0022-538X/91/084033-09$02.00/0
Copyright© 1991, AmericanSocietyfor Microbiology
ZEBRA
and
a
Fos-GCN4
Chimeric Protein
DNA-Binding
Specificities for Sites in the
Virus
BZLF1 Promoter
Differ in Their
Epstein-Barr
NAOMITAYLOR,' ERIK FLEMINGTON,2 JOHN L. KOLMAN,1RAYMOND P. BAUMANN,' SAMUEL H. SPECK,2 ANDGEORGE
MILLER'3
4*Departments of Molecular Biophysics and Biochemistry,'Epidemiology and Public Health,3 andPediatrics,4 Yale University School of Medicine, New Haven, Connecticut 06510, andDivision of Tumor Virology,
Dana-Farber CancerInstitute, andDepartmentof Pathology, Harvard Medical School, Boston, Massachusetts 021152
Received12October1990/Accepted16April 1991
Epstein-Barr virus (EBV) encodesa protein, ZEBRA, which enables the virustoswitchfrom alatenttoa
lytic lifecycle. Thebasicdomain of ZEBRA is homologoustotheFos/Junoncogenefamily, and both proteins
bindthe canonical AP-1site(TGAGTCA). However, ZEBRAdoesnotcontainaleucine zipper dimerization domain which has beenshown tobe necessaryforDNAbinding of Fos/Jun proteins. Additionally, ZEBRA bindstosites which deviatefromthe AP-1consensus sequence.Thus,itwasofinteresttodefinethedomain of theZEBRAprotein required forDNAbinding.We have determined bymutagenesisthatZEBRA residues 172 to227,representing the basic domainandaputative dimerization domain,arerequiredfor specific bindingto AP-1 and divergent sites. Mutagenesisofthe basic aminoacids 178 to 180 or187 to 189 abrogates ZEBRA binding to all DNA target sequences. These residues are conserved in Fos and arealso necessary for Fos DNA-binding activity.Wehave foundthataFos-GCN4 chimera and ZEBRA have differentcognatebinding specificities. The autoregulated BZLF1 promoter contains three divergent AP-1 sequences, ZIHA (TGAGCCA),ZIIIB(TTAGCAA),andZ-AP-1-octamer(TGACATCA).ZEBRAbindswithhigh specificityto ZIIIAand ZUIBbutweaklytothe Z-AP-1octamer. Conversely, theFos-GCN4 chimera recognizesonly the Z-AP-1 octamer. ZEBRA binds the ZIIIA and ZIIIB sites together in a noncooperative fashion, while Fos-GCN4 bindsthesesitesas a higher-ordercomplex. Additionally, we havefound thatflankingsequences influencebinding of Fos-GCN4toadegenerate AP-1site(TGAGCAA). The characteristic binding specificities of ZEBRA andcellular AP-1 proteinssuggest thatthey differentiallyaffect viraland cellular transcription. Epstein-Barr virus (EBV) encodes a protein, ZEBRA,
which enablesthe virustoswitch fromalatenttoalytic life cycle (8, 9, 41). EBV is latent in lymphoid cells butcan be activated invitroby phorbolesters, butyrate,orexpression of the ZEBRAgene(8, 9, 15, 29, 41, 46). The mechanism by which ZEBRA initiates the cascade of viral replication has been thefocus of much study. Numerousgroupshavefound that ZEBRA directly transactivates several promoters of lytic cyclegenes, includingaregion integraltotheEBVlytic originofreplication (7, 8, 13, 15-17, 20, 28, 41, 43).
Exon II of ZEBRA ishomologoustothe basic domain of the Fos/Junproto-oncogene protein family (11). In Fos/Jun complexes, thisdomain has been showntointeractdirectly witha DNA heptamer known asthe tetradecanoyl phorbol
acetate (TPA) response element or AP-1 site (TGAGTCA) (3, 26, 34). Itwasdetermined that ZEBRA shares theability of theFos/JuncomplextobindanAP-1consensussite(11). AnAP-1 sequence (MS-AP-1) is locatedin thepromoterof anEBVearlygene, BSLF2-BMLF1(MS-EA) (11). ZEBRA can transactivatetranscription throughthis site, although it is not yet clear whether AP-1 sequences are required for transactivation ofEBV genes in infected lymphocytes (18, 43).
ZEBRA also binds sites on the EBV genome which deviate from the AP-1 consensus sequence (11, 13, 27, 28,
37, 43). TheautoregulatedBZLF1 (ZEBRA)promoter
con-*Corresponding author.
tains three divergent AP-1 elements (13). Two sites, TGAGCCA (ZIIIA)andTTAGCAA(ZIIIB), 129 and 116bp upstream of the transcriptional start site are arranged in a tail-to-tail fashion andare separated by six nucleotides. An additional AP-1 sequence, TGACATCA (Z-AP-1 octamer),
islocatedfurther downstreamat-67.Thissequence,which wasfirst identified in thejun promoterby Angelet al. (2), mediates activationbyTPA andJun/AP-1. Recent work has shown thatalthoughthissite is TPAinducible in the ZEBRA promoter,it doesnotappeartobetransactivatedbyZEBRA (12, 13).
Sinceboth ZEBRA and Fos/Junareinvolved in transcrip-tional regulation of gene expression, it was important to
compare the DNA-binding activities of these proteins. Therefore,westudiedthe relativeDNA-bindingaffinitiesof ZEBRA and a Fos-GCN4 chimera for the divergent AP-1 sitesfound in the autoregulatedBZLF1 promoter.
MATERIALS ANDMETHODS
Plasmids. Thefull-length BZLF1cDNA(aminoacids[aa] 1to245) (30)wasclonedas aNaeI-BamHIfragmentintothe SmaI-BamHI sites of the trpE bacterial expression vector
pATH 11. TheRajiBZLF1 cDNA andtheexpressionvector
were the gifts of A. Sergeant and T. J. Koerner,
respec-tively. BZLF1 deletionmutantswereconstructedasfollows:
Z141-245
wasclonedasaNheI-BamHIfragment,Z172-245wascloned as aBsmI-BamHI fragment, Zl-198 wascloned as a NaeI-PstI fragment, and
Zl-227
was cloned as aBamHI-4033
on November 10, 2019 by guest
http://jvi.asm.org/
4034 TAYLOR ET AL.
HinclI
fragment.
Allfragments
were cloned into theappro-priate
sites ofpATH vectors.ZA111-159
was constructedby
bidirectional Bal 31 exonuclease
digestion
ofZl-245
attheunique
NheI site(aa 141). Exon Iofthe BZLF1 openreading
frame, WZ1_167,
was cloned into thepATH
vector fromWZhet (42), and exon II of
BZLF1, Z167-202,
was clonedfrom
genomic
BamHI Zas aPvuII-HincIIfragment.
TwoadditionalmutantZEBRA
proteins
wereconstructedby
site-directedmutagenesis
of the B95-8 BZLF1 cDNA cloned in SP64. This cDNA was thegift
ofP. Farrell(11).
Amino acids 178to 180 and 187 to 189 were mutated from
KRY to EELand RKCto EES,
respectively.
Site-directedmutagenesis
wasperformed
as describedby
themanufac-turer
(Bio-Rad).
AFos-GCN4 chimera which containsthe Fosbasic
DNA-binding domain,
aa 126to 162, fused inframetothe GCN4leucine
zipper domain,
aa251to281,was thekindgift
ofT.Kouzarides and E. Ziff(24). A Sau3AI-XbaI fragment
con-taining
theFos-GCN4chimerawasclonedinto theBamHI-XbaI sites ofpATH1.
