0022-538X/93/031716-05$02.00/0
Copyright
©D
1993, American Society forMicrobiologyIdentification of
a
Short Amino Acid Sequence
Essential
for
Efficient Nuclear
Targeting of the Epstein-Barr
Virus Nuclear
Antigen 3A
AUDE
LEROUX, MONIQUE
BEREBBI,
MARYAMMOUKADDEM,
MICHELPERRICAUDET,
AND
IRENE
JOAB*
Centre National de la Recherche
Scientifique
Unite6
Associee1301,
Institut GustaveRoussy,
39
nreCamille Desmoulins,
94805 Villejuif, France
Received 17September 1992/Accepted 14 December 1992
The Epstein-Barr virus nuclear antigen 3A is expressed in the nuclei of cells latently infected by the Epstein-Barr virus. We have previously shown thatafragment of 265 amino acidswasessential fortheproper subcellular localization of the Epstein-Barr virus nuclear antigen 3A. As described in thispaper,wehave used deletion analysistoidentifyadecapeptide,
RDRRRNPASR,
which is essential for nuclear localization of this protein.Furthermore, this decapeptide isafunctional nuclear localizationsignalasdemonstratedby itsability totargetexpression of13-galactosidase
inthe nuclei of transfected cells.Epstein-Barr virus (EBV) is
ahuman
herpesvirus
associ-ated with
nasopharyngeal carcinoma, Burkitt's lymphoma,
and otherlymphoproliferative
disorders. Infection of humanprimary
Blymphocytes by EBV,
invitro, confers
uponthe
cells theability
to growpermanently
in culture(12).
Thesegrowth-transformed
Bcells
expressnine viral proteins,
including six nuclear antigens (LP and EBNAs 1, 2, 3A, 3B,
and3C)
andthree
membraneproteins
(LMP, TP1,
andTP2)
(reviewed
inreferences 17
and21). These proteins
arelikely
to
be
required for the immortalization
of Bcells, the
main-tenanceandregulation
ofexpression
from the viralgenome, orthe control of
latency.
EBNA1 is
required
for autonomous maintenance of the circularized EBVgenome(20)
and activatesatranscriptional
enhancer
located in
the oriPregion
(24).
EBNA2is
essential forEBV-induced
B-lymphocyte
transformation(4, 10)
and transactivates viral and cellular geneexpression (5,
18,
31-33).
LMPalsocontributes
totheinduction
of cell activa-tionmarkers,
alters thegrowth
properties
ofrodent fibro-blast celllines,
andupregulates
theexpression of
the bcl2 proto-oncogene whichis
able to protect thecells from
apoptosis
(11, 30, 32). The
genesencoding
theEBNA3A,
EBNA3B,
andEBNA3C proteins exhibit
homologies
intheir
sequences,structures,
and
sizes.They
aretandemly located
in theBamHI
Eregion of the
EBV genome.The EBNA3
proteins
arehigh-molecular-weight,
high-proline-content
polypeptides
whicharefound
in thenucleus.
Their
C-termi-nalregions
containdifferent
repeating polypeptide domains
(13, 14, 16, 22, 23, 26, 28). Since
EBNA3C induces CD21
expression,
the EBNA3proteins
maybetransactivators of
B-lymphocyte
geneexpression(32).
To
investigate EBNA3A functional domains,
we con-structedarecombinant
plasmid
abletodirect
theexpression
ofthefull-length
EBNA3Aantigen.
Alarge deletion in
the N-terminal endof this protein modified its subcellular
local-ization(14).
We presenthere the identification of the aminoacidsequence
required
for nuclear targeting of the EBNA3Aprotein.
Nuclear
localization signals (NLSs) have already
beenfound inseveralnuclear
proteins; however,
nouniver-*Correspondingauthor.
sal
sequencefor nuclear
localizationhas
beendescribed
(reviewed in references 7 and
29).
