0022-538X/82/060790-08$02.00/0
Cloning and
Characterization
of DNA Complementary
to
the
Measles
Virus mRNA
Encoding Hemagglutinin and Matrix
Protein
SHMUELROZENBLATT,* CILA GESANG, VERED LAVIE, ANDFELICIA S. NEUMANN
Department of Virology, WeizmannInstitute of Science, Rehovot,Israel
Received 27 October1981/Accepted 1 February 1982
Since cloning and characterization of DNA complementary to measles virus
mRNA encoding for the nucleocapsid protein (M. Gorecki and S. Rozenblatt,
Proc. Natl. Acad. Sci. U.S.A. 77:3686-3690, 1980), two additional
measles-specific clones containing different classes ofsequenceshave beencharacterized.
Thecloned plasmids contain inserts of 480 and 530 base pairsasshown byagarose
gel electrophoresis and electron microscopy. The sizes of the mRNA species
complementary tothese insertsare 1,700 and 1,550 nucleotides, respectively,as
determinedby the Northern technique. The cloned DNA fragmentswerefurther
identified as reverse transcripts of the mRNA coding for the glycoprotein and
matrixprotein of measles virus. The major cell-free translation products of mRNA
selected byhybridizationtotheindividual cloned DNAs comigrated with the 70K
in vitro products and matrix proteins. One of the cell-free translation products
(70K) was also immunoprecipitated specifically with monoclonal antibodies
against measles virus glycoprotein.
The
measles
virus is a complex virus thatinteracts
withits
hostcell in
either an acute orapersistent infection.
Themeaslesviroid
consists
of six structural
proteins (9,
19-21) andasingle-stranded
RNA(molecular weight,
6 x106)
(9,
11, 17).
Six virus-specific
mRNAsdiffering
insize have been
recognized
inmonkey
cellsin-fected with measles virus
in the presenceof
actinomycin
D (7). Thepolypeptides
synthe-sized
in anmRNA-dependent reticulocyte
cell-free
system weresubsequently identified
asau-thentic
measlespolypeptides by electrophoretic
and
immunological
techniques
as well asby
analysis
oftheir
tryptic
peptide
patterns(15).
Thus, it
is nowpossible
todirectly
examinemeasles
virus-specific
geneexpression during
the courseof infection andtodetermine whether
acute versus persistent infections canbe
corre-lated with differential patterns of gene expres-sion.
A powerful approach to
studying
differentialgene
expression
ineucaryotic
cells and in cellsinfected
with virus involves theproduction
ofnucleic acid
hybridization probes
thatarehighly
radioactive and gene
specific.
In a previousreport (6), the method used to
obtain
measlesvirus nucleic acid hybridization products that
are
highly
radioactive was described. Arecom-binant
plasmid
that contains most of the nucleicacid sequences encoding measles virus
nucleo-capsid protein
was obtained. In the presentstudy,
we describe the isolation andcharacter-ization
of
tworecombinant plasmids
thatcontain
partial nucleic acid
sequences encoding parts ofthe measles
virus
glycoprotein (hemagglutinin)
and matrix
protein.
MATERIALSANDMETHODS
Cells and viruses. The CV-1 line of African green
monkey kidney cellswasobtained from Flow Labora-tories, Inc. (Rockville,Md.). The cellsweregrownin Eagle medium supplemented with10%calfserum.The Edmonston strain of measlesvirus, obtained from B.
Fields (Harvard Medical School),wasplaquepurified
three times in CV-1 cells. A stock was prepared by
infecting CV-1 cellsatamultiplicity of1/1,000. After thedevelopment ofa markedcytopathic effect,virus
was harvested from freeze-thaw lysates of the cells. Cell debriswasremovedbycentrifugationat1,000xg
for 10 min at 4°C, and the supernatants which
con-tained2 x 106to6x 106 PFU/ml (as titrated inVero
Africangreenmonkey cells)werestored at -90°C.