Protein expression. Fusion
proteins
were expressed as describedpreviously
(22, 42). Escherichia coliAG1contain-ing
thevariousTrpE-BZLF1deletionmutants wasgrownin 5 ml ofM9 mediumcontaining
1% Casamino Acids(Difco)
andinduced for4hin the presenceof
indoleacrylic
acid(20,ug/ml).
Proteins from0.5 ml of cells wereelectrophoresed
on a sodium
dodecyl
sulfate(SDS)-polyacrylamide gel
and stained with Coomassiebrilliant blue. The level of inducedprotein
in eachpreparation
was estimatedvisually,
andvolumes ofcells containing equivalent amounts ofprotein
(3.5
to 5ml)
werepelleted
andresuspended
in 1 ml of6 Murea
(1).
Cellsweresonicatedtwice for20seachtime. Theurea-solubilized
protein
fractionwasdialyzed
threetimes for45mineachtimeat4°C
against
15 mMN-2-hydroxyethylpi-perazine-N'-2-ethanesulfonic
acid(HEPES; pH 7.9)-75
mMKCl-0.2 mM EDTA-1 mM dithiothreitol-7.5%
glycerol.
Constructs in
pBluescript
KS were transcribed in vitro(31)
and thentranslatedinawheat germextract asdescribedby
the manufacturer(Promega).
Synthetic oligonucleotides.
Thesynthetic
double-strandedoligonucleotides
used inbinding
studies were as follows(sequence
reflects top strand in a 5'-to-3'direction;
coresequences are underlined and mutated bases are noted in
bold
type).
MS-AP-1: TCTTCATGAGTCAGTGCTTC MS-AP-1: TCTTCATTAGTCAGTGCTTC ZIIIA: CTAGCTATGCATGAGCCACAGATC
ZIIIA*: CTAGCTATGCATGAGCAACAGATC
ZIIIAm: CTAGCTATGCAGAATTCACAGATC
ZIIIB: CTAGCAGGCAITGCTAATGTACCGATC
ZIIIB*: CTAGCAGGCATTGCTCATGTACCGATC
ZIIIBm: CTAGCAGGATCCGCTAATGTACCGATC
Double:
GACTATGCATGAGCCACAGGCADGCJAATGTACCGA
(containsZIIIA +ZIIIB)Z-AP-1octamer:CTAGAAACCATGACATCACAGAGGATC Controloligonucleotide: TGGCATGCTGCTGACATCTGGC
All
oligonucleotides
had XbaI sites at the 5' end andBamHI sites at the 3'
end,
with the exceptionof the AP-1 and Doubleoligonucleotides,
which had CT and AGover-hangs
at the 5' and 3' ends, respectively. Radiolabeledoligonucleotides
were prepared by using the Klenowfrag-ment ofE. coliDNApolymerase and 32P-labeled
deoxynu-cleotides.
Gel retardationassays. DNA
binding
reactions wereper-formedasdescribed
previously
(11).Reaction mixtureswere incubated atroomtemperature(22°C)or4°C for30min.A 4 x10-13
M concentrationof the appropriate32P-end-labeled
double-strandedoligonucleotideand 2 ,ul ofprotein
(approx-imately 100ng)were incubated ina25-,ul volume of 15 mM HEPES(pH
7.9)-75
mMKCl-0.2 mM EDTA-4 mM dithio-threitol-7.5% glycerol-1 pigofpoly(dI-dC). Gel retardationcompetitionassayswereconducted with5-, 20-,and 50-fold molar excesses of cold competitor over the 32P-labeled MS-AP-1 probe. The Double oligonucleotide was used in
2.5-, 10-, and 25-fold molarexcesses since it contains two
potential binding sites (ZIIIA and ZIIIB). Samples were
electrophoresedthrough 4%polyacrylamide-0.5x TBEgels at room temperature or 4°C. Gels were then dried and
autoradiographed. Films fromtwooligonucleotide competi-tionexperiments were quantified by densitometer scanning on a Kodak
Biolmage
densitometer with Visage 2000 soft-ware. Disruption of complex formationwas determinedby comparing the ratios of shifted complex in lanes with and withoutcompetitor.RESULTS
Determination of the ZEBRA DNA-binding domain by deletionalmutagenesis. A BZLF1 cDNA obtained fromRaji
cells was cloned in frame with the trpE coding sequences contained inthepATH 11
expression
vector(Fig.
1B). The 75-kDafusionprotein
bounda20-bp oligonucleotide
which encompasses an AP-1 consensus sequence (MS-AP-1) de-rived from the promoter of the EBV early gene, MS-EA(Fig.
1CandD,
Z1-245).
Severaldeletionmutantsdefined thedomain of the protein which bound the AP-1 site (Fig. 1C and D). All of the mutants exhibited some temperature-sensitiveDNA-binding activity.Mutant
Z141-245
encoded thesmallest
protein,
which behaved like thewild-type
ZEBRA(Zl-245)
in bindingDNAatroomtemperatureand 4°C (Fig. lA).However,eventhismutantdidnotbindDNAaswellat4°C
as did thewild-type
Zl-245-
Theregion
of ZEBRA encoded by exon II, which shares homology with the Fos basic domain (11), was insufficient to elicit DNA-bindingactivity (Fig.
1CandD,Z168202).
Although
ZA111-159
boundDNA
unstably
(smearinFig. 1C)
at roomtemperature(22°C)and
Z172-245
didnotbindDNAunder theseconditions(Fig.
1C), both mutant proteins bound DNA at 4°C (Fig. 1D). Deletion ofthe18carboxyl amino acids ofZEBRAresulted in a mutant
(Zl-227)
which bound the AP-1 site at room temperaturebutnotat4°C(Fig.
1CandD).Furthermore,allofthe ZEBRAdeletionmutantsshowed thesamepatternof
binding
tothedivergent
AP-1sites,
ZIIIAandZIIIB,presentin the BZLF1 promoter (data not shown). Thus, ZEBRA amino acids 172 to 227 were
required
for DNAbinding,
althoughneighboringsequences mayhaveinfluenced
protein
folding
orstability
under sometemperatureconditions.Site-directedmutagenesisofthe ZEBRAbasicdomain. The
region
ofZEBRAencodedby
exonII ofBZLF1 (aa 168to202)ispartially homologoustothebasicregionof Fos(Fig. 2A).It hasrecentlybeen shown thataa144to146and 154to 156 are required for the DNA-binding ability of Fos (35).
These basic amino acids are conserved at
positions
178 to 180 and 188 to 190 of ZEBRA. Therefore, we determinedwhether thesamebasicamino acidsinthis
region
contributeto the DNAbinding of ZEBRA. These residues were mu-tatedfrom Lys-Arg-Tyrto Glu-Glu-Leu and from
Arg-Lys-Cys
toGlu-Glu-Ser, respectively (Fig.
2A).Thesemutationsabrogated
DNAbinding
totheconsensus(MS-AP-1)aswellas
divergent
AP-1sites(ZIIIAandZIIIB)(Fig.
2B).Equiv-alentamountsof the in vitro-translatedwild-typeandmutant
proteins
wereusedin eachreaction(datanotshown). Theseresults demonstrate that the samebasic residues in ZEBRA J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ZF RBRA
A CDNA
pATI' dcrivativcs
zI,414.
~i1J1-154
A:
1I- 4
C
4~~~~~
I~~~~~
+(4C)
;I. ,
II
22CIncubations
D
is-Z7..
f (22C)
B 7
r X -Z
Kd
4CIncubations
84~ ~ ~ 4 "O.
~~~w'*
....~~ ~ ~ ~ ~ ~ ~ ~ ..
and Fos are required for binding to AP-1 and related se-quences. However, deletional mutagenesis showed that
thesebasic residuesarenecessarybutnotsufficient for DNA binding (Fig. 1).
