0022-538X/80/03-1215/06$02.00/0
Viable Deletion
Mutant in the Medium and
Large
T-Antigen-Coding
Sequences
of the
Polyoma
Virus Genome
MARY M.BENDIG,tTHRESIATHOMAS,ANDWILLIAMR. FOLK*
Departmentof Biological Chemistry, The University of Michigan, Ann Arbor, Michigan 48109
Apolyomavirusmutantthat maps intheearly regionbetweenthe known
hr-tandts-amutantshasbeen isolated. Its66-base-pair deletion resultsinstructural
changes in both medium andlarge T-antigensbutcauses nosubstantial alterations
inviralreplicationorcelltransformation.
Theearly region of thepolyomavirusgenome
contains two known complementation groups
defined by mutants, the
host-range-nontrans-forming (hr-t) mutants and the
temperature-sensitivets-a mutants.Thehr-tmutantsmapin
the proximal portion of the early region. They
induce analteredhost rangeforlyticgrowthand
are defective in virus-induced cell transforma-tion andtumorigenesis(3, 8). Thets-amutants
mapin thedistalportionof theearly regionand
aredefective,atthenonpermissivetemperature,
inDNAreplication, control of viral RNA
tran-scription, and virus-inducedcelltransformation
(6,7, 9, 10,25,26).Inthisreportwedescribe the
isolation andpartial characterization ofa
dele-tionmutant, mutant45,thatmaps intheearly
region between the hr-t andts-amutants.
Mutant45 was detected while screening for
mutants with deletions at the endo BglI site
neartheorigin ofreplication.Theisolation
pro-cedure has beendescribed elsewhere(1).Inbrief,
form I DNA from unmutagenized Pasadena large-plaque polyoma virus was digested with
endo BglI (which cuts polyoma DNA at one
site) and then electrophoresed through a 1%
agarose gel.Theresultingform IIIlinearDNA
was extracted from the gel and digested first withSi nuclease (14 U ofenzyme per 2.5
,jg
ofDNAfor30minat
25°0)
and then withexonu-clease m (0.4 U ofenzyme per 1.2 ,ugof DNA
for 60 min at 3700). Secondary whole mouse
embryo cells were infected with the digested DNA, and fromtheresulting virus stock
individ-ual plaques werepicked and tested for
altera-tions in their endo HpaH digestion patterns.
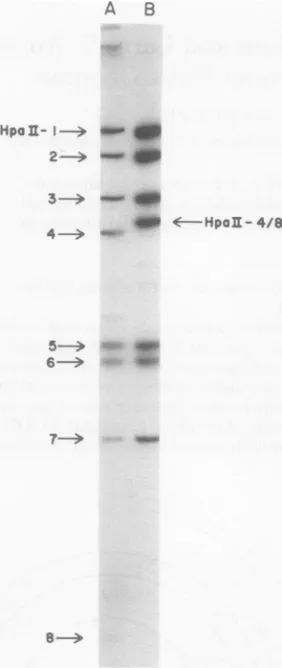
Mutant45gave awild-typesizeHpaII fragment
5 (HpaII-5contains the endoBglIsite, Fig. 1)
but wasmissingwild-typesizeHpaII fragments
4and 8. Anewfragment slightlysmallerin size
than that expected of a composite HpaII-4/8
fragmentwasobserved (Fig. 2), thussuggesting
that mutant 45 has adeletionremoving the endo
HpaII-4/8 site. The altered virus was plaque
t Present address: Institut fir Molekularbiologie H der Universitit Ziirich, 8093 Ziirich, Switzerland.
purifiedonce morebeforebeing further
charac-terized.
Digestion ofmutant45DNAwith
endonucle-asesAluI (4) and HaellI (12) indicated that a
60- to 100-base-pair deletion had occurred
be-tween(butnotincluding)theAvaI site and the
PvuII site locatedoneitherside of the
Hpall-4/
8 junction. Accordingly, mutant 45 DNA wascleaved with endonucleaseAvaIand incubated
FIG. 1. Map of the polyoma virus genome. The circular map is divided into the the eight HpaII
fragments (12). The single endo BglIsite and the
endoAvaI and endo PvuIIsites mapping near the
endoHpaII-4/8siteareindicated by arrows (11). On
the outeredge of the map, dotted lines define the locationof the hr-tmutants(14), the ts-a mutants (25), andmutant45;solidlinesdefine thepredicted coding regionsfor the small, medium, and large T-antigens (11, 30; R. Kamen, personal communication). On the inside ofthe map, theorigin of replication and the earlyandlate regions of the genome are indicated
(19, 21).
