Genetic
Expression
in
Heterozygous Replicative
Form
Molecules of fX174
V. MERRIAM,1 L. B. DUMAS,2 AND R. L. SINSHEIMER Division of Biology, CaliforniaInstitute ofTechnology, Pasadena, California 91109
Received for publication 4January1971
Heterozygous replicative form molecules of bacteriophage 4X174
deoxyribo-nucleic acid (DNA) have been constructed in vitro. These are composed of viral
strands extracted from purified preparations ofphage bearing ts mutations and
complementary strands of either half lengthorfulllength synthesized with purified
DNA polymerase, in vitro, on DNA from am3 phage. In infections with such
heterozygous DNA, involving mutations in each of four different cistrons, phage
with the genotype ofthe complementary strand comprised 1 to 20% ofthe total
phage produced by a spheroplast population. From single-burst analysis of the
progeny from DNA heterozygous in onecistron (B), itappearsthat those phage
with the genotype ofthe complementary strand arise as major components in a
small proportion of the infected cells rather than comprisingaminor componentin
most cells. The implications of such a pattern ofexpression are discussed with
respect tomechanisms of phage DNA synthesis.
The step-by-step
description
of nucleic acidsynthesis during 45X174 infection has formed a
coherent picture (14). The infecting viral strand
is convertedtoadouble-stranded
replicative
formmolecule (RF). This
parental
RF, i.e., thatcon-taining the
infecting
viralstrand,
is bound to amembrane site at which it is able to serve as
template
for ribonucleic acid(RNA)
synthesisand to undergo semiconservative
replication
viathe
rolling
circle mode. Thecomplementary
strand is elongated as
synthesis
ofnewcomple-mentary strands occurs on the double-stranded
circle;
anewviral
strand isbuilt
onthiselongated
complementary
strand(6, 12).
Thetail,
nowdouble-stranded
andcontaining
theoriginal
complementary
strand from theparental
RFand a newly synthesized viral strand, is
released
and closed into acircleasadaughterorprogeny
RF. If there are additional membrane sites
available, thisprogeny RF may utilize themand
become comparable to the true parental RF in
function; ifthere are noadditional sites available,
this progeny RFremainsin the
cytoplasm,
inertwith respect to semiconservative replication (15)
and perhaps to RNAsynthesis (10;J. W. Sedat,
Ph.D. Thesis, Calif. Institute of Technology,
1970). Later in infection, these progeny RF
molecules participate in progeny viral
single-strandsynthesis.
1Present address: Department ofBacteriology, University of California, Los Angeles, Calif.90024.
2Present address: Department ofBiological Sciences,
North-westernUniversity, Evanston, Ill. 60201.
The activities of the infecting viral strands
have been extensively documented, whereas the
activities of othercomponentsof thesystemhave
been inferred from population studies. In this
study we have undertaken to study specifically
the role of the parental complementary strand,
i.e., the initial partner of the parental viral
strand. For this purpose we have constructed
heterozygous RF molecules containingamutant
viral strand and a wild-type complementary
strand and used these to infect spheroplasts
under both permissive and nonpermissive (with
respect to the specific mutation) conditions.
The expected
physical
fate of the parentalstrands according tothe model described above
is showndiagrammatically in Fig. 1.The genetic
fate is
inferred
from standard semiconservativereplication. Thus, the initial replication of the
RF leads to release ofthe parental
complemen-tary strand into an RF with genotype dictated
by the complementary strand. In cells with a
single replication site, this RF would remain
inertinthecytoplasm until itenteredinto
single-strand synthesis. All subsequent RF molecules
producedwouldhave the genotype of theparental
viralstrand, which remains at thereplicatingsite.
OX
messenger RNA has the same sequence as theviral strand (9). Thus, phage-specific RNA
synthesizedpriortothe firstreplicationwouldbe
of the complementary genotype. If, as has been
proposed, onlythesite-bound RF can be used as
template
for messenger RNA, then subsequent603
on November 11, 2019 by guest
http://jvi.asm.org/
MERRIAM, DUMAS, AND SINSHEIMER
Initial
Replication
.ID
+
Subsequent Replications
+
Double
---~* Strand Synthesis
+
::::::
:P:::
.i.jtj
a
Single Strand Synthesis
a
a
a
FIG. 1. Diagramnmatic representation2 of DNA replication in
OX174.
Circles represenit RFmolecules, the lefthemispherecorresponiding to the viral strand, theriglhtto thecomplementarystranid. The parentalviral and com-plementary strandsare those marked withP. The genotypeofthecomplenmenltarystrandof theinifectingparental RFisindicatedbystippling.
to the first RF replication the RNA synthesized
would have the viral genotype.
One would predict then that most of the
phage progeny of a heterozygous parental RF
would beof the viralgenotype, whereas aminor
proportion of the phage would be of the
com-plementary genotype. Under conditions
non-permissive for the viral strand, there may be
significant rescue of the infection as a
conse-quenceof thepresenceofwild-type RNA formed
veryearly ininfection, but the distribution of the
genotypes should not beradically different from
the infection done under permissive conditions
MATERIALS AND METHODS
Media. Cells for phage production or assay were grown in tryptone-KCl broth (2). Cells to be used for spheroplasts were grown in 3XD medium (8), and the infection was carried out in PA and PAM media (8). Deoxyribonucleic acid (DNA) and phage
were stored in 0.05 M
tris(hydroxymethyl)amino-methane (Tris), pH 8.0. SSC is 0.15 M NaCl and
0.015M sodium citrate.