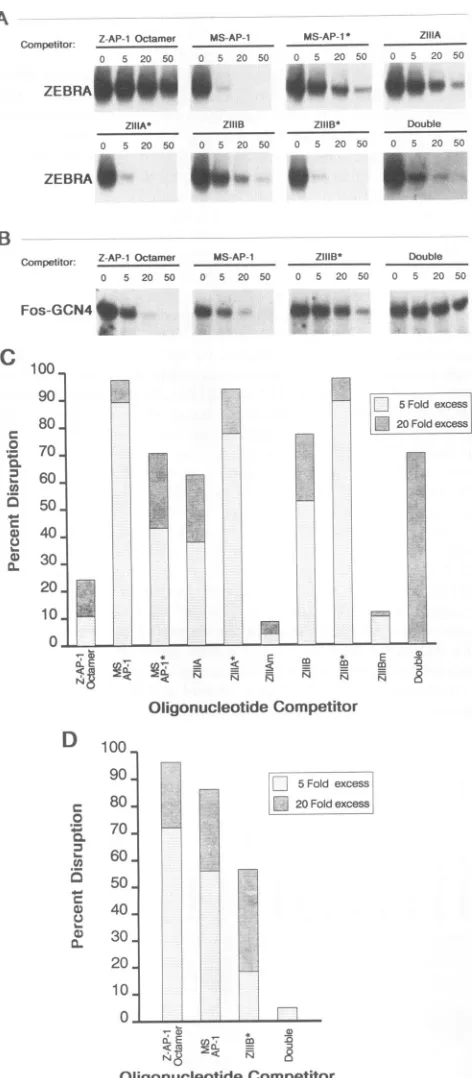
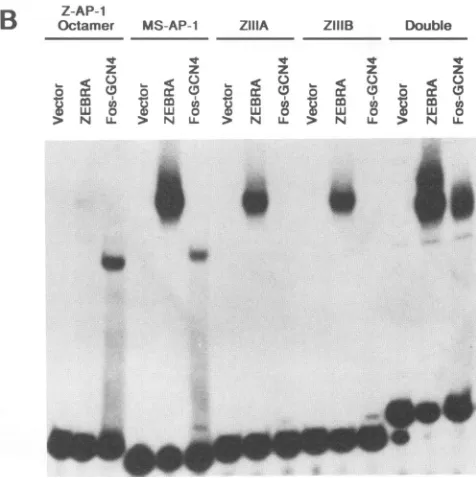
Binding ofZEBRAandFos-GCN4tooligonucleotides bear-ing sbear-ingle AP-1 and AP-1-like sequences. It was previously shown that ZEBRA footprinted sites ZIIIA at -129 (TGAGCCA) and ZIIIB at -116 (TTAGCAA) but not the Z-AP-1 octamersequence (TGACATCA) 67 bpupstreamof the BZLF1 transcriptional start site (13; Fig. 3A). A gel retardation assay wasusedtocomparetherelative affinities of ZEBRA and Fos-GCN4 for these sites in the BZLF1 promoter. TheFos basic domain has been showntobind the AP-1 consensus site as a homodimer when fused to the GCN4 leucine zipper (18, 24, 40). A Fos-GCN4 chimera, expressed as a TrpE fusion protein, bound the MS-AP-1 consensussite butnotZIIIBorZIIIA. ZIIIBdiffers from the consensusAP-1 siteat3bpandZIIIAcontainsasinglebase pair change from the consensus heptanucleotide AP-1 se-quence (Fig. 3B). Fos-GCN4 bound the Z-AP-1 octamer with high affinity, whereas ZEBRA interacted only weakly with this sequence. Thus, Fos-GCN4 binds the AP-1
oc-tamer and heptanucleotide sequences while ZEBRA binds the MS-AP-1, ZIIIA, and ZIIIB sites with relatively high affinity (Fig. 3B). The TrpE-ZEBRA and TrpE-Fos-GCN4 fusionproteins werederived from theurea-solubilized frac-tion oftotal E. coliproteinextracts.ZEBRAandFos-GCN4, expressed inthis system,bindDNAashomodimers (23). A control extractof bacterial protein did not exhibit specific bindingto anyofthese sites(datanotshown).
FIG. 1. DNA-binding activities of ZEBRAmutants. (A)
Struc-turesof ZEBRA deletionmutants. The ZEBRAcDNA is shownat
the top (30), and clones are designated accordingto the ZEBRA amino acidspresentintheconstruct.
Z,,111-159
isnamedaccordingtothe amino acids that have been deleted. Zl-167 was derivedfrom
genomic DNA (42). Translated sequences that are derived from
ZEBRA intron sequences are indicated by a hatched bar, and
sequencesfrom theexpressionplasmid polylinkerareindicatedbya
stippled bar. (B) Expression of TrpE-ZEBRA fusion proteins in bacteria. Total protein from bacteria that carried various TrpE-ZEBRA deletionproteinswasresolvedby SDS-polyacrylamide gel electrophoresis and stained with Coomassie brilliant blue. The
fusionprotein is indicated with dots in each lane. (C) Gel retardation
assayshowing binding ofTrpE-ZEBRA deletionmutants toa20-bp
32P-labeled oligonucleotide (4 x 10-' M) containing the AP-1
consensus sequence (11). Extract from bacteria transformed with
the trpE plasmid pATH 11 wasused as anegative control (vector lane). The various proteinswereeach incubated with the
oligonu-cleotideprobe and 1 p.gofpoly(dI-dC) in binding buffer for 30 min priortoelectrophoresis. Binding reactions and electrophoresiswere
performed at room temperature (22°C). Complexes were
electro-phoresedon a4%acrylamide gel in 0.5x TBE. (D) Gel retardation
assayof thesameTrpE-ZEBRA deletionmutantsperformedasfor panel C except that binding reactions and electrophoresis were carriedout at4°C.
Binding of ZEBRA and Fos-GCN4 to an oligonucleotide containing twoZEBRAresponseelements, ZIIIAand ZIIIB. Since the ZIIIA and ZIIIB sites are separated by only six nucleotides in the ZEBRA promoter, westudied the ability of ZEBRA and Fos-GCN4tobind both sitessimultaneously. AnoligonucleotidetermedDouble,whichincludes both sites intheorientation found in the BZLF1promoter,wasusedin gelband shiftassays. ZEBRAwasabletobindtwo siteson thisoligonucleotide simultaneously, asdemonstrated bythe
appearance of two band shift products in a mobility shift
assay(Fig. 3B). Surprisingly,the Fos-GCN4chimera,which didnotbindasingle ZIIIAorZIIIBsite,wasable to bindthe
Double oligonucleotide (ZIIIA plus ZIIIB), generating a moreslowly migrating complex (Fig. 3B).
C1 1 I
_1 - . - = -
-> N N N N N N NS N
Om S
< -C < ig
1- <z
zS
Cm
I
z Fl.-.,4,.
7 -4.
I
;I
" . =7
;.I
I .
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.57.291.79.407.2]4036 TAYLOR ET AL.
A
132-LSPEEEEKRRIRRERNKMAAAXCRNRRRELTDTLQA-167
i1l
II
I1: II167-SLEECDSELEIKRYKNRVASRKCRAKFKQLLQHYREV-203
---EEL
---EES
---B Z:A
,-% a',
OD r1.
