0022-538X/92/105988-11$02.00/0
Copyright © 1992,American Societyfor Microbiology
Two
Cellular
Single-Strand-Specific
DNA-Binding Proteins Interact
with
Two
Regions of the Bovine Papillomavirus
Type
1
Genome,
Including the Origin of DNA Replication
CORNELIAHABIGER, GERTRAUD STELZER, URSULA SCHWARZ,
ANDERNST-LUDWIGWINNACKER*
Institutfiir Biochemie der Universitat Munchen im Max-Planck-Institut fiir Biochemie, AmKlopferspitz 18a, D-8033Martinsried bei Munchen, Germany
Received 28 April1992/Accepted 1 July 1992
We haveidentified andpurified to nearhomogeneitytwospecific single-stranded DNA-binding factors (SPSF Iand II) with molecular masses of 42 and39 kDa,
respectively,
fromcalf thymus. Gel retentionanalysis and competition experiments demonstrate that the ubiquitous proteins SPSF I and II specifically interact with single-strandedDNAderivedfromtheminimalin vitroorigin of replicationof bovinepapillomavirustype 1 and a regionoftheviral genome proposed to be involved in plasmid maintenance. Bovine papillomavirus type 1 proteins do not interfere with DNA binding of SPSF I and II. Theexact locationofthebinding domainsof SPSF IandIIonthe DNA has been determined by methylation interference and T4 DNA polymerase footprinting. A potential cellular binding site for SPSF I and II is themajor
promoter (P2) of the human c-myc gene.The replication of the circular double-stranded bovine papillomavirustype 1 (BPV-1)genomeissubjecttocomplex control mechanisms. They guarantee the maintenance of about200copies per cell, all of which arereplicated extra-chromosomally once per cell cycle (5, 13). Unlike other DNAviruses, BPV-1 does notoverride cellular replication controls. Its bidirectional mode of replication hence pro-vides an excellent system with which to study the mecha-nismspreventing the reinitiation ofreplication from an origin that hasalready beenengaged in this process.
The minimal origin of BPV-1 DNA replication has been mapped recently to aregion between nucleotides 7900 and 50, containingan18-bppalindrome andanAT-rich sequence (39, 46). Earlier studies of the replicative intermediates of BPV-1 and DNase I-hypersensitive sites had suggested a
region between nucleotides 6600 and 7300 (30, 43). This region also contains one of the two plasmid maintenance sequences (PMS1; domain 1 and 2), which have been sug-gestedtofunctionascis-activeregulatoryelements support-ing extrachromosomal replication ofrecombinant plasmids in the presence of viral proteinsprovided in trans (18, 20). Other studies had shownastretch aroundbp
7730,
+100bp, designatedthe 2D-origin, tobe active as anorigin for viral plasmidreplication (45). A 69% subgenomic fragment(from nucleotides 6958 to 4450), which divides the BPV-1 early regions, andaportion of the long control region from the late regions and the other part of the long control region is sufficient forcellulartransformation and extrachromosomal replication oftheviral DNA. However, the transformation and replication efficiencies are reproducibly lower (25 to 50%) than those observed for the full-length BPV-1 genome(15,
17,
32).
Of all viralproteins, two, designated El (68 kDa) and E2 (48 kDa), have been shown to be both necessary and sufficient forBPV-1replication (19,38). Phosphoprotein El forms stable complexes with E2 (21, 23). Clusters of E2 binding sitesarelocatedin the vicinity of the important early
* Corresponding author.
viral promoters. These sites normallyfunctionas transcrip-tional enhancers, either alone or in cooperation with other binding sites for cellularfactors, suchas SP1(14).
An exactinterpretation ofexperiments aimedat elucidat-ing the functions of El and E2 is complicated by the observation that an El fusion protein (RecA-El) interacts with the functionaloriginofreplication(nucleotides 7900to
50)and also withafragment containingthe2D-originandan E2-responsive element 1 (E2RE1) contained therein (42). Moreover, the E1-E2complexalso binds to a BPV-1 frag-mentcontaining E2-responsiveelements(E2bindingsites 1 and2, distinct fromE2RE1)andtoPMS1, aswellas tothe fragment containing the2D-origin andE2RE1 (21, 23).
Inthis report,wedescribe the identification,isolation, and characterization oftwocellularproteins, designatedSPSF I (42kDa)and II(39 kDa), forspecific single-stranded DNA-binding factors Iand II. These proteins, isolated from calf thymus, interact specificallywithsingle-stranded DNA de-rived from theminimal in vitrooriginofreplicationand also with sequences of domain I of PMS1. Theexpressionof viral proteinsdoesnotinterfere withprotein binding.Thesites of SPSFIandIIinteraction withDNAhave beenmapped by methylationinterferenceand T4DNApolymerase footprint-ing. SPSFIand IIarethefirstexamplesofcellularproteins involved in interactions with DNA regions required for BPV-1DNAreplication. Theyareexpressedinavarietyof cell types and in different
species
andarethereforegeneral factors inhighereukaryotes.MATERIALSANDMETHODS
Cells.HeLa, NIH 3T3, C127, and CB1 cellswere grown asmonolayersin Dulbecco's modifiedEagle'smedium sup-plemented with 5% fetal calfserum. CB1 cells are mouse C127 cells which we obtained as a stably transformed cell line after calcium phosphate coprecipitation with plasmid pMLBPV(19).The BPV-1 genome ismaintained extrachro-mosomallyinaconstantnumber ofapproximately100to200 copies per cell, as proven by hybridization of the BPV-1 genomeagainstHirtextracts(9)fromcloned andpropagated
5988
on November 9, 2019 by guest
http://jvi.asm.org/
CELLULAR DNA-BINDING PROTEINS INTERACT WITH BPV-1 5989 CB1 foci.
F126,
an Epstein-Barrvirus (EBV)-negativeBur-kitt's lymophoma cell line, and IARC 549, an EBV-trans-formed human lymphoblastoid B-cell line, were maintained assuspensioncultures in RPMI 1640 medium containing8% fetal calfserum.
Preparation of cell extracts. Nuclear and cytoplasmic ex-tractswerepreparedfrom subconfluent cells by themethod of
Dignam
etal.(4)
with someimprovements. Specifically, Nonidet P-40(0.5%)
was added tobuffer A, which caused thecells,
in particular NIH 3T3, C127, and CB1, to swell more readily. Dounce homogenization thus could beavoided,
and thenucleicould bereleased by vortexing. The nuclear extraction bufferCcontained 0.35 M instead of 0.43 MNaCl.Oligonucleotides. Thesynthetic oligonucleotides (u, upper
strand; 1,
lower strand) representing the PMS1 domain 1region
(P-u,
P-i,
andPk3-i)
and the BPV-1 origin of replica-tion(ORI-u
and ORI-l) are shown in Fig. 1A. Oligonucleo-tideBE-i correspondstothe lower strandof the BPV-1 distal enhancer from nucleotides 4390 to 4440 (44). PyE-l repre-sents the lower strand of the B enhancer of polyomavirus from nucleotides 5160 to 5210 (27). Oligonucleotide EF2 contains anNFl-binding
site from adenovirus FL, nucleo-tides 166to 151(5'-GATCAACACAACACAGTIGGCACA
GTGCCAAAAGCCCCG-3'). Oligonucleotide
lex2 repre-sents the lower strand of the promoter region of the lexA gene ofSaccharomyces
cerevisiae (5'-CTCGAGTACTGTA TGTACATACAGTACTCGACTCGA-3'). All oligonucleo-tides werepurified
on denaturing urea-polyacrylamide gels before use.Gel retention and competition experiments. Gel retention assayswere
performed
asdescribedbySchneider et al. (34).Briefly,
a reaction mix of 30j.l
typically contained 5 to 20 fmolofsingle-stranded
oligonucleotides 5'-endlabeled with[-y-32P]ATP by
T4 polynucleotide kinase, 1 to 2 ,ug ofpoly[d(I-C)],
and up to 6 ,ug ofprotein. Thereaction buffer contained 25 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid, pH 7.9),5 mMdithiothreitol(DTT), 1 mMEDTA, 10% glycerol, and 150 mM NaCl. The reaction mixturewasadjusted
to10 mMMgCl2and incubated on ice for 5 to 10 min. Competition analyseswere usually carried outwitha50- to100-fold molarexcessofcompetitor DNA.DNA-protein complexes
wereseparated from free DNA on 8%polyacrylamide
gelsandvisualized by autoradiography of thedriedgels.Methylationinterferencefootprinting. 5'-end-labeled oligo-nucleotides were methylated with dimethyl sulfoxide as described
by
Maxam and Gilbert (22) and incubated withpartially
purified C127 extracts as for the gel retention assays. DNA-protein complexes and free oligonucleotides werevisualizedby autoradiographyand cut out from the gel. DNA was eluted from the gel by isotachophoresis (26), treated withphenol-chloroform,
and ethanol precipitated. Aftertreatmentwith piperidin, theproducts were analyzedon7to 10%urea denaturing sequencing gels.
