Copyright@1975 AmericanSocietyfor Microbiology Printed inU.SA.
Studies
on
Reverse Transcriptase
of
RNA Tumor Viruses
I.
Localization
of
Thermolabile
DNA
Polymerase and RNase H
Activities on One Polypeptide
INDER M. VERMA
TumorVirologyLaboratory, TheSalk Institute,SanDiego,California 92112
Received forpublication6September1974
Purified reverse transcriptase from avian myeloblastosis virus or Rous sarcoma virus consists of two subunits of average mol wt of 100,000 and 60,000. The
lower-molecular-weight subunit, a, has been isolated from avianmyeloblastosis virus, Roussarcomavirus, andatemperature-sensitivemutantofRous sarcoma virus, LA337. Subunit a manifests both the DNA polymerase and RNase H activities associated with purified reverse transcriptase of avian RNA tumor viruses. Thethermal inactivation of these enzymatic activities ofasubunit from LA337was comparedwith the isolated a subunit from thewild-typevirus.The
resultsshow thatbothDNApolymeraseand RNase H activities associated with the a subunit of LA337 are five to seven times more thermolabile than the
correspondingasubunit fromthewild-type virus. It isconcluded that (i) both thepolymeraseandnucleaseactivities resideonthe samepolypeptidechain, and
(ii)atleast the lower-molecular-weight subunit a is coded forby the viral RNA.
Avian RNA tumor viruses contain a DNA
polymerase whichcanfaithfully transcribe the viral RNA into complementary DNA (11, 12). The DNA polymerase can besolubilized from thevirions and purifiedbycolumn chromatog-raphy (3, 6, 8, 13). Purified DNA polymerase from avianmyeloblastosis virus (AMV)orRous sarcoma virus (RSV), uponanalysisonsodium dodecyl sulfate-containing polyacrylamide gels, dissociates intoequimolaramountsoftwo poly-peptide chains (subunits) with average mol wt of100,000 and 60,000 (4, 8, 11, 15). The
lower-molecular-weight subunit (a) hasbeen isolated
from AMV (4) and has been shown to contain
the threeknownenzymaticactivitiesassociated
with the DNA polymerase from avian RNA
tumor viruses: RNA-directed DNA synthesis,
DNA-directed DNA synthesis, and RNase H (which selectively degradesthe RNAmoiety of RNA-DNAhybrids)(11, 12). DNApolymerases
isolated from two temperature-sensitive mu-tants ofRSVwith defectsin a veryearly event
ofthe virusgrowth cycle (10) have been shown
to be morethermolabile than the DNA
polym-erase fromthewild-type parent (14). Since the
purified DNA polymerase studied in those
ex-periments contained bothsubunits, it could not be conclusively established that the
tempera-ture-sensitive lesion residesinthesubunit
con-taining the polymerase and nuclease activities. To demonstrate that the polypeptide chain
manifesting the synthetic and the degradative activities isthermolabile, the lower-molecular-weight subunit a has been isolated from the temperature-sensitive mutant LA337. The heat-inactivation properties of the DNA polym-eraseand RNase H activities ofasubunit from the mutant virus were compared with the a
subunitisolated from thewild-type parent. The results indicate that both the DNApolymerase
and the RNase H activities of a from LA337 are five to seven times morethermolabile than those oftheasubunit of thewild-typeparent.
MATERIALS AND METHODS
Materials. Tritium-labeled and unlabeled
deoxy-ribonucleoside triphosphates were obtained from
Schwarz/Mann and New England Nuclear.
Polynu-cleotides wereobtainedfromeither MilesLaboratory
or P-L Biochemicals. Oligonucleotides were 8 to 18
in length andwere obtained fromCollaborative
Re-search. AMV inplasmawasprovidedbythe Office of
ProgramResources andLogisticsofthe Virus Cancer
Program of the National Cancer Institute.
