Manipulation of Aphid Behavior by a Persistent Plant Virus
Maliheh Safari,a,b*Matthew J. Ferrari,b,cMarilyn J. Roossincka,b,c
aDepartment of Plant Pathology and Environmental Microbiology, Pennsylvania State University, University Park, Pennsylvania, USA
bCenter for Infectious Disease Dynamics, Pennsylvania State University, University Park, Pennsylvania, USA
cDepartment of Biology, Pennsylvania State University, University Park, Pennsylvania, USA
ABSTRACT Plants are frequently infected with cytoplasmic RNA viruses that persist
for many generations through nearly 100% vertical transmission without producing any symptoms. Movement between plant cells and horizontal transmission have not been observed with these viruses; instead, they are distributed to all host cells through host cell division. Jalapeño peppers (Capsicum annuum) are all infected with Pepper cryptic virus 1 (PCV-1; family Partitiviridae). We compared the effect of odor cues from PCV-1-infected (J⫹) and virus-free (J⫺) jalapeño peppers on the aphid Myzus persicae, a common vector of acute plant viruses. Pairwise preference experi-ments showed a stark contrast to insect-plant interactions in acute virus infections— that is, the virus-infected plants deterred aphids. The acute plant virus Cucumber mosaic virus (CMV) manipulates its host’s volatile emissions to attract aphid vectors and facilitate its transmission. We inoculated J⫹and J⫺ plants with CMV. Volatiles of J⫹and J⫺CMV-infected plants were more attractive to aphids than those of J⫹ and J⫺ mock-inoculated plants. However, in pairwise preference experiments with J⫹ CMV- and J⫺ CMV-infected plants, aphids preferred the J⫺ CMV volatile blend. Aphid reproduction on J⫹and J⫺plants was measured as an indicator of the effect of PCV-1 on host quality for aphids. Aphid reproduction on J⫹ plants was more than 2-fold lower than that on J⫺plants.
IMPORTANCE This study demonstrates that a persistent plant virus can manipulate
aphid behavior. This manipulation is in stark contrast to previously described effects of acute viruses on their hosts that facilitate their transmission. This study demon-strates a positive relationship between Pepper cryptic virus 1 and jalapeño pepper (Capsicum annuum) plants wherein the virus protects the plants from the vector of acute viruses and reduces aphid herbivory. This work reveals an important implica-tion of persistent plant viruses for pest and pathogen management in agriculture.
KEYWORDS aphid behavior, beneficial virus, partitivirus, plant volatiles
B
ased on their lifestyles, plant viruses are divided into two main groups: acute viruses and persistent viruses (1). Most well-studied plant viruses are acute viruses that cause disease in domestic plant hosts. In plants, infections with acute viruses can be resolved by recovery, death, or conversion to chronic infection. In contrast, persis-tent plant viruses replicate in their hosts for many generations, perhaps thousands of years. They do not cause obvious disease. The name “persistent” comes from their lifestyle in the host, which differs from that of viruses that are persistently transmitted by a vector (1–3). The persistent viruses are distributed to all plant cells through host cell division. They are not able to move between plant cells, and even grafting cannot transmit these viruses. There is no evidence for their horizontal transmission by a vector, whereas they are vertically transmitted to seeds at rates close to 100% (1, 4, 5). Unlike persistent animal viruses, persistent plant viruses cannot convert to an acute lifestyle because they lack the genes for movement.CitationSafari M, Ferrari MJ, Roossinck MJ.
2019. Manipulation of aphid behavior by a persistent plant virus. J Virol 93:e01781-18. https://doi.org/10.1128/JVI.01781-18.
EditorAnne E. Simon, University of Maryland,
College Park
Copyright© 2019 American Society for
Microbiology.All Rights Reserved. Address correspondence to Marilyn J. Roossinck, [email protected].
*Present address: Maliheh Safari, Department of Biochemistry and Biophysics, University of California, San Francisco, San Francisco, California, USA.
Received7 October 2018
Accepted28 January 2019
Accepted manuscript posted online13
February 2019
Published
crossm
17 April 2019
on November 6, 2019 by guest
http://jvi.asm.org/
Vectors play a crucial role in the transmission of acute plant viruses because plants are largely immobile (6). Plant-feeding insects are the most common plant virus vectors, and among insects, aphids are the most common vectors of agriculturally important plant viruses (7). Aphids probe any plant they land on to determine if it is a suitable food source (8), and during this probing process, many viruses can be transmitted. Because of the dependence of plant viruses on vectors for their transmission, viruses evolved ways to manipulate some host and vector features to facilitate transmission.
Studies on the impact of acute plant viruses on insect behavior began in 1951 and found that Aphis fabaepopulation growth was higher on virus-infected sugar beets than on healthy plants (9). Studies on two viruses from the familyLuteoviridae(Barley yellow dwarf virus[BYDV] andPotato leafroll virus) and their aphid vectors showed that virus infection changed aphid development time and reproduction (10). In addition, virus-induced changes in hosts of both luteoviruses encouraged aphid feeding on infected plants, which led to rapid population growth of aphids and subsequent dispersal of viruliferous aphids (11–14). These aphid behavioral changes were consis-tent with the transmission mode of luteoviruses by aphids. Luteoviruses are acquired by vectors feeding for hours to days on the phloem tissues of infected plants (6, 15, 16). Both viruses affected volatile organic compounds (VOCs) of their host plants, and vectors were responsive to host VOCs (12, 13, 17). The nonviruliferous aphid vector of BYDV preferred infected host plants, while viruliferous aphids preferred uninfected hosts; these differences promote pathogen spread. It has been demonstrated byin vitro feeding that aphid behavioral changes are mediated directly by virus acquisition (18). In soybean plants, the aphidRhopalosiphum maidisremains on healthy plants longer than on plants infected withSoybean mosaic virus. Leaving infected plants shortly after probing increases the probability that an aphid will find another plant and transmit the virus (19).