1215
I -T d-T
on November 10, 2019 by guest
http://jvi.asm.org/
[image:1.504.258.445.308.532.2]A B MUTANT 45
G A+G C+T C A+C
Hpal1- I-- ow
4
2 * _
3-->-M.w
- -HpolI-4/8
4
6---6.... .. i..0
C
Ac
A G
8-*
FIG. 2. Autoradiogramofpolyoma DNA digested
with endoHpaII. EndoHpaIIdigests of[32P/DNA from the wild type(slotA) andmutant 45 (slotB)
wereanalyzed byelectrophoresisina6%
polyacryl-amidegel(2).Arrowsontheleftpointtothewild-type
HpaIIfragments. Thearrow onthe rightpoints to
thecomposite HpaII-4/8fragment foundinmutant
45.
with[y-32P]ATPandpolynucleotide kinase and
sequenced by the chemical degradation
proce-dure ofMaxamand Gilbert(24).Comparisonof
its sequence with that of the wild-type parent
revealed that a 66-nucleotide deletion had
oc-curred between nucleotides 1090 and 1157
(Friedmannnumberingsystem[11]; Fig. 3).
Be-cause the basesdeletedare amultipleofthree,
thereadingframe isnotaltered.
Inspectionof the sequence in thisregion
sug-gests apossiblereasonfor the susceptibilityof
polyomaDNAtoS1nuclease. Between
nucleo-tides 1132and1183 occursapartialpalindrome
which could formahairpin (Fig. 4).Thismight
providearegionofsingle-strandedDNA
suscep-tibletoS1nuclease.
Theearly regionof thepolyomavirus genome
GG
TC AA
GG G
Tc
TA AA
Gc
FIG. 3. AutoradiogramofpolyomaDNAsubjected
tochemicaldegradationandgelelectrophoresis. Pol-yomaDNAs(mutant45anditswild-type parent)were
cloned in Escherichia coliwithpBR322(Bendiget
al.,manuscriptinpreparation).EndonucleaseAvaI
digestionofthepurifiedchimericDNAgenerateda
fragment containing the majority of the polyoma
earlyregion. Thiswaslabeled with[y-32P]ATPand
polynucleotidekinase andthendigestedwith
endo-nucleaseEcoRI. Theasymmetrically labeled DNAs
wereseparatedandsubjectedtochemical
degrada-tion (10min). Theproducts wereseparatedon 15% polyacrylamidegels.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.504.298.427.62.522.2] [image:2.504.90.231.65.399.2]I06 SMutant 45
CCT TGT GTT GCT GAG CCC GAT GACAGCATATCCI CCCC CCC AGA ACT CCT GTA TCCAGA
LargeT-antigen PRO CYS-VAL ALA-GLU PRO ASP ASP-SER*ILE-SER-PRO-ASP-PROPRO-ARG-THR-PRO-VAL-SER-ARG MiddleT-antigen ILE-LEU VAL LEU LEU SER-PRO MET-THR-ALA-TYR-PRO ARGTHR-PRO-PRO-GLU-LEU-LEU-TYR-PRO GLU
1194
AAS CGA CCA AGACC8ICGGA GCCACTGOA GGA GGA GGAGGA GGA GTA
LYS ARG-PRO-ARG-PRO ALAGLY*ALA-THR-GLY GLY GLY*GLYGLY*GLY VAL
[image:3.504.45.442.61.162.2]SER-ASP-GLN-ASP-GLNMLEU-GLU-PRO-LEU*GLU-GLU*GLU-GLU-GLU-GLUTYR
FIG. 4. Sequence of the region of wild-type polyoma DNA including themutant45deletion. The nucleotide sequenceofthe parentofmutant45wasincompleteagreement with that determinedbyFriedmannetal.(11)
andSoeda etal. (30).Arrowspointto theregion deleted in mutant45. Underlined sequences indicatea
partial palindrome which couldformanintrastrandloop. The HpaII-4/8 and PvuII sites indicated inFig.1 areenclosed in boxes. Amino acid sequencesformiddle andlargeT-antigensareindicated(11, 30).
