wtNL4.3. However, the ability of the double mutant to
downregu-late HLA class I was 42% reduced compared to that of wtNL4.3
(
P
⫽
0.007). Intracellular Nef expression was readily detectable in
cultures infected with control and mutant HIV-1, suggesting that
the attenuated HLA downregulation activity of the Nef double
mutant was not simply due to poor expression or to a stability
defect (
Fig. 7E
).
Impaired HLA class I downregulation of Nef double mutant
renders infected cells more susceptible to recognition by
HIV-1-specific T cells.
An impaired ability of the Nef-E24Q-Q107R
double mutant to downregulate viral peptide/HLA complexes
im-plies that cells infected with this virus should become more visible
to HIV-specific CD8
⫹T cells, regardless of HLA restriction. To
FIG 4Prediction of a novel putative B*13-restricted CTL epitope in p24Gag. (A) Locations and sequences of published and bioinformatically predicted
B*13:02-restricted epitopes are shown above the HIV-1 consensus B amino acid sequence spanning Gag135–155. (B) IFN-␥ELISpot responses, measured as the
number of spot-forming cells (SFC) per million PBMCs, to predicted epitopes and their N-terminal extensions in two B*13:02⫹chronically HIV-1 subtype B-infected patients who responded to this region. (C) NetMHCpan-predicted IC50for GI8 and variants harboring the B*13:02-associated polymorphisms
Gag-A146S and/or -I147L.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.135.452.67.320.2]test this hypothesis, we used a novel coculture assay that
fea-tures target cells (GXR-A*02 cells infected with control or
mu-tant HIV-1) and HIV-1-specific effector cells transiently
ex-pressing an A*02-restricted FK10
Gag-specific TCR
␣
/

, CD8
␣
,
and an NFAT-driven luciferase reporter construct (see Fig. S2
in the supplemental material). In this assay, TCR-dependent
signaling is quantified based on luminescence following
cocul-ture with A*02/FK10
Gag-expressing target cells.
Downregula-tion of HLA-A*02 by wild-type Nef from the infected target cell
surface is expected to lead to reduced TCR signaling in effector
cells, while target cells infected with HIV-1 containing
defec-tive Nef sequences that fail to downregulate HLA-A*02 should
lead to increased TCR signaling.
Consistent with this, we observed a significant negative
corre-lation between the extent of Nef-mediated HLA-A*02
downregu-lation on target cells and TCR-driven luciferase signal in
HIV-1-specific effector cells (Spearman’s
R
,
⫺
0.88;
P
⫽
0.03) (
Fig. 8
).
This was confirmed in three independent experiments
(Spear-man’s
R
,
⫺
0.82 to 0.89;
P
⫽
0.03 to 0.06). Importantly, control
experiments using uninfected GXR-A*02 target cells and parental
GXR25 target cells (lacking A*02) did not induce luciferase upon
coculture with effector cells (data not shown), indicating that
tar-FIG 5Replication capacities of NL4.3 viruses encoding B*13-associated polymorphisms in Pol. (A) Representative growth curves of single Pol mutants, expressed as the fold increase in infected cells from the day 2 value. (B) NL4.3-normalized RC values of each Pol single mutant, obtained from two independent experiments, each containing triplicate measurements. Histograms and error bars denote the means and standard deviations, respectively. (C and D) Experi-ments are the same as those described for panels A and B, respectively, but with data for combination Pol mutants.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.113.474.62.520.2]get cell recognition was HIV-1 epitope and HLA specific. Taken
together, our results suggest that the impaired HLA class I
down-regulation activity of the B*13:02-associated Nef-E24Q-Q107R
double mutant could render HIV-1-infected cells more
vulnera-ble to CD8
⫹T cell responses
in vivo
.
DISCUSSION
We engineered a total of 23 mutant HIV-1
NL4.3recombinant
vi-ruses harboring B*13-associated polymorphisms either alone
(
n
⫽
10) or in naturally occurring combinations (
n
⫽
13) and
investigated their effects on HIV-1 replication and Nef protein
function. Our results extend current knowledge in multiple ways.
First, we confirmed that escape via Gag-I437L incurs a substantial
(17%) fitness cost (
27
,
28
), and we extend this observation by
demonstrating that the adjacent K436R mutation rescues
in vitro
RC to wild-type levels. This compensatory relationship is
corrob-orated by the time course of selection of these polymorphisms
in
vivo
(I437L is ultimately selected in 26% of B*13:02
⫹individuals;
of these, 33% also developed K436R), the highly significant
nega-tive association reported between I437L and wild-type K436 in
HIV-1 subtype B sequences (
39
), and evidence that despite its
strong association with B*13:02, K436R alone does not confer
escape in the majority of HIV-infected individuals (
27
,
60
).