Cucumber mosaic virus(CMV) is a widespread plant pathogen with more than 1,200 host species (20). Aphid vectors can acquire CMV during brief probing of infected plant cells and transmit the virus effectively if the vector disperses rapidly to a new host plant (6, 16, 21). CMV-infected squash plants, despite their reduced size, have elevated levels of VOCs that attract aphid vectors to the infected plants (22). CMV infection initially attracts aphids but then reduces the quality and palatability of plants so that aphids disperse quickly after virus acquisition (22). Salicylic acid- and jasmonic acid-mediated host defense responses are not involved in diminishing plant quality for aphids; instead, CMV infection induces changes in the concentrations of carbohydrates and free amino acids in the plant tissue that cause a reduction in plant quality for aphids. All of these virus-induced changes in the plants favor CMV transmission: they attract aphids to feed on the infected plant, which then disperse quickly to efficiently transmit the virus (23). It was shown recently that although CMV infection induced quantitative and qualitative changes to the VOC emissions of tobacco plants, it did not change aphid preference for infected or uninfected tobacco plants (24). Inoculation of a squash isolate of CMV onto another host, pepper, did not result in the pattern observed on the native squash host. In addition, the virus-mediated changes in the new host were not consistent with CMV-vector interactions to aid its transmission (23). Hence, virus-host-vector interactions appear to be adaptive.
The most common persistent viruses in plants, and indeed the most common plant viruses overall in wild plants, belong to the familyPartitiviridae(25, 26). The partitivi-ruses were first reported as cryptic vipartitivi-ruses (5). ThePartitiviridaefamily includes viruses from diverse hosts: plants, fungi, and oomycetes (27, 28). Phylogenetic analysis of partitivirus RNA-dependent RNA polymerase implies that these viruses have been transmitted between plant and fungi (1). Plants cannot be cured of these viruses using various treatments (29), although occasionally a virus-free plant will develop due to a rare lack of vertical transmission.Pepper cryptic virus 1(PCV-1) belongs to the genus Deltapartitivirus and consists of two linear, monocistronic double-stranded RNAs (dsRNAs) that are encapsidated in separate isometric particles (30–32). PCV-1 was first reported from the Capsicum annuum cultivar Jalapeño M, and as with all persistent
on November 6, 2019 by guest
http://jvi.asm.org/
plant viruses, there is no evidence for its transmission by any vector, mechanical inoculation, or grafting, whereas ovules or pollen transmit the virus to over 98% of the next generation (30, 32).
Persistent viruses are very common in crop plants; hence, it seems likely that they were selected during domestication, and we hypothesize that these viruses provide some advantages to the plants. While there are many studies on the effects of acute viruses on plant volatiles that impact insect behavior, no studies have looked at the effects of persistent viruses. Here, we used three isogenic lines of jalapeño plants with and without PCV-1 to assess their attraction and plant quality traits for aphids.
RESULTS
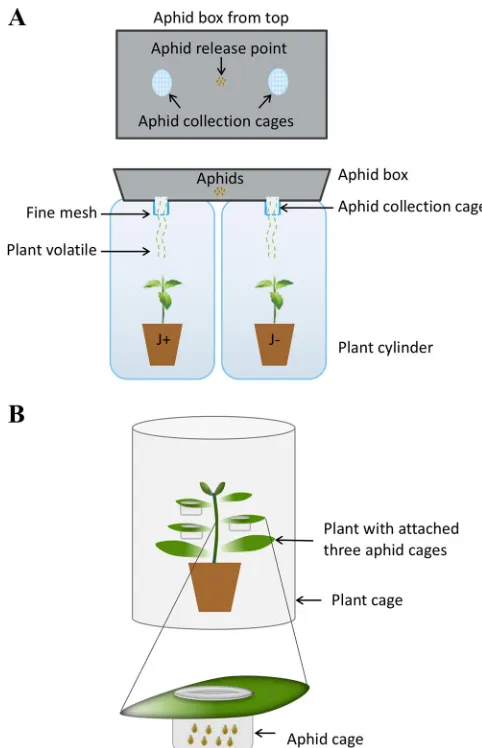
Aphid preference for volatiles of plants with or without PCV-1.To examine the response of Myzus persicae to VOC cues, we performed three pairwise preference experiments in which aphids were exposed to odors of (I) two empty cylinders, (II) an empty cylinder or a PCV-1-infected jalapeño plant (J⫹), and (III) an empty cylinder or a virus-free jalapeño plant (J⫺) (Table 1; Fig. 1a), without any visual cues. In experiment I, there was no significant difference between choosing either of two empty cylinders, while in experiments II and III, aphids selected cylinders containing plants. Figure 2a shows the mean percentages of aphids arrested in collection cages for each experi-ment, and the pairs involved in each choice test are shown bidirectionally. The 95% confidence intervals for the probability of selecting empty cylinder were 0.46 to 0.57 for experiment I, 0.23 to 0.34 for experiment II, and 0.22 to 0.33 for experiment III (Fig. 2b); only in the preference test with both empty cylinders (experiment I, empty versus empty) was 0.5 (i.e., no preference) within the 95% confidence intervals. Hence, aphid choice was not random, and they were responding to volatile cues. In experiments IV-1, IV-2, and IV-3, three isogenic lines of jalapeño with and without PCV-1 were used to discover how the virus affected aphid behavior. Similar patterns were observed in all three lines, and aphids demonstrated their preference for volatiles of virus-free plants (J⫺). The 95% confidence intervals for selecting J⫹over J⫺plants were 0.36 to 0.49 (IV-1, J1⫹versus J1⫺), 0.37 to 0.49 (IV-2, J2⫹versus J2⫺), and 0.31 to 0.41 (IV-3, J3⫹ versus J3⫺), as shown in Fig. 2b. Aphids showed discrimination against volatiles of PCV-1-infected plants, and 0.5 was not contained in any of the 95% confidence intervals. Using three different jalapeño lines verified that aphid behavior is affected by manipulation of plant volatiles by PCV-1 infection.