T4DNA polymerase footprinting. 5'-end-labeled
oligonu-cleotides, protein
extract(asindicated in Results), T4 DNApolymerase
buffer [16.6 mM (NH4)2SO4, 67 mM Tris-HCI(pH 8.8),
6.7 mMMgCl2,
10mM2-mercaptoethanol, 6.7 ,uMEDTA,
1 to2 ,ug ofpoly[d(A-T)],
and1 to2 ,ug ofpoly[d(I-C),
andcompetitor
DNA,ifneeded,wereincubated on ice for 10 min. Aftercompletionof thebinding reaction, 10 U of T4 DNApolymerase (Biolabs)was added, and the reaction mixturewasincubatedatroomtemperaturefor 15 min. Theproteins
were removedbyphenol extraction, and the DNAwas ethanol precipitated. The reaction products were ana-lyzed on 10% polyacrylamide sequencinggels.
Purification of SPSF I and SPSFII from calfthymus. All purification steps were carried out at
4°C.
Whole-cell ex-tracts from calf thymus were prepared by cutting 500 g of tissue into pieces, which were then suspended in 1 liter of buffer I(0.01%Nonidet P-40, 25 mM HEPES[pH7.9],
5mM MgCl2, 50 mMNaCl, 250 mM sucrose, 5 mMDPT, 0.5 mM protease inhibitor phenylmethylsulfonyl fluoride [PMSF], 1 ,uMleupeptin, 1% Trasylol) and treated in a Waring blender (adjusted to low, for 2min). The crude extract was cleared by centrifugation at 8,000 rpm for 10min. The supernatant was filtered through cotton cloths and added to DEAE-Sepharose CL-6B(Pharmacia) equilibrated with buffer 11 (25 mM HEPES [pH 7.9], 1 mM EDTA, 5 mM DTT, 10% glycerol), with the addition of 50 mM NaCI. This mixture wasstirred slowly for 6 h. Unbound proteins were removed by filtration, and the resin with the bound proteins (including SPSF I and II) was washed with 5 volumes of buffer II-50 mM NaCl. The washed DEAE-Sepharose was added to a column, and the proteins were eluted in a single step with 300 mM NaCl in bufferII. Fractions containing SPSF I and II were monitored by gel retention analysis. Active fractions were dialyzed against buffer II with 50 mMNaCland loaded onto a MonoQ column (HR 16/10; Pharmacia) equilibrated with buffer II-50 mM NaCI in a fast protein liquid chroma-tography(FPLC) system. After the column was washed with 10 bed volumes of bufferII-50 mMNaCl, the proteins were elutedby a linear gradient (50 mM to 1 M) ofNaCI. SPSF I-and II-containing fractions (centered at 550 mM NaCI)were pooled and dialyzed against buffer II with 150 mMNaCI or diluted with buffer II without salt to obtain a final concen-tration of 150 mMNaCl.DNA affinity chromatography columns were prepared by coupling the single-stranded oligonucleotide P-1 to CNBr-activated Sepharose (Pharmacia) following the instructions of the manufacturer. The column was connected to the FPLC system and equilibrated with buffer II containing 10 mMMgCl2 and 150 mM NaCl. The active MonoQ fractions, adjusted to 150 mM NaCl, were mixed with MgCl2 (end concentration, 10 mM) and with 1 ,ugof poly[d(I-C)] per ml and loaded onto the DNA affinity column. Washing with 10 volumes of buffer II-10 mM MgC12-150 mM NaCl was followed by elution with 5 bed volumes of a linear gradient (150 mM to 1 M) of NaCl. This protocol did not allow separation of the two factors because SPSF I eluted at about 950 mMNaCl and SPSF II eluted at about 850 mM NaCl. The pooled active fractions could be stored at
4°C
for up to 4 months without losing their binding activity. Aliquots were stored at -80°C.UVcross-linking. Binding reactions were performed essen-tially as described for gel retention except that the amount of proteinextract was scaled up 5- to 10-fold and the amount of 5'-end-labeled radioactive oligonucleotides was scaled up 3-to5-fold. The reaction mixture was placed on a transillumi-nator (254 nm, 8,000 p.W/cm2), irradiated at room tempera-turefor 4min, mixed with Laemmli buffer and separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel electro-phoresis (12) together with suitable protein size markers. Themolecular masses of the free oligonucleotides and the UV-cross-linked DNA-protein complexes were determined by comparison of Coomassie-stained dried gels and their autoradiographs (exposure to X-ray film [KodakXR5] with an intensifying screen at -80°C for 2 to 3 days).
Renaturation of proteins from
SDS-polyacrylamide
gels. Protein extract (2 to 3 mg) was loaded on a preparative 10% VOL. 66,1992on November 9, 2019 by guest
http://jvi.asm.org/
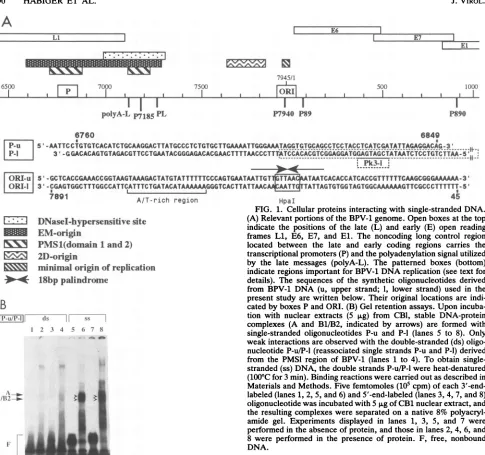
A
7945/1
I I I
AOI
I I6500 700 7500 500 1000
I I
II
I
polyA-L P7185 PL
I I
P7940 P89 P890
6760 6849
~ ~ ~ 1
|P-U '5 -AATTCCTGTGTCACATCTGCAAGGACTTATGCsC TCTGTGCTTGAAAATTGGGAAATAGGTGTGCAGCCTCCTACCTCATCGATATTAGAGGACAG-3 ...
IP-l
3'-CGGACACAGTGTAGACGT TCCTGAATACGGGAGACACGAACTTTTAACCCTT M[SCdAC eTC ; rXddYdii~e--KYyitcCTA_T
A-A'CCTTTAP1~~~~~~ ~ ~ ~
~~~~~~~~~~~~~~~~k-[ORI-U S'-GCTCACCGAAACCGGTAAGTAAAGACTATGTATTTTTTCCCAGTGAATAATTGT TAATCACACCATCACCGTTTTTTCAAGCGGGAAAAAA-3T ORll| 3'-CGAGTGGCTTTGGCCATTCATTTCTGATACATAAAAAAGGGTCACTTATTAAC TATTAGTGTGGTAGTGGCAAAAAAGTTCGCCCTTTTTT-5'
I I
I %RW a AfT-rich region
LIZ]
*lDNasel-hypersensitive
siteEM-origin
1v\!
PMS1(domain
1and2)
2D-origin1minimaloriginof
replication
*4
18bp palindrome
B
|I'-tL/l'-1| (kS ss 1
I2 3 4 5 6, 7 8
Am.. ~ ~ ..