Sim-ilarly, we confirmed that Gag-I147L, the most rapidly and
fre-quently selected B*13:02 escape mutation in HIV-1 (
39
), incurs a
minor (5%) replicative cost, which appears to be rescued by the
adjacent A146S and/or distal K436R mutation. Again, these
puta-tive compensatory relationships are corroborated by their time
course of selection (I147L is ultimately selected in 70% of B*13:02
individuals; of these, 13% also develop the A146S or K436R
mu-tation). Taken together, we hypothesize that replicative costs of
Gag-I147L and -I437L escape mutations, which usually occur
within the first year of infection and may be compensated only
later in fewer than half of cases, contribute to the long-term
en-hanced viremia control observed in B*13:02-expressing persons.
While B*13-mediated escape in Pol and Nef does not
substan-tially impair viral RC, the B*13-driven Nef-E24Q-Q107R double
mutant conferred a
⬎
40% reduction in Nef’s ability to
downregu-late cell surface HLA class I. This defect was nearly as profound as
that conferred by
⌬
Nef or the Nef-M20A mutation (which does
not occur naturally) in our assay system. Moreover, the inability of
Nef-E24Q-Q107R to downregulate HLA class I enhanced the
rec-ognition of infected cells by HIV-1-specific T cells
in vitro
. In other
words, this naturally occurring mutant combination, selected
in
vivo
under B*13-mediated CTL pressure, conferred a functional
cost to Nef’s immune evasion activity. Although attenuation of
Nef’s HLA class I downregulation function as a consequence of
CTL escape has been described previously (
35
,
36
), the mutations
required to confer these effects are rare
in vivo
. The
HLA-B*35-driven Nef-R75T and -Y85F escape mutations, when present
to-gether, reduced Nef’s HLA class I downregulation activity nearly
2-fold, but this combination occurs in
⬍
0.2% of natural HIV-1
sequences (
35
). Similarly, a Nef clone exhibiting
⬇
15% lower
HLA downregulation activity was isolated from an acutely
HIV-infected individual who subsequently controlled HIV-1 viremia
spontaneously to
⬍
2,000 copies/ml; however, this defect required
the selection of four Nef mutations in the context of two
indepen-dent HLA alleles (
36
). Although the E24Q-Q107R double mutant
is observed in only 4% of B*13-expressing individuals and
al-though this combination tends to arise late in infection, it
never-theless represents the most frequent
in vivo
CTL escape pathway
identified to date that significantly compromises this Nef
func-tion. Importantly, once this double mutant arises
in vivo
, its HLA
downregulation defect would serve to boost the levels of viral
epitopes complexed with all HLA-A and HLA-B alleles expressed
by the individual (not just those restricted by HLA-B*13), thus
rendering infected cells more susceptible to recognition by
nu-merous HIV-1-specific T cells (or at least those specific for viral
epitopes that had not yet developed CTL escape mutations). In
fact, we hypothesize that the reason this double mutant is
ob-served only rarely
in vivo
is because of its substantial functional
cost.
Several limitations of the study merit comment. First, we
fo-cused on escape in Gag, Pol, and Nef because all known
B*13-FIG 6Replication capacities of NL4.3 viruses encoding B*13-associated polymorphisms in Nef. (A) Representative growth curves of single and double Nef mutants, expressed as the fold increase in infected cells from the day 2 value. (B) NL4.3-normalized RC values of each Nef mutant, obtained from two independent experiments, each containing triplicate measurements. Histograms and error bars denote the means and standard deviations, respectively.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.138.452.64.257.2]restricted epitopes lie in these proteins. However, undiscovered
B*13 epitopes are likely to exist in other regions, as indicated by
the existence of B*13-associated polymorphisms in gp41, Tat, Vif,
and Vpr (
39
), and their impact on HIV-1 replication and protein
function remains unknown. Second, due to the relatively large
number of mutants assessed here, all replication capacity assays
were performed in an immortalized GFP reporter T cell line.
However, the magnitude of replication defects observed for
Gag-I437L and Gag-I147L corroborated previous results in PBMCs
(
27
,
28
), indicating that RC measurements in our assay system are
generally representative of those in other cell types. Third, due to
insufficient PBMCs from B*13
⫹individuals, we were unable to
fully characterize the novel GI8 epitope in p24
Gagalthough its
bioinformatically guided discovery (combined with reduced
pre-dicted binding affinities of its escape variants) supports it as such.