Phageandbacteria. OX174mutantsusedweream3 (cistron E), ts4 (cistron H), ts79 (cistron G), ts9 (cistron B), and ts41D (cistron F). Grown under nonpermissive conditions, am3 phage provide in-increasedburst sizesas aconsequenceof their failure tolysethe cells. CistronsF, G,and Hareresponsible
for structuralcomponents of the virus, whereas the function of the cistron B productisuncertain. There
isevidence that cistrons F, G, and H may comprise an operon (R. M. Benbow et al., in preparation). Stocks were prepared in Escherichia coli C, a
non-suppressing host, at 30C. Sucrose (10%) wasadded
to the medium at the time of infection with the ts mutants to prevent lysis. After 2 hr, infected cells were concentrated by centrifugation and lysed on resuspension in 0.05 M borate buffer; the debris was removed bycentrifugation. This procedure permitted increased titers and facilitated purification.
Thephagewereassayed onE. coli HF4714 (SuX), anamsuppressorstrain,atthepermissive temperature of 30C. Alltemperature-sensitivestockswereassayed forrevertantsbyplatingonSuXat41 C.Stockswith revertant frequencies > 10-5 were discarded. E. coli C,anonsuppressorstrain,wasalso used as indicator.
E. coliK12W6(su-hcr+rec+)was used for routine
spheroplast assays.E.coli AB2480(su-herA recA 13)
wasused inspheroplast assays where indicated.
DNA preparation: viral strand. The temperature-sensitive phage were concentrated as described and
the cell lysate was placed directly on a 5 to 20%
sucrosegradientin 0.05 Mborate.Aftercentrifugation
(SW25.1 rotor, 3hr,58,000 X g,5C), fractionswere collected and assayed for phage. Thepeak fractions werepooledanddialyzed for 1 hragainst0.05MTris, pH 8.0. Thephage suspensionat 1012 to 10's plaque-forming units per ml was then extracted once with water-saturated phenol and three times with ether; the etherwasremovedby bubbling with airat 37C. Complementary strand. The purified am3 phage
suspension was extracted three times with phenol at 45 C and treated with ether and bubbling as above.
604 J. VIROL.
op.
on November 11, 2019 by guest
http://jvi.asm.org/
This material served as template in reactions with
purifiedDNApolymerase;thecomplementarystrand
product was labeled with32P-labeled deoxyadenosine
triphosphate (5, 7). Two classes of material were
synthesizedin separateexperiments.(i)Inthe presence
of excess initiator, the product at onefold synthesis
consistedof segments ofapproximately one-half unit
length asdeterminedbysedimentation through alka-line sucrose. These aredesignated "random halves,"
as there is no reason to postulate specific initiation
points.(ii)Inthe absence of added initiator,
endog-enousinitiatoractivity permitted 0.33-fold synthesis.
Theproductwasprimarily unit-lengthlinearstrands,
designated here as "full-length linears" or simply
"linears."
The viral strandtemplate wasremoved byalkaline
sedimentation (SW25.1 rotor, 0.1 M KOH, 24 hr,
58,000X g, 5C). One sedimentation of the random
halves provided material containing only 10-5 viral circles (by weight) as measured bythe infectivity of
the preparation. With the linear strands, three
suc-cessive sedimentations were required to reduce con-taminationto10-3 (by weight).The finalproductwas
approximately homogenous but sedimented slightly
more slowly than marker full-length linear strands. This is a consequence of repeated selection of the
trailing edgeof the linear strandregion during
puri-fication.
The contamination by weightwas ascertained by
direct infectivity assay of the DNA preparations;
it is therefore a measure of circular viral strands. Any linear viral strandspurified alongwith the linear
complementary strands would escape detection. On
self-annealing, any circular or linear viral material
couldreactwith the linearcomplementarystrandsto forminfectious RF. In allexperiments, therefore,the
background infectivity of the self-annealed
comple-mentary material was measured to determine the totalcontribution ofcontaminating "viral material."
A correction for this contribution was subtracted
from the results attained with the heterozygous RF
moleculesbefore calculation of theexpression of the
complementarystrand.
Annealing.DNAsamplesin 0.05 M Tris,pH 8.0,
were mixed at concentrations between 1011 and 1012
strands per milliliter (0.3to 3,g/ml), and 24X SSC
wasaddedtogiveafinal concentration of2.4XSSC.
The solution was incubated at67 C for 1 hr; longer
incubation(upto15hr)gavenochangein theamount
of complementary strand expression. Under these
incubation conditions, single-stranded DNA circles
sustain0.1 hit/hr,asdetermined frominfectivityloss.
