JOURNALOFVIROLOGY, Feb. 1987, p. 465-471
0022-538X/87/020465-07$02.00/0
Copyright X) 1987, American SocietyforMicrobiology
Epstein-Barr Virus Nuclear Antigen
Forms
a
Complex That Binds
with High Concentration
Dependence
to
a
Single DNA-Binding
Site
GREGORY MILMAN* AND EUNSEONG HWANG
Department of Biochemistry, The Johns Hopkins UniversitySchoolofHygieneandPublic Health,
Baltimore, Maryland 21205
Received 9 July 1986/Accepted 17October 1986
A bacterially synthesized 28-kilodalton carboxyl-terminal fragment (28K-EBNA) of Epstein-Barr virus nuclear antigen shows highly concentration dependent binding to monomer, dimer, and trimer copies of
synthetic DNA-bindingsite
5' GATCTAGGATAGCATATGCTACCCCGGGG 3'
3' ATCCTATCGTATACGATGGGGCCCCCTAG 5'
inbacterial plasmids. Therateof the binding reaction is independent of the number of sites, but dependent uponthelengthof theDNA containingthesites. These dataareconsistentwith 28K-EBNA locating its binding
sites bya processoffacilitated transferorsliding along the DNA. The highly concentration dependent binding
suggeststhat multiple 28K-EBNA monomerpolypeptides formacomplex beforeor duringbinding. Binding
occursequallywell at 24and37°C,butnot at0°C. A28K-EBNAcomplex boundtoasinglesite has unoccupied
bindingsitescapable of interacting withadditionalDNA molecules. Such interactionisconfirmed byagarose gelelectrophoresis of protein-DNAcomplexes whichindicate thata28K-EBNA complex forms bridges between
twoDNA molecules. A bridge between the two binding regions in theEpstein-Barrvirus origin of plasmid
replication(oriP) wouldformaloopstructurewhich could beanimportantfeature for the regulatory function
ofauthentic Epstein-Barr virusnuclearantigen.
Epstein-Barr virus (EBV) transforms human B lympho-cytesinto permanently proliferating cell lines (19, 25). EBV
DNAreplicatesinthese cellsasamulticopy circularplasmid orepisome. Thecis-acting signals which enable EBV DNA plasmid replication are located on an 1,800-base-pair (bp) region(oriP)in the EBVBamHI Crestriction fragment (20, 23).oriPcontainstwolociessentialtoitsfunction (16). The leftend of oriP contains 20copies of 30-bp tandemrepeats which have a 12-bp palindromic consensus sequence TAGCATATGCTA. The right end of oriP contains four copiesofrepeatsnearlyidenticaltotheconsensus sequence, and the DNA sequence in this region can be drawn as a
116-bp stem-loop dyad symmetry structure reminiscent of other originsofreplication.
EBVnuclear antigen(EBNA-1) coded inthe EBV BamHI Krestrictionfragmentis theonlyviralproductrequiredfor
oriP-dependent plasmid replication (24). EBNA-1, like
polyomavirus and simian virus 40 T antigens, appears to performpleiotropic functions. Inadditionto arole in repli-cation, the protein may act as a transactivating
transcrip-tionalenhancer(16).Theamino half of EBNA-1polypeptide
contains a repeated Gly-Gly-Ala sequence whose length varies in different EBV isolates. Although these repeats
compriseoverathird of the B95-8 strainEBNA-1sequence, they do not seem necessary for EBNA-1 to function in plasmid replication (24). Phosphorylation and dephos-phorylation of EBNA-1 may regulate the physiological ac-tivityof theprotein (9).
Thepurificationandcharacterizationof EBNA-1 has been difficult because of the scarcity of the protein in cells. For
*Correspondingauthor.
example, Sculleyandco-workers(18) obtained only 5 ,ug of paritally purified EBNA-1 after2,500-fold enrichment from 20 g ofRaji cells. Moreover, their purification procedures required solubilizationindenaturingreagents(4 Mureaor6
Mguanidine hydrochloride) whichmay permanently inacti-vatethephysiological activities of EBNA-1. Higher levels of
EBNAmay be obtained from COS-1 cells transfected with DNAcodingfor EBNA-1 under thecontrolofasimian virus 40 virus promoter(17) orfromherpes simplexvirus
recom-binantscontainingachimeric EBNA-1 construct (10).