codes for at least three tumor antigens of
ap-proximately95,000(95K), 60K, and22K molec-ularweight (15,16,18,29).In ts-amutants,large
T-antigen (95K) isthermolabile, whereas the60
and 22KT-antigens remain unaffected (16, 18,
26,29).Incontrast,the hr-t mutationsaffect the
60and22Kantigens butnotthe 95KT-antigen
(16, 17, 27, 29). To determine whether mutant
45synthesizedearly viral proteins of normal size
andamounts, 3T6 cellswere infected, the
pro-teinswerelabeled with
['S]methionine,
and the T-antigens were immunoprecipitated andana-lyzed by sodiumdodecylsulfate-polyacrylamide
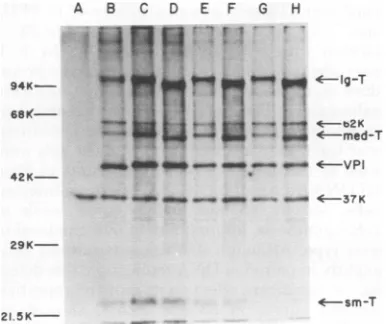
gel electrophoresis. Asseen inFig. 5,largeand
medium T-antigens from mutant 45-infected cells migrate ahead of those from wild type.
Small T-antigen, however, is thesamein both
mutant 45-andwild-type-infectedcells.(The37
and62Kprotein bandsarebelievedtobe
anal-ogous tothe37and 55Kspecies that Hutchinson
et al.
[16]
foundbytryptic peptide analysistobe unrelatedtosmall,medium,orlarge T-anti-gen.) Theprotein band labeled VP1 corresponds in sizetothemajorpolyomaviruscapsid protein, VP1.Cells infected wthasmall-plaquestrain of polyoma virus have an altered VP1 (Fig. 5B).
Size estimates of the mutant 45T-antigens
in-dicate that large T-antigen is 3,000 to 4,000
daltonssmallerthanwildtype andthat medium T-antigen is approximately 2,000 daltons smaller. The size of the deletionin mutant 45 DNA predicts a 2,600-dalton reduction in
pro-tein molecular mass. Chasing the 3S-labeling
medium for 6 or 12 .h with medium containing
an excess ofunlabeled methionine revealed no
significant differences between mutant 45 and
wild-type virus in the amounts andstability of theT-antigens (Fig.5).
Schaffhausen et al. have observed in vivo
phosphorylation of large and medium T-anti-gensandnophosphorylationofsmallT-antigen,
when infected cells are incubated with
[32P]-A B C D E F G H
t1..u K -- tI - .
94K- <*-t-- r Ig-T
68
K-342K-
_*
r * X
=med-T
00 w#m moI-E-VPIW-f
-
42K-____ _-__ _ 4 _-<37K
29K-*Qs ~~~~~sm-T
21.5K _
FIG. 5. Fluorogram of T-antigens labeled with
[36S]methionine.Culturesof3T6 cellswere infected
with Crawford small-plaque polyoma virus
(multi-plicity of infection = 10). The infected cells were
incubated at 33°C for 43 h, labeled for 2 h with
[3Slmethionine (100,iCi/ml), and harvested bythe procedure of Hutchinson et al. (16). In pulse-chase
experiments,cellswerewashed and chased with
me-diumcontaining10times thenormal concentration
of methionine before harvesting.
Immunoprecipita-tionofradiolabeled cellextracts wascarriedoutby
amodifiedStaphylococcus proteinAprocedure (16).
Anti-polyomatumor serum waskindlyprovidedby T. Benjamin. The immuneprecipitateswere electropho-resed through adiscontinuous sodium dodecyl sul-fate-polyacrylamide gel (5 and 12% acrylamide) (20) whichwasthenstained andpreparedfor fluorogra-phy (5). Theproteinmolecularweight markers, phos-phorylase b (94K), bovineserumalbumin(68K), oval-bumin(43K),carbonicanhydrase (29K),andsoybean trypsin inhibitor (21.5K), are indicated on the left. Slot A is from mock-infected cells, slot B is from
Crawfordsmall-plaque-infectedcells, slots C, E, and
Garefromwild-type large-plaque-infected cells, and slotsD, F, andHarefrommutant45-infected cells. SamplesinslotsEand Fwerechasedfor6hbefore harvesting, and those in slots G and H were chased for12h.