Biological assays.Phagewereassayedin the stand-ardmanner.Theinfectionofspheroplastswascarried
out according to Guthrie and Sinsheimer (8). Two
dilutions of each sample were always included to insure that the phage yieldwas linear (ona log-log
plot) for the DNA concentrations used. Progeny phage
were assayed at 30 and at 41C. The ratio of the
number ofphage plating at41C (corrected for the
yield from infections initiated by the self-annealed
complementary strand material) to the number of
phage plating at 30C is defined as the expression
of the complementary strand. Correction of the 30
C plating is appropriate, but insignificant.
When yields were to be examined under
nonper-missive conditions, the DNA and spheroplasts were mixed and incubated for the initial 15 min at 41 C. Prewarmed (41 C) PAM was added; half of the mixture was incubated at 41 C, and half was shifted downto 30 C. This pretreatment did not reduce the yield of either wild-type or temperature-sensitive phage formed after shift-down. After 2 to 3 hr, the spheroplasts were lysed by 10- to 100-fold dilution into 0.05 M borate buffer.
For single-burst analysis, the DNA was diluted in
0.05MTris,pH 8.0,andmixed with an equal volume
ofspheroplasts.Anappropriateamountof PAM was
added, and two drops of the solution were added
from a 5-ml pipette (0.1-ml portions) to each of 100 to 200 tubes. All unnecessary shear was avoided to
maintain the integrity of the spheroplasts. After 2
hr of incubation at the desired temperature, the
spheroplastswere lysed by theaddition to each tube
of 0.4 ml of 0.05 M Tris, pH 8.0. The whole sample
wasplated onE. coli SuX at 30 C. All progeny will
plate under these conditions. After 6 hr all plaques
were picked with sterile toothpicks and streaked on
SuXat41 C (platesprewarmedto37C) todetermine
temperature sensitivity and on E. coli C at 30 C to determine whether they wereamber. Forsuch
analy-sis, it is criticalto useyoung plaques, that is before
diffusion can lead to cross-contamination of the
plaques, and to have warm plates for the
temperature-sensitivity analysis.
In the single-burst experiments, a Poisson distri-bution of infected centers in the tubes is assumed. Thefraction ofplateswithout any plaques is therefore
used to calculate the average number of infected
centers per sample. Only series with initial concen-trations of <0.2 infected centers per portion were
analyzed. Theratio olDNA strands to spheroplasts
togive appropriateconcentrations of infective centers
was 1/200 at 41 C and 1/6,000 at 30 C. The actual
concentrations of DNA are on the order of106
strands/mlat41 Cand 105 strands/ml at 30 C, with
the actual concentration varying somewhat for
differentspheroplast preparations.
RESULTS
Annealing of viral and complementary strands.
The small
complementary
strands of4X174
have a
Cot
number of 5 X 10-3 moles per literpersecond (1) andthereforeanneal rapidly even
at low concentrations. At equal input of the
complementary
andviralstrands, there is a rapidinitial coalescence to yield a double-stranded
moleculewith nophysically detectable,free,
single-strandedmaterialremaining. A slowerregistering
within thisdouble-stranded molecule, which has
somesingle-stranded properties, leadsto a
prod-uct indistinguishable by sedimentation velocity
or neutral buoyant density from native RF
II,
thecirculardouble-stranded molecule containing
single-strand breaks in one or both strands. The
on November 11, 2019 by guest
http://jvi.asm.org/
MERRIAM, DUMAS, AND SINSHEIMER
infectivity ofthe single-strand mixture drops by
10- to40-foldduringannealing;the RF IIisolated
from infected cells is typically 1/20 as infectious
as the same number of single-stranded circles.
Concentration dependence of expression.
Ac-tivity,orexpression,of thecomplementarystrand
will bedefined in termsof itsability to causethe
appearanceofphage of thecomplementarystrand
genotype in an infection initiated by a
hetero-zygous RF.
Numerically
it is the proportion ofthe phage in the burst which plate at 41 C (see
below). With no guidelines for prediction, one
might expect such expression to be relatively
independent of the cistron examined and to be
maximal at the equivalence point of one gene
added in the complementary material for each
gene present in the viral material.
Viral DNA was mixed with various amounts
of
complementary
strand in 2.4X SSC andannealed at 67 C for 1 hr, a time sufficient to
yield
RF identical to native RF by the criterialisted above. This DNA was then used, without
further
purification,
to infectspheroplasts
at30Cfor2hr. Attheendofthistime, the
sphero-plasts were lysed by dilution and the progeny
were assayed at 30 and 41 C. The heterozygous
RF molecules have the genotype represented as
v(+ tsx)/c(amE +), i.e., the viral strand is
temperature-sensitive in cistron x,
wild-type
inall other cistrons; the
complementary
strand isamber in cistron E,
wild-type
in the remainderof the cistrons, in
particular
in x.Thus,
thosephage
plating
at 41 C either represent revertantsin the viral strand or have arise in some way
from the information of the
complementary
strand. In aU cases, the revertant
frequency
fortheviral strand is <
10-5.