Ns N rsJ
c-FOs
A
ZEBRA Z-M178-80 Z-M187-89
2 1,E tvIS At^
c~W. c c: OD K- OD cc
Ns N N .N rs N
ZEBRA (BZLFI) Promoter
ZIIIA ZIIIB
-140 GAAAc'l'A'I'GCATGAGCCACAGGCA'rl1'O;C'T'AATGTACCTCATAGACACACC
CTTTrGAC'TACG'T'ACT' CGGr G'G CCCG AACGATT.ACATIGGAGTATCTGTGTGG
Z-AP-1 OCTAMER
-90 T'AAA'I"I"I'A0CACG'G'CCCAAACCATGACATCACAGAGGAGGCTGGTGCCTT
A'ITTr I'AAATECGTlGCAGGGrGrT'I"ICGGTACTrGTrAGTGTCTCCTCCGACCACGGAA
-40 GGCTITT'AAAG0GGGAG0ATGTTCAGACAGGTAACTCACTAAACATTGCACCTT CCGAAA'I"IT'CCCCI'CTACAATCTGTCCATTGAGTGATTTGTAACGTGGAA
MS-EA (BSLF2-BMLF1) Promoter
ACGGTCACCTTCATGAGTc GTGcrTCGCCGGTCGCTGTGGGGCCAATCA
BZ_Z_P-1
[image:4.612.276.566.76.474.2]B Octamer MS-AP-1 ZIIIA ZIIIB Double
FIG. 2. Effects of point mutations in the basic domain ofZEBRA.
(A) Amino acid sequencesof human c-FosexonIII(aa 132to167) (44) and BZLF1exonII(aa 168to202) (5). The conserved residues
in this highly basic domain are indicated by lines (11). ZEBRA
mutantswereobtainedby site-directed mutagenesisofthe BZLF1 cDNAcloned inSP64. Amino acids 178to180 and 187to189were
changedasshown for Z-M178-80andZ-M187-89,respectively. (B) Gel retardation assay demonstrating theinability ofZEBRA con-structs mutated at aa 178 to 180 (Z-M178-80) or 187 to 189 (Z-M187-89) to bind MS-AP-1, ZIIIA, or ZIIIB. The wild-type (Z-WT) and mutated ZEBRAconstructs(Z-M178-180 and Z-M187-189)weretranscribed in vitrowithSP6 polymeraseandtranslatedin awheatgermextract.Equivalentamountsof theresultingtranslated
proteinswere each incubated withthefollowing 32P-labeled
oligo-nucleotide probes: MS-AP-1,ZIIIA, and ZIIIB(seeMaterials and
Methods for sequences). These complexes were then resolvedby gel electrophoresison a4%acrylamide gelin0.5x TBE.
Wenextstudied the order ofbindingtositesonthe Double oligonucleotide. Serial dilutions of ZEBRA and Fos-GCN4 wereincubated with 4x 10-13M 32P-labeledDoubleprobe. Atlowproteinconcentrations(0.25 pl),ZEBRAboundonly a single site on the oligonucleotide (one shifted complex),
while athigh concentrations (5 ,ul), itbegantointeract with both sequences on a single oligonucleotide molecule (two shifted complexes) (Fig. 4A). However, Fos-GCN4
gener-ated a single complex of slower mobility with the Double probe, regardlessofprotein concentration(Fig. 4B).
Nucleotide specificity of ZEBRA and Fos-GCN4 binding sites. Point mutations were introduced into the MS-AP-1, ZIIIA,and ZIIIBoligonucleotides inanattempt toestablish whichbase pairs mediate specific DNAbinding by ZEBRA and Fos-GCN4. The oligonucleotides created were MS-AP-1* (TTAGTCA), ZIIIA* (TGAGCAA), and ZIIIB*
(T-CAGCAA).
ZIIIA*and ZIIIB* contain an identical 7-bpcore recognition site (TGAGCAA) but differ in their sur-roundingsequences(see Materials and Methods). Addition-ally, the ZIIIAand ZIIIB oligonucleotides were mutatedat fourpositions togenerate the ZIIIAm and ZIIIBm oligonu-cleotides.
ZEBRA bound with relatively high affinity to the
MS-z z
O G , O < C)
0 = L 0N(Do
> N > N LL >
z z z
= 0 (
U m c
N LL > N LL N U.
FIG. 3. BindingspecificitiesofZEBRAandFos-GCN4 forAP-1 and BZLF1 promoter sequences. (A) Sequences of the ZEBRA (BZLF1)and MS(BSLF2-BMLF1)promoters.AP-1-likesequences are underlined (11, 13), and the transcriptional start site ofthe BZLF1 gene is marked. (B) Gel retardation assay showing the specificities of TrpE (vector), TrpE-ZEBRA(Zl-245),and TrpE-Fos-GCN4 (Fos-GCN4) proteins for oligonucleotides containing the AP-1and BZLF1 promoter sequences: Z-AP-1 octamer,ZIIIA,and ZIIIB (13). The Double oligonucleotide contains sites ZIIIA and ZIIIBin theorientation foundinthe BZLF1promoter. Equivalent
amounts ofbacterially expressedfusion proteins were incubated with eacholigonucleotide(4x 10-13M) for30minpriortoanalysis
oftheretardationcomplexes by gelelectrophoresis.
AP-1, ZIIIA, and ZIIIB sequences
containing
asingle point
mutation (MS-AP-1*, ZIIIA*, andZIIIB*) (Fig.
5A).
How-ever,itwasunabletobind either the ZIIAmorZIIIBmsite,
eachof which contains4-bp changes (Fig.5A). The lackof
binding to the ZIIIAm or ZIIIBm site confirms the
impor-tance of the identified core element. Fos-GCN4 was more fastidious in itsbindingsiterequirements; it
recognized
the ZIIIB*(TGAGCAA)sequencebutdidnotinteract with any of the other mutatedoligonucleotides
(Fig.5B).
ZIIIA* and ZIIIB* differ from the AP-1 consensus at twonucleotides,
while ZIIIB differs at three positions and ZIIIA differs at J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.330.568.230.469.2]ZEBRA DNA-BINDING SPECIFICITY 4037 A
0 - N G O
- a 0 0 - L
-_1W D:
*-.P
MS Double(ZIIIA*71113)
API
B N
L °~
- 0 o o
-MS Double(711
AP-1
FIG. 4. Binding of ZEBRA (A) and Fos-GCN4 (B) oligonucleotide. ZEBRA and Fos-GCN4 proteins we] gel retardation assay with 4 x 10-13 M 32P-labelec Double (ZIIIA plus ZIIIB) probe. Various volum
extract(indicated in microliters) wereincubated with
or Double probe in the presence of poly(dI-dC). Fos-GCN4 both bind to the MS-AP-1 site as a sii indicatedasS.Mobility shift complexes withaslowe
indicatedasD.
only one. ZIIIA* and ZIIIB* contain the same
nition sequence, but their surrounding sequ These results suggest that ZEBRA is considi tolerant of changes within the 7-bp recogniti4 than isFos-GCN4.
Competition analysis to determine the affinitic and Fos-GCN4 for AP-1 and ZEBRA promote
The relative affinities ofZEBRA and Fos-GCN4 for AP-1-° °, related sequences were assayed by competition analysis (Fig. 6). Complex formation between ZEBRAorFos-GCN4 and theMS-AP-1
probe
wasassayed
in thepresenceof threeM4-. D concentrations of cold competitor oligonucleotides. The
relativereduction ofAP-1complex formationisproportional
S tothe affinity of ZEBRAorFos-GCN4 for the
oligonucleo-tidecompetitor. Disruptionwasquantified by comparingthe integrated intensity of each shifted complexasafunction of the concentration of oligonucleotidecompetitor (Fig.6C and D).TheZEBRAcomplex formedwith theMS-AP-1 sitewas
most efficiently competed for with MS-AP-1, ZIIIA*, and ZIIIB* (Fig. 6A and C). A 5-fold molar excess of these
oligonucleotides
eliminated ZEBRAbinding
to 32P-labeledMS-AP-1,whereas a50-fold molarexcess ofZIIIA,ZIIIB, orDoubleoligonucleotidewasrequiredtoachieveasimilar A4ZIlIB) level ofinhibition (Fig. 6A and C). The
ZEBRA/MS-AP-1
complexwasminimally reducedbytheaddition ofa50-fold ItotheDouble molarexcessof Z-AP-1octamerandwasnotaffected
by
theretitrated ina addition ofunlabeledZIIIAm, ZIIIBm, or a control
oligonu-I MS-AP-1 or cleotide with no AP-1-like sequences (Fig. 6A and C and
tes of protein datanotshown). Thus,ZEBRA binds withhighest affinityto
itheMS-AP-1 TGAGTCA (MS-AP-1) and TGAGCAA(ZIIIA* andZIIIB*)
ZEBRA and sites and with a lower affinity to TTAGCAA (ZIIIB),
ngle complex, TGAGCCA(ZIIIA), TTAGTCA (MS-AP-1*), and the
Dou-,rmobilityare bleoligonucleotide (Fig. 6C and 7).
