0022-538XI82/070213-10$02.00/0
Effect
of Interferon on the Replication of Mink Cell
Focus-Inducing
Virus in Murine Cells: Synthesis, Processing,
Assembly, and Release of Viral Proteins
JOHN A. BILELLO,1*NELSON A. WIVEL,2AND PAULA M. PITHA1Biochemical Virology Laboratory, Johns Hopkins Oncology Center, Baltimore, Maryland 21205,1 and
Laboratory
of
CellBiology,National CancerInstitute,Bethesda,Maryland
202052
Received 14 December 1981/Accepted 15 March 1982
Treatmentof mink cell focus-inducing (MCF) virus (isolateAK-13)producing SC-1 cells with mouse fibroblast interferon (150 to 600 U/ml) led to a 100-fold
decrease in the release of infectious virus, whereas there was a 2.5- to 10-fold
decrease in variousparameters of virus particle release. Analysis of labeled virion
proteins indicatedthat a temporal change in virion protein composition occurred
after interferontreatment. After a 24-h exposure of chronically infected cells to
interferon,thevirionsproduced contained a85,000-dalton glycoprotein
(apparent-ly of nonviral origin) whichwas in excess of the virusenvelope glycoproteingp7O. Particles produced fromcells treated with interferon for 32 to 48 h were nearly
devoidofgp7Oand contained substantially lower quantities ofp30. Intracellular
processing of viralprecursor polyproteins to the mature virion structural proteins was not altered inthe presence of interferon. However, an accumulation of the
viral p30and pl2Eproteins was observed ininterferon-treated cells, consistent
withan increase incell-associated virions. Immunoprecipitation analysis of the
tissue culturefluids from
[35S]methionine-labeled
control and interferon-treated cells revealedamarked decrease inp30andpl5E/pl2E releasedafter interferon treatment. In contrast, gp7O did not accumulate in interferon-treated cells, but wasreleased into the culture medium in a form that was neither pelletable norassociatedwithpl5E/pl2E.
Interferons are afamily of cellular
glycopro-teins synthesized in response to viral infection
or nonviral inducers such as double-stranded
polyribonucleotides and certain halogenated
pyrimidols
(for review,seereference 16).Expo-sureofcellstointerferonresults in the
establish-mentofan"antiviral state"asmanifestedbythe
inhibitionof thereplication ofawide variety of DNA and RNA viruses. Although the
mecha-nism bywhich interferon inhibits viral
replica-tion isnotfullydefined, themajority ofviruses
are inhibited at the level ofmRNA translation
(16, 21, 22, 28). However, our own laboratory
and others have demonstrated that interferon
inhibits thereplication ofmurineretrovirusesin bothchronicallyandacutelyinfected cells, and this inhibitionoccursafter the
synthesis
ofviral RNAand proteins (4, 15, 32-35). Anumber of studies have suggestedthatinterferon interferes with retrovirus assembly and maturation, lead-ing to the "trapping" of the virions atthe cell surface(5,10, 34,36). Insystems wherevirionswere released frominterferon-treated cells, the
released particles had reduced
infectivity.
Ex-amination of the structural
proteins
ofvirionsreleased from interferon-treated cells had
indi-catedthatnewproteins, presumably incorrectly processed viral precursor polyproteins, were
incorporatedinto virions(9, 33).
Our previous results had indicated an
alter-ationin assemblyastheprimarydefect in
retro-virus production after interferon treatment of both chronicallyand denovo-infectedcells (32,
33, 36). Retrovirus assembly is a complicated
processrequiring the propertemporal and
spa-tialcleavage of threemajorprecursorproteins:
the gag precursor
(Pr67sar)
is cleaved to the virion core proteins (plO, p12, p15, and p30), gag-pol(Pr1809a59Pol)
is cleaved to the virionRNA-dependentDNApolymerase(and
presum-ablygag geneproducts),and the envprecursor
(gPr8Senv)
givesriseto the virionenvelopegly-coprotein gp7O and the virion matrix
protein
pl5E/pl2E (for review, seereference
40).
Viralpolyproteins function directly in the
assembly
process; for example, Pr675ag contains bothRNA-binding
proteins
which bind thegenomic
RNA and proteins whichspecifically
interact with the virion transmembraneprotein p15E,
affecting attachment of the viral core to the membrane. Theenvprecursor
gPr85nv
mustbe furtherglycosylated
and in aproperconfigura-213
on November 10, 2019 by guest
http://jvi.asm.org/
214 BILELLO, WIVEL, AND PITHA
tion in the membrane topermit both the interac-tion of plSE withPr679'9 and theformation ofa
stable disulfide linkage between pl5E andgp7O before proteolytic cleavage (7, 29, 31).
Wehaveattemptedto comparethesynthesis, processing, glycosylation, and release of viral proteins in thepresenceand absence of
interfer-on in order to determine whether interferon-induced alterations in these processes are
re-sponsible for the defective assembly. Our studies have concentrated on gp7O expression, since the envelope glycoprotein determines
vi-rus infectivity, neutralization, interference,and hostrangeproperties(reviewedinreference40). Additionally, the malignancy of murine
lympho-sarcomacell lines appears tocorrelate with the cell surface display of gp7O (37). Interferon-sensitive SC-1 cells infected with the envelope
gene recombinant mink cell focus-inducing
(MCF) virus isolated from AKR mice(19) were
used in this study. This virus-cell system has severaluniqueproperties relevant for studies of theeffect ofinterferononretrovirusreplication.
MCF viruses canbe titrated byfocus assayon mink cells whichareinsensitiveto mouse
inter-feron; thus, theinfectivity assayis notaffected
bythepossiblepresenceof residualmouse inter-feron in the system. Since earlier studies had indicated that interferonmayaffect viral precur-sorproteinprocessing,theuseof theSC-1/MCF
cell line with asubstantially slowerrate ofenv
geneprocessing (12) could amplifyminor inter-feron-induced differences in thispathway.
Last-ly, the in vivo administration of interferon has been showntoprolong thesurvival of AKR mice (18). Since this MCF virus is highly leukemo-genic and implicated in the appearanceof
s'pon-taneous T-cell lymphomas in AKR mice (19),information gathered from this in vitro study
may be relevant to the previously observed antitumor effect of interferon in this mouse
strain.