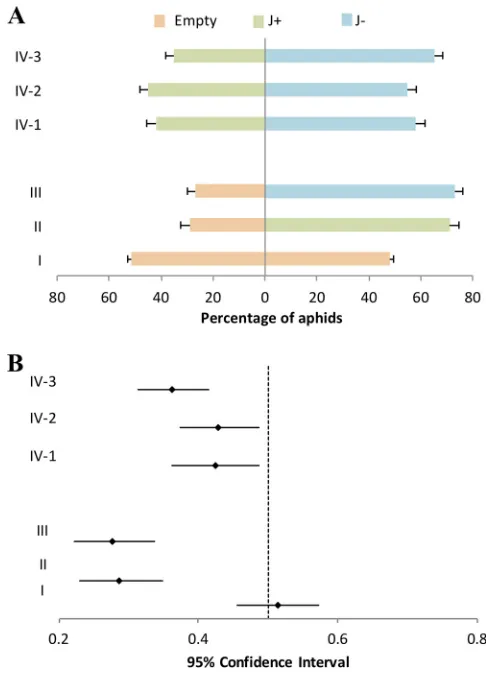
[image:3.585.40.379.83.204.2]Aphid preference for volatiles of plants with or without PCV-1 and CMV.Several studies have shown that CMV manipulates its host by inducing elevated levels of volatile emissions that attract aphid vectors for its transmission (22, 23). In order to explore the effect of PCV-1 on aphid attraction to CMV-infected plants, we inoculated J⫹and J⫺plants with CMV and performed aphid pairwise preference studies (Fig. 1a) in experiments V to IX. We compared aphid attraction to volatiles of mock-inoculated virus-free plants (J⫺ mock) versus CMV-infected plants (J⫺ CMV and J⫹ CMV) in experiments V and VI (Table 1). In both experiments, aphids preferred volatiles from CMV-infected plants (Fig. 3a). In arrangements VII and VIII, pairwise comparisons were TABLE 1Aphid preference experiment arrangements
Expt no. Arrangement
I Empty vs empty
II Empty vs J⫹
III Empty vs J⫺
IV-1 J1⫹vs J1⫺
IV-2 J2⫹vs J2⫺
IV-3 J3⫹vs J3⫺
V J⫺mock vs J⫺CMV
VI J⫺mock vs J⫹CMV
VII J⫹mock vs J⫹CMV
VIII J⫹mock vs J⫺CMV
IX J⫹CMV vs J⫺CMV
on November 6, 2019 by guest
http://jvi.asm.org/
between mock-inoculated PCV-1-infected plants (J⫹mock) and CMV-infected plants (J⫹ CMV and J⫺ CMV). A similar pattern was observed, and aphids significantly preferred the volatiles from CMV-infected plants (Fig. 3a). The 95% confidence intervals for the probability of selecting mock-inoculated plants in arrangements V to VIII ranged from 0.27 to 0.41 (Fig. 3b). However, in pairwise comparisons of aphid responses to volatile emissions from CMV-infected J⫹and J⫺plants (IX, J⫹CMV versus J⫺CMV), aphids preferentially selected the volatiles of J⫺ CMV plants (Fig. 3a). The 95% confidence interval for choosing J⫹CMV was 0.32 to 0.41 (IX, J⫹CMV versus J⫺CMV) (Fig. 3b).
Aphid reproduction on plants with or without PCV-1.Virus infection results in biochemical and physical changes in the host, and aphid reproduction was used as an indicator of the impact of PCV-1 on plant quality for aphids. We compared the reproduction of nonviruliferous aphids that developed from day one to adulthood on virus-free and PCV-1-infected plants over 14 days. Since it took 8 to 9 days to
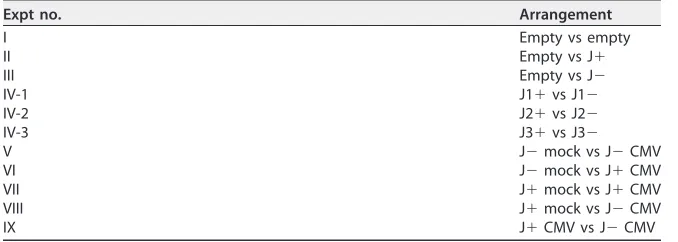
FIG 1Experimental design for aphid preference and reproduction tests. (a) Schematic of experimental set up for aphid pairwise preference for plant volatiles. Wingless, nonviruliferous 3- to 5-day-old aphids were placed in the middle of the aphid box, which was covered to eliminate any visual cues affecting aphid behavior. Aphids could make a choice between two collection cages that were connected to plant cylinders containing treatments. Each collection cage had one layer of fine mesh to trap aphids in the cage, and the number of aphids in each collecting cage was recorded 45 min after releasing aphids. (b) Aphid reproduction was tested on 6- to 8-leaf-old jalapeño plants. Eight adult wingless aphids were confined in a small clip cage and placed on a caged plant (3 clip cages per plant). These adult aphids were removed after 24 h, and offspring were left in each clip cage. Offspring molted to adults and produced new offspring. Aphid reproduction in each clip cage was calculated by dividing the number of offspring produced by the number of adult aphids on day 14.