The Fos-GCN4/MS-AP-1 complex was efficiently
com-peted for with a 5-fold molar excess of Z-AP-1 octamer,
20-fold molarexcessof MS-AP-1,or50-fold molarexcessof
7-bp recog- ZIIIB* (Fig. 6B and D). Surprisingly, the Double oligonu-ences vary. cleotide,whichwasboundbyFos-GCN4 inagelretardation erably more assay, was unable to disruptthe formation ofa complexat
on sequence the MS-AP-1 site (Fig. 4; Fig. 6B and D). None of the remaining divergentAP-1 sitescompetedwith the formation esofZEBRA ofaFos-GCN4/MS-AP-1 ata50-fold molarexcess(datanot
!r sequences. shown). A secondcompetition experimentwasalso
quanti-_D
0E < <E
L0 0 o0
*E
wt =
N v - A s <
III
FIG. 5. Gel retardationassayillustrating the binding specificities of ZEBRA and Fos-GCN4 formutantrecognitionsequences.MS-AP-1, ZIIIA, andZIIIBoligonucleotidesweremutatedat asingle position (noted by*); the basesubstitution is underlined.Twooligonucleotides mutated at multiple positions are designated ZIIIAm and ZIIIBm. The control oligonucleotide contains no AP-1-related sequences. Bacterially expressed TrpE-ZEBRA (A) orTrpE-Fos-GCN4 (B) was incubated with these32P-labeledoligonucleotides, and binding was
analyzed byamobility shiftassay. VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.57.290.72.248.2]4038 TAYLOR ET AL.
A
Z AP-1 Octamer MS-AP-1 MS-AP-1' ZIIIA 0 52050 0 5 20 50 0 5 20 50 0 5 20 50
ZEBRA w " w asz
ZlIlABZIIIB Zll Double
0 520 50 0 5 20 50 0 5 20 50 0 5 20 50
ZEBRA
B
Competitor: Z-AP-1Octamer MS-AP-1 ZIIIB' Double
0 52055 05205 0520 50 0 5 20 50
Eaawt.r 2Ifg.au.&add.
MS-AP-1 BSLF2 TCTrCAfGGTGCTTC
TGrrcA313TGCTTC
z
mO
N
U-1 2
2
-Z-AP-1 BZLF1 CTAGAMCCAG~wAGGATC 3 1
(Octamer)
ZIIIA BZLF1 CTAGCTATGCA1AGATC
ZIIIA* - CTAGCTATGCAGATC
2
-1
-Fos-GCN4 _
C
a 0
._
.0
Clcn 0
u CL
H
n
~~Ll
!
5Foldexcess!
E 20Fold excess
~~~~~~~~~~~~~~~~~~ ..
§< §< - 8
Oligonucleotide Competitor
r% U 100
90
a 80
0
m 70 U- 60 50
a
0 40
30
20
10
nI
5 Fold excess
L_ 20 Fold excess
<
Oligonucleotide Competitor
FIG. 6. Competition experiment to evaluate the affinities of
ZEBRA and Fos-GCN4 for AP-1 and ZEBRA promotersites. A
32P-labeled MS-AP-1 oligonucleotide (4 x 10-13M)wasincubated
with bacterially expressed TrpE-ZEBRA (A)or TrpE-Fos-GCN4
(B)inthepresenceof three concentrations of cold competitorDNA.
Complex formationwasresolved by gelelectrophoresis. Only the shiftedcomplexesareshown, and thenatureofthe competitor and itsmolarexcess overthe MS-AP-1 probeareindicated. (C)Results
quantified by scanning of the relative intensity of each shifted complex.Thepercentdisruption of theZEBRA/MS-AP-1complex isplotted for each oligonucleotide competitor.(D) Resultsquantified asforpanel C fordisruption of the Fos-GCN4/MS-AP-1 complex.
ZIIIB BZLF1 CTAGCAGGCAtd1GTACCGATC 2
-ZIIIB* - CTAGCAGGCAf1GTACCGATC 1 3
FIG. 7. Evaluation ofZEBRAandFos-GCN4 binding site affin-ities. Affinities were determined from the competition analysis in Fig. 6. Nucleotide substitutions in the recognition sequences are
noted by bold type. Arelative affinity of1 representsthe highest affinity; - indicates that the protein did not bind the sequence specifically.
fied;therelative affinities of ZEBRA and Fosforthe various
oligonucleotides were found to be the same in the two experiments (data not shown).
DISCUSSION
Comparison ofZEBRA and FosDNA-binding domains. The
construction andexpressionof ZEBRA deletion mutantsin E. coli enabled us to determine the domains within the ZEBRAproteinwhicharerequired forDNAbinding. Unlike
allotheridentifiedtranscriptionfactorswhichbind the AP-1 consensus sequence, ZEBRA does not contain a leucine zipperdirectly adjacent to the basic region (25, 45). How-ever,these studies indicate that theregion3' tothe ZEBRA basic domain (aa 199 to 227) is required for DNA binding.
The C-terminal 18 amino acids (228 to 245) may also con-tribute todimerformationorprotein stability,sincemutants
lacking this region bind DNA in a temperature-sensitive fashion(Fig. 1CandD).Thisregion,exonIII(aa 199to245),
is nothomologous toanyknown dimerizationdomain (45). However, it has been hypothesized thatthis domain forms an alpha helix and that dimerization involves a coiled-coil interaction(14, 23,27, 36). These resultsareconsistent with those of Packham et al., who found that mutants which deleted the same regions ofexonI or III bound DNA in a
salt-dependentmanner,whereas thewild-typeprotein bound
DNA under allconditions (37).
Site-directedmutagenesisofc-fos previouslyidentified the amino acids necessary for DNA recognition of theFos/Jun complex (35). Mutagenesis of Fos amino acids 144 to146or 154 to 156 completely abrogated DNA binding (35). The
corresponding amino acids are conserved in the basic do-main of ZEBRA exon
II,
and proteins mutated in thesepositions wereunableto bind anyof theDNA-binding sites
(MS-AP-1, ZIIIA, or ZIIIB) (Fig. 2). These results suggest that while the DNA-binding specificities of Fos-GCN4 and ZEBRA differ, the same two clusters of basic amino acids arenecessaryfor theDNA-binding abilities of bothproteins.
Furtherwork is neededtodefine the amino acids in the basic
region which confer theunique binding specificities of Fos-J. VIROL.
I
Li
#0
was -1#
4 '. 4p0*0
aw'O
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.340.532.71.248.2] [image:6.612.64.300.73.612.2]GCN4 and ZEBRA. We have conducted dimerization do-mainswapexperimentstoshow that DNAtargetspecificity, invitro, is contained exclusively within the basic region and is not influenced by the nature of the ZEBRA or GCN4
dimerization domain (23).