Recombinant plasmids
containing
deletionsin the
EBNA3A
open
reading frame. The cDNA
sequenceencoding
the
EBNA3A protein
downstream from theadenovirus 2major
late
promoter(19)
wasinserted in
theEcoRI site
ofthe
pUC13
vector,generating pUCE3
recombinant DNA.The
A795mutantwasconstructed bycutting pUCE3with
BglII
and
ligating
the endstogether.
Internal in-frame deletions between theseBglII
sites wereperformed
as follows.The
XhoI-StuI
restrictionfragment
ofpUCE3
waspartially
cleaved
by
Sau3A(Fig. 1).
Theresulting
XhoI-Sau3Afrag-ments were then inserted between the XhoI and
BglII
(second
site)
sites ofpUCE3.
In this way, three mutants(A654, A624, and A417)
weregenerated
that corresponded
tothe deletion of
218, 208,
and 139 aminoacids,
respectively,
from the EBNA3A openreading
frame. The A216 mutantwas constructed
by cleavage
withBglII
and BstEIIrestric-tion
enzymes,filling
in theends,
andligation.
Transient expression of
EBNA3A-deleted
proteins. The deletionmutantsdescribed aboveweretransfected intocells
from cell line 293by
the standardCaPO4
precipitation
method(8),
and the protein extracts wereassayed
in aWestern blot
(immunoblot) experiment (2).
The resultsshow that deletions within thecoding
sequenceledto theexpres-sion of shorter
polypeptides
that were still detectedby
an EBV-immune humanserumwhichwaspreviously
showntorecognize
EBNA3A(Fig.
2).
To
study specifically
the requirements for nuclearlocal-ization,
wefirstinvestigated
whetheratransientexpression-immunofluorescence
systemusing
the deleted variants ofEBNA3A
wouldreproduce
the nuclear localization seen aftertransfection ofpUCE3
into human293 and HeLa cells. Thecellswereassayed
forEBNA3A expression
by
indirectimmunofluorescence
48 h after transfection.The
anti-EBNAserumdescribed abovewasused forthese
experiments
(14).
Nomore than 10% of cells
expressed
EBNA3A, reflecting
the level oftransfection.Mock-transfected
cells showed a verylowlevel of
background fluorescence (data
notshown).
The deleted EBNA3A proteins were found to be nuclear when constructs A417 and A624(Fig.
3A)
were used in transfectionexperiments. Thus,
we concluded that amino1716
on November 9, 2019 by guest
http://jvi.asm.org/
Sau3A
A624 i
-Sau3A
A654
iSau3A
A795
F-H
BglI
A216
BglII
StuI
BglII StuI
BglII StuIl
BglII StulI
H
}F
BglIl* BstEII*
157-364
147-364
100-364
101-172 +change 173
nuclear
cytoplasmic
cytoplasmic
cytoplasmic
[image:2.612.81.526.66.350.2]*These restriction siteswereblunt-ended
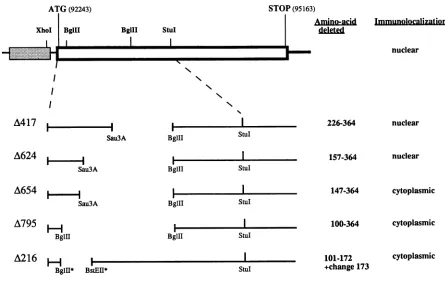
FIG. 1. Schematic representation of deletions in the EBNA3Aopen reading frame and subcellular localization of the corresponding
protein. Thetoplinerepresentsthe entire EBNA3Aopenreadingframe(openbox) taken fromacDNAplaced downstream of the adenovirus
2major latepromoter(shadedbox) (12). The coordinates in theEBVgenomeoftheinitiation andstopcodonsareindicated,aswellasthe
positionsofthe restriction sites used.Below areEBNA3A deletions: the solid lines representthesequences presentontherecombinant
plasmid. The numbers of thefirst and last amino acids lacking in the deleted proteinareindicated, aswellasthesubcellularlocalizationof
thecorresponding protein. In the A216mutantamino acid 173 is changed from GtoR.