Fourth, we assessed the impact of Nef-E24Q and/or Nef-Q107R
on only three of Nef’s
in vitro
functions (viral replication and
CD4/HLA class I downregulation), but the impact of Nef-E24Q
and Nef-Q107R on other Nef functions, including upregulation of
HLA class II invariant chain (CD74) (
61
), enhancement of virion
infectivity (
62
), and alteration of T cell receptor signaling (
63
),
remains to be determined. Moreover, to evaluate the effect of Nef
FIG 7Impact of B*13-associated polymorphisms on Nef-mediated CD4 and HLA class I downregulation. (A and B) Representative flow plots of CD4 (A) and HLA class I (B) downregulation for uninfected cells and cells infected with wtNL4.3,⌬Nef, and NefM20Acontrol viruses. Median fluorescence intensities (MFIs)
of CD4 and HLA class I expression (yaxis) are shown for GFP⫹(HIV-infected; red gates) versus GFP⫺(HIV-1 uninfected; black gates) cells. (C and D) Impact of control viruses and those harboring B*13-associated Nef polymorphisms on CD4 and HLA-A*02 downregulation, normalized to wtNL4.3 levels. Receptor downregulation values were obtained from a minimum of three independent experiments, each containing one measurement. Histograms and error bars denote the means and standard deviations, respectively. *,Pⱕ0.01. (E) Detection of Nef, GFP, and-actin protein expression by Western blotting in GXR-A*02 reporter T cells infected with NL4.3 viruses encoding control or mutant Nef sequences.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:11.585.136.452.67.507.2]mutations on HLA downregulation and its consequences for T
cell-mediated recognition of infected cells, we used HLA-A*02
and an FK10
Gag-specific TCR as representative indicators of this
process. Since HLA-B molecules may generally be more resistant
to Nef-mediated downregulation than HLA-A molecules (
64
) and
since the impact of Nef-mediated HLA downregulation may be to
some extent HIV gene and/or epitope specific (
65–67
),
recogni-tion of other CTL epitopes might differ from that observed here
for A*02-FK10
Gag. However, using a transient-transfection
sys-tem, we previously demonstrated a strong correlation between the
ability of natural Nef sequences to downregulate HLA-A*02 and
HLA-B*07 (
34
), and Gag-specific CTL in general may be less
sus-ceptible to Nef-mediated HLA downregulation than CTL
target-ing other HIV-1 proteins (
67
), possibly due to presentation of Gag
epitopes before Nef-mediated HLA downregulation is complete
(
68
). Together, these observations suggest that our luciferase
re-porter T cell approach should be broadly representative of CTL
recognition and may be a conservative measure of Nef function.
Finally, while we have demonstrated enhanced recognition of
target cells infected with Nef-E24Q-Q107R mutant virus using a
reporter T cell assay, we have not performed inhibition studies or
assessed other effector functions directly with primary CTL
clones.
The mechanism whereby Nef-E24Q and -Q107R together
con-fer such a profound HLA downregulation defect requires further
study. While these codons have not been explicitly identified as
being critical for Nef function, both lie within or adjacent to
im-portant motifs. Nef-E24 is located in the N-terminal alpha helix
within a region (amino acids 17 to 26) that, when deleted and
engineered along with a V10E substitution, renders Nef defective
for HLA downregulation (
69
); it also lies four codons downstream
of M20, which, when artificially mutated to alanine, produces a
severe HLA downregulation defect (
50
). Nef-Q107 lies adjacent to
arginine residues R105/R106 that are essential for several Nef
functions, including dimerization (
70–72
); it is therefore possible
that the creation of a triple arginine (RRR
105–107) via a Q107R
substitution may alter this protein-protein interface. Mutations at
the adjacent residue 106 (e.g., R106K and R106L) also confer
modest to severe HLA class I downregulation defects (
73
).
More-over, a recent crystal structure of Nef in complex with HLA class I
suggests that Nef codons 24 and 107 lie nearby and within
(respec-tively) neighboring antiparallel alpha helices, suggesting that their
presence in combination could modulate interactions between
these helices and possibly impact HLA downregulation function
(
74
). Further studies will be required to test this hypothesis and
identify compensatory pathways in natural sequences.