Binding sitespecificitiesof ZEBRA and Fos-GCN4.ZEBRA and Fos bind similar DNA sequences, and both ZEBRA expression and TPA treatment result in activation ofEBV lytic viral DNAreplication (8,9, 15, 41, 46). An underlying biological question is whether Fos, or other cellular AP-1 proteins, mediates TPA induction of the EBV lytic cycle. Whilewehavenotaddressedthisquestion directly,wehave begun by assessing the binding affinities of ZEBRA and Fos-GCN4 for several sites in the BZLF1 and MS-EA promoters. We chosetocompareZEBRA bindingtothat of Fos-GCN4, since the first indication that ZEBRA was a DNA-binding transactivatorcamefromarecognition of the homology between the basic domains of ZEBRA and c-Fos (11). While Fos generally binds DNA as a Fos/Jun het-erodimer, we used an artificial Fos-GCN4 construct, con-taining the Fos basic domain and the GCN4 leucine zipper, to allow Fos to bind DNA as ahomodimer. Manygroups have previously shown that the target site specificity ofa protein isnotaffected by thenatureof its leucine zipper (24, 32, 34, 40).
Several groups have recently examined ZEBRA binding activity in EBV promoters and the origin of EBV lytic replication (orilyt) (11, 13, 17, 21, 27, 28, 37, 43), but there hadbeennostudiesdirectly comparing the ability of Fos and ZEBRA to bind the same sites. Since our work was com-pleted, Chang etal. (6) reported onthebindingspecificities ofZEBRAand Fos/Junin ori lyt. Ourworkon the binding specificities ofZEBRAand Fosfor sites in the BZLF1 and MS-EApromotersprovides both complementary andnovel observations. Chang etal. demonstrated that ZEBRA binds an AP-1 consensus site with higher affinity than it binds divergent heptamer sites inori lyt, whereas we have found that it binds equivalently to an AP-1 site and a divergent
sequence (ZIIIA* or ZIIIB*) with high affinity (6). ZIIIA* and ZIIIB* contain the same coreheptamersequencefound
in ZRE2 inori lyt (6). Their datasuggestthatZEBRAbinds ZRE2 with high affinity (6). Both studies show that Fos/Jun and Fos-GCN4 are restrictive in binding divergent AP-1 heptamersites. However, wehavefound novel binding ofa Fos-GCN4 homodimer to a divergent heptamer sequence flankedbydyadsymmetry (ZIIIB*).
ZEBRAbounda muchbroaderrange of DNAsequences thandidFos-GCN4(Fig. 4). ZEBRAbound thepalindromic AP-1site,but italsoboundtheZIIIAandZIIIB sites,which do not contain perfect dyads. Unexpectedly, oligonucleo-tides withbase pair substitutions which disrupt the partial dyadnatureofZIIIAorZIIIB(TGAGCCAtoTGAGCAAor TTAGCAA to TGAGCAA) were bound by ZEBRA with increased affinity (Fig. 5 and 7). This departs from the proposed scissors-grip model ofDNA binding by dimeric proteins,inwhicheach proteinmonomerishypothesizedto bind anidentical DNA half-site (45). Although it is known that ZEBRA(expressedin vitroor asaTrpE fusion protein) binds DNA as a homodimer (6, 14, 23, 27), it is unclear whether both monomers contact or recognize symmetrical units on DNA. There are two possibilities which could explainZEBRAbindingto asymmetricsites: first, onlyone subunit ofthe ZEBRAhomodimer contacts DNA; or sec-ond, each subunit recognizes a wide variety of half-site
sequences,therebyrecognizingasymmetricsequences. Our preliminary data suggest that the second hypothesis
ac-countsforZEBRA's
asymmetrical
recognition ofDNA(23).It has also beenfound that the homodimeric glutocorticoid
receptorbinds DNAasymmetricallyandis more sensitiveto mutations in the right half of its binding site (reviewed in reference 4).
Fos-GCN4bindsas ahomodimertothepalindromicAP-1 site (19, 24, 40; Fig. 3). Both Fos/Jun heterodimers and artificial Foshomodimershavebeen showntorecognizethe AP-1consensussequenceasymmetrically (39).This suggests that the Fos basic domains in a homodimer interact with heptamer target sequences in a manner equivalent to a
Fos/Jun heterodimer. However, a Fos-GCN4 homodimer
did not bind any of the divergent AP-1 sites with high affinity. Itwasunabletorecognize ZIIIA,ZIIIB,orZIIIA*,
allof whichwererecognized byZEBRAhomodimers(Fig.4 and7). Fos-GCN4 didbind ZIIIB*(TGAGCAA), although it
didnotbindZIIIA*; botholigonucleotides containthesame
7-bp core recognition sequence. This result suggests that
flanking nucleotides influence binding ability. ZIIIB*, but not ZIIIA*, contains the bases CA andTG adjacent to the
recognition sequence increasing the dyad symmetry ofthe
site, CATGAGCAATG. The importance of flanking se-quences in the binding affinity of Fos was noted by Nakabeppuand Nathans (32). Theyfound that substitutions ofcytosineand
guanosine
foradenine andthyminebasesatflanking positions 1 and 10 of the cyclic AMP response element (CRE) resulted in greater than a 90% decrease of homodimericFosorFos/Junbinding activity (32).Our novel result indicates that the binding site specificity of Fos is
broader than originally proposed; Fos-GCN4 can bind an
asymmetric heptamer sequence with two mismatches from
the AP-1 consensus in an appropriate flanking
oligonucleo-tide environment.
While ZEBRA hasabroaderspecificitythanFos-GCN4 in
recognizingvariousheptamersequencesrelatedtothe AP-1 consensus, only Fos-GCN4 binds the Z-AP-1 octamer se-quence
(TGACATCA)
located in the BZLF1 promoter withhigh affinity.This is interesting sincethis sitewasoriginally defined as the Jun/AP-1 response element in thejun pro-moterand is TPA
responsive
(2). Thus,ZEBRAisunableto binda subset of the knownAP-1/TPA-responsiveelements.Fos/JunaswellasartificialFoshomodimersareknownto bind the octamer CRE
(TGACGTCA),
which differs from theZ-AP-1 octamer atposition5 (GtoA) (6, 12, 32, 33, 38, 39). Although the sites are similar, Deutsch et al. have shown thattheyhavedifferentsignal-responsive
properties;
that
is,
CREs are notresponsive
to TPA (10). Chang et al.have shown that ZEBRAdoesnotbindaCRE(6).
Although
ZEBRAdoes notbind the
TPA-responsive
Z-AP-1octamersite,bindingtothis site may beimportantin theinductionof the EBVlyticcascade. Forexample,Fosoranother cellular AP-1proteinmaybind this site in vivo(2, 3, 10, 26),
leading
to low levels of transcription from the BZLF1 promoter. ZEBRAcan then bindto the ZIIIA and ZIIIB sites in the BZLF1 promoter,
resulting
in autoactivation of this gene(13).
Binding of ZEBRA and Fos-GCN4 to an oligonucleotide with twobinding sites. We studied ZEBRA and Fos-GCN4 bindingto
multiple
sitesastheyarearranged
in the BZLF1 promoter.The arrangements of ZEBRA bindingsites in its ownpromoterand in thelytic-origin divergent
promoterare very similar (28). ZRE1 (TGTGTAA) and ZRE2(TGAG
CAA)
(identical
totheheptamercoresequenceintheZIIIA*andZIIIB*
oligonucleotides)
in thelytic
origin
arearranged
in a tail-to-tailfashionseparated
by
10bp
(28).