Our approach to characterizing EBNA-1 has been to synthesizeinbacteriaa28-kilodaltoncarboxyl-terminal frag-ment(28K-EBNA) whichpossesses propertiesof the intact molecule. The 28K-EBNA polypeptide is recognized by
anti-EBNA antibodies in humanserum(14) and is usefulin diagnosis of EBV related diseases (8). Filter-bindingassays
and DNase I footprinting demonstrated that 28K-EBNA
specificallybinds to therepeat sequencesinoriP(15).
To understand better the interaction of 28K-EBNA with its DNA-binding sequence, we constructed a synthetic EBNA-1-binding site containing the concensus palindrome sequencefrom oriP. Thesyntheticsequencewasinsertedin monomer, dimer,and trimerformsinto the Escherchia coli plasmidpUC8 (13). Inthispaperwedescribetheanalysisof 28K-EBNA binding to these synthetic repeats by filter binding assays and the examination of the association of
28K-EBNA with DNAbyagarose gelelectrophoresis.
MATERIALS AND METHODS
Bacteriallysynthesized
28K-EB?iA.
Thebacterialsynthesisandpurificationof 28K-EBNAhavebeendescribed(14). To
summarize, the 2,236-bp SmaI subfragment of the EBV
(P3HR-1) BamHI fragment K was inserted into the
high-465
Vol.61,No. 2
on November 10, 2019 by guest
http://jvi.asm.org/
PLASMIID DUC8
PvuI1 Pvull Ndel
1 430 2722
308
PLASMIID DUC8 (308 bD Pvul! 4ra nent)
EcoRI Hindll!
Pvull BamHl Pvul
Iwu I
1 187 308
177 207 PLASMID DRI (337 bD Pvull fraQment)
186 216 Ndel Hindll
Pvull EcoR! BamHI Pvul
I ~~~~~~~~~II I
I 1177 216 337
201 236
FIG. 1. RestrictionmapofpUC8 and pRl.
expression bacterial plasmid pHE6. The resulting plasmid, pNAK28, causes E. coli to produce large quantities of
28K-EBNA, apolypeptide containing the carboxylterminal
191 amino acids ofEBNA-1. The 28K-EBNA polypeptide waspurified to greaterthan 95%homogeneity by
phospho-cellulose andhydroxyapatite column chromatography.
Construction of synthetic EBNA DNA-binding sequences. DNA oligonucleotides 5' GATCTAGGATAGCATATGCT ACCCCGGGG 3' and 5' GATCCCCCGGGGTAGCATAT
GCTATCCTA 3' were synthesized on an oligonucleotide synthesize (Applied Biosystems), purified by acrylamide gel electrophoresis, and isolated from the gel. The oligomers wereannealed by heating to90°C and slow coolingtoroom temperature. The annealed oligomers were ligated into re-peatswith T4ligaseandthencleavedwithBamHIandBglII toproducetandem multiple repeat sequences.
32p incorporation into plasmid DNA. Plasmid DNA was
cleaved withPvuII, and theresultingblunt-endedDNAwas
end labeled with Klenow DNA polymerase by using only [32P]dGTP in the reactions (12a). The 32P-labeled small 300-bp fragments were purified by electrophoresis in 1.5%
low-melting-pointagaroseandseparated from theagaroseby phenol-chloroform extraction and ethanol precipitation.
End-labeled DNAwasusedfor the binding studies analyzed
by agarosegel electrophoresis.
DNAwasalsolabeled withT4 DNA polymerase (12a). A
2-pg sample of plasmidwascleaved with 30 U of Hindlll in
20 pulof T4polymerase buffer (33 mMTris acetate, 66mM
potassium acetate, 10 mM magnesium acetate, 0.5 mM
dithiothreitol, 0.1 mg of bovine serum albuminper ml, pH 7.9). The reaction mixturecontaining the cleaved DNAwas
incubated with 3.7 U of T4 polymerase (New England Biolabs) for exactly 9 minat 37°C to digest approximately 300 to 400 bases. The digestion was halted, and the DNA
was resynthesized by the addition of [32P]dATP and the other three unlabeled deoxynucleotide triphosphates. The labeledDNAwasseparated fromunincorporated[32P]dATP by column chromatography on Bio-Gel A-0.Sm (Bio-Rad
Laboratories). DNA labeled byT4polymerase wasused for
nitrocellulosefilterbinding assays.