A
4 3 2 1
FIG. 2. Immunological detec
line 293transfectedwithpUCE3 anti-EBNA-human serum (dilul complexes were visualized afti
labeledproteinA. (A)Lanes:1, (B)Lanes:1,pUCE3; 2, A624;3. kilodaltons are shown in the ce
mined withprestained standard
acids
157
to364
wereprobably
notessential for
the nuclear
localization of EBNA3A. In
contrast,the A654 and the A795
expressedproteins
wereexcluded from
thenucleus (Fig. 3B
andC,
respectively); immunostaining
wasobserved
through-B outthe
cytoplasm.
The A624plasmid harbors
afragment of
theEBNA3A
openreading frame which is 30
bp
longer than
1 2 3
the
oneincluded inthe A654
recombinant DNA. All
orpartofthe amino
acid
sequenceencoded
by these 30 bp would
thusseem tobe essential fornuclear
localization. Another
*
110possibility
is thatthis deletiondisrupted the protein folding
and the nuclear
localization
signal could thus be
inefficient.
*
84-
We constructed another recombinantplasmid,
the A216mutant
(Fig. 1),
whose deleted sequenceincluded
also the44 7
*fragment coding
for the10-amino-acid
sequence.It
wastransferred into HeLa cells, and the
intracellular localization
of theprotein
was determined. The fact that the EBNA3A4. 33
-_
encodedby
this mutant is not found in the nucleus either(data
notshown)
is further evidence of theimportance
ofthedecapeptide
sequencein nuclearlocalization.4 24 *_ In summary, amino acids 147 to 172were deleted in the
three
truncated EBNA3Aproteins
whichwerefound
in thetion ofEBNA3A in cells from cell cytoplasm and thus all or some of these amino acids are
andvariousconstructsby usingan implicated in the nuclear targeting of EBNA3A. The
com-tion, 1/100). The antigen-antibody parison of the cellular localization of A624 and A654proteins
er incubation with 1 ,uCi of 125I- indicates that the domain(s) responsible for EBNA3A nu-pUCE3; 2, A417; 3, A624;4, A795 car
that
isompos)
onsible or
essenu-A654.Molecularmassestimatesin clear localization is composed of at least one essential
nter of thefigure and weredeter- sequence which is contained in the 10 amino acids:
proteins.
RDRRRNPASR.
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.65.297.477.642.2]FIG. 3. Indirect immunofluorescence of HeLa cellstransfected withEBNA 3AmutantsA624(A), A654 (B), and A795 (C). Primary antibodywaspolyclonalEBV-positivehumanserum(dilution, 1/10)
which detectsEBNA3A protein (control); the secondary antibody
wasfluorescein-conjugatedgoat anti-humanimmunoglobulin G.
Insertionof
EBNA3A
sequencesintoaheterologous protein.,B-Galactosidase
wasused
as arecipient of the
putative
nuclearlocalization
signal of EBNA3A. To determine
whetherthesequence(amino acids 101
to172)
absentin the
A216 mutant is sufficient to induce nuclear localization, it
wasfused in
frame
tothe
coding sequenceof thef-galacto-FIG. 4. Indirect immunofluorescenceof
3-galactosidase
recom-binant protein in COS-1 cells. The plasmid pCH110 (Pharmacia) contains the LacZ gene from Escherichia coli placed under the control of the simian virus 40 promoter. Transfection ofpCH110in eucaryotic cells leads to expression of the ,B-galactosidase in the cytoplasm (A). We inserted the DNA fragments encoding amino
acids 101to172 and 147to157 of EBNA3A in the EcoRV sitesof pCH110 designated pCH110/220 (B) and PCH110/30
(C),
respec-tively. The primary antibody was a mouse monoclonal antibody directedagainstP-galactosidase,
and thesecondaryantibodywasafluorescein-conjugated goatpolyclonalantibodyto mouse immuno-globulinG.
sidase
gene
(designated
pCH110/216).