Selection of measlesvirus-specificRNA. Selectionof
RNA wascarriedoutbythemethod ofRicciardietal.
(13). ClonedDNA(10 ,ug)linearized with EcoRIwas
denatured at 100°C for 5 min and applied to
1-cm-square nitrocellulose filter pads presoaked in 0.9 M
NaCI-0.09M sodium citrate. The nitrocellulose filter
was air dried and then baked at 80°C for 2 h under
reduced pressure. Hybridization solutions (100 ,ul)
consisted of 10
p.g
of polyadenylic acid [poly(A)]-containing RNA [poly(A)-RNA], 10 mM
Tris-hydro-chloride(pH7.4),2 mMEDTA, 0.4% sodiumdodecyl
sulfate (SDS), 10 ,ug of Escherichia coli tRNA, and
50% (vol/vol) formamide. Hybridization was carried
out at37°Cfor 16 h.poly(A)-mRNAswereeluted and
790
on November 10, 2019 by guest
http://jvi.asm.org/
plaque-purified measles virus at 0.1 to 0.5 PFU/cell. RNA was extracted with phenol and chloroform-isoamyl alcohol and then precipitated with LiCl (16). poly(A)-mRNAs were thenpurified by oligodeoxythy-midylic acid-cellulose chromatography (2).
Fractionation, extraction,and in vitro translationof
RNA obtained under denaturing conditions. poly(A)-RNA of measles virus-infected cells (20 ,ug) was denatured in the presence of 15 mMmethyl-mercuric
hydroxide and fractionated in 1.5% agarose and 7.5 mMmethyl-mercuric hydroxide as described by Bai-ley and Davidson (3). A gelfraction containing RNA of 1,000 to 2,700nucleotides wassliced into 1-mm-thick
slices, and RNA was eluted from individual gel slices
by freeze-thawing. Agarose slices were placed in Ep-pendorf tubes containing 5 ,ug of rabbit liver tRNA, 0.1 M NaCl, 10 mM Tris-hydrochloride (pH 7.4), 1 mM EDTA,0.2%SDS, and 20 mM ,-mercaptoethanol (400
jil). The agarose slices were melted(95°C for 2 min),
blended in a Vortexmixer, andfrozen immediately in liquid nitrogen. After thawing, the agarose was re-moved by centrifugation at 12,000 x g for 20 min
(4°C).The RNA waspurified from thesupernatant by
successive extractions with phenol and chloroform-isoamyl alcohol followed by ethanolprecipitationand
subsequentlytranslated invitro inreticulocyte lysates
prepared by the method of Pelham and Jackson (12).
RESULTS
Identification of
cDNA clones.One
clone,
Cl-N,
carrying measles virus
complementary
DNA(cDNA)
sequencesfor
nucleocapsid proteins
has
previously been described (6).
Characteriza-tion of the clone revealed
aninsert
of
approxi-mately 1,420 base
pairs (Fig. 1A). The size of the
mRNA
complementary
tothe cDNA
was1,750
bases
[including
the
poly(A)
tail];
electron
mi-croscopy
studies
using
anR-loop technique
(Fig.
1B)
have shown that
atleast
80%
of the entire
mRNA
was presentin the clone. To
isolate
cDNA
clones with
sequencesthat do
notcode
for
nucleocapsid proteins,
the measles
virus
cDNA
insert in
Cl-N
wasremoved
from the
pBR322 vehicle by cleavage with PstI and
isolat-ed after fractionation
on a1.5%
agaroseslab
gel.
The
purified
DNAinsert
was
32p
labeled
in vitro
by
nick translation and
wasused
as ahybridiza-tion probe. Of the 39 cDNA clones
(5),
30
(containing
DNAinsertsvarying
insize from
400to 650 base
pairs) showed
stronghybridization
signals
tothis
probe, whereas
9demonstrated
weak hybridization
(to the same extent asbacte-rial
pBR322
DNAused
asinternal
negative
control).