F-1"~
~~if-HpaI
FIG. 1. Cellular proteins interacting with single-strandedDNA. (A) Relevant portions of theBPV-1 genome.Openboxesatthetop indicate the positions of the late (L) and early (E) open reading frames Li, E6, E7, and El. The noncoding long control region located between the late and early coding regions carries the transcriptionalpromoters(P)andthepolyadenylation signal utilized by the late messages (polyA-L). The patterned boxes (bottom) indicateregions important for BPV-1DNAreplication(seetextfor details). The sequencesof the synthetic oligonucleotides derived from BPV-1 DNA (u, upper strand; 1, lower strand) used in the present studyarewrittenbelow.Their original locations are indi-catedbyboxes PandORI.(B) Gel retentionassays.Upon incuba-tion with nuclear extracts (5 p.g) from CB1, stable DNA-protein complexes (A and B1/B2, indicated by arrows) are formed with single-stranded oligonucleotides P-u and P-l (lanes 5 to 8). Only weak interactionsareobservedwiththe double-stranded(ds) oligo-nucleotideP-u/P-l (reassociated single strandsP-uandP-1)derived from the PMSIregion of BPV-1 (lanes 1 to 4). To obtain single-stranded(ss) DNA, thedouble strandsP-u/P-lwereheat-denatured (100°C for3min). Binding reactionswerecarriedout asdescribedin Materialsand Methods. Five femtomoles(105cpm)of each 3'-end-labeled(lanes1,2, 5, and6)and5'-end-labeled(lanes3, 4, 7, and8) oligonucleotidewasincubated with5 p,g ofCB1nuclearextract,and theresulting complexeswereseparatedon anative8% polyacryl-amide gel. Experiments displayed in lanes 1, 3, 5, and 7 were
performedin the absence ofprotein,and those in lanes2, 4, 6,and 8 wereperformed in the presenceof protein. F, free, nonbound DNA.
polyacrylamide-SDS gel, parallel to one lane of marker
proteins. Following electrophoresis, the marker lane was
separatedfrom thegeland stained with Coomassie blue. The rest of the gel was cut into 18 equally spaced horizontal slices, with individual molecular weight fractions being indicated by the molecular weight standards in the Coomassie-stained markerlane. Each slice was suspended inelution buffer(50 mM Tris-HCl [pH 7.9], 0.1% SDS, 0.1
mg ofbovine serum albumin per ml, 1 mM DTT, 0.2 mM EDTA,0.1 mMPMSF, 2.5% glycerol) and eluted by rotation overnight at4°C.Afterremoval of thegel pieces by centrif-ugation, the solution was precipitated with
methanol-ace-tone (1:1) at -80°C for atleast 2 h or overnight. Proteins werepelleted by centrifugation at 15,000rpm and 4°C. The
air-driedproteinsweresolubilizedfirst in 3p,lof renaturation bufferI(0.1MTris-HCl[pH 7.5],8 Murea)and then diluted in125,ulof renaturation bufferII(20mMTris-HCl [pH 7.6], 10 mMKCl,2 mMDTT, 10 p.MPMSF). Renaturation of the
proteins requiredatleast 18 hat4°C.From15to20,ul of the protein solution was used for gel retention assays. The molecularweightsof SPSF I and II in activegelsliceswere calculated by comparison with the Coomassie-stained pro-tein marker lane.
RESULTS
Identification ofproteinsthat interact withsingle-stranded oligonucleotidesfrom BPV-1 DNA.Ourapproachtostudying and
identifying
putativecellular factors requiredfor BPV-1 DNAreplicationwas to search forproteins binding to the origin.The targetsofourefforts thus became domain1(D1) of the plasmid maintenance region PMS1, originally de-scribed by Lusky and Botchan (18, 20), and the minimal origin regionbetweenpositions 7911 and22,morerecently showntoberequiredfor BPV-1replicationin vivo(39)and in vitro(46). Thesetworegionsaredepicted inFig. 1Aand will be referredto asPMS1 forthePMS1Dlregionand ORI. . . .
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.71.556.58.513.2]CELLULAR DNA-BINDING PROTEINS INTERACT WITH BPV-1 5991 for the minimal
origin region.
The PMS1 D2regionwasnotstudied.
Inaninitial
experiment,
32P-labeleddouble-stranded DNAfragments (Fig.
1B, lanes 1to 4)from the PMS1 region and their heat-denatured counterparts (lanes 5 to 8) were incu-bated with anuclear extract from BPV-1-transformed C127 mouse cells(CB1 cells).
Onlyaweaknonspecific band was observed with the double-strandedfragments(Fig. 1B, lanes 2 and4),
while a strong triplet of retarded bands was observed with the denaturedfragments ingel retention gels. This indicated that only single-stranded DNA-specific fac-tors bind to thisparticular
region of the viral genome (Fig.1B,
lanes 6 and8).
The triplet bands (which can be recog-nized much better inFig.
2 and which subsequently were shown to result from the binding of two different proteins[see
below])
have beendesignated
A (for the upper band) andB1/B2
(for
the lowerdoublet).
The sequence
specificity
oftheseinteractionswasstudied [image:4.612.323.550.79.498.2]by
competition analysis. Single-stranded
oligonucleotides from the lower and upper strands of the PMS1 region (P-u[upper]
and P-l[lower];
96 nucleotides each; Fig. 1A), theorigin region
(ORI-u
and ORI-l; 100nucleotides each; Fig.1A),
and a number of control oligonucleotides were incu-bated withapartially purified
extractfrom calfthymus(seebelow).
Thecompeting oligonucleotides
were derived from the PMS1region (P-u
andP-l),
theorigin region (ORI-l), the upperstrand ofaregion
of the c-myc P2 promoter(MYC-u), the lower strand ofaregion
from the polyomavirus genome(PyE-l;
seeMaterials andMethods),
and the lower strand of the distal enhancer of BPV-1(BE-l;
see Materials andMethods).
In the case of the PMSI region (Fig. 2A), the lowerstrand(P-l)
displays
astrongretardationsignal (lane1) whichcanbeinhibitedby
itself(lane
2), thecomplementary upperstrand(P-u), ORI-l,
andMYC-u. Nocompetition was observed witholigonucleotides
PyE-l and BE-l. The upper strand(P-u) (Fig.
2B) displays
asignal
comparabletothat of the lower strand(P-l,
Fig. 2A) in strength, position, andcompetition
behavior. Bothcomplementarysingle strands of thisregion
thus possessbinding activity.The presence of bandsin
Fig.
2A,lane3,andFig. 2B, lane2,
which have a lowermobility
thanfree DNA (marked F)might
be taken as evidence for double-strand formation between P-u and P-l.Indeed,
the reactionconditions of thecompetition
experiments involving
complementary single strands would allow the formation of double strands. Since double-strandedfragments
donotpossessbindingactivity, it ispossible
that the observed absence ofashiftedsignalis not due to truecompetition
but rather the result of the inability of thenewly
formed double-strandedfragmenttobind pro-tein(Fig.
1B).
Thisinterpretation
canbe ruled out, however, becauseprotein-binding
single-stranded oligonucleotides such as ORI-l and MYC-u, under theseconditions, are not inhibitedby
theircorresponding
complementary strandslacking
aprotein-binding
site(data
not shown).In the case of the ORI
region,
only the oligonucleotide derived from the lower strand(Fig.
2C) displays a retarda-tionsignal which, again,
behaves like the oligonucleotides from thePMSlregion
describedabove, i.e.,it competes with itself and witholigonucleotides P-l,
P-u,andMYC-u but not withPyE-l
orBE-l. Thecorresponding oligonucleotidefrom the upper strand of the ORIregionisinactive and does not compete with itscomplementary
strand for the binding ofprotein (not
shown).
Similarbehavior is observed witholigonucleotidesderived from the c-myc P2 promoter. In thiscase(Fig. 2D), only the upper strand leads to the formation of shifted bands. The
A
F
I
com;tit=r- r_ =
conmpetitor -
='
-s, -c -. F:^p
c
F -I *_F
1 2 3 4 5 6 7 1 2 3 4 5 6 7
B
c}mpetitsr
P-YC-u
^'7
\ -- - = -,=
B-
~
~
ID
>
.-,R
-FF 1 2 3 4 5 6 7 1 2 3 4 5 6 7
FIG. 2. Three differentoligonucleotides arerecognized specifi-cally bySPSFI andII.Competitionanalysisrevealed highlyspecific interactionsofSPSFIand IIwiththe single-stranded oligonucleo-tides P-1 (A), P-u (B), ORI-I (C), and MYC-u (D). Each of the oligonucleotides (5fmol, 105cpm) was incubated with an SPSF I-andII-enriched MonoQ fraction (4 zg of protein) in the absence (lanes 1) orinthepresence(lanes 2 to 7) of a 100-fold molar excess of competitor oligonucleotide.Inlanes2to 5,only the SPSF I- and II-complexingoligonucleotides(asmarked on top of the gel) were used as competitors,whereastwounrelatedoligonucleotides were added to thebindingreactionmixture in lanes 6(BE-1,representing the BPV-1 distal enhancer) and7(PyE-1, representing the polyoma-virus Benhancer).TheSPSFI- andII-DNAcomplexes A, Bi, and B2 are indicated byarrows. F indicatesthepositionof free, unbound oligonucleotide.
upperstrand is inhibited by itself (lane 5),ORI-I (lane 2),P-l (lane 3), and P-u (lane 4) but not byBE-1(lane 6) andPyE-1 (lane 7). The complementary oligonucleotide is inactive and does not compete (not shown). The c-myc-derived oligonu-cleotidewastheonlynon-BPVfragmentamong avariety of promoter- and origin-derivedfragments which wesuspected would interactwith SPSF I andII.These includedfragments VOL.66, 1992
00-B B- r.