[3HJpoly(A)
wassynthesizedasdescribedpreviously(2). Escherichia coliDNApolymerasewas agiftfrom
A.KornbergtoD. Baltimore.
Purificationof virion DNApolymerase.Purified
virions of LA337and wild-type RSV, Prague strain
(subgroup C), wereobtainedas describedpreviously
(10). AMV was purified from chicken plasma as
described(13). About 4.5 mg ofpurifiedvirionsfrom
LA337and 14.0 mg ofpurified virions fromwild-type
RSVwerelysed with Nonidet P-40(ShellChemicals), 121
on November 10, 2019 by guest
http://jvi.asm.org/
andDNA polymerase was purified by column chro-matography on DEAE-Sephadex A-25 (13). En-zymatic assays were performed as described previ-ously (13). The peak fractions with enzymatic activi-ties weredialyzed overnight against buffer B
contain-ing 50 mM Tris-hydrochloride (pH 7.5), 0.1 mM
EDTA, 0.1 M ft-mercaptoethanol, 20% glycerol, and
0.15%Nonidet P-40. Thedialyzed enzyme was further
purified on a phosphocellulose column (13). AMV
DNA polymerase was purified as previously described
(13).
RNA-directed DNA synthesis was assayed using
poly(C)-oligo(dG) as template-primer; DNA-directed DNA synthesis was assayed usingpoly(dC) -oligo(dG) astemplate-primer. RNase H activities were assayed as described previously (2, 14). All enzyme assays were carried out for times during which synthesis or
degradationwasalinear function of time. The details
ofvarious reactions are described in thelegends to the
figures.
RESULTS
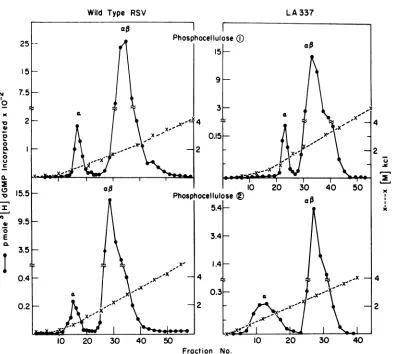
Isolation and characterization of a from AMV, wild-type RSV, and LA337. The lower-molecular-weight subunit a was isolated by
sequential chromatographyonphosphocellulose columns. Figure 1 shows the chromatographic profiles of DNA polymerases isolated from
LA337 and wild-type RSV. The major peak of
enzymaticactivityobtainedfrom the first
phos-phocellulose chromatography was dialyzed overnight against buffer B and rechromato-graphed on asecond phosphocellulose column. When assayed with poly(C) -oligo(dG) as
tem-plate-primer, two peaks of enzymatic activity
could be distinguished. The minor peak of
enzyme activity, a, elutingat asalt
concentra-tion of 0.1 M, represents 3 to 10% ofthe total
enzyme activity. The major peak of enzyme
activity, a,, elutes at a salt concentration of 0.24 M KCl. Similar separationof two enzyme
activities can be obtained from AMV. In all
subsequent experiments, a and af3 obtained from the second phosphocellulose columnwere
used.
The molecularsize of a and
a#
wasanalyzed by chromatography on Sephadex G-100col-umns. Figure 2represents a composite of three
sets ofexperiments and was drawn by aligning
the E. coli DNA polymerase I activity in all
three experiments. It can be seen that aC, from
AMV,which has anaverage mol wt of 150,000 to
170,000, elutes inthevoid volumeconsiderably
ahead of themarker,E. coli DNApolymeraseI,
which has a mol wt of 110,000 (7). This is
consistentwith the observation that the
holoen-zyme is composed of twosubunits, with atotal
molwtof170,000(6, 8,15).Subunitafromboth
AMV and wild-type RSV elutes from the
col-umnafterE. coli DNApolymeraseI.Analysis of
afrom AMV in sodiumdodecyl
sulfate-contain-ingagarose slab gels shows one major
polypep-tide band with an average mol wt of 60,000
(I. M. Verma and W. Gibson, unpublished
data). Thus, based upon Sephadex G-100
col-umn chromatography and polyacrylamide gel electrophoresis, afrom AMV orRSV wild-type
appearstoconsist ofonlyonepolypeptidechain
with a mol wt of 60,000 to 70,000. This is in
agreementwith the results ofGrandgenettetal.