This insertion
notonly
allows
the
expression
of
3-galactosidase,
asshown
by
im-munofluorescencestaining
with a mouse monoclonalanti-body
to1-galactosidase (Zymed
Laboratories),
but also leads to a nuclearlocalization of thisprotein (Fig.
4B).
Thison November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.325.544.67.530.2]the nuclear
targeting
of
aheterologous protein.
Some
proteins
contain
morethan
onefunctionally
redun-dant NLS
(27). For
example, Dang and Lee
(6) identified
tworegions of the human c-myc
protein
necessary
for nuclear
targeting of the
protein. One
region
of the human c-myc
protein
functions
as anNLS and the other induces
only
partial nuclear targeting. In contrast, the simian virus 40
large T
antigen contains only
oneNLS
(15).
The
progester-onereceptor
also has
anNLS similar
tothe NLS of
simian
virus 40
large T antigen. This NLS is constitutive, and when
it is deleted, the
ligand-free receptor becomes
cytoplasmic.
However, this receptor has
asecond
NLS
which
requires
the
binding of the hormone
tobe effective
(9).
In
the
polyomavirus
large
Tantigen,
the
twonuclear
targeting
sequences
arefunctional.
However,
only
one(VSRKRPRP)
is able
todirect
areporter
protein
tothe
cell nucleus
(25).
Interestingly, the smallest known
targeting signal,
that
of the
adenovirus
ElA
protein,
KRPR, is present in three EBNAs
(EBNA1, EBNA2,
and
EBNA3C). Recently,
Ambinder and
al.
(1) reported
the
identification of the EBNA1 NLS
which,
in
fact, contains this
peptide (LKRPRSPSS).
The
region
of
EBNA2
containing
this amino acid sequence is also
impli-cated in
nuclear transport.
However,
it is
likely
that another
sequence
inthis
protein
is able
tofunction
as anNLS
(3).
Thus,
eventhough
many
NLSs have been
identified,
nosingle
consensussequence
has
emerged
(reviewed
in
refer-ence
7).
Examination of the different
NLSs, however,
re-veals
two commonfeatures:
they
areusually
short
(less
than
12
amino
acids),
and
they
contain
ahigh
proportion
of
positively charged
amino
acids.
Inthis report
wehave
shown that the NLS
of EBNA3A
antigen (RDRRRNPASR)
contains
at most10
amino
acids,
including
five
arginine
residues
(R). Although
weidentified
aminimal EBNA3A
sequence that
directs
a3-galactosidase
fusion
protein
tothe
nucleus,
we cannottotally
exclude the
possibility
that
an-other sequence will be
implicated
in
the nuclear
targeting
of
EBNA3A. If
such
asequence
exists,
wehave demonstrated
that it is
notsufficient for
directing
the EBNA3A
protein
tothe nucleus. Our
results
(see
also
reference
14) strongly
suggest
that the EBNA3A
protein
contains
asingle
NLS
whose
role
mustbe
considered in studies of functional
domains of the EBNA3A
antigen.
Weare mostgratefultoD.Walls and T. R. O'Connor for
helpful
criticalreadingof the manuscript.This work was supported by the "Ligue Nationale
Francaise
contre le Cancer," the "Association pour la Recherche sur le Cancer,"theCNRS,and the "Fondation pour la recherche
medi-cale." Aude Le Rouxwassupported by agrant from the"Ligue
Nationale Francaise contre le Cancer"(comite
de laSarthe),
France.5.
Cordier, M.,
A.Calender,
M.Billaud,
U.Zimber,
G.Rousselet,
0.
Pavlish,
J.Banchereau,
T.Tursz,
G.Bornkamm,
and G. Lenoir. 1990. Stable transfection ofEpstein-Barr
virus(EBV)
nuclear
antigen
2inlymphoma
cellscontaining
the EBVP3HR1 genomeinducesexpression
ofB-cell activation moleculesCD21 and CD23. J. Virol.64:1002-1013.