Taken together with the previous literature, our results suggest
that B*13-mediated viremia control is achieved via multiple
mechanisms. The major mechanism, demonstrated by other
stud-ies, is likely to be effective sustained targeting of CTL epitopes in
multiple HIV-1 proteins (
11
,
21
). Fitness costs of the resulting
viral escape mutations, which presumably limit the net viral
“ben-efit” gained from evading the CTL response, also likely contribute.
Specifically, B*13 escape mutations may impair HIV-1 by two
distinct mechanisms: by reducing Gag fitness and by dampening
Nef’s immune evasion function by compromising its HLA
down-regulation activity. Functional costs of CTL escape in Nef have
been reported by others (
35
,
36
,
75
); our results extend these
ob-servations by suggesting that such costs may be biologically
rele-vant. More broadly, our study highlights the potential utility of
HIV-1 T cell vaccines designed to target vulnerable epitopes in Nef
where escape mutations impair its viral immune evasion activity.
ACKNOWLEDGMENTS
We thank Denis Chopera for engineering the pNL4.3-⌬nef vector used in the homologous recombination procedure, Takamasa Ueno for provid-ing control mutant Nef plasmids, Bemuluyigza Baraki for generatprovid-ing the GXR-A*02 cell line, and Mario Ostrowski and Brad Jones for providing the 5B2 CTL clone used to generate the FK10Gag-specific TCR. We thank
Chanson Brumme for assistance with data analysis. We also thank Mina John, Simon Mallal, P. Richard Harrigan, Martin Markowitz, Bruce Walker, Heiko Jessen, Evan Wood, Kanna Hayashi, and M.-J. Milloy for specimen and/or data access.
This work was supported by operating grants from the Canadian In-stitutes for Health Research (CIHR) (MOP-93536 and HOP-115700 to Z.L.B. and M.A.B.). The VIDUS and ACCESS studies were supported by the U.S. National Institutes of Health (VIDUS, R01DA011591 and U01DA038886; ACCESS, R01DA021525). This research was undertaken, in part, thanks to funding from the Canada Research Chairs program through a Tier 1 Canada Research Chair in Inner City Medicine which supports Evan Wood. X.T.K. was supported by a Master’s Scholarship from the CIHR Small Health Organizations Partnership Program (SHOPP), in partnership with the Canadian Association for HIV Re-search in partnership with Bristol-Myers Squibb Canada and ViiV Healthcare, and currently holds a CIHR Frederick Banting and Charles Best Canada Graduate Scholarship (doctoral). A.O. was funded by the Instituto de Salud Carlos III (PI12/00529). M.A.B. holds a Canada Re-search Chair (Tier 2) in Viral Pathogenesis and Immunity from the Can-ada Research Chairs Program. Z.L.B. was supported by a CIHR New In-vestigator Award and currently holds a Scholar Award from the Michael Smith Foundation for Health Research.
The funders of this study played no role in determining the content of the manuscript or the authors’ decision to publish.
REFERENCES
1.Kaslow RA, Carrington M, Apple R, Park L, Munoz A, Saah AJ, Goedert JJ, Winkler C, O’Brien SJ, Rinaldo C, Detels R, Blattner W, Phair J, Erlich H, Mann DL.1996. Influence of combinations of human major histocompatibility complex genes on the course of HIV-1 infection. Nat Med2:405– 411.http://dx.doi.org/10.1038/nm0496-405.
2.Saah AJ, Hoover DR, Weng S, Carrington M, Mellors J, Rinaldo CR, Jr, Mann D, Apple R, Phair JP, Detels R, O’Brien S, Enger C, Johnson P, Kaslow RA.1998. Association of HLA profiles with early plasma viral load, CD4⫹cell count and rate of progression to AIDS following acute HIV-1 infection. Multicenter AIDS cohort study. AIDS12:2107–2113.
FIG 8Impact of the Nef double mutant E24Q-Q107R on recognition of HIV-infected target cells by HIV-specific effector T cells. Representative data de-picting a negative correlation between the Nef-mediated ability to downregu-late HLA-A*02 on GFP reporter target cells infected with control and mutant Nef viruses and TCR-driven luciferase signal in A*02-FK10Gag-specific Jurkat
effector cells following coculture (Spearman’sR,⫺0.88;P⫽0.03).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:12.585.91.233.69.192.2]3.Tang J, Costello C, Keet IP, Rivers C, Leblanc S, Karita E, Allen S, Kaslow RA.1999. HLA class I homozygosity accelerates disease progres-sion in human immunodeficiency virus type 1 infection. AIDS Res Hum Retroviruses15:317–324.http://dx.doi.org/10.1089/088922299311277. 4.Carrington M, O’Brien SJ.2003. The influence of HLA genotype on
AIDS. Annu Rev Med 54:535–551. http://dx.doi.org/10.1146/annurev .med.54.101601.152346.