.44 A
::I=BI
B
on November 9, 2019 by guest
http://jvi.asm.org/
A
A
rP-i
.-i. F -
-_F;
'
wfl
k11B
si
4--0
:~~~~~~~~~~.*F
B
_ =
.r, ;t
--, _-
r--,I _; IIa Ii =
;, _ -
_-- ---A
_ ~~~~~~~-4TW131132
.- F
1 2 3 4 5 6 7 1 23 4 5 6 7
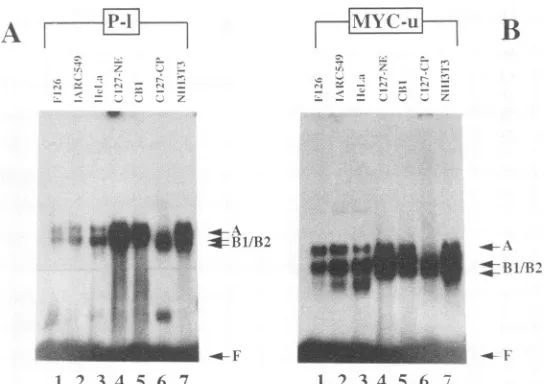
FIG. 3. SPSFI-andII-bindingactivities occur in extracts of cells from different sources. Oligonucleotides P-I(A) and MYC-u (B) were incubated, asdescribed in Materials and Methods, with different nuclear extracts (2 to 4 ,ug): human B cell lines F126 (lane 1) and IARC 549 (lane2); HeLa cells (lane 3);CB1 cells, aBPV-1-transformed C127mouse cell line (lane 5); NIH 3T3 cells (lane 7); and nuclear (NE) and cytoplasmic (CP)extractsofC127cells.Protein-DNAcomplexes A and B1/B2 and free, unbound oligonucleotide (F) are indicated by arrows. Because ofproteolytic activitiesin thecytoplasmic C127 extract, only a weak complex A could be formed in this particular experiment (lane 6 in A andB). The additionallower band inpanelA,lane 6, may have beenformed from apartiallydegradedprotein, which isstillable to binditscognate DNA.
of thesimianvirus40(SV40) origin of replication,afragment
containing theautonomousreplicationsequence(ARS) con-sensus sequence ofS. cerevisiae, the lexA and actin gene
promotersof S. cerevisiae, amousecellular DNAfragment interfering withSV40 DNA replication (8), and manymore
enhancerandpromotersequences.Asanexample,
oligonu-cleotidesPyE-landBE-lfromthepolyomavirus enhancerB and the distal BPV-1 enhancer areincluded inallpanelsof Fig.2.Therelative binding strengths of proteins SPSFIand II to the different binding sites were assessed by titration
competition experiments with increasing oligonucleotide concentrations. No significant differences between the dif-ferent fragments could be observed (data not shown), al-though allfouroligonucleotides haveonlytheweak homol-ogies indicated inFig. 9.
Theretardation activity displayed bythe single-stranded oligonucleotidesispresent notonlyincalfthymusbutalso in
avariety of othersources. As shown in Fig.3 for
oligonu-cleotides P-l(Fig. 3A) and MYC-u(Fig. 3B), binding activ-ities can be identified in extracts from human B-cell lines F126 (EBV negative) and IARC 549 (EBV transformed), fromHeLa, NIH3T3,andBPV-1-transformedmouseC127 cells (CB1 cells), and in nuclear (lane 4) and cytoplasmic (lane 6) extracts ofC127 cells. Thebinding activitiesoccur
ubiquitouslyinextractsof mammaliancells.Nocomparable activity, however,wasobservedin extractsfromS. cerevi-siae (not shown). Acomparison of the BPV-1-transformed
mouse C127 cell line, CB1, and its virus-free parent line demonstrates thatSPSFIand IIinteractions with DNAare
innowayinfluencedbytheexpression of viral proteins. Purificationof SPSF I and II.Since calf thymusappeared
tobethemostabundantsourceofthematerial,weinitiated apurificationofSPSFI and IIinvolvingthree majorsteps, anion exchange chromatography on DEAE-Sepharose and on MonoQaswellasDNAaffinitychromatography. Single-stranded DNA-Sepharose columnswere prepared by
cou-pling the single-stranded oligonucleotide P-l to
CNBr-acti-vatedSepharose. Thepurityandcomposition of the proteins were assessed by SDS gel electrophoresis (Fig. 4). Only three major bands are detectable following DNA affinity chromatography. The upper band (Fig. 4, marked by an asterisk) is an associated impurity which can be seen in fractions from all purification steps. The other two bands, which we designated SPSF I and II (for specific single-stranded DNA-bindingfactor), displaymolecular masses of 42and 39kDa, respectively.
SPSFIandIIbindspecificallytosingle-stranded DNA. Two typesofexperimentswereperformedtoshow that thesetwo
proteins represent single-stranded DNA-binding activities, UVcross-linking and reactivationfrom SDS gels. For UV cross-linking, extracts from various stages of the purifica-tion, all of whichwere active ingel retention assays, were incubated with
32P-labeled
oligonucleotide P-l, irradiated with UVlight of 254 nm, andsubjectedto SDSgel electro-phoresis. While crude and DEAE-Sepharose extractsyielded complex and broad-banding patterns, the MonoQ-and the DNAaffinity-purifiedextractsrevealedtwodistinct bands (Fig. 5A, lanes 3 and 5). The DNA affinity column flowthrough was inactive (lane 4). The two bands ran at
positions correspondingto 73 to 78 kDa,which represents approximately thesumofthemolecularmassesofeither of thetwoproteinsand thesingle-strandedoligonucleotide (97 bases; molecular mass, 31.7kDa). Inseparateexperiments,
wehave demonstrated that themobility ofoligonucleotides inthese SDSgels corresponds tothemobilityofdenatured proteins with similar molecular weights (not
shown).
The high specificity of these interactions was proven by UV cross-linkingof SPSFIandII tooligonucleotidePk3-l in the presence ofhomologous (Fig. SB,lanes 2 to6)
and heterol-ogous(lanes7to12) competitorDNA.Oligonucleotide Pk3-l represents the 5' portion ofoligonucleotideP-l, containing
one SPSF I and II binding site (Fig. 1A; see Fig. 9). As expected, no cross-linking products were detected in the presence ofa100-fold molarexcessoffragmentscontaining
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.175.449.77.269.2]CELLULAR DNA-BINDING PROTEINS INTERACT WITH BPV-1 5993
1 2 3 4 5 6 7 8 NI
97.4
* _66.2
4_5.0
"-~~~~~-.*SPSFI - ~ ---SPSFII
[image:6.612.75.289.76.221.2]--- - - ~~~~~~31.0
FIG. 4. Analysis of purification of SPSF I and II from calf thymus bySDS-polyacrylamide gelelectrophoresis. Lane 1 shows theprotein compositionof the initial whole-cell extract. Lanes 2 and 3 represent flowthrough and a DNA-binding fraction of the DEAE-Sepharose column, respectively.Lane 4is a sample from the pooled active DEAE-Sepharose fractions loaded onto the MonoQ column. Activity peaksoftheMonoQ columns (one of them is shown in lane 5)werepooled and applied to DNA affinity chromatography (lane 6). The flow-through of this column is shown in lane 7. Lane 8 representspurified SPSFI andII,asindicated by arrows. The upper band, marked byanasterisk, isanassociated impurity,which can be seen in fractions from all purification steps. The samples were analyzed on a 10% polyacrylamide-SDS gel and stained with Coomassieblue.Lane M,proteinsizemarkers (in kilodaltons).
SPSF I and II binding sites (Fig. 5B, lanes 2 to 6). The presenceofheterologous competitors (lanes 7 to 12) did not disturb theformation of the expected cross-linked products. In a second set ofexperiments, the two bands from the
A
I~-
,,
-C ;j a ;F <
SDS gelin Fig.4 and materialfrom other regions of the gel were eluted separately from a preparative gel, renatured, incubatedwith 32P-labeled fragment P-l, and subjected to gel retardation analysis. As shown in Fig. 6 (lanes 5 and 6), only the two fractions, corresponding to 40 to 45 and 35 to 39 kDa, displayed binding activity. The higher-molecular-weight material from the SDS gel yielded a shifted band of lowermobility (designated band A), while the lower-molec-ular-weight material yielded a higher-mobility signal desig-nated B1/B2. These twobands are identical in mobility to the signals observed with different DNA affinity-purified frac-tions(Fig.6,lanes 1 to4) as well as to those observed in Fig. 1 and 2. Competition analysis proved the highly specific binding capacities of the renatured proteins (data not shown). We conclude that two activeproteinsof 39 and 42 kDa present incalf thymus extracts are responsible for the observedsingle-strandedDNA-binding activities.