on November 6, 2019 by guest
http://jvi.asm.org/
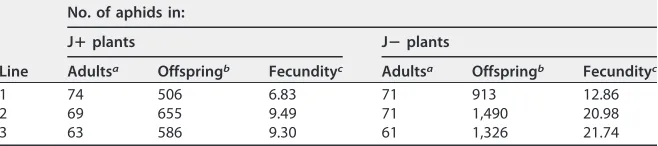
[image:4.585.82.323.70.444.2]complete the first reproduction cycle, only one generation of offspring was born within this period. Average reproduction on J1⫹(74 adult aphids) was 7, whereas reproduction on J1⫺ (71 adult aphids) was 13. For J2⫹ (69 adult aphids), repro-duction was 9.5 compared to 21 for J2⫺ (71 aphids). For J3⫹(63 aphids), repro-duction was 9.3 compared to 22 for J3⫺(61 aphids) (Fig. 4; Table 2). In all three lines, aphid reproduction on the PCV-1-infected plants (J⫹) was more than 2-fold lower than that on virus-free plants (J⫺). Statistical analysis indicated that the effect of the virus on aphid reproduction was significant (Pvalue of⬍0.001). This showed the positive role of PCV-1 in protecting its host from increasing population growth of aphids. In line 1, aphid reproduction for both treatments (J1⫹ and J1⫺) was lower than in the other two lines. Although the conditions used were the same, the experiments were not all done at the same time, so comparisons are valid only within an experiment, although we cannot rule out distinct plant qualities in the different lines. In all three lines, we saw the same trend of higher reproduction on
FIG 2Aphid preferences for volatiles of plants with or without PCV-1. The following control pairwise preference tests (I to III) were performed to test if aphids were responding to volatile cues: (I) empty versus empty; (II) empty versus J⫹; and (III) empty versus J⫺. In experiments IV-1 to IV-3, aphid preferences for volatiles of three different isogenic lines of jalapeño were tested, as follows: (IV-1) J1⫹ versus J1⫺; (IV-2) J2⫹ versus J2⫺; and (IV-3) J3⫹ versus J3⫺. (a) Numbers represent the mean percentages of aphids in collection cages at the end of each treatment, and pairs involved in each choice test are shown horizontally. In arrangement I, there is no significant difference between choosing either of two empty cylinders, while in arrangements II and III, cylinders containing plants were selected. In experiments IV-1, IV-2, and IV-3, three isogenic lines of J⫹and J⫺were used to test aphid attraction to PCV-1-infected or virus-free plants. A similar pattern was observed in all three lines, and aphids demonstrated their preference for virus-free plants (J⫺). (b) The 95% confidence intervals for the probability of selecting the empty cylinder in arrangements I to III were 0.46 to 0.57 (I, empty versus empty); 0.23 to 0.34 (II, empty versus J⫹); and 0.22 to 0.33 (III, empty versus J⫺). The 95% confidence intervals for selecting J⫹over J⫺plants were 0.36 to 0.49 (IV-1, J1⫹versus J1⫺); 0.37 to 0.49 (IV-2, J2⫹ versus J2⫺); and 0.31 to 0.41 (IV-3, J3⫹versus J3⫺).
on November 6, 2019 by guest
http://jvi.asm.org/
[image:5.585.82.325.70.408.2]J⫺than on J⫹plants. Thus, PCV-1 infection protects jalapeño plants from increas-ing aphid populations.
DISCUSSION
Behavioral responses that impact insect dispersal are important because of their effect on viral disease spread. Several studies have shown that acute viruses manipulate their host volatiles so that infected plants are more enticing to vectors, in spite of the plants’ small size and the negative effects on apparent plant quality. Virus-induced changes are in favor of virus transmission, attracting aphids to feed on plants and acquire the virus for its dispersal (12, 13, 17, 22). These virus-induced changes in vector behavior are adaptive and vary among viruses, hosts, and vectors (23). Here, we showed that partitivirus infection also influences aphid behavior (Fig. 2, IV-1, IV-2, and IV-3), but this interaction is in stark contrast to insect-plant interactions in acute virus infection; that is, the presence of this partitivirus leads to deterrence of aphids and protection of
FIG 3Aphid attraction to volatiles of mock- or CMV-infected J⫹and J⫺plants. Pairwise preference experiments were as follows: (V) J⫺mock versus J⫺CMV; (VI) J⫺mock versus J⫹CMV; (VII) J⫹mock versus J⫹CMV; (VIII) J⫹M versus J⫺CMV; and (IX) J⫹CMV versus J⫺CMV. (a) Numbers represent the mean percentages of aphids in collection cages at the end of each treatment, and pairs involved in each choice test are shown horizontally. In arrangements V and VI, J⫺mock volatiles were less attractive to aphids than were those of J⫺CMV and J⫹CMV plants. In arrangements VII and VIII, J⫹M volatiles were also less appealing to aphids than those of J⫹CMV and J⫺CMV plants. In arrangement IX, aphids preferentially chose volatiles of J⫺CMV over those of J⫹CMV. (b) The 95% confidence intervals for the probability of selecting J⫹and J⫺mock in arrangements V to VIII were 0.32 to 0.0.41 (V, J⫺mock versus J⫺CMV); 0.30 to 0.40 (VI, J⫺mock versus J⫹CMV); 0.32 to 0.41 (VII, J⫹mock versus J⫹CMV); and 0.27 to 0.36 (VIII, J⫹mock versus J⫺CMV). The 95% confidence interval for choosing J⫹CMV was 0.32 to 0.0.41 (IX, J⫹CMV versus J⫺CMV).
on November 6, 2019 by guest
http://jvi.asm.org/
[image:6.585.84.327.71.442.2]plants from the vector of acute viruses, as well as from the damage of aphid herbivory itself. Based on these results, it seems that during thousands of years of virus replication in the plant host, the virus-plant interaction has evolved to be beneficial for the plants by discouraging the vector of acute viruses.