Upon in vitro translation of
the mRNAselected
by hybridization
toindividual
DNAclones
(data
notshown),
thenine
cloneswhich
did
nothybridize with Cl-N could
be dividedinto
twogroups:eight of
themarereferred
to asthe G
group, and one isreferred
to asthe "M"of individual clones with
PstI(Fig.
2)generated
two
fragments in
each clone, onecorresponding
in
size
to linear pBR322 DNA, and a secondfragment consisting of
1,420, 540, and 480base
pairs (Cl-N, Cl-M, and Cl-G, respectively; Fig.
2A). The DNA was blotted onto nitrocellulose
papers,
fixed by baking in
vacuum,and
hybrid-ized with
purified inserts of clone
N32P-labeled
in vitro by nick translation. Hybridization
wasobserved only
tothe
Cl-N insert;
nohybridiza-tion
wasobserved with Cl-G
orCl-M,
evenafter
a
long
exposure(Fig.
2B).The size of the
cDNAinserts
wasfurther
confirmed by electronmicro-scopic studies. Figure
3 shows electronmicro-graphs of heteroduplex
moleculesformed by
reassociation of
pBR322 and the cDNA clones,each
onelinearized by digestion with
EcoRI,which
does not cut the insert. The lengths of theinserted
measlesvirus
sequences appearin themicrographs
tobe 1,420±
20, 530 + 40, and 490 + 40base
pairs in Cl-N, Cl-M, and Cl-G,
respec-tively, which
is inexcellent agreement with thelength determined
by agarose gelelectrophore-sis.
Heteroduplex molecules formed by
reasso-ciation of Cl-N, Cl-M,
and Cl-Gin
all threevariations
haveconfirmed
that thereis no DNAhomology
amongCl-N, Cl-M,
orCl-G (Fig.
3d,e,
and
f).
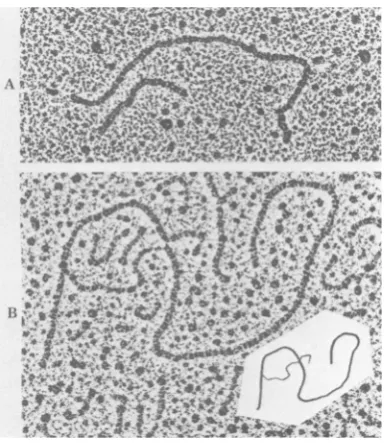
FIG. 1. Electron microscopy of (A Cl-N DNA
%J W
-
ILdigested with PstI and (B) R-looping of Cl-N (1 ,ug)
linearized with EcoRI, denatured, and annealed with mRNA(5 ,ug of measles virus-infected cells (5).Insert
shows interpretive drawing.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.487.257.450.403.625.2]__
FIG. 2. Sizingof measles virus DNA clones. Plasmid DNAs digested withPstIwereelectrophoresed on a 1.5% agarose slab gel stained with ethidium bromide (A), transferred onto nitrocellulose paper (18), and hybridized with
32P-purified,
nick-translated (14) measles virus DNA insert of clone N (B). Lanes: 1, pBR322; 2,Cl-N;3,Cl-M; 4, Cl-G. Right-hand lane:32P-labeledSimianvirus 40 DNA digested withHinfl,usedas markers.
b.p., Basepair.
Correlation of
the cloned measles DNAs withcomigrated
with
60,000-dalton nucleocapsids
[image:3.487.51.438.371.626.2]viral gene products and mRNA species. DNA
of
prepared and
purified from cells infected with
the
individual clones
(Cl-N,
Cl-M,
and
Cl-G)
measles
virus.
Furthermore, the translation
was
separately
digested with EcoRI,
immobi-
product was
immunoprecipitated
with
anti-mea-lized onto
nitrocellulose
filters,
and
hybridized
sles
virus antibodies
(Fig. 4B, lanes 4 and 5). As
with poly(A)-RNA from measles
virus-infected
expected, no
translatable RNA was eluted from
cells. RNA was then eluted
from the
filters and
filters containing either
pBR322 DNA
(Fig.