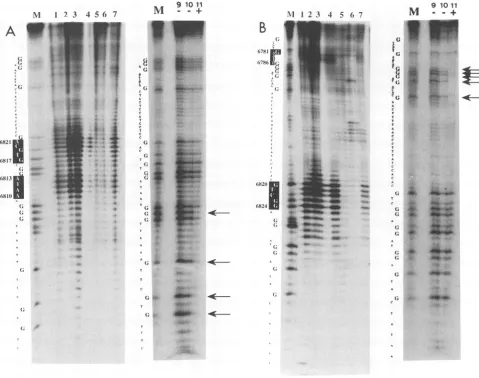
Points ofcontact between SPSFIandIIandsingle-stranded oligonucleotides. The binding specificities of the two proteins were established in footprint experiments. For a T4 DNA polymerase footprint, the 5'-end-labeled oligonucleotide P-u wasincubatedwith a DNA affinity-purified preparation (Fig. 7A) and subsequently exposed to T4 DNA polymerase. Digestion with its endogneous 3'--5' exonuclease activity displaystwostopscentered around positions 6819 and 6812 in the centerof the PMSI region (Fig. 7A, lanes 1, 2, and 3). Incompetition experimentswith the SPSF I-and II-binding oligonucleotides (Fig. 7A, lanes 5 and 6), the stops disap-pear,whiletwounrelatedoligonucleotides (Fig. 7A, lanes4 and7)donotinfluence theprotectionpattern. Inlane 4, the band iscompressed because too little material was applied to thegel.Methylation interference footprints (Fig. 7A, lanes 9 to11)wereperformedwithaDEAE-Sepharoseextractfrom C127 cells. In the free DNAportion, all Gresidues remain
B
0:
0__-]- Pk3-1
Comp..oitor / - Aa- 7A - ,. 97.4
97.4
7*
-o-SPSt
SPSF
I e6(.2
U'SPSFII
SPSFII-=*-415.11
131.F 1 2 3 4
--.. 2=|i.;011|E
:,.>'f;_31_
,e, t:' w.S.z.
^/><J;> SeSs<e
66.2
45.0
31.0
1 2 3 4 .5 6 7 8 9 10 11 12
FIG. 5. Twodistinctpolypeptides are cross-linked tooligonucleotides P-1and Pk3-1 byUVlightirradiation. (A) To perform theUVcross-linking experiment, calfthymus protein extracts from various purificationsteps were incubated with 20 fmol (4 x 105cpm) ofoligonucleotide P-1. DNA-boundproteinswerecross-linked to the labeledoligonucleotidebyirradiation withUVlight (254 nm, 8,000pLW/cm2)for 4min. The reaction productsweresubjectedtoelectrophoresison a10%polyacrylamide-SDSgel, stained withCoomassie blue to visualize the protein markers, dried, andexposedtoX-rayfilms.In lanes 1 and2,proteinsfrom a crude extract (CE) and from a DEAE-Sepharose fraction (DS) wereUVcross-linked
toP-1. Lane 3shows the reactionproductsof an SPSF I- and II-enriched MonoQ fraction(MQ)with P-1.Lane 4 representsflowthrough,and lane 5 shows a sample ofpooled activity peaks from DNA affinity chromatography columns. The cross-linking products of SPSF I and II with oligonucleotideP-1areindicatedby arrows in lane 3 (MonoQ fraction) and in lane 5 (DNA affinity-purified proteins). (B) UVcross-linkingofan
SPSF I- and II-enriched MonoQ fraction to oligonucleotide Pk3-1 in the presence of a 100-fold molar excess of SPSF I- and II-binding oligonucleotidesashomologouscompetitors (lanes2to6,asmarkedontopof thegel)andavarietyofheterologouscompetitors(lanes7to12). Thecross-linking productsbetween Pk3-1 and SPSF I and IIareindicatedby arrows. Lane M, size markers(inkilodaltons).
.-.
"ff&lARd-VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.114.498.430.622.2]I 2 3 45
*
I-*11B/B2
FIG. 6. Renaturation of purified SPSF I and II from SDS-polyacrylamide gels. Size-fractionated, renatured proteins (de-scribed in Materials andMethods)fromanSPSFI- andII-enriched MonoQ fractionwereassayed for theirDNA-bindingactivity. The twoactive fractions, bindingto oligonucleotide P-l, areshown in comparison tothemobilityof SPSF I and II from different DNA affinitychromatography purifications (lanes 1 to4). Lane5 repre-sentsafractionof renaturedproteincorrespondingto40to45kDa, which forms DNA-protein complex A with oligonucleotide P-1 (indicatedbyanarrow). ComplexesB1/B2(indicated byanarrow) in lane6 arethe results of interaction of the 35-to39-kDafraction of renaturedproteins with P-l. FreeDNAis indicatedbyF.
unprotected,while incomplexed DNA,the four G residues indicated by arrows are protected. One of these (position
6807)
resides closetooneoftheregions
protectedin the T4 DNA polymerase footprint, while the other three G's are located upto 15bases upstream(5').Similar
experiments
wereperformed
with the complemen-tarystrand P-1. Two T4 stopswere observed, around posi-tion 6783(nearthestartof thegel)and aroundposition 6823 (Fig. 7B, lanes 1 to 3). Correspondingly, five G residues (indicated by arrows)werefoundtobeprotectedin methyl-ation interferenceexperiments. Theyaresituated inaregion close tothe upper T4 stop, while the G residues around the lower T4 stop are not protected. In a methylation interfer-ence experiment with the shorter fragment Pk3-1, four G residues (indicated by open arrowheads in Fig.9)
wereprotected (not shown). We conclude that the T4 DNA polymerase andthe methylation interference data coincide and overlap and that two binding sites for SPSF I and II within the PMS1 region thusare located between positions 6780 and 6825
(see Fig. 9).
Asalready
indicatedby
the UV cross-linking experimentshowninFig.
5B,onebinding site,
locatedon oligonucleotide Pk3-1,issufficientforinteracting with SPSFIand II.Thus,thetwoseparateprotectedregions
of theT4footprint analysis
eachrepresentaseparatebinding
site.Asecondbindingsite forproteinsSPSF I and IIonBPV-1 DNAlocated around the minimaloriginof BPV-1replication
was analyzed by T4 DNA polymerase footprinting with oligonucleotide ORI-1, derived from the lower strand of this region.Asdiscussedabove,thecomplementary oligonucle-otide ORI-u does not bind. The footprint (Fig. 8, lane 4) shows two T4 DNA polymerase stop regions, one around
position
7925 andone around position11. InFig. 8, lanes 2 and 3, a control experiment with a flowthrough from the DNAaffinity
column, which is devoid of DNA-binding activity, is shown. Lanes 5to 10 ofFig.8 showcompetitionexperiments
with homologous oligonucleotides (lanes 5 to8),
whichremoveboth T4 stops, and with nonhomologousoligonucleotides (lanes9and 10).The dataaresummarized in Fig. 9 and show that thebinding site for SPSFI and II mapsbetween positions 7925 and 11, around anHpaI site locatedwithin theminimaloriginof BPV-1 DNAreplication.
DISCUSSION
Wehave
purified
twosingle-strand-specific DNA-binding
proteinsfromcalfthymus.
Theproteins, designated
SPSFI and SPSF II, have molecular masses of approximately42 and39kDa,respectively,
asshownbySDS-polyacrylamide
gelelectrophoresis
of the DNAaffinity
chromatography-purified proteins(Fig. 4),
byUVcross-linking experiments
(Fig. 5), and bythe behavior of thetwo renaturedproteins
reisolatedfromdenaturing gels (Fig.
6).
Bothproteins
canbe isolated fromavarietyof othersources(Fig. 3)
and appearto beubiquitous
inextractsof mammalian cells.SPSF I and II
specifically
bindtosingle-stranded
oligonu-cleotidescontaining
sequences derived from one strand of the BPV-1 minimalorigin
ofreplication,
from either strand of domain1 of theplasmid
maintenance sequence(PMS)
of BPV-1, and fromonestrand ofthe P2 promoterofthe human c-myc gene. We haveperformed gel
retentionanalysis,
methylation interference,and T4 DNApolymerasefootprint
experiments
to determine theprecise
points
of contact between thetwoproteins
andtheir DNA targets.AsshowninFig. 2,SPSF I-andII-enrichedfractions
give
rise to threespecific DNA-protein complexes
(A
and adoublet,
B1/B2)
ingel
retention assayswitholigonucleotides
containingbinding
sequencesfor SPSF I and II. Because of their similar elutionprofiles (see
Materials andMethods),
the twoproteins
couldnotberesolvedproperly during
columnchromatography.
In ordertoassign
eachprotein
to one of the observedcomplexes,
wehave therefore used renatured SPSF I and IIpreparations
obtained fromSDSgels
(Fig. 4).