33,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.253.446.233.395.2]1218
phosphate (27). To determine whether the
al-tered medium andlarge T-antigens frommutant
45 retain their sites for phosphorylation, 3T6
cells were infected and labeled wth
[32P]phos-phate, and the T-antigenswere immunoprecip-itated. Figure 6 shows thatin bothmutant 45
andwild-type viruses,large T-antigen is heavily phosphorylated, whereas medium T-antigen has
alow level ofphosphorylation.
Mutant 45 was tested for defects in known earlygene functions. As mentionedpreviously,
one class ofearly gene mutants, the ts-a
mu-tants, affect a viral function required for the
initiation of viral DNAreplication (9). The rel-ative amounts of viral DNA synthesis in cells infected with either mutant orwild-type virus
wasdetermined byinfecting duplicate plates of
confluent 3T6 cells at amultiplicity of10PFU/
cell. The infected cellswereincubatedat
330C,
labeled with [32P]phosphate from 12 to 36 h postinfection, and harvested via the Hirt
proce-dure.Samplesfrom the Hirtsupematants were
exhaustivelydigested with endo EcoRI and elec-trophoresedthrough1%agarosegels.
Densitom-etertracings ofautoradiogramsof thegelswere
used to calculate the relativeamounts of form
III DNAfrommutant 45-andwild-type-infected
cells. Mutant 45 had slightly lower levels of DNAsynthesis, approximately60% thelevel of
wild type. Although this suggests mutant 45is
slightly impaired in DNAreplication,this defect
hasnosignificanteffectonits growthproperties.
Growth curves at 33 and 39.5°C showed no
significant differences between mutant 45 and wild-type viruses(datanotshown).
Bothclasses ofpreviouslycharacterized early region mutants are defective in virus-induced
cell transformation(3,10).Mutant45 wastested for itsabilitytotransform BHK cellsorFisher
rat3T3 cells toanchorage independenceas
de-terminedby growth in softagar. From the
re-sults of thesetransfornationassays(Table 1),it
appearsthatmutant45transformsasefficiently
aswild-typevirus. Inaddition, preliminary
ex-periments indicate that mutant45 induces
an-giomatous lesions of the liver andlungsofSyrian
hamsters wheninjectedintonewborns.
Although mutant45 isstill abletocarry out
important earlygenefunctionssuch asthe
ini-tiationof viralDNAreplicationand the
trans-formation of
cells,
twoearlygeneproducts,me-dium andlargeT-antigens,have reduced
molec-ular weights. DNA sequencing of theearly
re-gion,mapping oftheearly mRNA's,andtryptic
peptide analysis of early gene products have permitted the identification of theapproximate
coding regionsfor the three T-antigens (11,30;
R. Kamen, personal communication). As
illus-trated in Fig. 1, the deletion inmutant 45lies
A B C
94K >|
~-Ig
-T
68K-:med-T
43K-..._
FIG. 6. Autoradiogram of T-antigens labeled with
[32Plphosphate. Culturesof 3T6 cells were infected withwild-typeandmutant 45viruses (multiplicity of
infection = 10). After 42 h at 33°C, the cells were washed twicewithphosphate-freemedium and then labeledfor4hwith[32P]phosphate(100,Ci/ml).Cell extracts wereprepared and immunoprecipitatedas describedfor35S-labeled cells. The
immunoprecipi-tates were analyzedon a 4 and8%discontinuous sodiumdodecylsulfate-polyacrylamide gel. Thegel
wasstained with Coomassiebrilliantblue,destained,
dried, and autoradiographed. Molecular weight
markers are indicated on the left. Slot A is from
mock-infectedcells,slot B isfromwild type, and slot Cisfrommutant 45.
within thecoding regions for medium andlarge T-antigens and thus would be expectedtoalter mediumand largeT-antigens butnot toaffect smallT-antigen.
The amino acidswithin middle andlarge
T-antigensthataredeleted in mutant45 areshown
in Fig. 4. The most noteworthy sequence of
aminoacids,astretch of sixglutamic acid
resi-dues in middle T-antigen, is just beyond the
limits ofthedeletion.