5A,
translated in an
mRNA-dependent
rabbit reticu-
lane
4)
orsingle-stranded phage
M-13 DNA
locyte cell-free translation system. As shown in
containing
cDNA
inserts of the same
polarity
asFig. 4A, lanes 4 and 5, the
major cell-free
measles virus
mRNA(Fig.
4A
and
B, lanes
6).
translation
product (RNA
hybridized
toCl-N)
Confirmatory
evidence
wasprovided
by
immu-~ ~ .
..
!~~~~~~~~~~~~~~~~~~ ~,
41 :- .
FIG. 3. Electronmicroscopy of heteroduplexDNAs.pBR322 and three clonedDNAs(Cl-M,Cl-G,
Cl-N;
1 ,ugofeach)werelinearizedwithEcoRI,denatured,andannealed(5)asfollows: pBR322and(a)CI-G,(b)Cl-M,(c)Cl-N;(d)Cl-NandC1-M;(e)Cl-Nand
Cl-G;
(f)Cl-MandC1-G.Insetsshowinterpretive drawings. Bar,0.5,um.
on November 10, 2019 by guest
http://jvi.asm.org/
< per, and each of the identical blots was
hybridized
to one of the threeprobes.
Thenucleocapsid
andglycoprotein
cDNAprobes
hybridized to mRNA with an approximate size
of1,750 bases (Fig. 7A and B); and the matrix
clonehybridized to mRNA of 1,550 bases
(Fig.
7C). The difference in hybridization of the
ma-trix clone and clone G is further demonstrated
inFig.
7C' where the blothybridized
with Cl-M2 3 4 5 A
*i
FIG. 4. Products ofthe in vitrotranslation ofRNA
selectedby hybridizationtoM-13phageDNA witha
plus polarity (N) insert (complementary to mRNA)
and minus polarity insert. [35S]methionine-labeled
polypeptides synthesized in a reticulocyte cell-free
system (11) werefractionated on an SDS-10o poly-acrylamide gel. The dried gel was fluorographed at
-90°C (4). (A) Translation products. (B) Immunopre-cipitates. Translationproducts under direction of: no
added RNA(lanes 1), total cytoplasmicpoly(A)-RNA
of uninfectedcells(lanes 2),ormeaslesvirus-infected
cells(lanes 3);RNAselectedby hybridizationof total
cytoplasmicpoly(A)-RNAtoimmobilized Cl-N DNA
(lanes 4); phageDNAwithplus polarityN insert(lanes
5); or minus polarity insert (lanes 6). Molecular
weights (M.W.) shownwerethefollowingmethylated (methyl-14C) proteins of known molecular weight,
fromtop tobottom:myosin phosphorylase B, bovine
serumalbumin, ovalbumin, and carbonic anhydrase.
noprecipitation techniques with monoclonal
antibodies (obtained from B. Bloom and V. ter
Meulen) raised against nucleocapsids and glyco-proteins (Fig. 6). We concluded from these
results(Fig. 4, 5, and 6) that the cDNAsinserted
in the recombinant clones are specific for the
measlesvirus nucleocapsid, the
non-glycosylat-ed precursor of the glycoprotein (hemaggluti-nin), and the matrix protein.