AscanbeseeninFig.
6,thisexperiment
allowedus toassign
the uppercomplex
A(seen
inFig.
2)
tothe 42-kDaprotein
SPSF I and the doubletB1/B2
(seen
inFig. 2)
tothe39-kDa SPSF II. The association of SPSF II with the doublet complex B1/B2is observedthroughout
allpurification
steps, including DNAaffinity chromatography.
This observation rulesoutthe presence ofproteins
other than SPSF I and II and proves that bothproteins
bindseparately
to the same single-strandedoligonucleotides.
We also know that bothproteins copurify
under allconditions tested and that com-plexesAandB1/B2behaveindistinguishably
incompetition
experiments.
These observations donotprovebutstrongly
indicate thattheyarestructurally
relatedtoeach other. The doublet retentionsignal
observed with SPSF II may be due to aposttranslational species
of SPSF II.In a first step to
gaining
information about thebinding
specificities
of SPSF I and II, weperformed competition
experimentswith differentoligonucleotides
fromavariety
of sources, such as thepolyomavirus
enhancerB,
the SV40 origin of DNAreplication,
asynthetic
S. cerevisiaeorigin
carrying
a consensusARS sequence, and variouseukaryotic
promoterregions.
None of theseboundtoSPSF I andII,
nor didtheycompetein double-strandedorsingle-stranded
form with SPSF I- and SPSFII-DNA complexes.
Theonly
oligonucleotide
other than the BPV-1-derivedfragments
foundtointerferewith SPSF I- andII-DNAcomplex
forma-tionwasthe upper strand ofthe P2 promoter from the human c-myc gene.As shown inFig. 2,
itwascapable
ofcompeting
for DNAbinding
with any of theother threeoligonucleotides
containing
an SPSF I and IIbinding
site. A sequencecomparison
of the DNAregions containing
theputative
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.126.235.73.218.2]CELLULAR DNA-BINDING PROTEINS INTERACT WITH BPV-1 5995
NI 1 23 4 56 7
A
6821j
6817
6813
681(. _
As
(;
4-A _~~~~.
A X1
G~~~~~~A.s G~~~
g roll
M +
_
_s
._. ..
i = _r,._
I ... W.
f s
A {; js *
C
S t?:
*
C h:o
T .4,
C .
C Rea C ,,^ C .
C C
(; * _'
^..s:7e.:.
C .s _
(s *
_H#--T =; _;S_
(; t
(; _ _
A w
1' X tZ
A .6 . ;e
A,.^
_;;
1. _e _
(; w
{;
r w _>
T b
A t.
A t*'
.
A g
_t-T *
T ?/
c .*.
_
; *.} _
Y
_
{i
*@jeb
Ni 1 2 3 4 5 6 7
B
6781 7Z8
(682(I
6824 I
(
4'
.X. *
t- .
rtl: rII
I O
I _
1 . 4
_..mf
I OMM _tw
afowt
( = _
M910 11
_4 _
_
; ^.
A( .
FIG. 7. T4 DNA polymerase and methylation interference footprinting experiments with SPSF I and II. For T4 DNA polymerase
footprinting (lanes1to7 inpanelsAandB), differentamountsofprotein(approximately 50to100pg, asestimated from gel retention assays)
ofaDNAaffinity-purified SPSFI and IIpreparationwereincubatedwith5'-end-labeled oligonucleotide P-u (A)orP-1 (B) inabinding reaction
as described in Materials and Methods. The products of the subsequent T4 DNA polymerase treatment were analyzed on 10% polyacrylamide-SDS-ureagels. Lanes 1to3(AandB)showtheT4DNApolymerase cleavagepatternofSPSFI-andII-boundDNA inthe
absence ofcompetitorDNA. A 100-foldmolarexcessofhomologous competitorwasaddedtothereaction mixtures inlanes5(A, P-u; B, P-i) and 6 (A and B, ORI-I). The reaction products obtained in thepresenceofheterologous competitorsareshown in lanes 4 (A and B, BE-I)
and 7 (A and B, PyE-l). Nucleotides shown in white letters against black indicate the exact positions of T4 DNA polymerase stops.
MethylationinterferencestudieswerecarriedoutwithanactiveDEAE-Sepharoseextractfrom C127 cells. Inlanes 9 and 10 (A, P-u; B, P-i),
thepiperidinecleavagepatternoffree, unboundDNAisshown, whereas in lane11, DNA from complexes A andB11/B2wasanalyzed. Arrows
indicatethepositions of G residues which,ifmethylated,preventproteinbinding. Gsequencesizemarkers for oligonucleotides P-u (A) and
P-i (B)are shown in lanes M.
SPSF I- and II-binding regions in the four known active
fragmentsidentified certainsequence homologieswhichare
subdivided into short homology boxes (Fig. 9). We even
attempted to define a consensus sequence. Its functional significance,apartfromservingas aDNAtargetforproteins SPSF I andII,remains unknown.
Thepresenceof cellularprotein-binding sites around viral origins of DNA replication is not uncommon. In many
instances, they represent binding sites for transcription factors(3). Some elements involved in transcriptional
regu-lationarelocatedclosetotheSPSF I- andII-bindingsite in the BPV-1 PMSI region: the polyadenylation signalforlate BPV-1 gene expression, the promoters P7185 and PL, and protein E2 recognition element E2RE2. A constitutive
en-hancer sequence which is negatively regulated by the E2
repressor through competitive binding for a cellular factor was identified between nucleotides 7162 and 7275 (40).
Recently, this element was reported to be necessary for transformation andreplication functions(41). The constitu-tive enhancer appears to be a master regulator of BPV-1 transcription and is required for maintenance of BPV-1 replication in latently infected cells, although Ustav and Stenlund (38)have reported that it is not requiredin
tran-sient-replication assays. cis-Acting regulatory elements close to the BPV-1 origin of replication are E2RE1, E2-binding sites 11 and 12, and the promoters P7940 and P89. Whetherand how someoftheseelements, inparticular the constitutive enhancer, which is positioned closest to the VOL. 66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.70.549.73.451.2]A
.1.
'1.
A A
A
A A
A
1311
C
*1
.1.
;
C
FIG. 8. T4 DNA polymerase footprinting with SPSF I- and
II-complexed oligonucleotide ORI-1. The experiment was carried outas described in the legend to Fig. 7. Lane 1 shows T4 DNA
polymerase-cleaved oligonucleotide without protein as a control. Different amounts of a complexformed between the flowthrough
material fromaDNAaffinity chromatographycolumn andfragment ORI-Iweredigestedwith T4 DNApolymerase (lanes2and3).Lane 4shows the reactionproductsof SPSF I- andII-complexed ORI-Iin the absence ofcompetitors. Lanes5(ORI-1),6 (P-u),7(P-l), and 8
(MYC-u) show the results obtainedwitha100-fold molarexcessof
oligonucleotides that were able to bind SPSF I and II upon gel retention.Inlane9, oligonucleotide BE-1wasusedas aheterologous
competitor, while lane 10 shows the result obtained with
oligonu-cleotidePyE-las aheterologous competitor.ThesequenceofORI-I
is written frombottomtotopalonglaneM,which indicates the G
sequencesize markers revealedbyMaxam andGilbertsequencing. Nucleotides showninwhitelettersagainstblack indicate theexact
positionsof T4 DNApolymerasestops.
bindingsite inPMSl,interact with SPSF I andIIwill haveto
beinvestigatedinthe future.
Data about therole ofPMS sequencesseemtobe contra-dictory. Earlier resultsconcerningthe role of PMS and the function of El (18-20) were difficult if not impossible to reproduce (38).Weattemptedtodetermine therequirements
forPMSI domains 1 and 2by performing transient-replica-tionassays bythe method of Ustav and Stenlund(38),with mutantplasmidsdeleted for domain 1and/ordomain 2(44a). Without domain 1, the plasmid was able to replicate as efficientlyaswild-type BPV-1,whereas the domain 2 dele-tion mutant failed toreplicate.In accordance with the results of Vande Pol andHowley (41)and Ustav and Stenlund(38), we assume that deletion of domain 2, which encompasses regions E2RE2, PL,
P7185.
and the constitutive enhancer, destroys important transcriptionalelements. Unless BPV-1 proteins El and E2 are provided in trans, this deletion prevents extrachromosomalreplication
of the transfected BPV-1plasmid.
These data would thus indicate that PMSI domain 1plays
norole inplasmid
maintenance and replica-tion.Interaction ofSPSF I and IIwiththisregion might
have otherfunctions than an involvement in maintenance of the extrachromosomalplasmid
state. Inthis context, it may be relevant thatonlythe full BPV-1 genome is able todisplay
100% transformationandreplication efficiency(15,
17,32).