MATERIALSAND METHODS
Cells and viruses. Interferon-sensitive SC-1 cells chronically infected with the AK-13 isolate of AKR-MCF virus (19) were used. Virus was titered by enumerating cytopathic foci on mink lung cells (19).
Interferon.Experiments were performed with a
sin-glelot ofpurified mouse interferon induced in L cells with Newcastle disease virus (specific activity, 1.1 x 107reference units per mg of protein). The interferon was assayed in L929 cells by the cytopathic assay of Finter (13), using vesicular stomatitis virus as the
challenge virus. Interferon titers are expressed in internationalreference units; 1 U of theinternational standard titrated as 1 U in our assay. Subconfluent monolayers of chronically infected SC-1 cells were treated, 24 h after trypsinization and passage, with the indicated amount of interferon for the indicated length oftime.
Reversetranscriptase assay. The virus was pelleted
from thesupernatant fluid at100,000 x g in anSW40
rotor for 90 min, and the virion-associated reverse transcriptase was assayed as described previously
(34).
Electronmicroscopy. We prepared monolayers for transmission electron microscopy as described previ-ously (36). Thin sections were examined in a Hitachi HU-11Emicroscope withanacceleratingvoltage of 75 kV. Pellets of the fixedmonolayersweredividedinto multiple fractions, and sections from the separate fractions were used for countingtominimizesampling error and avoidrepetitive counting of a few cells.
Radiolabeling. In experiments where metabolically radiolabeledvirus or virion proteinsweretobe charac-terized, cells were labeled fora16-h interval either 8 or 32 h after interferon treatment (24- or 48-h virions, respectively). Labeling with [35S]methioninewaswith 50 plCi/ml in medium containing 5% of the normal level of methionine. Tritiated monosaccharides (AmershamCorp.,Arlington Heights, Ill.)wereused at 20,uCi/ml inamediumcontaining 10% the normal concentration of glucose.
Pulse-chaseexperimentswereperformedat24or48 hafter interferon treatment. Cellswere preincubated for1 hin mediumlacking methionine before labeling for 30 min with 125 ,uCi of [35S]methionine per ml (Amersham;specific activity, 1,100to1,300 Ci/mmol). Chase wasincomplete mediumcontaininga100-fold excessof unlabeled L-methionine. Cellswerewashed twice with phosphate-buffered saline before prepara-tion of cellextracts.
We performed lactoperoxidase radioiodination of cell surface proteins after washing the monolayers twice with buffer(phosphate-buffered saline[pH 7.4]
or 50 mM
N-2-hydroxyethylpiperazine-N'-2-ethane-sulfonic acid [HEPES; pH 7.4]-buffered saline as indicated). Cells were labeledat roomtemperature for 10 min in 1 mlof the indicated saline solution contain-ing 5 p.goflactoperoxidase (Calbiochem-Behring, B grade) 0.5 mCi of1251,and 10 pl1 of 0.03%H202. Ifno chasewasdesired,monolayerswerewashed andlysed
in extraction buffer. Chase was performed after the reaction was stopped byaddition of cold wash buffer
containing0.5 mMtyrosine.Cells were washed twice and transferred to fresh medium. After the indicated length of chase, the cells were washed and superna-tantsandcells were analyzed as described above.
Immunoprecipitation analysis of labeled proteins. Preparation of cell extracts and immunoprecipitation with monospecific goat antisera to purified retrovirus proteins was described previously (3). To compensate fordifferences in protein synthesis, each immunopre-cipitation was done with approximately 108 dpm of cell extract; the volume of each samplewas adjusted to 1 mlwith extraction buffer. Analysis of viral pro-teins released into the medium was done with 1.0 ml of clarifiedsupernatant(4,000x g, 5min) adjusted to 1% Triton X-100 and0.5%deoxycholate. Cell extracts and
supernatants were precipitated with normal goat se-rumbefore specific immunoprecipitationwith a previ-ously determined amount of antiserum. Staphylococ-cus aureus immunoabsorbant was used to collect antigen-antibody complexes by centrifugation. Pellets were washed twice with 20 mM Tris (pH7.4), 100 mM NaCl, 1 mM EDTA, and 0.5% Nonidet P-40 and once with the same mixture containing 2.5 MKCl.
Particles were analyzed aftercentrifugation through
on November 10, 2019 by guest
http://jvi.asm.org/
EFFECT OF INTERFERON ON MCF VIRUS REPLICATION
a 2%o sucrose cushion. Seven milliliters of radiola-beled tissue culturesupernatantwaslayeredover5 ml
of 20% sucrosein Tris-buffered saline (TBS; 20 mM Tris,pH 7.4,and 100 mMNaCI) and centrifugedfor 90
minat35,000rpminanSW41rotor at4°C.Tubeswere
fractionated fromthetop,and theupper4mlwasused
asapost-centrifugation sample.Justbeforeremoval of thesucroselayer, the sides ofthe tubeswerewashed
threetimes with TBS.Atapointwhere 1 ml ofsucrose
remained above thepellet, the sides of the tubeswere
again washed three times with TBS. The remaining
sucrose was removed, and the pellet was gently
washed twice with TBS.Pelletsweresuspended in the extraction buffer used for immunoprecipitation and analyzed either directly on polyacrylamide gels or
afterimmunoprecipitation with specific antisera. Anal-ysis of virus prepared by bandingonsucrosegradients
gaveidentical results.
Polyacrylamide gelelectrophoresis and quantitation of labeled protein bands. Samples were analyzed on
12.5 or 15% sodium dodecyl sulfate-polyacrylamide
slabgels, using the discontinuoussystemdescribed by Laemmli (24). Fluorographywasperformed according
tothe procedure ofBonnerand Laskey (8), and the dried gelswereexposedtopreflashed Chronex4X-ray filmat-70°C.
Autoradiographs of the dried gelswerequantitated
by theuseofanintegratingspectrophotometer (Ortec model 4310) whenanappropriateexposuretimewas
available in which the film gave a linear response.
Routinely, labeled bandswereexcised fromthedried
gels, dissolved in 0.3 ml of 15% hydrogen peroxide, and counted in Aquasol II liquid scintillation fluid (NewEngland Nuclear Corp.). Thetwomethodsgave
comparable results.