Mobility retardation in agarose electrophoresis.
PvuII-cleaved pUC8 DNA was mixed with PvuII-cleaved pRl, pR2, orpR3 DNA before bindingtoprovide anhomologous
comparison DNA lacking an EBNA-1-binding sequence.
The DNAs and 28K-EBNA were mixed in 4 ,u ofbuffer
containing40 mMTrisacetate(pH
8.3)-260
mMNaCI-1 mMMgCl2-1
mM mercaptoethanol-1 mM EDTA-12.5 ,g ofbovine serumalbumin per ml. The reactionswere incubated for 1 hat 24°C and then mixed with 1
RI
ofelectrophoresisbuffer(40 mM Tris acetate, pH 8.3) containing 50%glycerol
andbromphenolblue indicatordyeandplacedin the wells of
a 1.5% low-melting-pointagarose (Bethesda Research
Lab-oratories) gelinamini-gelapparatus(Hoeffer).Thesamples
were electrophoresed at 5 W (approximately 100 V) for
approximately 2 h until the tracking dye reached
approxi-mately 1 in. (ca. 2.5 cm) from the end of the gel.
PvuII-cleaved lambda DNAwasusedto provideDNA size
mark-ers.Thegelsweresoakedin0.5pgof ethidium bromide per mlfor 10 min and thenphotographed. Forautoradiography,
thegels were dried ontoWhatman no. 1 filter paper witha
vacuumgeldrier(Hoeffer) without heatfor1hand thenwith heatfor 15 min.
Nitrocellulosefilterbinding of 28K-EBNADNAcomplexes. Samples of28K-EBNA and 3 fmol of each 32P-labeled DNA (1,000 to 10,000 cpm) were incubated in 5 pI of buffer containing50 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid) (pH 7.5),200 mMNaCl,5 mMMgCl2,
1 mMdithiothreitol, 250
pg
of BSA perml,and 125pLg
of E. coli tRNA per ml. Nitrocellulose(Schleicher
and Schuell Co.; BA85)was cut into0.5-by0.5-cm squares and soaked for 30 min in 0.5 M KOH, rinsed 10 times in water, andequilibrated in 100 mM Tris hydrochloride (pH 7.5). The filters were placed on a fritted glass holder attached to a
vacuum. The28K-EBNA DNAmixtureswereappliedtothe
nitrocellulose, andthe filterswere washed with 1to 5 mlof
buffercontaining25 mMHEPES(pH7.5), 150 mM
NaCl,
5mMMg2Cl,and10% glycerol.Thefiltersweredried,and the
amount of bound 32P-DNA was determined by scintillation counting.
RESULTS
Plasmidscontaining EBNA-binding sequences. Thetandem EBNA-1-binding sites inoriPcontain different left(AGGA)
andright (CCCR)consensus sequences surroundinga12-bp palindromic repeat. The two synthetic DNA strands were
annealed toproduce astructure with similar asymmetry:
AGGA PALINDROME CCC
5' GATCTAGGATAGCATATGCTACCCCGGGG 3'
3' ATCCTATCGTATACGATGGGGCCCCCTAG 5'
Theannealed DNA monomers were ligated with T4 ligase to
form high-molecular-weight, randomly associated DNA
re-peats. Toobtaintandem repeats forinsertion intoplasmids,
theannealed monomer repeats wereconstructed with half of
a
BglII
site ononeend and halfofaBamHIsite on the other.A nontandemligationeventforms acomplete site for one of these enzymes. The high-molecular-weight ligated DNA was cleaved with both restriction enzymes to produce only tandem repeats.