Thecontribution fromtheself-annealed
complementary
strand materialis more
significant,
amounting
to 1 to10%
thevalues from infections initiated
by
theheterozy-gous RF. All values are therefore corrected for
the number of
phage
produced
by
theself-an-nealed complementary material. The fraction of
phage platingat 41C, afterthis
correction,
com-paredtothetotal phage
plating
at30C,isdefinedas the expression ofthe
complementary
strand.The activity of the
complementary
strand istherefore defined here
only
in terms of progenygeneration; no
implication regarding
RNAformed during infection is made.
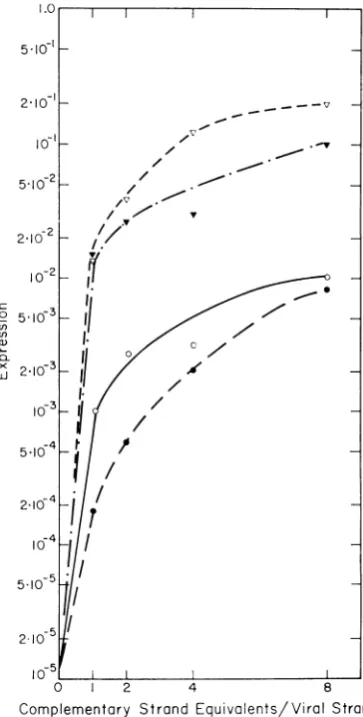
Figure 2 shows the dependence of
expression
for cistrons G and B on the relative number of
the
complementary
strandequivalents.
Maximumexpression doesnot occur ata1 :1
input;
expres-sion
actually
increases withincreasing
comple-mentary strand input in both cistrons examined
to 10-fold that observed atthe
equivalence
point,
whether the
complementary
random halves orlinears are used. A 10-fold difference in expression
in these two mutant cistrons is apparent at all
levels. To extend this study, expression was
examined for four cistrons at a 1:1 input by
1.0
5210-l _
2-10-510 .o-2
510-3
2-10-3
05
l16
0 2 4 8
Complementary Strand Equivalents/Viral Strand FIG.2. Expression in
heterozygouis
RFmolecuiles
as afunction ofthe relativenunberofviral and
com-plementary strands in the annealing mixture. Viral
strands extractedfrom purified temperature-sensitive phage (cistronsGandB)were mixed at -1J11
strandsl
ml in 2.4X SSCwith amounts of complementarystranzd
material equivalent to 1, 2, 4, and 8 times the viral strand present. Both random half and linear
com-plementary strands were used. Each mixture was
incuibatedat 67 Cfor I hr, dilutedwith 0.05 M Tris
(pH 8.0), and used to infect spheroplasts. Expression oftlhe complementary strand is defined by the ratio ofthe number ofphage
platinig
at 41 C (after correc-tion; see Materials and Methods) to the total phageplatinigat30C.Symbols: 0,cistron G,complementary
strand linear; *, cistron G, complementary strand halves; 7, cistron B, complementary strand linear;
Y, cistron B, complemenztary strand halves.
606 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.495.268.450.137.496.2]REPLICATIVE MOLECULES OF PHAGE
OX174
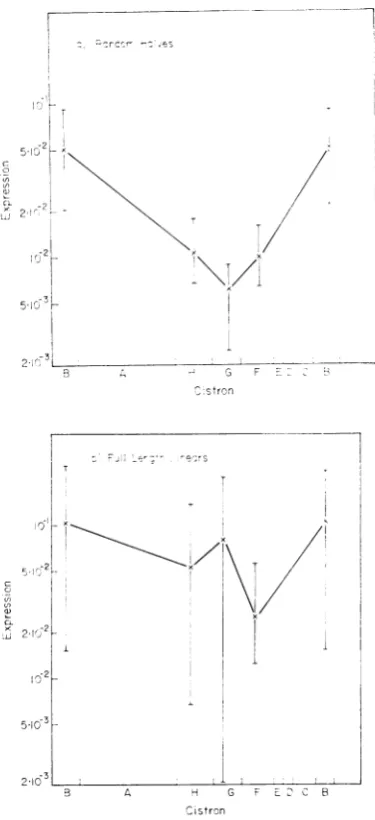
weight (Fig. 3a and b). For the random halves
(Fig. 3a), expression is significantly lower in
cistrons F, G, and H than in cistronB;with the
full-length linears (Fig. 3b), the average values
show less variationalthough therange of values
in different assays is greater. The source of this
variation, which isgreater betweensamples than
within samples, is unknown.
Thus, neither expectation (the maximum
ex-pression atequivalenceandequivalent expression
of all cistrons) wasrealized. The comparison of
expression for random halves and linears in the
four cistrons suggests that all portions of the
genome are not
completely comparable
undertheseconditions.
Activity of the complementary strand under
nonpermissive conditions. If the infected
sphero-plasts are incubatedat 41C, cellsin which
only
temperature-sensitive protein appears do not
yield phage. Thus, the phage
yield
forsingle-strandedDNA fromphage bearinga tsmutation
drops at 41 Cbya factor of 103. In infections by
v(+ tsB)
/c(amE
+)
where theexpression
ofcistron B under
nonpermissive
conditions issufficient to give
yields significantly
greater thanthe
background
fromtemperature-sensitive
DNAalone, the
plaques
may beanalyzed individually.