5.Gao X, Bashirova A, Iversen AK, Phair J, Goedert JJ, Buchbinder S, Hoots K, Vlahov D, Altfeld M, O’Brien SJ, Carrington M.2005. AIDS restriction HLA allotypes target distinct intervals of HIV-1 pathogenesis. Nat Med11:1290 –1292.http://dx.doi.org/10.1038/nm1333.
6.Pereyra F, Jia X, McLaren PJ, Telenti A, de Bakker PI, Walker BD, Ripke S, Brumme CJ, Pulit SL, Carrington M, Kadie CM, Carlson JM, Heckerman D, Graham RR, Plenge RM, Deeks SG, Gianniny L, Craw-ford G, Sullivan J, Gonzalez E, Davies L, Camargo A, Moore JM, Beattie N, Gupta S, Crenshaw A, Burtt NP, Guiducci C, Gupta N, Gao X, Qi Y, Yuki Y, Piechocka-Trocha A, Cutrell E, Rosenberg R, Moss KL, Lemay P, O’Leary J, Schaefer T, Verma P, Toth I, Block B, Baker B, Rothchild A, Lian J, Proudfoot J, Alvino DM, Vine S, Addo MM, Allen TM, et al.
2010. The major genetic determinants of HIV-1 control affect HLA class I peptide presentation. Science330:1551–1557.http://dx.doi.org/10.1126 /science.1195271.
7.Kiepiela P, Leslie AJ, Honeyborne I, Ramduth D, Thobakgale C, Chetty S, Rathnavalu P, Moore C, Pfafferott KJ, Hilton L, Zimbwa P, Moore S, Allen T, Brander C, Addo MM, Altfeld M, James I, Mallal S, Bunce M, Barber LD, Szinger J, Day C, Klenerman P, Mullins J, Korber B, Coovadia HM, Walker BD, Goulder PJ.2004. Dominant influence of HLA-B in mediating the potential co-evolution of HIV and HLA. Nature
432:769 –775.http://dx.doi.org/10.1038/nature03113.
8.Naruto T, Gatanaga H, Nelson G, Sakai K, Carrington M, Oka S, Takiguchi M.2012. HLA class I-mediated control of HIV-1 in the Japanese population, in which the protective B*57 and HLA-B*27 alleles are absent. J Virol86:10870 –10872.http://dx.doi.org/10 .1128/JVI.00689-12.
9.Brumme ZL, John M, Carlson JM, Brumme CJ, Chan D, Brockman MA, Swenson LC, Tao I, Szeto S, Rosato P, Sela J, Kadie CM, Frahm N, Brander C, Haas DW, Riddler SA, Haubrich R, Walker BD, Harrigan PR, Heckerman D, Mallal S.2009. HLA-associated immune escape path-ways in HIV-1 subtype B Gag, Pol and Nef proteins. PLoS One4:e6687.
http://dx.doi.org/10.1371/journal.pone.0006687.
10. Tang J, Tang S, Lobashevsky E, Myracle AD, Fideli U, Aldrovandi G, Allen S, Musonda R, Kaslow RA.2002. Favorable and unfavorable HLA class I alleles and haplotypes in Zambians predominantly infected with clade C human immunodeficiency virus type 1. J Virol76:8276 – 8284.
http://dx.doi.org/10.1128/JVI.76.16.8276-8284.2002.
11. Honeyborne I, Prendergast A, Pereyra F, Leslie A, Crawford H, Payne R, Reddy S, Bishop K, Moodley E, Nair K, van der Stok M, McCarthy N, Rousseau CM, Addo M, Mullins JI, Brander C, Kiepiela P, Walker BD, Goulder PJ.2007. Control of human immunodeficiency virus type 1 is associated with HLA-B*13 and targeting of multiple Gag-specific CD8⫹ T-cell epitopes. J Virol 81:3667–3672. http://dx.doi.org/10.1128/JVI .02689-06.