6.
Dang,
C.V.,
and W. M. F. Lee. 1988. Identification of the human c-mycprotein
nuclear translocationsignal.
Mol. Cell. Biol. 8:4048-4054.7.
Garcia-Bustos, J.,
J.Heitman,
and M. N. Hall. 1991. Nuclearprotein
localization. Biochem.Biophys.
Acta1071:83-101. 8.Graham,
F.L.,
and A.J.vander Ed.1973. Anewtechnique
forthe assayof
infectivity
of humanadenovirus 5 DNA.Virology
52:456-467.
9.
Guichon-Mantel, A.,
H.Loosfelt,
P.Lescop,
S.Sar,
M.Atger,
M.
Perrot-Applanat,
and E.Milgrom.
1989. Mechanisms of nuclearlocalization of the progesterone receptor: evidence for interaction betweenmonomers.Cell 57:1147-1154.10.
Hammerschmidt, W.,
and B.Sugden.
1989.Geneticanalysis
ofimmortalizing
functions ofEpstein-Barr
virus in human Blymphocytes.
Nature(London)
340:393-397.11.
Henderson,
S.,
M.Rowe,
C.Gregory,
D.Croom-Carter,
F.Wang,
R.Longnecker,
E.Kieff,
and A. Rickinson. 1991. Induc-tionofbcl-2
expression by
Epstein-Barr
viruslatent membraneprotein
1protectsinfected Bcellsfromprogrammed
cell death. Cell 65:1107-1115.12.
Henle,
W.,
V.Diehl,
G.Kohn,
H. ZurHausen,
and G. Henle. 1967.Herpes-type
virus and chromosome marker in normalleucocytes
aftergrowth
with irradiated Burkitt cells. Science 157:1064-1065.13.
Hennessy, K.,
F.Wang,
E.Woodland-Bushman,
andE. Kieff. 1986. Definitiveidentificationofa member of theEpstein-Barr
virus nuclear
protein
3family.
Proc. Natl. Acad. Sci. USA 83:5693-5697.14.
Joab, I.,
D. T.Rowe,
M.Bodescot,
J. C.Nicolas,
P.J.Farrell,
and M. Perricaudet. 1987.
Mapping
of the genecoding
forEpstein-Barr-virus-determined
nuclearantigen
EBNA3A andits transient
overexpression
in ahuman cell lineby using
anadenovirus
expression
vector. J.Virol. 61:3340-3344. 15.Kalderon, D.,
B. L.Roberts,
W. D.Richardson,
and A. E. Smith.1984. A short amino acid sequence able to
specify
nuclear location. Cell39:499-509.16.
Kerdiles, B.,
D.Walls,
H.Triki,
M.Perricaudet,
and I.Joab. 1990. cDNAcloning
and transientexpression
ofEpstein-Barr
virus-determined nuclear
antigen
EBNA3B in humancells and identification of noveltranscripts
from itscoding
region.
J. Virol. 64:1812-1816.17.
Kieff, E.,
and D. Leibowitz. 1990.Epstein-Barr
virus and itsreplication,
p. 1889-1990. In B. N.Fields,
D. M.Knipe,
R. M.Chanock,
M. S.Hirsch,
J. L.Melnick,
T. M.Monath,
and B.Roizman
(ed.),
Virology.
RavenPress, Ltd.,
NewYork. 18.Knutson,
J. C. 1990. Level ofc-fgr
RNA is increasedby
EBNA-2,
anEpstein-Barr
virus generequired
for B-cellimmor-talization. J. Virol. 64:2530-2536.
19.