RESULTS
Interferon treatmentof MCF virus-producing SC-1cellsleadstotheproduction of noninfectious
virus. SC-1 cells chronically infected with the AK-13 isolate of MCF viruswere treated with
150 to600 U ofpurified mouse fibroblast
inter-feron for 24 or 48 h. The levels of infectious
viruspresentinthemedium ofinterferon-treated
cultures were reducedby 100-fold, but reverse
transcriptase assay of the same supematants
indicated onlya10-foldreduction in virus
parti-cles. Metabolic labeling of virions with [35S]methionineindicateda2.5-to10-fold
reduc-tion in virusparticles(Table 1). Theseresultsare
consistent with earlier studies with
AKR-L1-and Rauscher virus-infected cells which had
indicated that noninfectiousvirusparticleswere
formed inthepresenceof interferon.
Analysis of the numberofcell-associated
vi-rusparticles produced by interferon-treated
SC-1/MCFcellswasperformed byelectron
micros-copy.Transmissionelectronmicroscopy(TEM) of representative sections of SC-1/MCF cells
indicatedthat the block in virus release occurred at astep afterbudding. The numberofplasma membrane-associated virus particles visible by TEMofthinsectionswasincreasedby
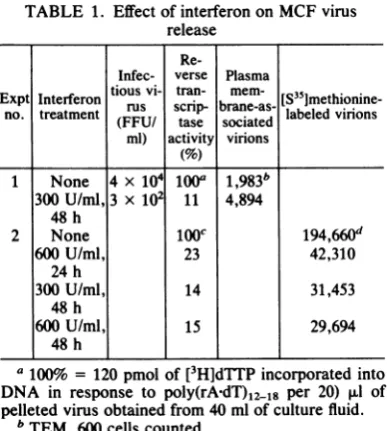
approxi-TABLE 1. Effect ofinterferononMCF virus release
Re-Infec- verse Plasma
Exp
Inerfrontious
vi- tran- mem-[S35]methionine-Expt Interferon rus scrip- brane-as- labeled virions no. treatment (FFU/ tase sociated
ml) activity virions
1 None 4 x104
ioo'
1,983b300U/ml,3 x 102 11 4,894 48 h
2 None lOOc 194,660d
600 U/ml, 23 42,310
24 h
300U/ml, 14 31,453
48 h
600U/ml, 15 29,694
48 h
a 100% = 120
pmol
of
[3H]dTTPincorporated into DNA in response to poly(rAdT)12_18 per 20)RI
of pelleted virus obtained from 40 ml of culture fluid.bTEM, 600 cellscounted.
c
100o
= 223,000 cpmincorporated.dTotal hot trichloroacetic acid-precipitable cpm in
samples of[35S]methionine-labeledvirusparticles
pre-paredasdescribed in Materials and Methods.
mately 2.5-fold in interferon-treated cells when
compared with untreated SC-1/MCFcells(Table
1).We havepreviouslyshown that theestimates of virus particlesonthe cell surface quantitated by scanning electron microscopy and computer-generated image analysisarein complete agree-mentwithTEM estimates(36). Ultrastructural-ly, there was no detectable difference between virions assembled in the presence and absence of interferon.
Viral protein synthesis and processing in con-trol and interferon-treated cells. Viral protein
synthesis and processingwerestudiedby pulse-chase labeling and immunoprecipitation
analy-sis. Since other authors (27) have indicated that interferon may exert its inhibitory effect on
membranebudding viruses by virtue ofa
tunica-mycin-like activity associated with or induced
by interferon,
tunicamycin-treated
cells werealso included for comparison. Figure 1 shows the synthesis and processing of
gp7O
in SC-1cells infected with MCF virus as well as in
infected cells treated with interferon or tunica-mycin. Unlike tunicamycin treatment, which leadstothe accumulationofa
nonglycosylated
precursor to the virion envelopeglycoprotein
(Pr69enlV),
treatment with interferon didnotleadto aninhibition of
glycosylation
orprocessing.
The majordifference observed inthe
synthesis
and processing of the
envelope
glycoprotein
precursor related to the accumulation of thenonglycosylated virion matrix
protein
pl5E/
VOL.43, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.265.459.78.294.2]L
FIG. 1. Anti-gp7. immunoprecipitates of cell
ex-tractsfrom SC-1/MCF cells treated with interferonor
tunicamycin.SC-1/MCF cellsweretreatedwith 600 U ofpurified mouseinterferon per ml for 48 h before a
30-mm pulse-labeling with [35S]methionine as de-scribed in the text. Parallel control cultures were
eitherlabeledatthesametimeortreatedfor1hbefore
labelingwith 1 pLgoftunicamycinperml. Cellextracts
werepreparedeitheratthe end of the pulseoraftera
90-min chase in medium containinga100-foldexcess ofunlabeled L-methionine. Immunoprecipitation with anti-gp7Owasasdescribed in thetext.
theimmunoprecipitation shown,where thesame number ofcounts was used for each
precipita-tion.The observed lack of effect oftunicamycin
onprocessingofPr67919andrelease ofp30into the supernatants of tunicamycin-treated SC-1
cells infected with MCF virus correlated with the resultsfound in othersystems(Bilello,
War-necke, Koch,andOstertag, submitted for publi-cation). Chase of the reverse transcriptase
pre-cursor,
PMWag-PO',
also appeared to proceednormally in interferon-treated cells. The
-195,000-molecular-weight protein which ran above PMWag-Po' and was found in anti-gp70,
-p30,and-pl5E immunoprecipitateswas an SC-1 cytoskeletal protein which contaminated
im-munoprecipitates made from interferon-treated
cells. Both thecharacterization andsignificance
of the interferon-induced increase in the 195-kilodalton proteinareunderinvestigation.