12. Li S, Jiao H, Yu X, Strong AJ, Shao Y, Sun Y, Altfeld M, Lu Y.2007. Human leukocyte antigen class I and class II allele frequencies and HIV-1 infection associations in a Chinese cohort. J Acquir Immune Defic Syndr
44:121–131.http://dx.doi.org/10.1097/01.qai.0000248355.40877.2a. 13. Zhai S, Zhuang Y, Song Y, Li S, Huang D, Kang W, Li X, Liao Q, Liu
Y, Zhao Z, Lu Y, Sun Y.2008. HIV-1-specific cytotoxic T lymphocyte (CTL) responses against immunodominant optimal epitopes slow the progression of AIDS in China. Curr HIV Res6:335–350.http://dx.doi.org /10.2174/157016208785132473.
14. Jia M, Hong K, Chen J, Ruan Y, Wang Z, Su B, Ren G, Zhang X, Liu Z, Zhao Q, Li D, Peng H, Altfeld M, Walker BD, Yu XG, Shao Y.2012. Preferential CTL targeting of Gag is associated with relative viral control in long-term surviving HIV-1 infected former plasma donors from China. Cell Res22:903–914.http://dx.doi.org/10.1038/cr.2012.19.
15. Gonzalez-Galarza FF, Takeshita LY, Santos EJ, Kempson F, Maia MH, da Silva AL, Teles e Silva AL, Ghattaoraya GS, Alfirevic A, Jones AR, Middleton D.2015. Allele frequency net 2015 update: new features for HLA epitopes, KIR and disease and HLA adverse drug reaction associa-tions. Nucleic Acids Res43:D784 –D788.http://dx.doi.org/10.1093/nar /gku1166.
16. Brumme ZL, Brumme CJ, Chui C, Mo T, Wynhoven B, Woods CK,
Henrick BM, Hogg RS, Montaner JS, Harrigan PR. 2007. Effects of human leukocyte antigen class I genetic parameters on clinical outcomes and survival after initiation of highly active antiretroviral therapy. J Infect Dis195:1694 –1704.http://dx.doi.org/10.1086/516789.
17. Wei Z, Liu Y, Xu H, Tang K, Wu H, Lu L, Wang Z, Chen Z, Xu J, Zhu Y, Hu L, Shang H, Zhao G, Kong X.2015. Genome-wide association studies of HIV-1 host control in ethnically diverse Chinese populations. Sci Rep5:10879.http://dx.doi.org/10.1038/srep10879.
18. O’Brien SJ, Gao X, Carrington M.2001. HLA and AIDS: a cautionary tale. Trends Mol Med 7:379 –381. http://dx.doi.org/10.1016/S1471 -4914(01)02131-1.
19. Emu B, Sinclair E, Hatano H, Ferre A, Shacklett B, Martin JN, McCune JM, Deeks SG.2008. HLA class I-restricted T-cell responses may contrib-ute to the control of human immunodeficiency virus infection, but such responses are not always necessary for long-term virus control. J Virol
82:5398 –5407.http://dx.doi.org/10.1128/JVI.02176-07.
20. Antoni G, Guergnon J, Meaudre C, Samri A, Boufassa F, Goujard C, Lambotte O, Autran B, Rouzioux C, Costagliola D, Meyer L, The-odorou I.2013. MHC-driven HIV-1 control on the long run is not sys-tematically determined at early times post-HIV-1 infection. AIDS27:
1707–1716.http://dx.doi.org/10.1097/QAD.0b013e328360a4bd. 21. Harrer EG, Bergmann S, Eismann K, Rittmaier M, Goldwich A, Muller
SM, Spriewald BM, Harrer T.2005. A conserved HLA B13-restricted cytotoxic T lymphocyte epitope in Nef is a dominant epitope in HLA B13-positive HIV-1-infected patients. AIDS 19:734 –735. http://dx.doi .org/10.1097/01.aids.0000166099.36638.56.
22. Yusim K, Korber BTM, Brander C, Haynes BF, Koup R, Moore JP, Walker BD, Watkins DI (ed).2014. HIV molecular immunology 2014. Theoretical Biology and Biophysics, Los Alamos National Laboratory, Los Alamos, NM.http://www.hiv.lanl.gov/content/immunology/pdf/201 4/immuno2014.pdf.