Levrero,
M.,
V.Barban,
S.Manteca,
A.Ballay,
C.Balsamo,
M. L.Avantaggiati,
G.Natoli,
H.Skellekens,
P.Tiollais,
and M. Perricaudet. 1991.Defective and nondefective adenovirusvec-tors for
expressing
foreign
genes in vitro and in vivo. Gene101:195-202.
on November 9, 2019 by guest
http://jvi.asm.org/
20. Middleton, T., and B. Sugden. 1992. EBNA1 can bind the enhancerelement to the initiatorelement of Epstein-Barr virus plasmid origin of DNA replication. J. Virol. 66:489-495. 21. Miller, G. 1990. Epstein-Barr virus biology,pathogenesis, and
medical aspects, p. 1921-1957. In B. N. Fields, D. M. Knipe, R. M.Chanock, M. S. Hirsch, J. L. Melnick, T. M. Monath, and B.Roizman (ed.), Virology. Raven Press, Ltd., New York. 22. Petti, L., and E. Kieff. 1988. A sixth Epstein-Barr virus nuclear protein (EBNA3B) is expressed in latently infected growth-transformedlymphocytes. J. Virol. 62:2173-2178.
23. Petti, L., J. Sample, F. Wang, and E. Kieff. 1988. A fifth Epstein-Barr virus nuclear protein (EBNA3C) is expressed in latently infected growth-transformed lymphocytes. J. Virol. 62:1330-1338.
24. Reisman, D., and B. Sugden. 1986. trans activation of an Epstein-Barrviral transcriptional enhancer by the Epstein-Barr viral nuclear antigen 1. Mol. Cell. Biol. 6:3838-3846.
25. Richardson, W. D., B. L. Roberts, and A. E. Smith. 1986. Nuclearlocationsignals in polyoma virus large T. Cell 44:77-85. 26. Ricksten, A., B. Kallin, H. Alexander, J. Dillner,R.Fahraeus, G. Klein, R.Lerner,and L.Rymo.1988. BamHl E region of the Epstein-Barr virus genome encodes three transformation asso-ciated nuclear proteins. Proc. Natl. Acad. Sci. USA 85:995-999. 27. Robbins, J., S. M. Dilworth, R. A. Laskey, and C. Dingwall. 1991. Two interdependent basic domains in nucleoplasmin
nu-clear targeting sequence: identification of a class ofbipartite nuclear targeting sequence. Cell 64:615-623.
28. Shimizu, N., M. Yamaki, S. Sakuma, Y. Ono, and K. Takada. 1988.ThreeEpstein-Barrvirus(EBV)-determined nuclear anti-gens induced by the BamHl E region of EBV DNA. Int. J. Cancer 41:744-751.
29. Silver, P. A. 1991. How proteinsenterthe nucleus.Cell 64:489-497.
30. Wang, D., D. Leibowitz, and E. Kieff. 1985. An EBV membrane protein expressed in immortalized lymphocytes transform es-tablished rodent cells. Cell 43:831-840.
31. Wang, F., C. Gregory,M. Rowe, A. Rickinson, D. Wang, M. Birkenbach, H. Kikutami, T. Kishimoto, and E. Kieff. 1987. Epstein-Barr virus nuclear antigen 2 specifically induces expres-sion of the B-cell activation antigen CD23. Proc. Natl. Acad. Sci.USA 84:3452-3456.
32. Wang, F., C. Gregory, C. Sample, M. Rowe, D. Leibowitz, R. Murray, A. Rickinson, and E. Kieff. 1990. Epstein-Barr virus latent membrane protein(LMP1) and nuclear proteins 2 and 3C
areeffectors of phenotypic changes in B lymphocytes: EBNA-2 and LMP1cooperatively induce CD23. J. Virol. 64:2309-2318. 33. Zimber-Strobl, U., K. Suentzenich, G. Laux, D. Eick, M. Cordier, A.Calender, M. Billaud, G. Lenoir, and G. Bornkamm. 1991. Epstein-Barr virus nuclear antigen 2 activates transcrip-tion ofthe terminalprotein gene. J. Virol. 65:415-423.