(4), although their estimate of the molecular
weight of a of AMV is somewhat higher. The
molecular size ofa isolatedfromLA337was not
characterized because ofinsufficient material. It has, however, been shown previously that ad
obtained either from LA337or wild-type RSV
has the same molecular size and number of
subunits (14).
DNA polymerase and RNase H assay ofa
subunit of LA337 and wild-type RSV. The
three known enzymatic activities associated
with theDNA polymerase of avian RNAtumor
viruses were compared using poly(C) oligo(dG)
to measure RNA-directed DNA synthesis,
poly(dC) oligo(dG) toassayDNA-directed DNA
synthesisand [3H]poly(A) poly(dT) to assay for
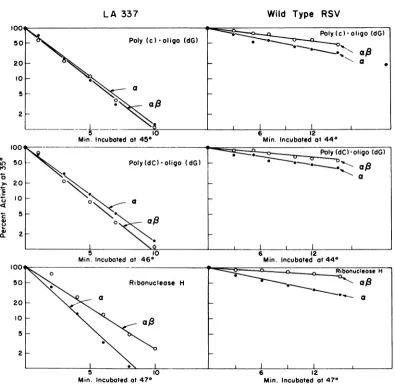
RNase H activity (Fig. 3). The average time of
inactivationofone-half of the activity of a from
LA337andwild-type RSV for these three
activi-ties has been calculated (Table 1). The a
subunit isolated from the virions of LA337
showed a five- to sevenfold greater
thermolabil-ity than the a subunit isolated from wild-type
RSV for all three activities.
Comparison ofa and a(3 from LA337 and wild-type RSV. Both the RNA-directed DNA
synthesis and DNA-directed DNA synthesis
activities associated with a subunit of LA337
are slightly more resistant tothermal
inactiva-tion than the activities of the corresponding
holoenzyme, af (Fig. 4). Subunit a from the
wild-type parent is, however, slightly more
thermolabile than the af3 complex. The RNase
H activity of a from both LA337 and wild-type
parent arereproduciblymore thermolabile than
that of the two-subunit complex.
DISCUSSION
DNA polymerase isolated from two
condi-tional temperature-sensitive mutants of RSV
(LA335 and LA337) has been shown to be
three-to fivefold more thermolabile than the DNA
polymerase from thewild-type parent (14). The
thermal inactivationprofileofthe DNA
on November 10, 2019 by guest
http://jvi.asm.org/
Wild Type RSV
123
LA337
x
xI
Fraction No.
FIG. 1. Sequential column chromatography ofDNApolymerase from wild-type RSVandLA337. Themajor peak of enzymatic activity from the first phosphocellulosecolumnwaspooled, dialyzed overnight against buffer
B, andrechromatographedon asecondphosphocellulosecolumn. Twopeaks of enzymatic activities,aanda#,
could be distinguished. Peak awasstored in 50%glycerolat -20C,whereasa# wasstoredat -70C. The
recoveryof the enzymatic activity from the second phosphocellulosecolumnwasover50%.Enzymeassayswere
performed utilizing poly(C) -oligo(dG) as template-primer and 3[H]dGTP (100 counts per min/pmol) as
substrate. Peaksaand a#from LA337 virionswere isolated inan identicalfashion, exceptthedialysiswas
carried out for 5 to 8 h instead of overnight to minimize thermal inactivation ofthe enzyme during the
purification procedure. Therecoveryof totalenzymeactivity after first phosphocellulose columnwas over20%.