Cl-N, Cl-M, and Cl-G DNAswere32p labeled
in vitro by nick translation and were used as
hybridization probestothree identical
diazoben-B
.- .>
...k
FIG. 5. Productsof theinvitrotranslationoiKINA
selectedbyhybridizationtopBR Cl-M, Cl-G,and
Cl-N DCl-NAclones. [35S]methionine-labeled polypeptides
synthesizedinareticulocyte cell-freesystem(11)were fractionatedon anSDS-10% polyacrylamide gel. The
driedgelwasfluorographedat-90°C (4). (A)
Transla-tionproducts. (B) Translation productsafter
immuno-precipitation with anti-measles guinea pig serum as
describedpreviously (10).Translationproductsunder
direction of: noadded RNA(lanes 1), total
cytoplas-mic poly(A)-RNAofuninfectedcells(lanes 2), RNA
from measles virus-infected cells isolated 24 h after
infection(lanes 3), RNA selectedby hybridizationof
total cytoplasmic poly(A)-RNA to immobilized
pBR322 DNA (lanes 4), Cl-N DNA (lanes 5), Cl-M DNA (lanes 6), and Cl-G DNA(lanes 7). Molecular
weight (M.W.)markersasinFig.4.
B
40.
S"
"1"-NW _;_...._
low.,Oa
4"mwxmw
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.487.52.240.76.364.2] [image:4.487.256.447.226.508.2]794 ROZENBLATT ET AL.
FIG. 6. Products of in vivoand in vitro translation of RNA selected hybridization topBRCl-N andCl-G immunoprecipitated with monoclonal measles virus-specific antibodies. [35S]methionine-labeled proteins of measlesvirus-infected cells and polypeptides synthesized in a reticulocyte cell-free system (11) were fractionated
on an SDS-10% polyacrylamide gel. The dried gel was fluorographed at -90°C (4). Gels show translation products of poly(A)-RNA of measles virus-infected cellsimmunoprecipitated with guinea pig anti-measles virus
sera(A, F). Immunoprecipitationwith rabbit monoclonal antibodiesagainst nucleocapsid protein: (B) in vivo-labeledproteins of measles virus-infected cells; (C) in vitro products ofRNAselected by hybridization of total cytoplasmic poly(A)-RNAtoimmobilized Cl-N DNA immunoprecipitatedasin B;(D) in vivo-labeled proteinsof measles virusimmunoprecipitated with rabbit monoclonal antibodies against glycoprotein; (E) in vitro products ofRNAselected byhybridization of total cytoplasmic poly(A)-RNAtoimmobilizedCl-G DNA immunoprecipi-tated as in D; (G) products as in C immunoprecipitated with glycoprotein antibodies; (H) products as in E
immunoprecipitated with nucleocapsid antibodies.
was
hybridized later
withCl-G.
Simultaneous
hybridization
with Cl-N and Cl-G could notresolve the
two mRNA species (data not shown)._a
s-
_
impCorrelation
of measles virus mRNAspecies
andtheir
corresponding
geneproducts.
poly(A)-RNA
(20
,ug)
wasfractionated
under
denaturing
condi-tions from
measles
virus-infected
cells
(methyl-.1Mmm ,, .*f-_.m-:i-.
won_
FIG. 7. Sizing of measlesvirus mRNAbyhybridizationofCl-N, Cl-M and Cl-Gtototalpoly(A)-RNA from
cells acutely infected with measles virus. Poly(A)-RNA from measles virus-infected cells (lanes 1) and from
uninfected cells(lanes 2)wasfractionatedunderdenaturing conditions (1)in1.5% agarose gel. The RNA was
transferredtodiazobenzyloxymethyl paper and hybridized with nick-translated (14) plasmid [32P]DNA; Cl-N (A), Cl-G (B), and Cl-M (C)werehybridizedlater withCl-G(C').32P-labeled18S and 28SrRNA wereusedas markers.
AN-,
owxmw
.P%.., 1:
...-
...::.
21 1.
O"
ot-.I
....
..:,.