Severalother viablemutantsofpolyomavirus
with deletions in themiddleof theearlyregion
have recently been reported. Magnusson and
Berg have isolated several mutants with the
deletionsinHpaII-7or -8with lowered
capaci-ties fortransformingrat 1fibroblasts (23).The
precise locationofthese deletionsisnotknown,
however.Griffinand Maddock(13)haveisolated
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.294.439.64.318.2]TABLE 1. Transformationassays' Transformed
Expt Cell Virus MOI colonies/infected cells
1 BHK Mock 0 0/5.6x104
Wild type 21 73/5.6x104
Mutant 45 61 143/5.6x104
2 BHK Mock 0 0/4.0x104
Wild type 80 94/4.0x 104 Mutant45 70 56/4.0x 104
3 FR3T3 Mock 0 0/2.0x 104
Wild type 60 90/2.0x104
Mutant45 280 296/2.0x 104 aBaby
hamster
kidneycells (BHK-21/13from G. DiMayorca) orFisherrat 3T3celis (FR3T3 from R.G. Martin) (28) were mock infectedorinfected with
polyoma virus atthe indicatedmultiplicities (MOI).
Infected cells were plated in a layer of 0.33% agar mediumover abaselayerof 0.5% agar medium(22). After28daysat370Cfor BHK cellsor33°Cfor FR3T3 cells, large macroscopically visible colonies were scoredas transformed.
and characterizedtwomutants (dl-8anddl-23)
inthisregion.Thedl-8 mutant mapson oneside
ofmutant 45toward theorigin, andits
approxi-mately100-base-pairdeletionmayslightly
over-lapthe66-base-pair deletion in mutant 45. The
approximately 100-base-pair deletion in dl-23
maps onthe other side of mutant45, but there
is likely to belittle or no overlap between the
deletions in mutants dl-23 and 45. Together,
mutant45,dl-8anddl-23 delete about 250
con-tiguous base pairs (approximately 10% of the
early region). Like mutant45,bothdl-8 and -23
have truncated medium and large T-antigens
and normal-size small T-antigen; however,
un-like mutant 45, dl-8 and dl-23 exhibit altered
replication ortransformation properties. These
viabledeletion mutants constitute a new class of
mutants in theearly regionofthepolyomavirus
genome. Itis of interestthat thisregion shares
nohomologywiththat of simian virus40 orBK
virus (11, 30). It seemsthat their large
T-anti-gens are functional without such a sequence
component.
Wethank PatriciaVoilmerandRoseann White forhelp in preparing thesecondarywholemouseembryocells.Thegift of polyoma anti-T serum from T. Benjamin is gratefully acknowledged.
SupportwasprovidedbyPublicHealthServicegrantCA 13978from the NationalCancerInstitute, training grant GM 07135 toM.M.B.fromthe National Institute ofGeneral Med-icalSciences, andAmericanCancerSociety Faculty Research Award FRA152 toW.R.F.
LITERATURE CITED
1. Bendig, M. M., and W. R. Folk. 1979. Deletion mutants ofpolyomavirusdefining a nonessential region between theorigin ofreplicationand the initiation codon for early proteins. J. Virol. 32:530-535.
2. Bendig, M. M., W. R. Folk, and W. Gibson. 1979.
Polyoma viruses with mutations at endonuclease HindUI site 1: alterationsatthe COOH terminus ofVP1. J.Virol. 30:515-522.
3. Benjamin, T. L. 1970. Host range mutants ofpolyoma virus.Proc. Natl.Acad. Sci.U.S.A. 67:394-399. 4. Berkner, K.L., and W. R. Folk.1979.Amapof thesites
inthepolyoma genome cleaved by endonuclease AluI. Virology 92:682-696.
5. Bonner, W.M., and R. A.Laskey.1974. Afilm detection method for tritium-labeled and nucleic acids in poly-acrylamidegels. Eur. J.Biochem. 46:8388.
6. Cogen, B. 1978. Virus-specific early RNA in 3T6 cells infectedby a tsA mutant of polyoma virus. Virology 85: 222-230.
7. Eckhart, W.1969.Complementationandtransformation by temperature sensitive mutants of polyoma virus. Virology 38:120-125.
8. Feunteun, J., LSompayrac,M.Fluck,and T.
Ben-jamin.1976.Localization of genefunctions inpolyoma
virusDNA. Proc.Natl. Acad. Sci. U.S.A. 73:4169-4173. 9. Francke, B., and W. Eckhart. 1973. Polyoma gene functionrequiredfor viral DNAsynthesis.Virology55: 127-135.