On the secondphosphocellulose column over 75% ofthe input enzymatic activity was recovered. Peak a
constitutedabout 10% of the totalenzymeactivity. Bothaandaftobtainedfrom LA337 virionswerestoredat
-70 C.
eraseactivity from the wild-typerevertants(wt
LA335 andwtLA337) is indistinguishable from
that of wild-type virus (14). Mason et al.
(Virology, in press) have recently characterized
the biological and biochemical properties of
these mutants, their revertants, and several
recombinants with leukosis viruses and have
shown that the lesion in these mutants is the
increasedthermolability of theirDNA
polymer-aseactivity. Varmusetal. (Cold SpringHarbor
Symp. Quant. Biol., in press) have reported
that less viral DNA is produced in LA335 and
LA337at the nonpermissive temperature than
isproduced by thewild-type virus.
Sinceall three knownenzymeactivities
asso-ciated with the RNAtumorvirusDNA
polym-erase werethermolabile, it was suggested that
all three activities were located on the same
polypeptide chain (14). To establish that the
polypeptide chainmanifesting all threeenzyme
CY
N
0
x
10 0 0
0. CL
0
'IO
toE
IL
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.51.453.66.420.2]VoLw volume UNAP'olymerose1
a) I ~~~~~~~~~~0
o
10
40-20 25 Q.
p h coln3lofAMVa aofrA M
Peakafw-RaRSV
20tie 0m ri-yrclrd (p 7.) 02M
polmeas 0aIde sasanadmre.Dx
-~0/
E
xo -~~~
O-x~~ ~ ~~~~-~~>
10 15 20 25 30
Effluent m2
FIG. 2. Size determinationofaandaffromsecond
phosphocellulosecolumn. Peaksaanda#fromAMV
were obtained as described in the legend to Fig. 1.
Peakafrom wild-typeRSV andaand az fromAMV were separately chromatographed on a Sephadex G-100 column (1 by 70 cm). The elution buffer
contained50mMTris-hydrochloride (pH 7.9),0.2M
KCI,0.1 mMEDTA, 10 mM mercaptoethanol, and
0.1% Nonidet P-40. In each case, E. coli DNA
polymeraseIwas added as astandard marker.
Dex-tran blue was used to determine the void volume. Peaks a and a# were assayed for enzyme activity
usingpoly(C) oligo(dG)astemplate-primer(15). The
Poly(c)-oligo(dG)
activities isthermolabile and isencoded by the viralRNA, I isolatedasubunit from LA337and
compared its thermal inactivation properties with isolated a subunit from wild-type RSV. The a subunitwasisolated from purified DNA polymerase which had bothsubunitsin equimo-lar ratio. This approach assures that the iso-lated lower-molecular-weight subunit a is part
oftheviral DNA polymerase. The mechanism
ofseparation of a from
a#
(two subunitcom-plex) is not clear. A comparison ofthe tryptic peptides of isolatedaand,8(large subunit)from
either AMV or wild-type RSV reveals thatthe two subunits share at least a portion oftheir amino acid sequences (Gibson and Verma,
un-published data). Thus, based upon the
struc-tural relatednessofthetwo subunits and poly-acrylamide gel electrophoretic profile of aged
reverse transcriptase from AMV (K. Molling,
Cold Spring Harbor Symp. Quant. Biol., in
E. coli DNA polymerase was assayed utilizing poly-(dA)-oligo(dT) as template-primer and 8[H]TTP as substrate (15). The figurewasdrawn by aligning the E. coli DNA polymerase activities of the three
sepa-rateexperiments. The recovery of enzyme activity of afifrom the column was over90%1, whereas a and E. coli DNA polymerase showed a recovery of over 30% of
theinputenzymeactivity.