,t,
C '", .:..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.487.131.359.72.240.2] [image:5.487.109.391.441.603.2]MEASLES VIRUS cDNA CLONES 795
mercuric hydroxide
agarose gel). Gel segments(3
cmlong) containing fractionated mRNA in asize range of 1,000 to 2,700
nucleotides
weresliced, and mRNA waseluted
from each gel slice
as
described
above. RNA recovery in thiselu-tion
procedure was 60%, as determined by32p-labeled
rRNA markers. Eluted mRNA wastranslated in
anmRNA-dependent
rabbitreticu-locyte cell-free
translation system. Measuring[35S]methionine incorporation
induced by each mRNAfraction,
twomajor peaks were obtainedcorresponding
in size to 16S and 14S (data notshown). The
[35S]methionine-labeled
polypep-tides immunoprecipitated
with guinea piganti-measles virus
sera werecharacterized
byelec-trophoresis
on a10% SDS-polyacrylamide gel
(Fig.
8).Under
thefractionation conditions
used,
migration of
rRNA markers was found tobe
linear with
respect to thelogarithm
oftheir
molecular weight. The peak fraction for each
translation
product could easily be determined.Fractions for polypeptides
were asfollows:
frac-tion 9 (2,115
nucleotides) for
a 65Kprotein,
fraction
14(1,735nucleotides)
for a 60Kprotein,
fraction
15 (1,677nucleotides) for
a 70Kand
a 40Kprotein, and
fraction 18
(1,515nucleotides)
for
a35K
protein (Fig.
9and
Table1).
DISCUSSION
A
clone
containing measles virus
DNAse-quences
which
specify
mostof the nucleotide
sequences
coding for measles virus
nucleocap-N~ ~~~ .._~ ~ ~ ~~~ ~~~~~~~... Ci~~~~~~~~~~~~~~~~..
,N,
M.. _0
N- _~mmqAm_&_
_ _ W
e..
...sls.."...mi. i_No ?--.X
do
am.FIG. 8. Measles virus polypeptides obtained by in vitro translation of fractionated mRNA. Equal reaction volumes of
[35S]methionine-labeled
in vitrotranslation products, obtained with mRNA eluted from each gel slice,wereimmunoprecipitated with anti-measles guinea pig serum. (a) no exogenous mRNA; (b) mRNA from uninfected CV-1 cells; (c) mRNA from measles virus infected cells; (1 through 30) mRNA eluted from successive slices ofagel segment containing fractionated mRNA of 2,700 (fraction 1) to 1,000 nucleotides (fraction 30). VOL.42, 1982I-, 1,
j A
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.487.91.409.253.613.2]C. 'i "I I-1- (i ' ?C
S-.Iii
.. a.
FIG. 9. Summary of the peak fraction foreachpolypeptide.
sid
protein
wasreported elsewhere
(6). The
isolation and
characterization of cDNA clones
that
appear tospecify the nucleotide
sequencescoding
for measles virus matrix
protein
and the
non-glycosylated
precursorfor
hemagglutinin
protein
arereported
here.
Aseries of 39 clones
containing measles virus
sequences wereidenti-fied
by colony
hybridization, using
ameasles
virus-enriched
[32P]cDNA
probe (S.
Rozenblatt,
F.
S.
Neumann,and C.
Gesang, submitted for
publication). The advantage of this
probe
is that
it
recognizes viral
nucleic acid
sequencesspecif-ically and thus directly
discriminates
amongclones
containing plasmids
with virus DNA
in-serts.
Translation in vitro of the measles virus
RNA
purified by
hybridization selection
has
shown that the
cloned
double-stranded
cDNA,
contained within
Cl-M, encodes
aportion
of the
measles
virus matrix
protein.