Sites ZIIIA (TGAGCCA) and ZIIIB (TTAGCAA) arearranged
in theon November 10, 2019 by guest
http://jvi.asm.org/
4040 TAYLOR ET AL.
same manner in the BZLF1 promoter, separated by 6 bp. This arrangement may be a common mode of targeting ZEBRAtranscriptional controlinthe EBV genome.
We found that two ZEBRA homodimers can bind ZIIIA and ZIIIB simultaneously on the Double oligonucleotide
(Fig. 3). We have not determined which site isfilledinitially,
although footprinting data have demonstrated that ZEBRA binds ZIIIB withhigheraffinitythan ZIIIA (13, 27).Gel shift
experiments with limitingdilutions ofZEBRAproteinshow
that botharebound only afterasignificant portion of single sites arefilled. Furthermore, competition experiments
indi-cate that ZEBRA does not have a greater affinity for the
Double oligonucleotide than for either single site (Fig. 6A andC).Thus, ZEBRAbindsnoncooperativelytothese sites. However, binding of ZEBRA to these two adjacent sites
may allow forsynergistictransactivation.
Fos-GCN4, which did not bind oligonucleotides bearing single ZIIIA orZIIIB sites, bound the Double oligonucleo-tide(containing ZIIIAandZIIIB)as ahigh-molecular-weight complex (Fig.3and4). Theassociation ofthisslow-mobility complex can be disrupted by addition ofZEBRA protein,
suggesting that the two proteins compete for a similar sequence ontheDoubleprobe(unpublished data). However,
the Doubleoligonucleotide didnotinterferewithFos-GCN4/
MS-AP-1 complex formation(Fig. 6B andD). One explana-tion for this phenomenon is that these associations are noncompetitive because the twoprobes bind different
Fos-GCN4complexes. The MS-AP-1 probeisbound by a Fos-GCN4homodimer,whereas theDoubleprobemaybindonly
a Fos-GCN4 complex with more than two monomers and
perhaps with otherproteins.
The interaction ofZEBRA with cellular AP-1 sequences mayresultin thedisplacement ofFos orothercellular AP-1
factors, thereby precluding theiractivity. Alternatively, by binding AP-1 sites, ZEBRA may transactivate (orpossibly
transrepress) cellular genes in a manner analogous to that with cellular AP-1 transcription factors. AP-1 proteins are
probably not able to activate transcription of EBV genes
through ZEBRA-responsive elements because atleast one
AP-1-binding protein, Fos-GCN4, is unable to bind those sites. The divergent nature oftheZEBRA-responsive sites makes it unlikely that cellular AP-1 proteins can have an
ongoingrolepromotingviraltranscriptionandactivatingthe EBVlytic cycle. Additionally, theabilityof Fostobind the TPA-responsive Z-AP-1 octamerand CRE sites,which are notboundby ZEBRA,indicatesthat ZEBRA cannotsimply
substitute for Fos. The ability ofthese related proteins to
bindsimilaraswellasdistinct sites adds yet another level of
complexity to the relationship of these transactivators in
regulating cellular and viral geneexpression.
ACKNOWLEDGMENTS
We aregratefulto E. Manet, A. Sergeant, and P. Farrell for the gifts ofBZLF1cDNAs, T.Kouzarides and E.Zifffor the Fos-GCN4 construct, and T. J.Koernerfor the pATHexpression plasmids. We also thank D. Crothers for his support and advice. N.T. would like toacknowledgethe support of E. & E. Lichtenstein.
This work was supported by Public Health Service grants CA12055, CA16038, and A122959 (to G.M.); RO1CA143 and R01CA52004 (to S.H.S.); IF32CA08482-01 (to E.F.); and CA09159-15 (toR.B.) from the National Institutes of Health and by grant NP 669K (toG.M.)from the American CancerSociety.N.T. is supported by the Medical Scientist Training Program of the NationalInstitutes of Health.
REFERENCES
1. Angel, P.,E. A.Allegretto,S.Okino,K.Hattori,W.J.Boyle,T. Hunter,and M.Karin.1988.Oncogene junencodesa sequence-specifictrans-activator similartoAP-1. Nature (London)332: 166-171.
2. Angel, P., K. Hattori,T. Smeal,and M. Karin. 1988. Thejun proto-oncogeneispositively autoregulated byitsproduct,Jun/ AP-1. Cell55:875-885.
3. Angel,P., M. Imagawa, R.Chiu, B. Stein, R.J. Imbra,H.J. Rahmsdorf,C.Jonat,P.Herrlich,and M.Karin.1987.Phorbol ester-induciblegenescontainacommoncis elementrecognized byaTPA-modulatedtrans-actingfactor. Cell 49:729-739. 4. Beato, M. 1989. Gene regulation by steroid hormones. Cell
56:335-344.
5. Biggin, M.,M. Bodescot,M.Perricaudet, and P.Farrell.1987. Epstein-Barr virus gene expression in P3HR1-superinfected Rajicells. J. Virol. 61:3120-3132.
6. Chang, Y.-N.,D.L.-Y.Dong, G. S.Hayward, andS. D. Hay-ward. 1990. TheEpstein-Barrvirus Zta transactivator:a
mem-ber of thebZIPfamilywithuniqueDNA-binding specificityand
a dimerization domain that lacks the characteristic heptad leucinezippermotif.J.Virol.64:3358-3369.
7. Chavrier, P.,H.Gruffat,A.Chevallier-Greco,M. Buisson,and A.Sergeant.1989. TheEpstein-Barrvirus(EBV) earlypromoter DRcontainsacis-actingelementresponsivetothe EBV
trans-activatorEB1andanenhancer with constitutive and inducible activities.J. Virol.63:607-614.
8. Chevallier-Greco, A., E. Manet, P. Chavrier, C. Mosnier, J. Daillie, and A.Sergeant. 1986. BothEpstein-Barrvirus(EBV) encoded trans-acting factors, EB1 and EB2, are required to
activatetranscriptionfroman EBVearlypromoter. EMBO J. 5:3243-3249.
9. Countryman, J.,andG.Miller. 1985. Activationofexpression
of latent Epstein-Barr herpesvirus after gene transfer with a small clonedsubfragment ofheterogeneous viralDNA. Proc. Natl.Acad.Sci. USA81:7632-7636.
10. Deutsch,P.J.,J.P.Hoeffier,J.L.Jameson,andJ.F. Habener. 1988. Cyclic AMP and phorbol ester-stimulated transcription
mediatedbysimilarDNAelements thatbinddistinctproteins.
Proc.Natl. Acad.Sci. USA 85:7922-7926.
11. Farrell,P.J., D. T. Rowe, C. M. Rooney, and T.Kouzarides. 1989. Epstein-Barr virus BZLF1 trans-activator specifically
bindsto a consensusAP-1site and is relatedtoc-fos. EMBOJ. 8:127-132.
12. Flemington, E., andS. Speck. 1990. Identification of phorbol esterresponseelements in thepromoter ofEpstein-Barrvirus putativelyticswitchgene BZLF1. J. Virol. 64:1217-1226. 13. Flemington, E.,and S.Speck. 1990. Autoregulationof
Epstein-Barr virusputative lyticswitch gene BZLF1. J.Virol. 64:1227-1232.
14. Flemington, E.,andS. H.Speck.1990. Evidenceforcoiled-coil dimerformation by an Epstein-Barr virus transactivator that lacksaheptadrepeatof leucine residues.Proc.Natl. Acad. Sci. USA87:9459-9463.
15. Gradoville, L., E. Grogan, N. Taylor, and G. Miller. 1990. Differences in the extent of activation of Epstein-Barr virus
replicativegeneexpressionamongfournon-producercell lines
stablytransformedbyoriP/BZLF1plasmids. Virology 178:345-354.