Release of viralproteinsfromcontrol and
inter-feron-treated cells. Our data indicated that the effect ofinterferon on MCF viralprotein
synthe-sis and processing was minimal. However, an
intracellularaccumulationofp30andpl5E/pl2E
wasobserved,whereas no increase in the
intra-cellular level of gp7O occurred. Since other studies have indicated thatgp7Ocanbe released
EISERASE
NTLERFERO - - _
-TLUrICvli CN
pl2E in interferon-treated cells. When cell ex-tracts wereimmunoprecipitated with antiserum
to the virion plSE protein (Fig. 2), there was
little difference in the amount plSE and a
marked increase in pl2E in interferon-treated
cells, whereas in control cells only plSE was
detected. Previous studies (23; Bilello,
unpub-lished data) as well as those described below
indicated thatpl5E isprimarily cell associated and pl2E islocalized in virions, where it is the
major p1SE-related protein. It was recently
shownthatpl5Eis aprecursorofpl2Eand that apeptide from the carboxyterminusofpl5E is removed during itsprocessing topl2E(17).
Analysis of the virion p30-related proteins
indicated no major differences in the cleavage
patternofPr67gag (Fig. 3). Ininterferon-treated cells, a small but reproducible accumulation of
p30 (three- to fivefold increase) was measured
by quantitation of the autoradiograph ordirect
determination of label in the p30 band. The
increasein cell-associatedp30isaccentuated in
GpR85EN
GP/uO
-.- .sm fa ins
p[E,9ENf
-FIG. 2. Anti-pl5E immunoprecipitates ofcell ex-tractsfrom SC-1/MCF cells treated with interferon or
tunicamycin.Anti-pl5E immunoprecipitates were pre-pared as described in the legend to Fig. 1 and analyzed on a 15% acrylamide-0.2% bisacrylamide slab gel to enhance the separation between pl5E and pl2E.
on November 10, 2019 by guest
http://jvi.asm.org/
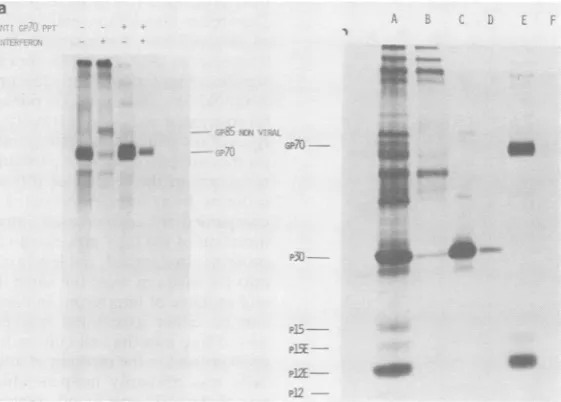
[image:4.504.55.246.80.314.2] [image:4.504.260.451.369.618.2]EFFECT OF INTERFERON ON MCF VIRUS REPLICATION 217
[image:5.504.74.243.79.294.2]em~~~~~~~~~~~~~~~~~~~~~~~..
FIG. 3. Anti-p30 immunoprecipitates of cell
ex-tractsfrom SC-1/MCFcelltreated with interferon or tunicamycin. Anti-p30 immunoprecipitates were
pre-pared from extracts of control, tunicamycin-treated, andinterferon-treatedcells as described in the legend toFig. 1.
either freeorassociated with virus particles (6;
Bilello,unpublisheddata), wedecided to
exam-inetheeffectof interferon on the release of viral proteins as well as the incorporation of these
proteins into virus particles. Control and
inter-feron-treated cells(600U/ml;48h)werelabeled for 30 minwith[35S]methionineand thenchased for2, 4, and 24 h in medium containing a 100-foldexcessofunlabeled L-methionine. Analysis
of gp70 in thetissue culture fluidatvarious time
points duringthe 24-h chase indicated that there
was nodifference in the release of labeled gp7O into the medium in the presence andabsence of interferon (Fig. 4a). Similar analysis of p30
re-leaseindicatedamarked decrease (73 to87%)in the release of p30intothe supernatant(Fig.
4b)
ofinterferon-treated cells.
Figure5showsanimmunoprecipitation
analy-sis oftissue culture fluidsfrom cells labeled with [35S]methionine before and after centrifugation
toremove virusparticles. In lanes A throughD,
equal volumes (1 ml) of labeled tissue culture fluid were immunoprecipitated with either anti-p30 (panel a) or anti-gp7O (panel b) serum. A comparison of the levels of p30 present in the supernatant from untreated MCF
virus-produc-ing SC-1 cellsbefore (lane A) and after (laneC)
centrifugation indicated that nearly all (88 to
94%) of thep30 in the supernatantwas associat-ed with particles. Tissue culture fluids from interferon-treated SC-1/MCF cells contained verylittle p30, and nearly all of it appeared tobe sedimentable. Similar analysis using anti-gp7O
serumindicated that although gp7O was present inthe supematants of both control and interfer-on-treated cells, areduction (38 to 43%) in the amount of gp7O present after pelleting of the virus was observed only in supernatants from
untreated cells (Fig. Sb).
b
a
N Iso
gas
FIG. 4. Immunoprecipitationofsupernatant fluids from[35S]methionine-pulse-labeledcontrol and interferon-treatedcellsatvarious times aftertransfertomediumcontaininga100-foldexcessof unlabeled L-methionine. (a)
Anti-gp7O immunoprecipitates; (b)anti-p30 immunoprecipitates.
VOL.43,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.116.410.443.648.2]218 BILELLO, WIVEL, AND PITHA
a~
a fVkI Dr,
IrMT'EFs,Jj _
-4.
FIG. 5. (a) Anti-p30 [35S]methionine-labeled cell from MCF virus-infected S fromuntreated cells;B,sup(
with 600 U of interferon p supernatant fluid as in lar
200,000 x gfor 90minover removevirusparticles; D,s
fugedasdescribedabove.(b
pitates.
Analysis of virion strui formed on particles rele
interferon-treated cells
either [35S]methionine o
rides. A study of virion interferontreatment rev( inp30releasedescribed
al in virions labeled bel interferon treatment (da
er, the amount of viric
substantially lower,and
new glycoprotein ofapj tons which was the ma
labeledwith[3H]glucosa
kilodalton glycoprotein
pitated with antiserum virusparticles, butnotN viralproteins (33; Fig.6
data), and represented presentin controlvirion
Figure 6b shows the
[35S]methionine-labeled
48 h of treatment with inhibitoryeffectonvirus
production was maxima virionsreleasedfromun
virion structural proteii
protein component, wh
werealsopresent in a ra foundinothervirusprep in the middle ofa groul proteins. The identity of
confirmed
by
immunoprecipitations
of the virion.