The elements which contributeto 100% activity of the full genomehavenotyetbeenidentified and may include SPSF I andIIbindingsites.SPSF I and II were isolated as
single-stranded
DNA-specific proteins. However, they can also bind with low affinitytoveryshortdouble-strandedoligonucleotides
(c50
bp)containing
theirspecific binding
sites. Theaffinity
to double-strandedfragments strongly
depends
onfragment
length. Double-strandedfragments longerthan about 100bp
are nolongerrecognized bySPSF Iand II. Other
sequence-specific single-stranded DNA-binding proteins,
with strong indications thatthey
are involved in initiation of DNA synthesis,have also been showntodisplay
weakaffinitiesto double-stranded DNA sequences. The S. cerevisiaeproteins
ACBP(10)
and ssARS-T(33),
which appear to be indistin-guishable, interactspecifically
with the T-rich strand of the ARS core sequence. Both groups reported lowaffinity
for theirrespective proteins
todouble-stranded ARS sequences. Additionally, an SV40 origin-binding protein,purified
from primate cells,alsointeracts withsingle-anddouble-stranded DNA(37).
Recently, a proteinfrom HeLacells,
Pur, was described(1)
to bindspecifically
tosingle-stranded
se-quencesin5'-flanking regions
of the c-myc gene(1.6
kb from thestartoftranscription
ofP1)
and alsotothecorresponding
double-stranded sequences. Thebinding
site of Pur is lo-cated inanucleaseSi-sensitive
region
nearDNase I-hyper-sensitive siteII,
(35). Interestingly,
the SPSF I and IIbinding
sites in the P2 promoter of the c-myc geneaswellas thebindingsites in thePMSIregion
of BPV-1(30)
also reside in the directvicinity
of DNaseI-hypersensitive
sites.We have wondered how a
single-stranded
DNA-specific
proteinmight
findbinding
sites inadouble-strandedplasmid.
Sincethiscanbereadily explained
viasecondary
structuralfeatures,
we subcloned afragment
from nucleotides 6685(AccI)
to 6945(BglII) containing
the SPSF I- and II-PMSI binding sites into a pGEM4Zvector and treated this con-struct and the controlplasmid
with endonuclease VII fromphage
T4(11).
This enzymespecifically recognizes
second-ary structures insingle-
and double-stranded DNA. Thepreliminary
results of thisexperiment (unpublished
data)
indicated that indeed these DNA sequences form stablesecondary
structures, which may includepartially
single-stranded DNAregions,
and thus could represent efficientbinding
sites for SPSF I and II.Afinal
point
regarding
theaction ofSPSFIand II relates to the observed presence of twobinding
sites within a distanceof 800bp.
Iftheserepresented
twopartsofanorigin
;.dowf
-z'
N-1 1 2 3 4 5 6 7 8 9 11)
G
7924 7927
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.103.251.73.468.2]CELLULAR DNA-BINDING PROTEINS INTERACT WITH BPV-1 5997
P- 5'-TGTGTCACATCTGCAAGGACTTATGCCCT'
6760
G
_T
_T
_G
CCTCCTACCTCATC TATTA CA-3'6849
5ITAccATA
ACACg
T^TTACCT"TTGCAGTGTGACAC 36859 6849 6760
4'-TTTTTTC5 T AAlTTTATTCACT-rirCeAgnTC7TTT89TTCCTTTC1QTCC,
2414
consensus sequence
2451i
5t
--3A
YSS KA
FIG. 9. Contact pointsofSPSF I andII.White letters in black boxes in the four oligonucleotides binding to SPSF I and II indicate T4 DNA polymerase stops. Arrows above thesequences indicate the positions of protected G residues, as determined by methylation interference footprinting. Numbers below the sequences refer to positions in the original DNAs (BPV-1 DNA and myc). The underlined sequence in oligonucleotideMYC-u representsthe original DNA sequence from nucleotides 2414 to 2451 of the humanc-mycgene; the flanking sequences arerandom.Sequencehomologies, identifiedby comparison of thefour SPSF I- andII-bindingoligonucleotidesequences, are subdivided into fourshort homology boxes. A proposed consensus sequence is written below.
ofreplication,this wouldhardly be uncommon. The origin of replication of EBV, another DNA virus replicating as an extrachromosomal plasmid in the latent state (16, 29), and the prokaryotic origin of plasmid R6K (25) consist of two partswhichare separated by more than 1 kb.
Inaddition to a putative role in DNA replication, SPSF I and IImight also be involved in the regulation of transcrip-tion. Several single-stranded DNA-binding proteins have beenreported to be involved in transcriptional regulation. A nuclear protein fromDrosophila melanogaster specifically interacts with the mRNA-like strand of the long terminal repeat of retrotransposon 1731,which plays a role in regu-lation of 1731 expression (6). CNBP, which sequence-spe-cificallybindssingle-stranded DNA,recognizes the noncod-ing strand of the sterol regulatory element found in the HMG-coenzyme A reductase promoter (28). Factor SRE-BF, isolated from Chinese hamster ovary nuclear extracts, interacts with double- and single-stranded DNA sequences from acis-acting element,designatedSRE-1,whichhas been identified in several promoters of genes necessary forsterol regulation (36). A protein from skeletal muscle extracts, designated muscle factor 3, interacts with single-stranded
DNA regions from three different muscle gene DNA ele-ments: the MCAT gene sequence motif, the skeletal actin muscle regulatory element, and acreatine kinase enhancer probe (31). The single-stranded DNA-binding stimulatory factor DBSF,purified fromS. cerevisiae, enables the puri-fied human estrogen receptor to bind as a dimer to its cognate palindromic DNA sequence and thus to activate genetranscription (24).Protein H16 from nuclearextractsof noninfected monkey cells binds a unique single-stranded
DNAsequence in theearly promoter of SV40 and has been shown to stimulate transcription invitro bypurified RNA polymerase II (7). Aprotein purified from rat liver nuclei, called PYBP, bindsonlyto single-strandedpyrimidine-rich DNA, present, for example, in the human transferrin gene promoter (2). A direct involvement of SPSF I and II in transcriptional regulation by bindingdirectlyto RNAcould beruledout.Invitro-transcribedRNAfrom the SPSFI-and II-PMSlbinding sites did notinteract with purified SPSF I andII (datanotshown).
Insummary,ourdata indicate thatSPSFIandII,purified from calf thymus, belong to a class of single-stranded DNA-binding proteins,whichbind with sequencespecificity
tosequences mainly located close to or directlywithin an
origin of DNA replication. The precise functions of these proteins have yet to be elucidated. Several models exist which propose functions as helix-destabilizing proteins at theinitiationsite of DNAsynthesis(1)or asspecificinitiator proteins (10,33). Furtherinvestigations willbe necessaryto
obtain insightinto these interactions. ACKNOWLEDGMENTS
We thankB6rriesW.Kemper,University of Cologne, for provid-ingT4endonucleaseVII and HorstIbelgauftsforcritically reading themanuscript.
Thiswork wassupported byDeutsche Forschungsgemeinschaft grant Fa138/3-4.
REFERENCES
1. Bergemann, A. D., and E. M. Johnson. 1992. The HeLa Pur factor binds single-stranded DNA at a specific element
con-servedin geneflanking regions and origins ofDNAreplication. Mol.Cell. Biol. 12:1257-1265.
2. Brunel,F., P. M.Alzari, P. Ferrara, and M. M. Zakin. 1991. Cloning and sequencing of PYBP, a pyrimidine-rich specific single strand DNA-binding protein. Nucleic Acids Res. 19: 5237-5245.
3. DePamphilis, M. L. 1988.Transcriptionalelements as compo-nents of eukaryotic origins of DNA replication. Cell 52:635-638.
4. Dignam, J. D., R. M. Lebowitz, and R. G. Roeder. 1983. Accurate transcription initiation by RNA polymerase II in a solubleextractfromisolatedmammaliannuclei. Nucleic Acids Res. 11:1475-1489.
5. Dvoretsky,I., R.Shober,S. K.Chattopadhyay,and D. R.Lowy. 1980. Aquantitativein vitro focus assay for bovinepapilloma virus. Virology103:369-375.
6. Fourcade-Peronnet, F., S. Codani-Simonart, and M. Best-Bel-pomme. 1992. A nuclear single-stranded-DNA binding factor interacts with thelong terminalrepeatsofthe 1731Drosophila retrotransposon. J. Virol. 66:1682-1687.
7. Gaillard, C., and F. Strauss. 1990. Sequence-specific single-strand-binding protein for the simian virus 40 early pro-moter stimulates transcription invitro. J. Mol. Biol. 215:245-255.