10. Fried, M. 1965. Isolation of temperature-sensitive mu-tantsofpolyoma virus.Virology25:669-671. 11. Friedmann,T.,A.Esty,P.LaPorte,and P.Deininger.
1979. Thenucleotide sequence and genome organization of thepolyoma earlyregion:extensive nucleotide and aminoacidhomologywithSV40. Cell17:715-724. 12.Griffin, B. E.1977.Fine structure ofpolyomavirus DNA.
J. Mol.Biol.117:447-471.
13.Griffin, B.E., and C. Maddock. 1979. Newclasses of viabledeletion mutants in theearly region of polyoma virus. J.Virol. 31:645-656.
14.Hattori, J.,G.G.Carmichael,and T. L. Benjamin.
1979.DNA sequencealterations in hr-t deletion mu-tantsofpolyoma virus. Cell 16:505-513.
15.Hunter,T.,M. A.Hutchinson,andW.Eckhart. 1978. Translation ofpolyoma virus T-antigens in vitro. Proc. Natl. Acad.Sci. U.S.A.75:5917-5921.
16.Hutchinson, M. A., T.Hunter,and W.Eckhart. 1978. Characterization ofT-antigensinpolyoma-infectedand transformed cells. Cell 15:65-77.
17. Ito,Y.,J. R.Brocklehurst,N.Spurr,M.Griffiths,J.
Hurst,and M.Fried. 1977.Polyomavirus wild type andmutantT-antigens.Inst.Natl. Sante Rech. Med. 9: 145-152.
18. Ito, Y., N.Spurr,and R. Dulbecco.1977. Characteri-zation ofpolyoma virusT-antigen. Proc.Natl. Acad. Sci. U.S.A. 74:1259-1263.
19. Kamen, R., and H. Shure.1976.Topographyofpolyoma
virusmessengerRNA molecules.Cell 7:361-371. 20. Laemmli,U. K. 1970. Cleavage ofstructural proteins
duringtheassemblyof the headofbacteriophageT4. Nature(London) 227:680-685.
21. Lund, E., B. E.Griffin, and M.Fried.1977. Polyoma virus defective DNAs.U.Physical map of a molecule withrearranged and reiterated sequences (D74). J. Mol. Biol.117:497-513.
22. Macpherson,I.1969.Agar suspension culture for quan-titation of transformed cells, p. 214-219. In K. Habel and N. P. Salzman (ed.),Fundamental techniques in
virology.Academic Press Inc., New York.
23. Magnusson,G., and P. Berg. 1979. Construction and
analysesofviable deletionmutantsofpolyomavirus. J. Virol.32:523-529.
24.Maxam,R., and W. Gilbert 1977. A newmethodfor
sequencingDNA.Proc.Natl. Acad. Sci.U.S.A.
74:560-564.
25. Miller, L. K., andM.Fried. 1976.Construction of the genetic mapofthepolyoma genome. J. Virol. 18:824-832.
26. Paulin,D., andF.Cuzin.1975.Polyomavirus Tantigen.
on November 10, 2019 by guest
http://jvi.asm.org/
1220 NOTES
I.Synthesis ofamodifiedheat-labile T antigen incells
transformed withtheta-amutant.J. Virol.15:393-397. 27. Schaffhausen, B. S.,J.E.Silver,and T. LBenjamin.
1978.Tumorantigen(s)incellsproductively infected by wild-typepolyoma virus andmutantNG-18. Proc.Natl. Acad. Sci. U.S.A. 75:79-83.
28. Seif, R., and F. Cuzin. 1977. Temperature-sensitive growthregulation inonetypeoftransformedratcells induced by thetsa mutantofpolyoma virus. J.Virol.
24:721-728.
29. Silver, J.,B. Schaffhausen, and T. Benjamin.1978. Tumor antigens induced by nontransformingmutants
ofpolyoma virus. Cell15:485-496.
30. Soeda, E., J. R. Arrand, N. Smolar, and B. E. Griffin. 1979.Sequence from earlyregionof polyoma virus DNA containingviral replication origin andencoding small, middle and (part of) large T-antigens. Cell17:357-370.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/