23. Martinez-Picado J, Prado JG, Fry EE, Pfafferott K, Leslie A, Chetty S, Thobakgale C, Honeyborne I, Crawford H, Matthews P, Pillay T, Rousseau C, Mullins JI, Brander C, Walker BD, Stuart DI, Kiepiela P, Goulder P.2006. Fitness cost of escape mutations in p24 Gag in associa-tion with control of human immunodeficiency virus type 1. J Virol80:
3617–3623.http://dx.doi.org/10.1128/JVI.80.7.3617-3623.2006. 24. Crawford H, Prado JG, Leslie A, Hue S, Honeyborne I, Reddy S, van der
Stok M, Mncube Z, Brander C, Rousseau C, Mullins JI, Kaslow R, Goepfert P, Allen S, Hunter E, Mulenga J, Kiepiela P, Walker BD, Goulder PJ.2007. Compensatory mutation partially restores fitness and delays reversion of escape mutation within the immunodominant HLA-B*5703-restricted Gag epitope in chronic human immunodeficiency virus type 1 infection. J Virol81:8346 – 8351.http://dx.doi.org/10.1128/JVI .00465-07.
25. Brockman MA, Brumme ZL, Brumme CJ, Miura T, Sela J, Rosato PC, Kadie CM, Carlson JM, Markle TJ, Streeck H, Kelleher AD, Markowitz M, Jessen H, Rosenberg E, Altfeld M, Harrigan PR, Heckerman D, Walker BD, Allen TM.2010. Early selection in Gag by protective HLA alleles contributes to reduced HIV-1 replication capacity that may be largely compensated for in chronic infection. J Virol84:11937–11949.
http://dx.doi.org/10.1128/JVI.01086-10.
26. Schneidewind A, Brockman MA, Yang R, Adam RI, Li B, Le Gall S, Rinaldo CR, Craggs SL, Allgaier RL, Power KA, Kuntzen T, Tung CS, LaBute MX, Mueller SM, Harrer T, McMichael AJ, Goulder PJ, Aiken C, Brander C, Kelleher AD, Allen TM.2007. Escape from the dominant HLA-B27-restricted cytotoxic T-lymphocyte response in Gag is associated with a dramatic reduction in human immunodeficiency virus type 1 rep-lication. J Virol81:12382–12393.http://dx.doi.org/10.1128/JVI.01543-07. 27. Prado JG, Honeyborne I, Brierley I, Puertas MC, Martinez-Picado J, Goulder PJ.2009. Functional consequences of human immunodeficiency virus escape from an HLA-B*13-restricted CD8⫹T-cell epitope in p1 Gag protein. J Virol83:1018 –1025.http://dx.doi.org/10.1128/JVI.01882-08. 28. Boutwell CL, Carlson JM, Lin TH, Seese A, Power KA, Peng J, Tang Y,
Brumme ZL, Heckerman D, Schneidewind A, Allen TM.2013. Frequent and variable cytotoxic-T-lymphocyte escape-associated fitness costs in the human immunodeficiency virus type 1 subtype B Gag proteins. J Virol
87:3952–3965.http://dx.doi.org/10.1128/JVI.03233-12.
29. Wang YE, Li B, Carlson JM, Streeck H, Gladden AD, Goodman R, Schneidewind A, Power KA, Toth I, Frahm N, Alter G, Brander C, Carrington M, Walker BD, Altfeld M, Heckerman D, Allen TM.2009. Protective HLA class I alleles that restrict acute-phase CD8⫹T-cell re-sponses are associated with viral escape mutations located in highly
on November 7, 2019 by guest
http://jvi.asm.org/
served regions of human immunodeficiency virus type 1. J Virol83:1845– 1855.http://dx.doi.org/10.1128/JVI.01061-08.
30. Brumme ZL, Li C, Miura T, Sela J, Rosato PC, Brumme CJ, Markle TJ, Martin E, Block BL, Trocha A, Kadie CM, Allen TM, Pereyra F, Heckerman D, Walker BD, Brockman MA.2011. Reduced replication capacity of NL4-3 recombinant viruses encoding reverse transcriptase-integrase sequences from HIV-1 elite controllers. J Acquir Immune Defic Syndr56:100 –108.http://dx.doi.org/10.1097/QAI.0b013e3181fe9450. 31. Brockman MA, Chopera DR, Olvera A, Brumme CJ, Sela J, Markle TJ,
Martin E, Carlson JM, Le AQ, McGovern R, Cheung PK, Kelleher AD, Jessen H, Markowitz M, Rosenberg E, Frahm N, Sanchez J, Mallal S, John M, Harrigan PR, Heckerman D, Brander C, Walker BD, Brumme ZL.2012. Uncommon pathways of immune escape attenuate HIV-1 inte-grase replication capacity. J Virol 86:6913– 6923. http://dx.doi.org/10 .1128/JVI.07133-11.