The absolute number of ts-mutant
phage
in a41 C
lysate
fromspheroplasts
infectedby
av(+ tsB)
/c(amE
+)
exceeds thatexpected
fromthe ts-mutant DNA alone
by
10- to 100-foldand comprises 13% of the
plaques analyzed.
Itappearsthatthe
wild-type
complementary
strandin the
parental
RFmaythenrescuetheinfection,
probably through
the formation ofwild-type
messenger RNA
prior
tothe firstreplication
ofthe
parental
RF.Importance ofrepair to expression. If all
ex-pression
inheterozygotes
arises from randomrepair of one strand to
yield orthoduplexes
ofthe two
possible
types(3), expression
should begreatly suppressed
in hcrrec-spheroplasts.
Heterozygotes of the four cistrons were used to
infect AB 2480
(rec
13hcrA)
and K12W6(rec+-hcr+).
As shown in Table 1, expression in allcases is
comparable
in the two hosts.Repair
canthen
play only
a minor roleinexpression.
Single-burst analysis. The fact that
expression
isessentially
thesame in hcrrec cellsasinhcr+rec+cells indicates that the entity responsible for
expression of the complementary genotype is a
heterozygous RF molecule. As outlined in the
introduction, one may make predictions as to
the composition of a burst from a single cell
infected with such amolecule. The data thusfar
presented have concerned onlytotal populations
ofphageresultingfrom the burstsofmany cells.
Repair occurringin some cells coulddistort the
apparent pattern of expression, notably for the
RFmoleculescontainingcomplementary material
of less thanfulllength,thatistherandomhalves.
Some cellular conversion of one or two
half-length pieces to a full-length linear
complemen-tary strand is obviously necessary. If only one
K~~~~~~~
- C:crr.ies
2~~~~
T
5,Ir-2 kXs
w 210
2-1C ,\
3
^
~~~~G
F_stron
211; - lIr ne<
3 A H G F ED C B
Cistron
FIG. 3. Expression in heterozygous RF molecules in cistronsB, F, G, and H. Viral DNA from purified temperature-sensitive phage (cistron B = ts9; F = ts4;G = ts79;H= ts41D)was mixed at 1011
strandsl
ml with101J
strand equivalents of wild-typecom-plementarystrandof (a) halflengthl or (b) full length.
Expression is defined as in Fig. 2. Lines indicate the rangeofvalues found in anumberofassays; the average values are indicated and joined. Abscissa distances
represent the estimated relative sizes of the cistrons (Benbow et al., in preparation).
607
VOL. 7, 1971
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.495.256.444.151.562.2]MERRIAM, DUMAS, AND SINSHEIMER TABLE 1. Effect of repairon expression in
heterozygousRFmoleculesa
Expressionin Cistron
hcr+rec+
hcr-rec-F 0.061 0.019
G 0.066 0.100
H 0.026 0.014
B 0.082 0.140
a
Heterozygous
RF molecules, prepared at a10:1 ratio of complementary linear strand to viralstrand, wereassayed in K12W6 spheroplasts
(hcr+rec+) and inAB2480spheroplasts (hcr-rec-).
Values in the rec-hcr- host are corrected for the relative efficiency of control single-stranded DNA in the two hosts.
piece were retained, RF molecules of genotype
v(+ tsx)/c(amE tsx) and v(+ tsx)/c(+ +)
mayresult inadditionto theexpected v(+ tsx)/
c(amE +). Site-specific changes may also occur
for less obvious reasons with the
full-length
linear strands. It is clear that a more profound
analysis would result from examination of
bursts from single infected spheroplasts. In
addition, the cistron E marker in the
comple-mentary strand material, which has been
men-tioned butnotutilizedto this
point,
willprovide
additional insight into intracellular events. The
general prediction is that in bursts of
single
spheroplasts
the progeny corresponding to theviralgenotypeshould be inexcessandthatmost
bursts willincludeasmall number ofprogenywith
thecomplementary genotype.
Single
bursts ofinfections initiated withheter-ozygous RF molecules of genotype c(+ tsB)/
c(amE +)
containing
random halves or linearcomplementary strands were examined under
bothpermissiveand
nonpermissive
conditions asdescribed above. The burstsizes ofthese
sphero-plasts wereextremelyvariable, both fromcontrol
single-stranded DNA infections and from
heter-ozygousRFinfections.The average burst in both
cases was
approximately
40 phage per infectedspheroplast;
the range was from 1 to 200phage
perburst.
Thegenetic
analyses
of thesingle
burstsappearin Table 2.