.
....preparations
...
with antisera to thepurified
pro-teins
p30
(lane C) andgp70
(lane E).
Whenparticles frominterferon-treated cells were
ana-lyzed, amarked difference in the virion compo-sitionwasfound. Verylittlep30or
lower-molec-ular-weight virion proteins were sedimentable
from the supematants from interferon-treated
cells
(Fig.
6b,lanes BandD).Immunoprecipita-tion of MCF
particles
from untreatedvirus-in-fectedSC-1 cells with
anti-gp70
seraprecipitated
gp7O as well as the virion matrix
protein
pl2E
(and asmallamount ofunprocessed
pl5E)
(Fig.
6b, lane E). In contrast,
only
asmall amountofgp70andnopl2Ewas
immunoprecipitated
fromparticle preparations from interferon-treated
MCF-infected SC-1 cells (lane F).
Analysis
of immunoprecipitates of virion phosphoproteins labeled with32P,failedto 1-free tissue culture fluids indicate any alteration of the32P-labeled
species C-i cells. A, Supernatant(Pr67ag p12,
and other minor species) present enatantfrom cellstreated in thevirion;
however, the approximate level ofer ml for 48 h; C, same
32P-labeled
p12
protein
was
reduced fivefold
in
a20%
sucrosencushion
to MCF virions released from interferon-treatedame as lane B, butcentri- SC-i cells (data notshown).
)Anti-gp7Oimmunopreci- Interferon-mediated inhibition of murine
leu-kemia virus releasepersists aslongasinterferon is present in the culture medium, and virus production resumes within 16 to 24 h after
cturalproteins was per- interferon removal (34). A reversal
experiment
-ased from control and was performed as follows. Cells were treatedlabeled for 16 h with with interferon for 48 h and then washed and
r tritiated monosaccha- refed with medium without interferon. At 24 h
is produced early after after removal of interferon, the cells were la-ealed that thereduction beled for 16 h with [35S]methionine, and the
above was only margin- labeled supernatants were analyzed for virion tween 8 and 24 h after proteins. Supernatant fluids from parallel cul-ta not shown). Howev- tures labeled between 32 and 48 h after
interfer-in-associated gp70 was on treatment were also immunoprecipitated to
the virions contained a indicate the extent of the block in retrovirus proximately 85,000 dal- release before reversal. Interferon treatment
re-jor virion
glycoprotein
sulted in a marked decrease inpl5E/pl2E
re-Lmine (Fig. 6a). The 85- lease into the supernatant as well as a lack of could be immunopreci- association of gp7O with either pl2E or
pl5E
prepared against total (Fig. 7). Transfer of the interferon-treated cells with antiserato purified to medium without interferon resulted in the and Bilello,unpublished increased release ofp30 (data not shown) and
a cellular glycoprotein the association ofgp7O with
pl5E
and pl2Einks as a minor species. immunoprecipitates ofsupernatants (Fig. 7) or
protein composition of virions (data not shown).
virions harvested after Glycosylation of gp7O after interferon
treat-i interferon, when the ment of AKR/MCF-infected SC-1 cells. One
pos-sinfectivityandparticle siblemechanism for inhibition of the infectivity d. Lane A shows MCF of membrane budding viruses was the proposed itreated SC-1 cells. The tunicamycin-like activity induced by interferon n, p30, was the major (26). Although the experiments described above
iereas pl5E and pl2E indicated thatgp7Owas glycosylated, processed,
ttioconsistent with that and shed,we were interested to ascertain wheth-arations. gp7Oisvisible er there were any detectable alterations in the
p ofnon-virion-specific carbohydrate moiety of
gp7O
which would leadf thelabeled bands was to its inability to be assembled into virions.
.. _
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.51.243.67.247.2]EFFECT OF INTERFERON ON MCF VIRUS REPLICATION 219
a
m
..::x
FIG. 6. (a)Glycoprotein composition of virus particles produced by control and interferon-treated cells.
SC-1/MCFcells werelabeled for16hwith tritiated glucosamineandpelleted througha20% sucrosecushionas
described in thetext.Thefirsttwolanes show the glycoproteincomposition of particlesproduced by control and
interferon-treated cells. The last two lanes indicate anti-gp7O immunoprecipitates from the two particle
preparations. (b) [35S]methionine-labeledparticles released from controlcells and cells treated for 48hwith600
Uof interferonperml.(A)Controlparticles; (B) particlesfrom interferon-treated cells; (C)anti-p30 precipitation
ofA; (D) anti-p30precipitation ofB; (E) anti-gp7O precipitationof A; (F) anti-gp70 precipitation ofB.
When gp70 labeled with
[3H]glucosamine,
[3H]mannose, or [3H]fucose was analyzed in
virionsor intissueculturefluids before and after
centrifugation, apatternsimilar to thatobserved for [35S]methionine-labeled gp7O was found
(datanotshown). Although we observed minor
differences in labeling with monosaccharides,
wefound noevidencefor an overt alteration in
the glycosylation of free or virus-associated
gp70. Differences in the amount of carbohy-drate-labeled MCF virusgp7O released by
con-trolandinterferon-treated SC-1/MCF cellswas
generally proportional to the extent of overall inhibition of protein synthesisseenin interferon-treated cells (30to50% of control).
Expressionofgp7Oonthesurfaceof interferon-treatedSC-1/MCF cells.Our studies of
metaboli-cally labeled viral proteins did not exclude the
possibility that interferon treatment leads to a redistribution of viral glycoproteins in the cell membranewhichmayleadtoinefficient
assem-bly. It was previously reported that unlike in other retrovirus systems, cell surface iodination ofMCF-infected SC-1 cells leads tothe
detec-tion of both labeled gp7O and gPr85env after
immunoprecipitation (12).We
performed
similarexperiments to determine whether we could
determineaninterferon-induced alterationinthe
labeling of gp7O, its precursor, or both. Our results indicated that detection of
gPr85eenV
onthe cell surface appearedto beprimarily due to
the low viability of the cells in phosphate-buff-ered saline. Labeling in HEPES-bufferedsaline,
which maintains -100% cell viability and a
minimum of cell breakage (as indicated by a
failure to label the intracellular protein actin), resulted in labeling ofgp7O exclusively. Under theseconditions, nodifferencewasobserved in the accessibility ofgp7O to lactoperoxidase io-dination between control andinterferon-treated
cells(datanotshown).