16. Hammerschmidt, W., and B. Sugden. 1988. Identificationand characterization oforilyt,alyticoriginofreplicationof
Epstein-Barrvirus.Cell55:427-433.17. Hardwick,J.M.,P. M.Lieberman,andS. D.Hayward.1988.A
newEpstein-Barrvirustransactivator, R,inducesexpressionof
acytoplasmicearlyantigen.J. Virol.62:2274-2284.
18. Holley-Guthrie, E. A., E. B. Quinlivan, E.-C. Mar, and S. Kenney.1990.TheEpstein-Barrvirus(EBV)BMRF1promoter forearlyantigen(EA-D)isregulated bythe EBV trans-activa-tors, BRLF1 andBZLF1, in acell-specific manner. J. Virol. 64:3753-3759.
19. Hope,I.A.,and K.Struhl.1987.GCN4,aeukaryotic
transcrip-tional activatorprotein,bindsas adimertotargetDNA.EMBO J. 6:2781-2784.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
20. Kenney, S.,J.Kamine, E. Holley-Guthrie,J.C. Lin, E. C. Mar, andJ. Pagano.1989. The Epstein-Barr virus (EBV)geneBZLF1
immediate-earlygeneproductdifferentially affects latentversus
productive EBVpromoters.J.Virol. 63:1729-1736.
21. Kenney, S., J. Kamine,D. Markovitz, R. Fenrick, and J. Pagano.
1988. An Epstein-Barr virus immediate-early gene product
trans-activates gene expression from the human immunodefi-ciency long terminal repeat. Proc. Natl. Acad. Sci. USA 85: 1652-1656.
22. Kleid, D. G., D. Yansura, B. Small, D. Dowbenko, D.M. Moore,
M.J. Grubman, P. D. McKercher, D. 0.Morgan, B.H.
Rob-ertson,andH. L.Bachrach. 1981. Cloned viral protein vaccine for foot and mouth disease responses in cattle and swine. Science214:3834-3838.
23. Kolman, J., N. Taylor,and G. Miller.Unpublished data. 24. Kouzarides, T., and E. Ziff. 1989. Leucine zippers of fos, jun
andGCN4 dictatedimerization specificity and thereby control DNAbinding. Nature (London)340:568-571.
25. Landschulz, W. H., P.F. Johnson, and S.L. McKnight. 1988. The DNA-binding domain of theratliver nuclearprotein C/EBP isbipartite. Science 243:1681-1688.
26. Lee, W., P.Mitchell, and R.Tjian. 1987. Purified transcription factor AP-1 interacts with TPA-inducible enhancer elements. Cell49:741-752.
27. Lieberman, P. M., andA. J. Berk. 1990. In vitro transcriptional activation, dimerization, and DNA-binding specificity of the Epstein-Barr virus Zta protein. J. Virol. 64:2560-2568. 28. Lieberman, P. M., M. J. Hardwick, J. Sample, G. S. Hayward,
and S. D. Hayward. 1990. The Zta transactivator involved in inductionof lytic cycle geneexpression in Epstein-Barr virus-infectedlymphocytes bindstoboth AP-1 and ZRE sites intarget promoterand enhancer regions. J. Virol. 64:1143-1155. 29. Luka, J., B. Kallin, and G. Klein. 1979. Induction of the
Epstein-Barr virus (EBV) cycle in latently infected cells by n-butyrate. Virology 94:228-231.
30. Manet, E., H. Gruffat, M. C. Trescol-Biemont, N. Moreno, P. Chambard, J.F.Giot, and A. Sergeant. 1989.Epstein-Barr virus bicistronic mRNAs generated by facultative splicing code for
twotranscriptional trans-activators. EMBO J. 8:1819-1826. 31. Melton, D. A., P. A. Krieg, M. R. Rebagliati, T. Maniatis, K.
Zinn, and M. R. Green. 1984. Efficient in vitro synthesis of biologically active RNA and RNA hybridization probes from plasmids containing a bacteriophage SP6 promoter. Nucleic Acids Res. 12:7035-7056.
32. Nakabeppu, Y., and D. Nathans. 1989. The basic region of Fos mediatesspecific DNA binding. EMBO J.8:3833-3841.
33. Nakabeppu,Y., K. Ryder, and D. Nathans. 1988. DNA binding
activitiesofthreemurineJunproteins: stimulation byFos. Cell 55:907-915.
34. Neuberg, M., J. Adamkiewicz, J. B. Hunter, and R. Muller. 1989. A Fos protein containing the leucine zipper forms a homodimerwhichbinds totheAP1binding site.Nature (Lon-don)341:243-245.
35. Neuberg, M., M. Schuermann, J. B. Hunter, and R. Muller. 1989. Twofunctionallydifferent regions inFos arerequired for the sequence-specific DNA interaction of the Fos/Jun protein complex.Nature(London)338:589-590.
36. O'Shea, E. K., R. Rutkowski, and P. S. Kims. 1989. Evidence that theleucine zipperisacoiled-coil. Science 243:538-543. 37. Packham, G., A. Economou, C. M. Rooney, D. T. Rowe, and
P.J. Farrell. 1990. Structureandfunction of the Epstein-Barr virus BZLF1protein. J.Virol. 64:2110-2116.
38. Rauscher, F. J., P. J. Boulalas, B. R. Franza, Jr., and T. Curran. 1988. Fos and Jun bindcooperatively tothe AP-1site: recon-struction invitro.GenesDev.2:1687-1699.
39. Risse, G., K. Jooss, N.Manfred, H. J. Bruller,and R. Muller. 1989.Asymmetricalrecognition of the palindromicAP1binding site (TRE)byFosprotein complexes. EMBOJ. 8:3825-3832. 40. Sellers,J.W., and K. Struhl.1989.ChangingFosoncoproteinto
aJun-independent DNA-binding protein with GCN4 dimeriza-tionspecificity by swapping 'leucine zippers.'Nature(London) 341:74-76.
41. Takada, K., N.Shimizu, S. Sakuma,andY. Ono. 1986. trans-activation of the latentEpstein-Barrvirus(EBV)genomeafter transfection of theEBV DNAfragment.J.Virol.57:1016-1022. 42. Taylor,N., J. Countryman, C. Rooney, D. Katz, and G. Miller. 1989. Expression ofthe BZLF1latency-disriptinggenediffers in standard and defective Epstein-Barr viruses. J. Virol. 63: 1721-1728.
43. Urier, G.,M.Buisson, P. Chambard, and A.Sergeant.1989. The Epstein-Barr virus early protein EB1 activates transcription from differentresponsive elements includingAP-1binding sites. EMBOJ.8:1447-1453.
44. van Straaten, F., R. Muller, T. Curran, C. van Beveren, and I.M. Verma.1983.Completesequenceofahumanc-oncgene: deduced amino-acidsequenceof the human c-fosgeneprotein. Proc. Natl. Acad. Sci. USA 80:3183-3187.
45. Vinson, C. R., P. B.Sigler,andS. L. McKnight. 1989. Scissors-gripmodel forDNArecognition by afamily of leucine zipper proteins. Science 246:911-916.
46. zur Hausen,H.,F.J.O'Neill,U. K.Freese, and E. Hecker.1978. Persisting oncogenic herpesvirus induced by the tumor pro-moter TPA.Nature(London)272:373-375.

![FIG. 4.oligonucleotide. Binding of ZEBRA (A) and Fos-GCN4 (B) ZEBRA and Fos-GCN4 proteins we]](https://thumb-us.123doks.com/thumbv2/123dok_us/1313845.84789/5.612.57.290.72.248/fig-oligonucleotide-binding-zebra-fos-gcn-zebra-proteins.webp)