The acute plant virus CMV can manipulate its host’s volatile emissions to attract aphid vectors and facilitate its transmission, although these interactions are complex and dependent on specific relationships between virus strains and host plants. We inoculated J⫹and J⫺plants with CMV; volatiles of J⫹and J⫺CMV-infected plants were more attractive to aphids than those of J⫹and J⫺mock-inoculated plants (Fig. 3, V, VI, VII, and VIII). This is consistent with other studies showing that the VOC emissions from CMV-infected plants are more attractive to aphids than those from uninfected plants (22, 23). PCV-1 did not counter the effect of CMV on the plant VOCs, perhaps because of a strong elevation of VOCs by CMV. However, in pairwise prefer-ence tests between J⫹CMV- and J⫺CMV-infected plants, aphids preferred the J⫺CMV volatile blend (Fig. 3). Therefore, the presence of PCV-1 influenced aphid choice when both plants were CMV infected. This outcome verified our hypothesis that partitivirus infection decreases the likelihood of acute virus transmission, even in the presence of an acute virus.
Aphid reproduction on J⫹and J⫺plants was measured as an indicator of the effect of PCV-1 on host quality for aphids. Aphid reproduction on J⫹plants was more than 2-fold lower than that on J⫺plants (Fig. 4). There are several factors that can affect aphid reproduction. Turgor pressure can affect aphid feeding and therefore
[image:7.585.87.326.71.240.2]reproduc-FIG 4Aphid reproduction on plants with and without PCV-1 tested on three isogenic lines of jalapeño plants. Reproduction was calculated by dividing the number of offspring by the number of adults on each clip cage after 14 days. Results are shown as averages⫾standard error (SE) for 12 clip cages per treatment. Numbers of aphids and fecundity measures are shown in Table 2. P values: line 1, 2.485⫻10⫺6; line 2, 5.812⫻10⫺5; and line 3, 1.928⫻10⫺6. Statistical analysis indicated that the effect of the virus on aphid reproduction was significant (Pvalue⫽2.6⫻10⫺10). Asterisks indicate significant differences between control J⫹and J⫺plants (***,P⬍0.001).
TABLE 2Numbers of adults and offspring and aphid fecundity for three isogenic lines of jalapeño infected with PCV-1 (J⫹) and virus-free (J⫺) plants
Line
No. of aphids in:
Jⴙplants Jⴚplants
Adultsa Offspringb Fecundityc Adultsa Offspringb Fecundityc
1 74 506 6.83 71 913 12.86
2 69 655 9.49 71 1,490 20.98
3 63 586 9.30 61 1,326 21.74
aNumber of adults in clip cages 14 days after placement.
bNumber of offspring found in clip cages. In 14 days, only one generation of aphids occurs.
cFecundity is a measure of the ratio of offspring to adult aphids, as described in Materials and Methods.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.42.371.639.712.2]tion (33). However, in this study we minimized turgor pressure effects by watering plants regularly and similarly. Host plant quality during aphid growth and development (both nymph and adult stages) is another key factor of aphid reproduction (for a review, see reference 34). In poor host quality conditions, aphids resorbed their eggs for their own survival (35, 36). Nitrogen also plays an important role in plant quality and impacts aphid reproduction. Early in the growing season, phloem amino acid content is high, and aphid reproduction is higher than that when leaves mature and phloem amino acid levels drop (37–40).
Virus infections cause biochemical and physical changes in the host and conse-quently affect the reproduction of aphids. The populations of the aphids Schizaphis graminum,Rhopalosiphum padi, andSitobion avenaeincreased on BYDV-infected plants in comparison with those on uninfected plants (11, 41, 42). Moreover, aphid reproduc-tion was higher on wheat cultivars sensitive to BYDV than on a BYDV-tolerant wheat cultivar (42). Aphid population growth was correlated with the virus-vector relationship. The acquisition of BYDV requires continual aphid feeding on the phloem of infected plants. Thus, BYDV-induced changes in plants encourage sustained feeding and lead to aphid settling and rapid population growth, followed by dispersal of viruliferous aphids (11–14). Increased carbohydrate and amino acid contents of infected plants can lead to a higher reproduction rate in the vector. Total amino acid content was increased in spring wheat after infection with BYDV (43). Also, BYDV-infected plants had higher carbohydrate contents than healthy plants (44). On the other hand, CMV, which is transmitted in a nonpersistent manner, induced changes in the concentrations of carbohydrates and free amino acids in the plant tissue that lead to reduction of plant quality for aphids. When plant quality and palatability are reduced, aphids are dispersed quickly and efficiently transmit CMV to a healthy plant (6, 16, 21, 22).
The phytohormone ethylene is involved in the regulation of host defense responses against herbivores, necrotrophic pathogens, and nonpathogenic microbes (45, 46). In Turnip mosaic virus, the expression of nuclear inclusion a protease (NIa-Pro) is involved in decreasing host defenses and increasing vector reproduction. NIa-Pro is responsible for disruption of ethylene responses in plants and inhibition of aphid-induced callose production (47, 48). It is possible that PCV-1 infection directly or indirectly (by changing the plant microbiome) induces callose deposition, which is an important plant defense against aphid feeding, finally decreasing aphid reproduction on infected jalapeño plants.
Hence, while acute plant viruses manipulate the behavior of aphid vectors in their favor, enhancing their transmission, PCV-1, a persistent plant virus that is not horizon-tally transmitted, protects its plant host from aphids by reducing the attractiveness of the plant and the quality of the plant as an aphid host. This has the secondary effect of protecting the plants from aphid herbivory, which can also cause significant damage.
MATERIALS AND METHODS
Plants and insects.Three isogenic lines of jalapeño peppers (Capsicum annuum) with and without PCV-1 (denoted as J1⫹/J⫺, J2⫹/J2⫺, and J3⫹/J3⫺, respectively) were obtained from Rodrigo Valverde (Department of Plant Pathology and Crop Physiology, Louisiana State University). Plants were sown in 10-cm plastic pots containing Sun-Gro horticulture soil and grown in an insect-free environmental room at 24°C under fluorescent light (a ratio of 16:8 for the light/dark photoperiod).