PolytdC) oligo (dG) Ribonuclease H
Min. Incubotedat450 Min. Incubotedat460
5
Min.Incubatedot470
FIG. 3. Thermalinactivationprofiles ofDNApolymeraseand RNase H activitiesofisolatedasubunitfrom
LA337andwild-typeRSV.Assaysofheatlability wereperformed asdescribed (15).Briefly, the enzymewas
preincubated in reaction mixture 1, which contained 100 mM Tris-hydrochloride (pH 8.3), 20 mM
dithiothreitol, 12 mM Mg acetate, and 120 mMNaCl. The reaction mixture was incubated at the high
temperature showninthefigure,andatvariousintervalsportions (0.05ml)werewithdrawn and addedto0.05 mlof reaction mixture 2, which containedtemplate-primerand thesubstrates. This mixturewaskeptonicefor amaximum of10to 15 min, thenfurther incubatedat35Cfor30min.Acidprecipitable radioactivitywas
determinedasdescribedpreviously (1). (a)Approximately0.005to0.05unitsofpurifiedasubunitwasused per assay. (A unit of enzyme activity is defined as theamount ofenzyme required to incorporate 100pmol of
s[H]dGMPat37C in15min.) Onemicrogramofpoly(C)and0.5jigofoligo(dG)wereusedastemplate-primer,
and10nmolofs[H]dGTP (1,000countspermin/pmol)wasusedassubstrate.The enzymewaspreincubatedat 45C. (b) The samereaction conditions as described above were employed, except 1.0 gg of poly(dC) was substitutedforpoly(C),and thepreincubationwascarriedout at46C.(c)RNaseHassayswereperformedas
previously described [Vermaetal.,Nature(London),inpress].About1 to2unitsofenzymeactivitywasused,
and thepreincubationswerecarriedout at47C.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.68.255.59.248.2]TAMZ 1. Average one-half time for inactivation of DNA polymerase and RNase H activity of isolated a
from LA337 and wild-type RSV
t%,(min)"
RNA-
DNA-Source directed directed RNase H
Source DNA DNA R
e[H]
synthesis synthesis poly(A)-poly(C)* poly(dC) poly(dT) oligo(dG) oligo(dG)
LA337 1.8 2.1 1.5
Wild-type RSV 8.8 9.5 8.0
a
t%
isthe time required for inactivation of50%ofactivity at the temperatures indicated in Fig. 3.
LA 337
0
If)
re)
0 >-.
Z
.1-C c
press), it appears that a is derived from
f.
Experiments performed on the mechanism of
RNase H (5) and heat stability of isolated a subunit in the presence or absence of template (A. Panet, I. M. Verma, and D. Baltimore,
Cold Spring Harbor Symp. Quant. Biol., in press) suggest that , subunit enhances the affinity ofaforbindingtothetemplate.Weare
now investigating whether 6 is enzymatically activeand how a isgenerated from,.
ACKNOWLEDGMENT1S
I am verygratefulto W.S. Masonforgenerously providing me with mutant andwild-typevirus. I thank D.Baltimorefor hissustainedinterest inthisworkand helpinthepreparation
Wild Type RSV
Min. Incubated at470 Min. Incuboted at470
FIG. 4. Comparison of thermal inactivation profiles of DNA polymerase and RNase H activities ofaanda#, fromLA337 andwild-typeRSV. Assays for heatlabilitywereperformedasdescribed. About0.5 to 1.0unitsof
a,@wasused per assay. Preincubationswerecarried outattemperaturesshown in thefigure.
125
VOL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.68.463.237.624.2]of this manuscript. I am grateful to Amos Panet for many valuable discussions.
Thiswork wascarried out in thelaboratoryofD. Baltimore at the Department of Biology and Center for Cancer Re-search, Massachusetts Institute ofTechnology, Cambridge, Mass. It was supportedbyacontract fromthe VirusCancer Program, Public HealthService grant lP01-CA-14051 from the National Cancer Institute to D.Baltimore, PublicHealth Service grant CA-16561-01 fromthe National Cancer Insti-tuteandaresearchgrant, 320, fromthe Jane Coffin Childs Memorial Fund to I.M.V.