Theturnoverofgp7Oonthe surfaceof control and interferon-treated cells was examined by following the fate ofiodinated
gp7O
at various times of chase. Control and interferon-treatedcells were iodinated and chased for 0.5or 3 h.
Theamountof labeled
gp7O
associated with the cellorpresentin virionswasthendeterminedby
immunoprecipitation
as described above. Nodifference was observed in the levels of
gp7O
associated with control and interferon-treated
cells. However, theamount of125I-labeled
gp7O
present in virions released at 3 h after
labeling
wassignificantly lowerininterferon-treatedcul-tures(Fig. 8), whichcorrelates withourresults
with metaboliclabeling.
DISCUSSION
The resultspresentedhereindicatethat
inter-ferontreatmentof SC-1 cells
chronically
infect-"AM
ia
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.122.403.83.284.2].Pi P15E
a BC D
GP70
--P15E
-
P12E---"'IS'.
_4
St
..FIG. 7. Effectofthe transfer of interferon-treated cellstonormal mediumonthe release of viralproteins
intothetissue culturemedium.Anti-pl5E immunopre-cipitates of[35S]methionine-labeledvirionproteins in the supernatants of control, interferon-treated, and interferon-treated cells shifted for 24 h to normal growth medium for24hbefore labeling.(A)
Superna-tants from control cells labeled with [35S]methionine from32to48 hasdetailed in thetext.(B)Supernatants fromSC-1/MCFcellslabeledbetween 32 and 48 h after
treatmentwith 600 U ofinterferonperml.(C)
Super-natants from a culture of untreated cells grown in
parallel to those described in A, but grown for an
additional 24h infresh normalgrowthmedium before
a16-hlabeling with[35S]methionine.(D)Supernatants from an interferon-treated culture grown for48 h in paralleltotheinterferon-treatedcells analyzed in lane B, buttransferredtofresh mediumwithoutinterferon for 24 h before a 16-h labeling in the absence of
interferon.
ed with MCF virus leads to the formation of noninfectious virions which are deficient in
gp70. No evidence was obtained for the
inhibi-tion of viral protein synthesis or processing. Similarly, wecould notdetect apreferential or
increased incorporationof uncleaved virion
pre-cursorproteins into virionsreleased from
inter-feron-treated cells. Intracellular levels of the
fully processed p30orpl5E/pl2E proteinswere
elevated ininterferon-treated cells.Precipitation
of the cell-free supernatants from interferon-treatedcells withanti-pl5Eserafailed todetect
significantlevels of eitherpl5Eorpl2E (Fig.7). Althoughtheamountofp30released from
inter-feron-treatedcellswasmarkedly reduced, near-lyall of thep30 released from control or interfer-on-treated cells was in a particulateform. The reduction in the release ofp30 and pl5E/pl2E
proteins from interferon-treated cells was
ac-companied by an increased intracellular
accu-mulationof the fully processed virion structural proteins. Incontrast,the levels ofgp7Oreleased into the medium werethe samein the presence
andabsence of interferon. In control cells,gp7O
can be either assembled into particles or
re-leased free into the cell culture fluid. However,
gp7Opresentin the medium ofinterferon-treated
cells was primarily nonpelletable, and little if
anyof thegp7Owasfound associated withvirus
particles.
The second major difference found was that
no gp7O/pl2E complex was detected in either virions or tissue culture fluids from interferon-treated cells. The gp7O/pl2E complex can be readily demonstrated by reciprocal co-immuno-precipitation of untreated SC-1/MCF cells, since disulfide bonds between gp7O and pl2E are
maintainedthroughout theimmunoprecipitation
procedure, and the individual proteinsare
sepa-rated by reduction of the complex with 13-mer-captoethanol before electrophoresis. There are two major interpretations of these data: first,
FIG. 8. Anti-gp7O immunoprecipitates of cell
sur-face-iodinatedcellextracts and particles released from control and interferon-treated cells. (1) Cell extract preparedimmediately after iodination; (2) cell extracts aftera30-minchase; (3)particles after a30-minchase; (4) cell extract after a 3-h chase; (5) particles after a 3-h chase.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.73.221.71.394.2] [image:8.504.258.447.449.612.2]221
that gp7O is particle associated in interferon-treated cells, but because of the lack of
associa-tionwithpl5E/pl2Eisverysusceptible to shear forces generated during centrifugation; and sec-ond, that gp7O is not assembled into particles and isreleased from pl5E/pl2E after proteolytic cleavage ofgPr85e"v. Several lines of evidence favor the latter hypothesis: (i) virus particles associated with thecells have a decreased
infec-tivity (36); (ii) the amount of gp7O detected in virions isindependent of virion preparation and purification; (iii) failure to detect a gp70/pl2E
complex in the medium before centrifugation;
and (iv) the startling difference in the kineticsof release for gp7O compared with p30 and pl5E/ pl2E.
Themolecular basis for thefailure of gp7O to be incorporated into retroviruses produced by interferon-treated cells is yet unknown. Mahesh-wari et al. (26) observed that interferon treat-mentof L-cells infected with vesicular stomatitis virus resulted in the production of vesicular
stomatitis virus particles which had markedly reducedlevels of theglycoprotein (G) and mem-brane protein (M). They also reported that
[3H]glucosamine
incorporation into the gp7O ofMoloney leukemia virus particles was reduced (12). Maheshwari et al. later related these find-ings to an inhibition of the formation of
aspara-gine-linked oligosaccharide chains, an effect similar to that induced upon exposure of cells to
tunicamycin(6). We would like to suggest that
since they analyzed only virus particles, the apparentlack of[3H]glucosamine-labeled
enve-lopeglycoproteinmayhave been due to afailure to incorporate viral glycoproteins into virions. In the SC-1/MCF system, as well as in other
cellswehave studied,noinhibitionofN-linked
glycosylation in the presence of interferon has beenobserved (J. Bilello and P. M.Pitha,
manu-script in preparation). The envelope
glycopro-tein both from control and interferon-treated
cells contains mannose, fucose, and glucos-amine. Aboud et al. (1) recently reported an
early decrease in the rate of [3H]glucosamine incorporation into the envelope
glycoprotein
precursor in interferon-treated cells.