8. Harti, M., T. Wilinow, and E. Fanning. 1990. cis-Active
ele-mentsfrom mousechromosomal DNA suppress simian virus 40 DNAreplication. J.Virol. 64:2884-2894.
9. Hirt,B.1967.Selective extraction ofpolyomavirus DNA from infectedmousecell cultures. J. Mol. Biol.26:365-369. 10. Hofinann, J. F.-X., and S. M. Gasser. 1991.Identification and
purification ofa protein that binds the yeast ARS consensus
. ...
P-l l Ol:l
IORI-I
IMYC-u S'--TGCAGTGCATAAATTGTTGCCAGAGGCIm TCAATGC-3'
VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
[image:10.612.75.556.77.199.2]sequence. Cell 64:951-960.
11. Kosak, H. G., and B. W. Kemper. 1990. Large scale preparation of T4endonucleaseVIIfromover-expressing bacteria. Eur. J. Biochem. 194:779-784.
12. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
13. Law, M. F., D. R.Lowy,I. Dvoretzky,and P. M. Howley. 1981. Mouse cellstransformedby bovinepapillomaviruscontain only extrachromosomal viralDNA sequences. Proc. Natl. Acad. Sci. USA 78:2727-2731.
14. Li, R., J. D. Knight, S. P. Jackson, R. Tjian, and M. R. Botchan. 1991.Direct interactionbetween Spl andthe BPV enhancer E2 protein mediates synergistic activation of transcription. Cell 65:493-505.
15. Lowy, D. R.,I. Dvoretzky, R. Shober, M. F. Law, L. Engel, and P. M. Howley. 1980. In vitro tumorigenic transformation by a defined sub-genomicfragment of bovinepapillomavirus DNA. Nature(London)287:72-74.
16. Lupton,S.,and M. Levine.1985. Mapping genetic elements of Epstein-Barr virus that facilitate extrachromosomalpersistence in Epstein-Barr virus-derived plasmids in human cells. Mol. Cell. Biol. 5:2533-2542.
17. Lusky, M., L. Berg, H. Weiher, and M. R. Botchan. 1983. Bovinepapillomavirus containsanactivator ofgeneexpression atthe distalend oftheearly transcription unit. Mol. Cell. Biol. 3:1108-1122.
18. Lusky, M., and M. R. Botchan. 1984. Characterization ofthe bovine papilloma virus plasmid maintenance sequences. Cell 36:391-401.
19. Lusky, M., and M. R. Botchan. 1985.Geneticanalysis of bovine papillomavirustype 1trans-acting replication factors. J. Virol. 53:955-965.
20. Lusky, M., and M. R. Botchan. 1986. Transientreplication of plasmids: cis and trans requirements. Proc. Natl. Acad. Sci. USA83:3609-3613.
21. Lusky, M., and E. Fontane. 1991. Formationof the complex of bovinepapillomavirus El and E2proteins is modulatedby E2 phosphorylation and depends upon sequenceswithin the car-boxyl terminus ofEl. Proc. Natl. Acad. Sci. USA 88:6363-6367.
22. Maxam, A. M., and W. Gilbert. 1977. A new method of sequencingDNA. Proc. Natl.Acad.Sci. USA74:560-564. 23. Mohr, I. J., R.Clark,S.Sun,E.Androphy, P. MacPherson, and
M.R. Botchan.1990.Targeting the El replicationproteintothe papillomavirus originofreplication bycomplexformation with the E2transactivator. Science250:1694-1699.
24. Mukherjee,R., and P. Chambon. 1990. Asingle-stranded DNA-binding protein promotes the binding of the purifiedestrogen receptortoitsresponsive element. Nucleic AcidsRes. 18:5713-5716.
25. Mukherjee, S., H. Erickson, and D. Bastia. 1988. Enhancer-origininteraction inplasmid R6Kinvolves aDNAloop medi-atedby initiator protein. Cell 52:375-383.
26. Ofverstedt,L.G., K.Hammarstrom,N.Balgobin, S. Hjerten, U. Petterson, and J.Chattopadhyay. 1984. Rapidandquantitative recoveryofDNAfragments from gelsbydisplacement electro-phoresis(isotachophoresis). Biochim. Biophys. Acta 782:120-126.
27. Piette, J., and M. Yaniv. 1986. Molecular analysis of the interaction between an enhancer binding factorand its DNA target.Nucleic Acids Res. 14:9595-9611.
28. Rajavashisth, T. B., A. K. Taylor, A. Andalibi, K. L. Svenson,
andA.J. Lusis. 1989. Identification of azinc-fingerprotein that bindstothe sterolregulatory element. Science 254:640-643.
29. Reisman, D., and B. Sudgen. 1986. trans activation of an Epstein-Barr viraltranscriptional enhancer by the Epstein-Barr viralnuclearantigen1.Mol. Cell. Biol. 6:3838-3846.
30. Rosl, F., W. Waldeck, and G. Sauer. 1983.Isolationofepisomal bovinepapillomavirus chromatinandidentification ofaDNase I-hypersensitive region. J.Virol.46:567-574.
31. Santoro, I., T. Yi, and K. Walsh. 1991. Identification of single-strandedDNA-binding proteins that interact with musclegene elements. Mol. Cell. Biol. 11:1944-1953.
32. Sarver, N., J.C.Byrne,and P. M.Howley. 1982. Transforma-tionandreplication inmousecellsof abovine papillomavirus-pML2 plasmidvector that can be rescued in bacteria. Proc. Natl.Acad. Sci. USA 79:7147-7151.
33. Schmidt, A. M.A., S. U. Herterich, and G. Krauss. 1991. A single-strandedDNAbinding proteinfromS. cerevisiae specif-ically recognizes the T-rich strandofthe core sequenceof ARS elementsanddiscriminatesagainstmutantsequences.EMBO J. 10:981-985.
34. Schneider, R., I. Gander, U. Muller, R. Mertz, and E.-L. Winnacker.1986.Asensitive andrapid gel retentionassayfor nuclear factor I and other DNA-binding proteins in crude nuclearextracts. Nucleic AcidsRes.14:1303-1317.
35. Siebenlist, U.,L. Hennighausen,J.Battey,andP.Leder.1984. Chromatinstructureandprotein binding in the putative regula-tory region of the c-myc gene in Burkitt's lymphoma. Cell 37:381-391.
36. Stark, H.,0.Weinberger, and J.Weinberger. 1992. Common double- and single-stranded DNA binding factor for a sterol regulatory element. Proc. Natl. Acad. Sci. USA 89:2180-2184.
37. Traut,W., and E. Fanning. 1988.Sequence-specificinteractions betweenacellularDNA-binding proteinandthe simian virus40 origin ofreplication.Mol. Cell. Biol.8:903-911.
38. Ustav, M., and A. Stenlund. 1991. Transient replication of BPV-1requires twoviralpolypeptidesencodedbytheEland E2 openreading frames. EMBO J. 10:449-457.
39. Ustav, M., E. Ustav, P. Szymanski, and A. Stenlund. 1991. Identification of theorigin of replication of bovine papillomavi-rusand characterization of the viraloriginrecognition factorEl. EMBOJ. 10:4321-4329.
40. Vande Pol, S. B., and P. M. Howley. 1990.Abovine papilloma-virus constitutive enhancer is negatively regulated by the E2 repressor through competitive binding fora cellular factor. J. Virol.64:5420-5429.
41. Vande Pol, S. B., and P. M. Howley. 1992. The bovine papillo-mavirus constitutive enhancer is essential forviral transforma-tion,DNAreplication, and the maintenance oflatency.J.Virol. 66:2346-2358.
42. VanWilson,G., andJ. Ludes-Meyers. 1991. Abovine papillo-mavirusEl-related proteinbindsspecificallytobovine papillo-mavirus DNA. J. Virol.65:5314-5322.
43. Waldeck, W.,F.Rosl,and H.Zentgraf. 1984.Originof replica-tion in episomal bovine papilloma virustype 1 DNAisolated from transformed cells.EMBO J. 3:2173-2178.
44. Weiher, H., and M. R. Botchan. 1984. Anenhancer sequence from bovine papilloma virus DNA consists of two essential regions. Nucleic AcidsRes. 12:2901-2916.
44a.Weissinger, F.Unpublisheddata.
45. Yang, L., and M. R. Botchan. 1990. Replication of bovine papillomavirus type 1 DNA initiates within an E2-responsive enhancer element. J. Virol.63:5903-5911.
46. Yang,L.,R.Li,I.J.Mohr,R.Clark,andM. R. Botchan.1991. Activation of BPV-1 replication in vitro by the transcription factor E2. Nature(London)353:628-632.