The
major
radioac-tive
polypeptide product
synthesized
in
acell-free
reticulocyte
translation
systemcomigrated
with
authentic
matrix
protein and
wasthe
only
polypeptide
immunoprecipitated by
antisera
against
measles virus. The double-stranded
TABLE 1. Correlation between measles virus mRNAsandpolypeptides
Molwt
Frac- Molwt No. of coding Molwt
tion of baesi capacity of
tionfl* mRNA(10
5)
bAmNAmRNAa
Ofa
protein(10-s)
(1O-3)9 7.2 2,115 84.0 65
14 5.9 1,735 68.1 60
15 5.7 1,677 65.7 70
40
18 5.15 1,515 58.9 35
aCalculating coding capacity: RNA length was cor-rectedfor the poly(A) tail considered to be 100 nucleo-tides; the average molecular weight of an amino acid
was 125.
cDNA contained within the clone G encodes a
portion of the measles virus non-glycosylated precursorforhemagglutinin protein. Two major
radioactive polypeptide products were
synthe-sized inacell-free reticulocyte translation
sys-temusing the RNAs selected by hybridization;
thelarger product migrated atabout 70K, and
the second migrated at 40K. This result was
obtained with seven independently isolated
cDNAclones ofgroupG.The only polypeptide
immunoprecipitated with anti-measles virussera
was the 70K polypeptide. Since polypeptides
synthesized in a cell-free reticulocyte are not
glycosylated, it is not surprising that the
non-glycosylated precursor for the hemagglutinin
migrated faster than theauthentic protein during
electrophoresis. The nature of this polypeptide
was determined by theuse ofmonoclonal
anti-measles virus glycoprotein sera. The nature of
the40Kpolypeptide is notknown.
A previous attempt was made to correlate
mRNA species with virus polypeptides on the
basis of molecular weight (8). In the present
study, the sizes ofindividual mRNA species of
measles virus were determined by two
tech-niques. In one approach, poly(A)-RNA from
cells acutely infected with measles virus was
fractionated on agarose gels under denaturing
conditions
(Glyoxal)
and transferred todiazo-benzyloxymethyl paper. The viral sequences
were then detected by hybridization with
32p-labeled individual cDNA clones (Cl-G, Cl-N,
and Cl-M). This study has shown that the sizes
of the mRNAs encoding the non-glycosylated
precursorforhemagglutinin and the major viral
nucleocapsid protein are similar (1,700 bases),
although the molecular masses of the two
pro-teins,asdeterminedbyelectrophoresis in
SDS-polyacrylamide gel electrophoresisare70K and
60K, respectively. The mRNA coding for the
matrixprotein is 1,550 bases long. The second
more general approach involved the separation
of different mRNA species into homogenous
w
..M.
^S
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.487.130.367.70.209.2] [image:7.487.50.242.515.633.2]density gradients and sedimented
together
with-in the 16S to 18S region. Electrophoresis on
agarose-methyl mercuric hydroxide
gels proved
to
be a suitable fractionation method; the
mRNAs could be resolved and
recovered from
the
gel while retaining their
biologically active
form. The present study indicated the presence
of
four mature functional mRNA species: 2,115,
1,735, 1,680, and 1,515 nucleotides in size, in
direct correlation with the size of their encoded
proteins. A 65K protein encoded
by a
2,115-nucleotide mRNA is of an unknown nature and
could be a
candidate for the Fo fusion protein.
The 60K protein, corresponding to
an mRNA of
1,735 nucleotides, and the 35K
protein,
corre-sponding to an mRNA of 1,515 nucleotides, are
most
likely the nucleocapsid (N) and the matrix
(M)
proteins,
respectively. The 70K to 72K
protein
encoded by a fraction
containing mRNA
of 1,680 nucleotides is the non-glycosylated
pre-cursor
for the G protein hemagglutinin. This
same mRNA fraction was also found to encode a
40K
polypeptide; the nature of the 40K protein
is not yet clear.
ACKNOWLEDGMENTS
S.R. thanks E. Winocour for his support and encourage-mentand B. Bloom and V. ter Meulen forprovidingmeasles virus monoclonal antibodies.
This work wassupportedin partbygrants(toS.R.)from the VolkswagenwerkStiftung,theU.S.-Israel Binational
Founda-tion,andthe IsraelMultipleSclerosisSociety. S.R.holds the
CharlesH. RevsonCareerDevelopmentChair.