Although
wehavenotbeenabletoconfirmthese
observa-tions, there may be differences in
methodology
andthecellsystem studied. Onepossible mech-anism for the failure to incorporate
gp7O
intovirus particlescomesfromnucleotideandamino acid sequenceanalysisof
gp7O (39;
S.Oroszlan,
personalcommunication),where itappears that the majorityof the
carbohydrate
acceptor sitesare at the
carboxy-terminal
end ofgp7O.
It isconceivable that an altered
carbohydrate
sidechain could interfere with the formation orthe
stability of disulfide bonds between
gp7O
andpl5E/pl2E, allowing the release of
gp7O
uponproteolytic cleavage ofgPr85e"v. Alternatively, conformation changes at the membrane either
duetointerferon-induced alterations in gp7O or membrane stiffening (11, 30) may prevent the stable association between gp7O and pl5E/12E
orlead to anincreased release of gp7O at points distal to viralmaturation sites. Experiments are inprogress to distinguishbetween these
alterna-tives.
Apart from their effect on virus replication,
interferons affect a number ofcellular functions and processes. Although we have found no
interferon-induced alteration in the expression
ofgp7Oat the cellsurface, a number of studies have indicated thatinterferon alters the expres-sion ofhistocompatibilityand otherantigens (for review, see reference 41). We also have found noalteration in the total amount of gp7Oreleased orin thekineticsof this process, whereas others havereported changes in the release of plasmin-ogen activator (20, 38) and carcinoembryonic
antigen (2). We would like to stress that our
observations oncell surface expression and re-lease of gp7O apply only to this system. The studies ofplasminogen activator releasealluded
to above have indicated that interferon can
eitherpotentiate or inhibit the release of a single protein, depending on the cell type or system studied.
Studies of themembrane budding virus vesic-ularstomatitis virus have indicated that specific host cell surface proteins can be incorporated into virions even in the absence of the viral envelope (G) glycoprotein (25). We have noted thatafter treatment ofSC-1/MCF-infected
cells
withinterferon,there is atemporal difference in theaccumulation of a cellular85-kilodalton
gly-coprotein into virions. This furtherstrengthens
our suggestion that the restriction in virus
as-sembly is not selectively localized at the virus structuralproteins,but maybeaconsequence of
an interferon-induced change in plasma mem-brane ofthe infected cells, e.g., a decrease in
fluidity.
ACKNOWLEDGMENTS
We thank S. Oroszlanfor his generous giftofanti-pl5E
serumand David Carbone for assistance withradioiodination studies.
Thisstudywassupported byPublic HealthServicegrant 5ROI-CA19061-06(to P.M.P. from the National Institutes of Health) and by American CancerSociety grantIN-11R (to
J.A.B.). P.M.P. isaLeukemiaSocietyscholar.
LITERATURECITED
1. Aboud,M.,R.Knmchl,M. akhhv lf,andS.Salzberg.
1981.Intracellularproductionof virusparticlesand viral components in NIH3T3 cellschronically infected with Moloney murineleukemiavirus: effect of interferon. J. Virol.40:830-838.
2. Attallah,A. M.,C. F.Needy,P.D. Noguchi,and B. L. EHsberg. 1979. Enhancementofcarcinoembryonic antigen VOL.43,1982
on November 10, 2019 by guest
http://jvi.asm.org/
BILELLO, WIVEL, AND PITHA
expressionby interferon. Int. J. Cancer 24:49-52. 3. Bilello, J. A., G. CoUletta, G. Warnecke, G. Koch, D.
Frisby,I. B.Pragnell, andW. Ostertag.1980.Analysis of the expression of spleen focus forming virus
(SFFV)-related RNA and gp55, a Friend and Rauscher
virus-specific protein.Virology 107:331-344.
4. Billiau,A.1977. Effect ofinterferononRNA tumorvirus. Tex.Rep. Biol. Med. 35:406-419.
5. Billiau, A., H. Heremans, P. T. Allen, J.
DeMaeyer-Guignard, and P.DeSomer. 1976. Trappingof oncorna-virus particlesat the surface of interferon treated cells.
Virology73:537-542.
6. Bolognesi, D. P., A. J. Langlois,and W. Shaefer. 1975.
Polypeptidesof mammalian oncornaviruses.IV. Structur-al components of murine leukemia virus released as solubleantigensin cell culture. Virology68:550-555. 7. Bolognesi, D. P., R. C. Montelaro, H. Frank, and W.
Schaefer. 1978. Assembly of type C oncornaviruses: a model.Science 199:183-186.
8. Bonner,W.M.,andR. A. Laskey. 1974.Afilm detection method fortritium-labeled proteins and nucleicacids in
polyacrylamidegels. Eur.J. Biochem.46:83-88. 9. Chang,E.H.,andR. M.Friedman. 1977. Alarge
glyco-proteinof Moloneyleukemia virus derived from interfer-on treated cells. Biochem. Biophys. Res. Commun. 77:392-398.
10. Chang, E. H., S. J. Mims, T. J. Triche, and R. M. Friedman. 1977. Interferon inhibitsmouseleukaemia vi-rusrelease: anelectron microscope study. J. Gen. Virol. 34:363-367.
11. Chatterjee, S., H. C. Cheung, and E. Hunter. 1982. Interferon inhibits Sendai virus-induced cell fusion an effectoncell membrane fluidity. Proc. Natl. Acad. Sci. U.S.A. 79:835-839.
12. Famulari, N. G., and K. Jelalian. 1979. Cell surface
expressionof dualtropic MCFmurine leukemia virus. J. Virol. 30:720-728.
13. Finter,N. B.1966.Interferonassaysandstandards,p. 87-93. N. B. Finter (ed.), Interferons. North-Holland
Pub-lishingCo., Amsterdam.
14. Friedman, R.M., R. K. Maheshwari, F. T. Jay, and C. Czarniecki. 1980. Mechanism of interferon inhibition of viruses that bud from the plasma membrane. Ann. N.Y. Acad. Sci. 350:533-543.