32. Yoon K, Jeong JG, Yoon H, Lee JS, Kim S.2001. Differential effects of primary human immunodeficiency virus type 1 nef sequences on down-regulation of CD4 and MHC class I. Biochem Biophys Res Commun284:
638 – 642.http://dx.doi.org/10.1006/bbrc.2001.5026.
33. Mwimanzi P, Markle TJ, Martin E, Ogata Y, Kuang XT, Tokunaga M, Mahiti M, Pereyra F, Miura T, Walker BD, Brumme ZL, Brock-man MA, Ueno T.2013. Attenuation of multiple Nef functions in HIV-1 elite controllers. Retrovirology10:1.http://dx.doi.org/10.1186 /1742-4690-10-1.
34. Mann JK, Byakwaga H, Kuang XT, Le AQ, Brumme CJ, Mwimanzi P, Omarjee S, Martin E, Lee GQ, Baraki B, Danroth R, McCloskey R, Muzoora C, Bangsberg DR, Hunt PW, Goulder PJ, Walker BD, Har-rigan PR, Martin JN, Ndung’u T, Brockman MA, Brumme ZL.2013. Ability of HIV-1 Nef to downregulate CD4 and HLA class I differs among viral subtypes. Retrovirology 10:100. http://dx.doi.org/10.1186/1742 -4690-10-100.
35. Ueno T, Motozono C, Dohki S, Mwimanzi P, Rauch S, Fackler OT, Oka S, Takiguchi M.2008. CTL-mediated selective pressure influences dy-namic evolution and pathogenic functions of HIV-1 Nef. J Immunol180:
1107–1116.http://dx.doi.org/10.4049/jimmunol.180.2.1107.
36. Kuang XT, Li X, Anmole G, Mwimanzi P, Shahid A, Le AQ, Chong L, Qian H, Miura T, Markle T, Baraki B, Connick E, Daar ES, Jessen H, Kelleher AD, Little S, Markowitz M, Pereyra F, Rosenberg ES, Walker BD, Ueno T, Brumme ZL, Brockman MA.2014. Impaired Nef function is associated with early control of HIV-1 Viremia. J Virol88:10200 –10213.
http://dx.doi.org/10.1128/JVI.01334-14.
37. Garcia JV, Miller AD.1991. Serine phosphorylation-independent down-regulation of cell-surface CD4 by nef. Nature350:508 –511.http://dx.doi .org/10.1038/350508a0.
38. Schwartz O, Marechal V, Le Gall S, Lemonnier F, Heard JM.1996. Endocytosis of major histocompatibility complex class I molecules is in-duced by the HIV-1 Nef protein. Nat Med2:338 –342.http://dx.doi.org /10.1038/nm0396-338.
39. Carlson JM, Brumme CJ, Martin E, Listgarten J, Brockman MA, Le AQ, Chui CK, Cotton LA, Knapp DJ, Riddler SA, Haubrich R, Nelson G, Pfeifer N, Deziel CE, Heckerman D, Apps R, Carrington M, Mallal S, Harrigan PR, John M, Brumme ZL.2012. Correlates of protective cel-lular immunity revealed by analysis of population-level immune escape pathways in HIV-1. J Virol86:13202–13216.http://dx.doi.org/10.1128 /JVI.01998-12.
40. Martin E, Carlson JM, Le AQ, Chopera DR, McGovern R, Rahman MA, Ng C, Jessen H, Kelleher AD, Markowitz M, Allen TM, Milloy MJ, Carrington M, Wainberg MA, Brumme ZL.2014. Early immune adap-tation in HIV-1 revealed by population-level approaches. Retrovirology
11:64.http://dx.doi.org/10.1186/s12977-014-0064-1.
41. Brumme ZL, Brumme CJ, Carlson J, Streeck H, John M, Eichbaum Q, Block BL, Baker B, Kadie C, Markowitz M, Jessen H, Kelleher AD, Rosenberg E, Kaldor J, Yuki Y, Carrington M, Allen TM, Mallal S, Altfeld M, Heckerman D, Walker BD.2008. Marked epitope- and allele-specific differences in rates of mutation in human immunodeficiency type 1 (HIV-1) Gag, Pol, and Nef cytotoxic T-lymphocyte epitopes in acute/ early HIV-1 infection. J Virol82:9216 –9227.http://dx.doi.org/10.1128 /JVI.01041-08.
42. Pond SL, Frost SD, Muse SV.2005. HyPhy: hypothesis testing using phylogenies. Bioinformatics 21:676 – 679. http://dx.doi.org/10.1093 /bioinformatics/bti079.