LITERATURE CITED
1. Alwine,S.C., D. J. Kemp, and G. R. Stark. 1977. Method
for detection ofspecificRNAsin agarosegels bytransfer todiazobenzoyloxymethyl-paper and hybridization with DNAprobe. Proc. Natl. Acad.Sci. U.S.A.74:5350-5354.
2. Aviv,H., and P. Leder. 1972.Purification ofbiologically
active globin messenger RNA by chromatography on
oligothymidylic acid-cellulose. Proc. Natl. Acad. Sci.
U.S.A.65:1408-1412.
3. Bailey,J. M.,and N.Davidson.1976.Methylmercuryas a reversibledenaturingagent for agarosegel
electrophore-Electronmicroscopicevidence forsplicing of SV40 late mRNA.Cell 13:783-790.
6. Gorecki, M., and S. Rozenblatt. 1980. Cloning of DNA
complementary to the measles virus mRNA encoding
nucleocapsid protein. Proc. Natl. Acad. Sci. U.S.A.
77:3686-3690.
7. HaUl, W. W., W. R. Kiesling, and V. ter Meulen. 1978. Membraneproteinsof subacutesclerosing
panencephali-tis and measlesviruses. Nature (London) 272:460-462. 8. HaUl, W. W., W. R. Kiesling, and V. ter Meulen. 1978.
Biochemical comparison of measles and subacute
scleros-ingpanencephalitis viruses, p. 143-156. In R. D. Barry and B. W. J. Mahy(ed.),Negativestrand viruses and the host cell. Academic Press, Inc., London.
9. Hall, W. W., and S. J. Martin. 1973. Purification and characterization of measles virus. J. Gen. Virol. 19:175-188.
10. Lamb, R. A., P. R. Etkind, and P. W. Choppin. 1978.
Evidence forninthinfluenza viral polypeptide.Virology
91:60-78.
11. Morgan, E. M., and F. Rapp. 1977.Measles virus and its associateddiseases. Bacteriol.Rev.41:636-666. 12. Pelham, H. R. B., and R. J. Jackson. 1976. An efficient
mRNA-dependent translation system from reticulocyte
lysates. Eur. J. Biochem. 67:247-256.
13. Riccardi, R. P., J. S. Miller, and B. E. Roberts. 1979.
Purification andmapping of specific mRNAs by
hybrid-ization selection and cell-free translation. Proc. Natl.
Acad.Sci.U.S.A. 76:4927-4931.
14. Rigby, P. W. J., M. Dieckmann, C. Rhodes, and P. Berg. 1977. Labelling deoxyribonucleic acid to high specific
activityin vitroby nick-translation with DNApolymerase
I.J. Mol. Biol. 113:237-251.
15. Rozenblatt, S., M. Gorecki, H. Shure, and C. Prives. 1979.
Characterizationof measlesspecific proteins synthesized
in vivo and in vitro fromacutelyandpersistentlyinfected cells. J.Virol. 29:1099-1106.
16. Rozenblatt,S., and E. Winocour. 1972.Covalentlylinked
cell andSV40-specificsequencesin an RNA from
produc-tivelyinfected cells. Virology 50:558-566.
17. Schleuderberg, A. 1971. Measles virusRNA. Biochem.
Biophys.Res. Commun. 42:1012-1015.
18. Southern, E. M. 1975. Detection ofspecific sequences among DNAfragmentsseparatedby gelelectrophoresis.
J. Mol. Biol. 98:505-517.
19. Tyrrel, D. L. J., and E. Norby. 1978.Structural
polypep-tidesof measles virus. J. Gen. Virol. 39:219-229.
20. Waters, D. J., and R. H. Bussell. 1973. Polypeptide
compositionof measles and caninedistempervirus.
Virol-ogy55:554-557.
21. Wechsler, S. L., and B. Fields. 1978. Intracellular synthe-sis of measles virus specified polypeptides. J. Virol. 25:285-297.