15. Friedman, R.M.,and J. M.Ramseur. 1974. Inhibition of murine leukemia virusproduction in chronically infected AKR cells: anoveleffect ofinterferon.Proc.Natl.Acad. Sci. U.S.A.71:3542-3544.
16. Gordon, J., and M. A. Minks. 1981. The interferon
renaissance: molecular aspects of induction and action. Microbiol. Rev. 45:244-246.
17. Green,N.,T.M.Shinnick, 0. Witte,A. Ponticelli, J. G.
Sutcliffe,andR.A.Lerner. 1981. Sequence-specific anti-bodies show thatmaturation of Moloney leukemia virus
envelopepolyprotein involves removalof a COOH-termi-nalpeptide.Proc.NatI. Acad. Sci. U.S.A.78:6023-6027. 18. Gresser, I., J. Coppey, and C. Bourali. 1968. Interferon and murine leukemia. VI. Effect of interferon prepara-tionson thelymphoid leukemia of AKR mice. J. Natl. CancerInst.43:1083-1089.
19. Hartley, J. W., N. K. Wolford, L. J. Old,and W. P. Rowe. 1977. Anewclass ofmurineleukemia virusassociated with development of spontaneous lymphomas. Proc. Natl.Acad. Sci.U.S.A. 74:789-792.
20. Hovi, T., 0. Saksela, and A. Vaheri. 1981. Increased secretion of plasminogen activator by human
macro-phages after exposure to leukocyte interferon. FEBS Lett.129:233-236.
21. Joklik,W.K., and T. C.Merigan. 1966. Concerning the mechanism of action ofinterferon. Proc. Natl. Acad. Sci. U.S.A.56:558-565.
22. Jungwirth, C., J. Horak, G. Bodo, J. Lindner, and B. Schultze.1972.Thesynthesis ofpoxvirus-specificRNAin
interferon treated cells. Virology48:59-70.
23. Karshin, W. L., L. J. Arcement, R. B. Naso, and R. B. Arlinghaus. 1977. Commonprecursorfor Rauscher leuke-mia virus gp69/71, p15(E), and p12(E). J. Virol. 23:787-798.
24. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature(London) 227:680-682.
25. Lodish,H.F.,and M. Porter.1980.Specificincorporation ofhostcell surfaceproteinsintobuddingvesicular stoma-titis virusparticles. Cell 19:161-169.
26. Maheshwari, R. K., D. K. Banerjee, C. J.Waechter, K. Olden, andR. M. Friedman. 1980. Interferon treatment inhibitsglycosylation ofaviralprotein. Nature(London)
287:454-456.
27. Maheshwari, R. K., A. E. Demsey, S. B. Mohanty, and R. M. Friedman. 1980. Interferon treated cells release vesicular stomatis virus particles lacking glycoprotein spikes: correlation with biochemical data. Proc. Natl. Acad. Sci. U.S.A. 77:2284-2287.
28. Metz, D. H., and J. Esteban. 1972.Interferon inhibits viral
protein synthesis in L cells infected with vacciniavirus. Nature(London) 238:385-388.
29. Oroszlan, S., L. E. Henderson, T. D. Copeland, A. M. Schultz,and E. M. Rabin. 1980.Processingandstructure of murine leukemia virus gag and env gene precursor proteins, p. 219-239 In G. Koch and D. Richter (ed.),
Biosynthesis,modification, andprocessingofcellularand viral polyproteins. Academic Press,Inc., New York. 30. Pfeffer, L. M., F. R.Landsberger,and I.Tamm. 1981.
-Interferon-induced time-dependentchangesin theplasma
membrane lipid bilayer of cultured cells. J. Interferon Res. 1:613-619.
31. Pinter, A., and E.Fleissner.1977. The presenceof disul-fide linkedgp70-pl5E complexes in AKR murine leuke-mia virus. Virology 83:417-422.
32. Pitha, P. M. 1980.The effects of interferoninmousecells infected with MuLV. Ann.N.Y.Acad. Sci. 350:301-313. 33. Pitha, P. M., B. Fernie, F. Maldarelli,T. Hattman, and N. A. Wivel. 1980. Effectof interferon on mouseleukemia virus. V. Abnormal proteins in virions of Rauscher MuLV produced in the presence of interferon. J. Gen. Virol. 46:97-110.
34. Pitha, P. M., W. P. Rowe, and M. N. Oxman. 1976. Effect of interferon on exogenous, endogenous and chronic murine leukemia virus infection.Virology 70:324-338. 35. Pitha, P. M., S. P. Staal, D. P.Bolognesi,T. P. Denny,and
W. P. Rowe. 1977. Effect of interferon onmurineleukemia virus infection. II. Synthesis of viral components in exogenous infection. Virology 79:1-13.
36. Pitha, P. M., N. A. Wivel, B. F. Fernie, and H. P. Harper. 1979. Effect of interferon on murine leukemia virus infec-tion. IV. Formation of non-infectious virus in chronically infected cells. J. Gen. Virol. 42:467-480.
37. Reading, C. L., K. W. Brunson, M. Torrianni,and G. L. Nicholson. 1980. Malignancies of metastatic murine
lym-phosarcoma cell lines and clones correlate with a de-creased cell surface display of the RNA tumor virus envelope glycoprotein gp7O. Proc. Natl. Acad. Sci. U.S.A. 77:5943-5947.
38. Schroder, E. W., I. Chou, S. Jaken, and P. H. Black. 1978. Interferon inhibits the release of plasminogen activator fromSV3T3cells. Nature (London)276:828-829.
39. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981. Nucleotide sequence of Moloney murine leukemia virus. Nature (London) 293:543-548.
40. Stephenson, J. R. 1980. Type-C virus structural and trans-formation-specific proteins, p. 245-297. In J. R. Stephen-son (ed.), Molecular biology of the RNA tumor viruses. Academic Press, Inc., New York.
41. Taylor-Papadimitriou,J. 1980. Effects of interferon on cell growth and function, p. 13-46, InI.Gresser (ed.), Inter-feron 80, vol. 2. Academic Press, Inc., New York.
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 5.[35S]methionine-labeled](https://thumb-us.123doks.com/thumbv2/123dok_us/1457339.98433/6.504.51.243.67.247/fig-s-methionine-labeled.webp)