A colony ofM. persicaewas obtained from Gary Thompson (Penn State University) and maintained on turnips (Brassica rapa). To produce sufficient aphids for experiments, the aphid colony was transferred to fresh turnip plants, and colonies were maintained with a natural photoperiod at 25°C. Aphid colonies were reestablished by moving two aphid-infested leaves to a new turnip plant every 10 to 15 days.
CMV inoculation.Two-week-old pepper plants were kept in the dark for 24 h and then mechanically inoculated using purified virus particles of Fny-CMV (49) diluted in 50 mM NaHPO4(pH 7.0). Leaves were dusted with carborundum to increase inoculation efficiency, followed by rubbing with acid-etched glass. Mock inoculation employed the same protocol, but with buffer only. Plants were used for aphid preference experiments 20 days after inoculation, when infected seedlings showed symptoms of virus infection.
Aphid preference for volatiles of plants with or without PCV-1.Aphid preference tests of plant volatile cues were performed using the experimental setup shown in Fig. 1a. Two cylinders (13⫻28 cm) containing different treatments were connected to an aphid box (11⫻17 cm) via two aphid collection cages (2.5⫻4 cm). The aphid box was covered to eliminate any visual cues. Each collection cage had one
on November 6, 2019 by guest
http://jvi.asm.org/
layer of extrafine mesh at the bottom to allow aphids to respond to VOCs without any contact cues and to trap aphids in the cage.
For the pairwise preference tests, 50 wingless nonviruliferous 3- to 5-day-old aphids were collected using a fine brush and starved for 1 h, after which they were transferred to the middle of the aphid box (Fig. 1a). The number of aphids in each collection cage was recorded 45 min after releasing the aphids. The percentage of the aphids that made a choice was calculated for each treatment. Pairwise preference tests were performed in the following three different control arrangements: (I) empty versus empty, (II) J⫹versus empty, and (III) J⫺versus empty. Aphid preferences for three isogenic lines of J⫹and J⫺were tested: IV-1, J1⫹versus J1⫺; IV-2, J2⫹versus J2⫺; and IV-3, J3⫹versus J3⫺. Plants used in these tests were matched for size and development stage (four- to six-leaf stage). The preference experiment for each plant pair was repeated at least twice, and each pairwise arrangement was done 12 times using four to six plants. All experiments were done at 25°C in daylight to ensure that plants released a consistent volatile blend.
Aphid preference for volatiles of plants with or without PCV-1 and CMV.To investigate aphid preference for volatiles of jalapeño plants in the presence of CMV (an acute virus) and PCV-1 (a persistent virus), the same experimental setup as above was used (Fig. 1a). The pairwise preference tests were performed with the following five different plant arrangements: (V) J⫺mock versus J⫺CMV; (VI) J⫺ mock versus J⫹CMV; (VII) J⫹mock versus J⫹CMV; (VIII) J⫹mock versus J⫺CMV; and (IX) J⫺CMV versus J⫹CMV. All experimental conditions were as above.
Aphid reproduction on plants with or without PCV-1. Experiments were done using 6- to 8-leaf-stage jalapeño plants in a pesticide-free greenhouse at 23 to 26°C supplemented with light for a 16-h day length. Eight adult wingless aphids were confined in a small clip cage (2.5⫻4 cm) and placed on a caged plant. Three clip cages were randomly attached to each plant, and for each line, four plants were used (Fig. 1b). The adult aphids were left to reproduce inside the cage for 24 h, and then adults were removed and offspring were maintained in clip cages to mature and reproduce. During 14 days, offspring molting to adults, reproducing, and producing new offspring were monitored in the clip cages. At day 14, the numbers of offspring and adults in each individual clip cage were recorded, and aphid reproduction was calculated by dividing the number of offspring produced by the number of adult aphids. This experiment was replicated 12 times for each jalapeño line, and in total, reproduction was measured for more than 60 aphids for each treatment.
aphid reproduction⫽no. of offspring no. of adults
Statistical analysis.Statistical analysis was performed using R version 3.3.3. A general linear model (GLM) with binomial error was used to analyze aphid preference data for all pairwise experiments, estimating the proportion of aphids that moved to the preferred option and 95% confidence intervals for each pairwise test. For statistical analysis of aphid reproduction, the lme4 package in R was used, and two models were defined. The null model for aphid reproduction included different plants and lines assumed as random effects. A second model included the presence of PCV-1 considered a fixed effect, with the same random effects as in the null model. A likelihood ratio test of the two models was performed (using analysis of variance [ANOVA] in R) to test the significance of the fixed effect.
ACKNOWLEDGMENTS
We thank Rodrigo Valverde (Louisiana State University, LA, USA) and Gary Thomp-son (Penn State University, PA, USA) for providing jalapeño seeds and the aphid colony, respectively, and Juan Jose Lopez-Moya (Center for Research in Agricultural Genomics, Spain) for technical help with the aphid setup. We also specially thank Andrew G. Stephenson and Heike Betz for their helpful discussions.
This work was funded by the Pennsylvania State University College of Agricultural Sciences and the Huck Institute for Life Sciences.
REFERENCES
1. Roossinck MJ. 2010. Lifestyles of plant viruses. Philos Trans R Soc Lond B Biol Sci 365:1899 –1905.https://doi.org/10.1098/rstb.2010.0057. 2. Safari M, Roossinck MJ. 2014. How does the genome structure and
lifestyle of a virus affect its population variation? Curr Opin Virol 9:39 – 44.https://doi.org/10.1016/j.coviro.2014.09.004.
3. Roossinck MJ. 2013. Plant virus ecology. PLoS Pathog 9:e1003304.
https://doi.org/10.1371/journal.ppat.1003304.