LITERATURE CITED
1. Baltimore, D., A. S. Huang, and M. Stampfer. 1970. Ribonucleic acidsynthesisofvesicular stomatitis virus. II. AnRNApolymerase in the virion. Proc. Nat. Acad. Sci. U.S.A.66:572-576.
2. Baltimore, D., and D. Smoler. 1972. Association of an endoribonuclease with the avian myeloblastosisvirus DNApolymerase. J. Biol.Chem.247:7282-7287. 3. Faras, A. J., J. M. Taylor, J. P. McDonald, W. E.
Levinson, and J. M. Bishop. 1972. Purification and characterization of thedeoxyribonucleicacid polymer-aseassociatedwith Roussarcomavirus.Biochemistry
11:2334-2342.
4. Grandgenett,D.P.,G. F.Gerard,and M.Green.1973. A single subunit from avian myeloblastosis virus with both RNA-directed DNApolymerase activityand ribo-nuclease H activity. Proc. Nat. Acad. Sci. U.S.A. 70:230-234.
5. Grandgenett,D.P., and M. Green.1974.Different mode of action of ribonuclease H in purified a and a,B ribonucleicacid-directeddeoxyribonucleicacid polym-erasefrom avian myeloblastosisvirus. J. Biol. Chem. 249:5148-5152.
6. Hurwitz, J., and J. P. Leis. 1972.RNA-dependentDNA polymerase activity of RNA tumor viruses. I.Directing
influence of DNA in the reaction. J. Virol.9:116-129. 7. Jovin, T. M., P. T. Englund, and L. L.Bertsch. 1969. Enzymatic synthesis ofdeoxyribonucleic acid. XXVI. Physical andchemical studies of a homogeneous deoxy-ribonucleic acid DNA polymerase. J. Biol. Chem. 244:2996-3008.
8. Kacian,D. L., K. F. Watson, A. Burny, and S. Spiegel-man. 1971. Purification of DNApolymerase of avian myeloblastosis virus. Biochim. Biophys. Acta 246:365-383.
9. Leis,J.,I.Berkower,and J.Hurwitz. 1973. RNA-depend-entDNApolymerase activity in RNA tumor viruses, p. 287-308.In R. D. Wells and R. B. Inman(ed.), DNA synthesis in vitro. University Park Press, Baltimore. 10. Linial, M., and W.S.Mason. 1973.Characterizationof
twoconditional earlymutantsofRoussarcoma virus. Virology 53:258-273.
11. Temin, H.M., and D. Baltimore. 1972. RNA directed DNAsynthesis of RNA tumor viruses, p. 129-186. In K. M.Smith and M. A. Lauffer (ed.), Advances in virus research, vol. 17. Academic Press Inc., New York. 12. Tooze, J. (ed.). 1973. The molecular biology of tumor
viruses. Chapter 11. ColdSpringHarborLaboratory, New York.
13. Verma, I. M., and D. Baltimore. 1973. Purification of the RNA-directed DNApolymerase from avian myeloblas-tosis virusand its assay withpolynucleotidetemplates, p. 125-131. In L. Grossman and K. Moldave (ed.), Methods in enzymology, vol. 29. Academic Press Inc., NewYork.
14. Verma, I. M., W. S. Mason, S. D. Drost, and D. Baltimore. 1974. DNA polymerase activity from two temperature-sensitive mutants ofRous sarcoma virus isthermolabile. Nature (London) 251:27-31. 15. Verma, I. M., N. L. Meuth, H. Fan, and D. Baltimore.
1974. Hamster leukemia virus: lack of endogenous DNA synthesis and unique structure of its DNA polymerase. J. Virol. 13:1075-1082.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 3.preincubatedLA337previouslymldithiothreitol,assay.45aandsubstitutedtemperaturedetermineds[H]dGMPand maximum C](https://thumb-us.123doks.com/thumbv2/123dok_us/1577199.110313/4.504.68.255.59.248/fig-preincubatedla-previouslymldithiothreitol-assay-aandsubstitutedtemperaturedetermineds-h-dgmpand-maximum.webp)