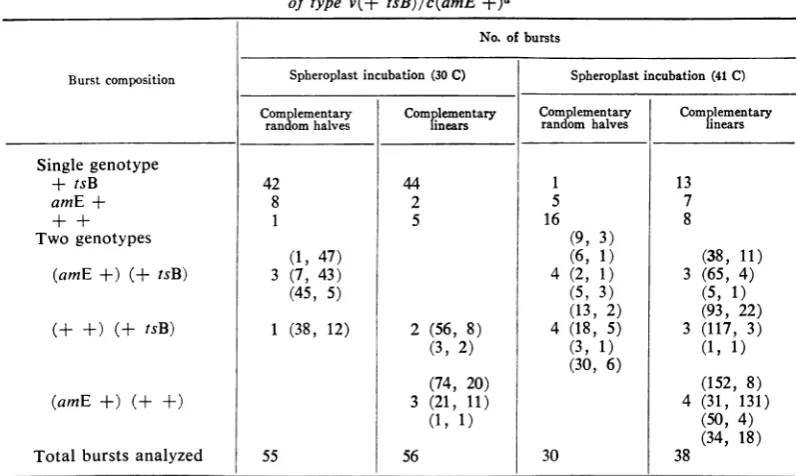
Though
mixed bursts are observedwith both types ofRF,under both
permissive
andnonpermissive conditions, in all cases the
single
genotype bursts are in excess. Under
permissive
conditions, the
majority
ofcellsyield phage
only
of theviral genotype, + tsB. Such bursts would
atise ifall of the
complementary
strandmaterialwere eliminated
by
the cellprior
to RFreplica-tion. Under
nonpermissive
conditions,
none ofthese bursts shouldbe observed; however, there is
rescue of such bursts in the case of heterozygous
RF molecules containing linear complementary
strands. The presence of the complementary strand may therefore be inferred in such cells
(al-though it does not appear in the progeny) and
thus, by extrapolation, in some of those cells
producing onlyviral genotype bursts under
per-missiveconditions.
Asthe burst-size distributionis comparable for
spheroplasts yielding single or mixed genotype
bursts, it may be concluded that the 10 to 20%
expression ofthecomplementarystrand character
in the cistron B region under permissive
condi-tions arises from 10 to 20% of the cells yielding
all or mostlycomplementary type phage, rather
thanmostofthecells yielding10 to 20%
comple-mentary type phage and 80 to 90% viral type
phage.
Most bursts under nonpermissive conditions
contain the selected (+) complementary strand
markerin the cistronBregion. In the unselected
cistron region, the markerfrom the
complemen-tary
material,
amE,ispresentincombinationwith thecomplementary
strand cistron B region (+) in9of29cases intheprogenyof RF containingrandomhalvesand in 14of25 cases in the
prog-enyofRFcontaininglinears.There is thus greater
linkage between the cistron E and cistron B
re-gionwhenthe complementarystrand is of greater
length. Thisobservationis inaccord with the
pro-posalthat some DNAelimination occurs in the
RFmolecules containing several random halves
per
complementary
strand.A significant portion of the bursts under all
conditions ismixed. Those appearing under
per-missive conditions might be attributed to tubes
containing more than one infected center; the
number appearing under nonpermissive
condi-tionsis muchtoogreattobeso
explained
and maybeassumedto arisefromasingle spheroplast.In
mostof these bursts the cistronmarkerfrom the
complementary
strand is inexcess. Intotal,thesedata indicate that in those bursts where phage
with the genotype of the complementary strand
occur at all, they comprise all or most of the
population
ofthe burst. DISCUSSIONThe current model for DNA synthesis in
4X174-infected
cells is outlined in theintroduc-tion. Such a description is consistent with the
apparent failure of direct parental-to-progeny
transfer and the
requirement
for theintegrity
ofthe
parental
viral strand late inwild-type
infec-tion. The role of the
parental
complementary
strand in this model is quite
subsidiary.
If theparental RF is heterozygous, as described here,
608 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
TABLE 2. Single-burst analysis ofspheroplasts infected withheterozygousRFmolecules oftype v(+ tsB)/c(amE+)a
No.of bursts
Burstcomposition Spheroplastincubation (30 C) Spheroplast incubation (41 C) Complementary Complementary Com lementary Complementary
randomhalves linears randcom halves lnears
Single genotype
+ tsB 42 44 1 13
amE + 8 2 5 7
+ + 1 5 16 8
Twogenotypes (9, 3)
(1, 47) (6, 1) (38, 11)
(amE +) (+ tsB) 3 (7, 43) 4 (2, 1) 3 (65, 4)
(45, 5) (5, 3) (5, 1)
(13, 2) (93, 22)
(+ +) (+ tsB) 1 (38, 12) 2 (56, 8) 4 (18, 5) 3 (117, 3)
(3, 2) (3, 1) (1, 1)
(30, 6)
(74, 20) (152, 8)
(amE +) (+ +) 3 (21, 11) 4 (31, 131)
(1, 1) (50, 4)
(34, 18)
Totalbursts
analyzed
55 56 30 38a An RFsamplecontaining1:1 viral andcomplementary strand materialwasdiluted such that stan-dardtreatmentduringthespheroplastassayyieldedasolutioncontaining <2 infectedspheroplasts/ml.
Anequalvolume ofspheroplasts was addedto aportion ofthe diluted RF solution and the mixture
wasincubatedat41 C for 15 min.Eightvolumesof PAM wereadded,and thesamplewasquickly
dis-tributedin0.1-mlsamples (<0.2infectedcenters/tube) priorto2 hrofincubation of thespheroplasts
at30 Corat41 C. Theinfected cellswerethenlysed osmotically byaddition to each tube of 0.4 ml of
Tris, pH8.0. The entire tubewas platedonEscherichia coli SuXat30 Ctopermitall progenyto plate.