4. Blanc S. 2007. Virus transmission– getting in and out, p 1–28.InWaigman E, Heinlein M (ed), Viral transport in plants. Springer-Verlag, Berlin, Germany.
5. Boccardo G, Lisa V, Luisoni E, Milne RG. 1987. Cryptic plant viruses. Adv Virus Res 32:171–214.https://doi.org/10.1016/S0065-3527(08)60477-7. 6. Whitfield AE, Falk BW, Rotenberg D. 2015. Insect vector-mediated
trans-mission of plant viruses. Virology 479-480:289 –290.
7. Ng JCK, Perry KL. 2004. Transmission of plant viruses by aphid vectors.
Mol Plant Pathol 5:505–511.https://doi.org/10.1111/j.1364-3703.2004 .00240.x.
8. Powell G, Tosh CR, Hardie J. 2006. Host plant selection by aphids: behavioral, evolutionary, and applied perspectives. Annu Rev Entomol 51:309 –330.https://doi.org/10.1146/annurev.ento.51.110104.151107. 9. Kennedy JS. 1951. Benefits to aphids from feeding on galled and
virus-infected plants. Nature 168:825– 826.https://doi.org/10.1038/168825a0. 10. Jiménez-Martínez ES, Bosque-Pérez NA, Berger PH, Zemetra RS. 2004. Life history of the Bird Cherry-oat aphid, Rhopalosiphum padi (Homoptera: Aphididae), on transgenic and untransformed wheat chal-lenged with Barley yellow dwarf virus. J Econ Entomol 97:203–212.
https://doi.org/10.1093/jee/97.2.203.
11. Montllor CB, Gildow FE. 1986. Feeding responses of two grain aphids to barley yellow dwarf virus-infected oats. Entomol Exp Appl 42:63– 69.
https://doi.org/10.1111/j.1570-7458.1986.tb02188.x.
on November 6, 2019 by guest
http://jvi.asm.org/
12. Eigenbrode SD, Ding H, Shiel P, Berger PH. 2002. Volatiles from potato plants infected with potato leafroll virus attract and arrest the virus vector Myzus persicae (Homoptera: Aphididae. Proc R Soc B 269: 455– 460.https://doi.org/10.1098/rspb.2001.1909.
13. Jiménez-Martínez ES, Bosque-Pérez NA, Berger PH, Zemetra RS, Ding H, Eigenbrode SD. 2004. Volatile cues influence the response of Rhopalo-siphum padi (Homoptera: Aphididae) to Barley yellow dwarf virus-infected transgenic and untransformed wheat. Environ Entomol 33: 1207–1216.https://doi.org/10.1603/0046-225X-33.5.1207.
14. Srinivasan R, Alvarez JM, Eigenbrode SD, Bosque-Pérez NA. 2006. Influ-ence of hairy nightshadeSolanum sarrachoides(Sendtner) andPotato leafroll virus(Luteoviridae: Polerovirus) on the host preference ofMyzus persicae(Sulzer) (Homoptera: Aphididae). Environ Entomol 35:546 –553.
https://doi.org/10.1603/0046-225X-35.2.546.
15. Gray S, Gildow FE. 2003. Luteovirus-aphid interactions. Annu Rev Phy-topathol 41:539 –566.https://doi.org/10.1146/annurev.phyto.41.012203 .105815.
16. Blanc S, Drucker M, Uzest M. 2014. Localizing viruses in their insect vectors. Annu Rev Phytopathol 52:403– 425. https://doi.org/10.1146/ annurev-phyto-102313-045920.
17. Medina-Ortega KJ, Bosque-Pérez NA, Ngumbi E, Jiménez-Martínez ES, Eigenbrode SD. 2009.Rhopalosiphum padi(Hemiptera: Aphididae) re-sponses to volatile cues from Barley yellow dwarf virus-infected wheat. Environ Entomol 38:836 – 845.https://doi.org/10.1603/022.038.0337. 18. Ingwell LL, Eigenbrode SD, Bosque-Pérez NA. 2012. Plant viruses alter
insect behavior to enhance their spread. Sci Rep 2:578.https://doi.org/ 10.1038/srep00578.
19. Fereres A, Kampmeier GE, Irwin ME. 1999. Aphid attraction and prefer-ence for soybean and pepper plants infected with Potyviridae. Ann Entomol Soc Am 94:542–548.https://doi.org/10.1093/aesa/92.4.542. 20. Edwardson JR, Christie RG. 1991. Cucumoviruses, p 293–319.InCRC
Handbook of viruses infecting legumes. CRC Press, Boca Raton, FL. 21. Martín B, Collar JL, Tjallingii WF, Fereres A. 1997. Intracellular ingestion
and salivation by aphids may cause the acquisition and inoculation of non-persistently transmitted plant viruses. J Gen Virol 78:2701–2705.
https://doi.org/10.1099/0022-1317-78-10-2701.
22. Mauck KE, De Moraes CM, Mescher MC. 2010. Deceptive chemical signals induced by a plant virus attract insect vectors to inferior hosts. Proc Natl Acad Sci U S A 107:3600 –3605. https://doi.org/10.1073/pnas .0907191107.
23. Mauck KE, De Moraes CM, Mescher MC. 2014. Evidence of local adapta-tion in plant virus effects on host-vector interacadapta-tions. Integr Comp Biol 54:193–209.https://doi.org/10.1093/icb/icu012.
24. Tungadi T, Groen SC, Murphy AM, Pate AE, Iqbal J, Bruce TJA, Cunniffe NJ, Carr JP. 2017. Cucumber mosaic virus and its 2b protein alter emission of host volatile organic compounds but not aphid vector settling in tobacco. Virol J 14:91.https://doi.org/10.1186/s12985-017 -0754-0.
25. Roossinck MJ. 2012. Plant virus metagenomics: biodiversity and ecology. Annu Rev Genet 46:357–367.
26. Roossinck MJ. 2014. Metagenomics of plant and fungal viruses reveals an abundance of persistent lifestyles. Front Microbiol 5:767.https://doi .org/10.3389/fmicb.2014.00767.
27. Shiba K, Hatta C, Sasai S, Tojo M, Ohki ST, Mochizuki T. 2018. Genome sequence of a novel partitivirus identified from the oomycetePythium nunn. Arch Virol 163:2561–2563. https://doi.org/10.1007/s00705-018 -3880-0.
28. Nibert ML, Woods KM, Upton SJ, Ghabrial SA. 2009.Cryspovirus: a new genus of protozoan viruses in the familyPartitiviridae. Arch Virol 154: 1959 –1965.https://doi.org/10.1007/s00705-009-0513-7.
29. Szego˝ A, Tóth EK, Potyondi L, Lukás N. 2005. Detection of high molecular weight dsRNA persisting inDianthusspecies. Acta Biologica Szegedien-sis 49:17–19.
30. Valverde RA, Gutierrez DL. 2008. Molecular and biological properties of a putative partitivirus from jalapeño pepper (Capsicum annuumL.). Rev Mex Fitopatol 26:1– 6.
31. Valverde RA, Fontenot JF. 1991. Variation in double-stranded ribonucleic acid among pepper cultivars. J Amer Soc Hort Sci 116:903–905.https:// doi.org/10.21273/JASHS.116.5.903.
32. Sabanadzovic S, Valverde RA. 2011. Properties and detection of two cryptoviruses from pepper (Capsicum annuum). Virus Genes 43:307–312.
https://doi.org/10.1007/s11262-011-0634-4.
33. Wearing CH. 1967. Studies on the relations of insect and host plant: II. Effects of water stress in host plants on the fecundity ofMyzus persicae (Sulz.) andBrevicoryne brassicae(L.). Nature 213:1052–1053.https://doi .org/10.1038/2131052a0.
34. Awmack CS, Leather SR. 2002. Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol 47:817– 844.https://doi.org/10 .1146/annurev.ento.47.091201.145300.
35. Leathers V, Tanguay R, Kobayashi M, Gallie DR. 1993. A phylogenetically conserved sequence within viral 3=untranslated RNA pseudoknots reg-ulates translation. Mol Cell Biol 13:5331–5347.https://doi.org/10.1128/ MCB.13.9.5331.
36. Sequeria R, Dixon AFG. 1996. Life history responses to host quality changes and competition in the Turkey-oak aphid,Myzocallis boerneri (Hemiptera: Sternorrhyncha: Callaphididae). Eur J Entomol 93:53–58. 37. Dixon AFG. 1970. Quality and availability of food for a sycamore aphid
population, p 271–287.InWatson A (ed), Animal populations in relation to their food resources. Blackwell Science, Oxford, UK.
38. Leather SR, Wade FA, Godfray HCJ. 2005. Plant quality, progeny se-quence, and the sex ratio of the sycamore aphid,Drepanosiphum pla-tanoidis. Entomol Exp Appl 115:311–321.https://doi.org/10.1111/j.1570 -7458.2005.00257.x.
39. Weibull J. 1987. Seasonal changes in the free amino acids of oat and barley phloem sap in relation to plant growth state and growth of Rhopalosiphum padi. Ann Applied Biology 111:729 –737.https://doi.org/ 10.1111/j.1744-7348.1987.tb02031.x.
40. Leather SR, Dixon AFG. 1981. The effect of cereal growth stage and feeding site on the reproductive activity of the bird-cherry aphid, Rho-palosiphum padi. Ann Applied Biology 97:135–141. https://doi.org/10 .1111/j.1744-7348.1981.tb03006.x.
41. Araya JE, Foster JE. 1987. Laboratory study on the effects of barley yellow dwarf virus on the life cycle ofRhopalosiphum padi(L.). Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 94:578 –583.
42. Fereres A, Lister RM, Araya JE, Foster JE. 1989. Development and repro-duction of the English grain aphid (Homoptera: Aphididae) on wheat cultivars infected with barley yellow dwarf virus. Environ Entomol 18: 388 –393.https://doi.org/10.1093/ee/18.3.388.
43. Ajayi O. 1986. The effect of barley yellow dwarf virus on the amino acid composition of spring wheat. Ann Applied Biology 108:145–149.https:// doi.org/10.1111/j.1744-7348.1986.tb01975.x.
44. Fereres A, Araya JE, Housley TL, Foster JE. 1990. Carbohydrate compo-sition of wheat infected with barley yellow dwarf virus. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 97:600 – 608.
45. Erb M, Meldau S, Howe GA. 2012. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci 17:250 –259.https://doi.org/10 .1016/j.tplants.2012.01.003.
46. Howe GA, Jander G. 2008. Plant immunity to insect herbivores. Annu Rev Plant Biol 59:41– 66.https://doi.org/10.1146/annurev.arplant.59.032607 .092825.
47. Casteel CL, De Alwis M, Bak A, Dong H, Whitham SA, Jander G. 2015. Disruption of ethylene responses byTurnip mosaic virusmediates sup-pression of plant defense against the green peach aphid vector. Plant Physiol 169:209 –218.https://doi.org/10.1104/pp.15.00332.
48. Casteel CL, Yang C, Nanduri AC, De Jong HN, Whitham SA, Jander G. 2014. The NIa-Pro protein ofTurnip mosaic virusimproves growth and reproduction of the aphid vector,Myzus persicae(green peach aphid). Plant J 77:653– 663.https://doi.org/10.1111/tpj.12417.
49. Roossinck MJ, Palukaitis P. 1990. Rapid induction and severity of symp-toms in zucchini squash (Cucurbita pepo) map to RNA 1 of cucumber mosaic virus. MPMI 3:188 –192.https://doi.org/10.1094/MPMI-3-188.