After 6 hr ofincubation, allplaqueswerepicked andtested onSuX at41 C and onE. coliC at 30 C. Wild-typephagein thecistronBregionwill platein the former case;wild-typephage in thecistron E
regionwill platein thelatter. By directexamination one maythereforedescribe the entire genome of
eachphagein the burst. Burstswereidentifiedascontainingasinglegenotypeor morethanonegenotype
andsorecorded. Figuresinparenthesesfor mixed burstsgivethe actual numbers of thetwophagetypes
observedin the mixed bursts in theorderindicated.Thetotals representpooleddata fromanumber of
similarexperimentsdoneatdifferenttimes. In anygivenexperiment,theP(O) was >0.8,foranaverage
of <0.2 infected centers/tube.
the
wild-type
complementary
strand would bereleased in the first daughter RF molecule. It
could then expressitselfgenetically onlyin
single
strandsmade from that RF. It couldbe
expressed
phenotypically through RNA synthesis prior to
RF replication. Under conditions nonpermissive
to theviral
strand,
this RNAmight
beadequate
toeffect some rescueoftheinfection.
Itis
thought
thatmost,ifnotall,
spheroplasts
have onlyonesite for DNA
replication.
Geneticrecombinationisobserved butnot
complementa-tion (11;C. A.HutchisonIII, Ph.D.Thesis,
Cali-fornia Institute of
Technology,
1969). Theactualdegree ofRFreplication withinaspheroplasthas
notbeenmeasured. Ifonlyasinglereplication of
parental
RF occurs, theviraltype RFwouldre-mainatthemembrane where it might not
partici-pate in progenysynthesis.Theonly daughter RF
would then be of
complementary
genotype andonly
complementary
typephage would appearinthe burst. Additional
replication
of the parentalRF would produce additional viral-type RF
molecules and would be expected to reduce the
relative contributionof thecomplementary
geno-type RFto the pool. Theproportion of
comple-mentarygenotypeRFmolecules woulddrop as a
function ofthereplication of the parentalRF and
presumably
so wouldtheproportion of progeny ofcomplementary
strandgenotype.The essential result obtained in these
experi-mentsisthe expression of the genotype (at the ts
locus) of thecomplementarystrands in infections
withheterozygousRF. This expression is at least
one order of magnitude greaterthan can be
ex-plained away asa consequence of gene reversion
or homozygous RF contamination. It does not
on November 11, 2019 by guest
http://jvi.asm.org/
MERRIAM, DUMAS, AND SINSHEIMER
seem to be at alldependent upon the presence or
absence of a host cell DNA repair system (3, 13).
Theexpression ofcomplementary strand
geno-type is quantitatively significant. In allt cases the
progeny from a population of spheroplasts
in-fectedwithheterozygousRFmoleculescontained
between 1%and 10%-, phage with the genotype of
the complementary strand. It has been observed
with mutants in each of four different cistrons.
It is observed equally well whether the
comple-mentary strands are present as approximately
viralDNAlengthpieces orhalf-viralDNAlength
pieces.
Thatthe genotype of thecomplementarystrand
is present inonlyaminority of the progenywould
beexpectedfrom the model. However,
examina-tion ofsingle bursts of infected cells disclosed a
patternfar from expected. Rather than most cells
yielding mixed bursts with afew complementary
type phage, the majority of infected cells yield
progenyexclusively ofthe viralgenotype (i.e., ts).
Only a small class of cells yielded bursts with
phage of complementary genotype; these
con-tained in many cases progeny ofexclusively
com-plementary genotype.
Insome ofthose cellswhichyielded onlyviral
typephage, thepresenceatsomestageof the
com-plementary strand may be inferred from the
rescue ofthese bursts under conditions
nonper-missive for the viral strand phenotype. In the
majority of suchcells, however, itisatleast
possi-blethat thecomplementarystrand may havebeen
eliminated.
Various hypotheses may be advanced to
ac-countforthese observations. Asrecombinationis
observed in
spheroplasts,
it is evident that morethan one RF can penetrate these bodies. The
failure to observe
complementation
suggests alimitation upon templates for transcription, but
this is not
necessarily
true forreplication.
Thestructures of the products of viral and
comple-mentary strand
annealing
used in theseexperi-ments are notknown. Thus,in some instances a
viral strandmaybeannealedtodifferent
portions
of two
complementary
strands(or
viceversa).
Thesepossibilities, combined withseverely
limitedRFreplicationandgrossly unequaluseofoneor
two out ofa small number ofRF molecules as
templates
for progeny DNAsynthesis,
couldgive
rise to the observedresults.
Another explanation would involve the
com-plete failure of RF replication in a fraction of
cells.Inthiscaseall progeny virusbutone
(which
could
occasionally
be missed) would be ofcom-plementary strand genotype.
A more interesting
explanation
would allot amoresignificant roletothe
complementary
strandin the determination of progeny virus genotype
than is expected from the model previouLsly
de-veloped.
Suchexperiments
asthose recentlyper-formed (V. Merriametal.,in
pr-ess),
in which thedecay
ofa tritiumatom attachedtothe 5-carbon ofcytosine
in the viral strand of a parental RFwas unab!e to induce mutation, whereas the
de-cay ofa similar tritium atom in the
complemen-tary strand of a parental RF was markedly
effective,
are also suggestive of anassignment ofmuch greater significancefor thecomplementary
strand ofthe parentalRFinthe determination of
progeny genotype.
The results recently described by Dressler and
Wolfson
(4)
led themto proposeamodelofOX
RFreplicationviaelongationof the viral strand.
Such a model would give the complementary
strand the master role in the determination of
progeny genotype. This model is in direct
con-tradiction to one proposed earlier (13) derived
from a different line ofevidence.
Unfortunately,
theexperimental datapresented byDressler and
Wolfson donotunambiguously resolve the
issue;
therearetwomolecularproductsfrom each DNA
replication and the interpretation of their data
depends uponanassumptionastowhichproduct
is observed in their studies.
Adoption of such an alternative model would
thenrequire aspecial explanation, such as
elimi-nation of the complementary strand, for the
majorityofspheroplasts inwhich the viral strand
genotype is dominant among the progeny.
It is conceivable that thefirst daughter RF is
distinct fromsubsequent daughter RF and shares
with the parental RF the responsibility for RF
replication. The relative involvement of thetwo
might
vary,dependentupon externalfactors.ACKNOWLEDGMENTS
Thisresearch wassupported by Public Health Service granit GM 13554 fromi the National Institute of General Medical Sciences.
One ofus(V.M.) wishes to thank Dan Ray for the use of the facilities of UCLA for the comiipletion of some of these experi-ments.
LITERATURE CITED
1. Britten, R. J., and D. E. Kohne. 1968. Repeated sequences in DNA. Science 161:529-540.
2. Denhardt,D.T.,andR. L.Sinsheimer. 1965. The process of infection with bacteriophiage OX174. Ill. Phage matura-tion andlysis aftersynchronized infection. J. Mol. Biol. 12:641-646.
3. Doerfier,W.,andD. S.Hogness. 1968. Gene orientation in bacteriophage lambda as determlined fromgenietic activi-ties ofheteroduplex DNA formed in ritro. J. Mol. Biol. 33:661-678.
4. Dressler, D., and J. Wolfson. 1970. Therollingcircle for-OX DNA replication. Ill. Synthesis of supercoiled duplex rings.Proc. Nat.Acad.Sci.U.S.A.67:456-463. 5. Dumas, L. B., G.Darby, and R. L. Sinsheimer. 1970. The
replication ofbacteriophage ,X174 DNA in ritro.
Tem-610 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
peratureeffectsonrepair synthesis and displacement
syn-thesis. Biochim. Biophys. Acta228:407-422.
6. Gilbert, W., and D. Dressler. 1968. DNA replication: the rolling circle model. Cold Spring Harbor Symp. Quant. Biol. 33:473-484.
7. Goulian, M., and A. Kornberg. 1967. Enzymatic synthesis of DNA. XXIII. Synthesis ofcircular replicative form of phage4X174DNA. Proc. Nat. Acad. Sci. U.S.A. 58:1723-1730.
8. Guthrie, G. D., and R. L. Sinsheimer. 1963. Observationson
the infection of bacterial protoplasts with the deoxyribo-nucleicacid of bacteriophage MX174. Biochim. Biophys. Acta72:290-297.
9. Hayashi, M., M. N. Hayashi, and S. Spiegelman. 1963. Restriction ofin *iro genetic transcription to oneof the complementary strands of DNA. Proc. Nat. Acad. Sci. U.S.A.50:664-672.
10. Hayashi, M. N., and M. Hayashi. 1968. The stability of
native DNA-RNA complexes during in vivo 4X174 trans-cription. Proc. Nat. Acad. Sci. U.S.A. 61:1107-1114. 11. Jansz, H. S., C. van Rotterdam, and J. A. Cohen. 1966.
Geneticrecombination of bacteriophageOX174in sphero-plasts. Biochim. Biophys. Acta 119:276-288.
12. Knippers, R., M. Whalley, andR.L. Sinsheimer. 1969. The
process of infection with bacteriophage 4X174. XXX. Replication of double-stranded4,X174. Proc. Nat. Acad. Sci. U.S.A. 64:275-282.
13. Russo,V.E. A.,M. M.Stahl, and F. W.Stahl. 1970. On the transfer ofinformation from oldtonewchains ofDNA duplexes in phage lambda: destruction of heterozygotes. Proc.Nat. Acad. Sci. U.S.A. 65:363-367.
14. Sinsheimer, R. L. 1970. "Contagium Vivum Fluidum," after seventy years. Koninklijke Nederlandse Akademie van
Witenschappen, Proc., Series C, 73:307-316.
15. Sinsheimer, R. L., R. Knippers, and T. Komano. 1968. Stages in the replication of bacteriophage 4X174 DNA
invivo.Cold Spring Harbor Symp. Quant.Biol.33:443-447.