The tandem repeats wereinserted into the BamHl site of
plasmid pUC8 (13), and plasmids pRl, pR2, and pR3 were constructedcontainingone, two,and three synthetic EBNA
binding sites, respectively. The distance between synthetic EBNA-binding sites is the same as that in the "tandem repeatregion" (15) of authentic EBV DNA. The number of
insertedrepeats wasdetermined by cleavage withPvuIIand
electrophoresisof the cleaved DNA on 1.5% agarose. Maps ofimportant restriction sites in pUC8 and pRl are shown in
Fig. 1.
on November 10, 2019 by guest
http://jvi.asm.org/
HIGHLY CONCENTRATION DEPENDENT EBNA BINDING TO DNA Rate of 28K-EBNA binding is independent of number of
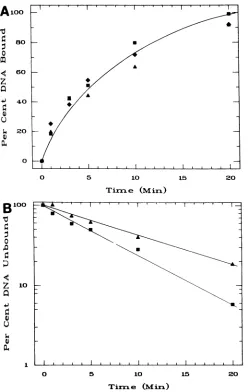
sites on the DNA. HindIII-cleaved, 32P-labeled linear plas-mids pRl, pR2, pR3, and pUC8 were incubated at 24°C with 2.6 ,ugof 28K-EBNA per ml for different times. The percent DNA bound as a function of time is shown in Fig. 2A. The
time course ofbindingfor all three plasmids was the same. The binding reaction appears to follow first-order kinetics, and the curve is a regression logarithmic fit to the data. A
linear curve resulted when the average percent DNA
un-boundfor all three plasmids was plotted on a semilog scale (Fig. 2B). Under these experimental conditions, the half-time for binding of 28K-EBNA to the plasmids was
approx-imately 5 min.
Onceformed, the binding complex was very stable. The amount of DNA bound was not greatly affected by dilution
of 28K-EBNADNAcomplexes in solutions containing up to
A1oo
0
z
0 Q)
80
60
40
20
0
0 5
Tix.
B
1000
0
z 10
a
[image:3.612.312.554.86.222.2]I.)
TABLE 1. StabilityofEBNA-DNAcomplexa
Treatment % of DNA bound
100 mM NaCl... 100
200 mMNaCl... 100
400 mMNaCl... 74
733mMNaCl... 0
1mMP04- ... 10O 7mMP043 ... ... 86
67 mMP043 ... ... 67
333mMP04-3... ... 47
pH7.5 ... .. 100
pH 5.0... .. 79
pH 9.5 ... .. 72
aSamplescontaining3fmolof32P-labeledpRl and464 pmol of28K-EBNA (2.6 ,ug/ml)in 5ILIwereincubated for 1h at24°C. The incubation mixtures werethendilutedwith 10p.lof solution containing sodium chloride, sodium phosphate (pH 7.5), 100 mM sodium acetate (pH 5.0), or 100mMsodium carbonate (pH 9.5) to attain the final concentrations listed above. The mixtures were incubated for10 min, and the percentages ofDNA bound compared with undiluted samples were determined by the nitrocellulose filter-binding assay.
X411'
400 mM NaCl, in buffers at pH 5 and 9.5, or in highA concentrations of
phosphate (Table
1).
Factorsdetermining the rate of 28K-EBNA binding to DNA.
The binding time was dependent on the length of DNA
containing the binding sequence. The 2,751-bp 32P-labeled pRlplasmid described above was cleaved withPvuIItogive
a 236-bp fragment containing the binding site. The 28K-EBNAbindingto this smallerfragment also displayed first-order kinetics withahalf-time forbinding of8 min(Fig. 2B). The rateofthebinding depended upon both the
concen-tration of 28K-EBNA and the temperature (Fig. 3). At a
j I 28K-EBNA concentration (13.2
p.gIml)
that was fivefold10 15 20 higher than that used in the experiment shown in Fig. 2, the
binding of 28K-EBNAtopRl occurredapproximately five-e (Miri) fold faster. The half-time for binding at this 28K-EBNA
,~.l concentration was a little under 1 min. In contrast to the
100
0
z
m
a
4.1
U
0 5 10 15 20
Timrle (Mimi)
FIG. 2. Rateof28K-EBNA bindingtopRl, pR2,andpR3.(A)
Linear32P-labeled plasmids pRl (-), pR2(*), andpR3 (A)were incubated for the indicated timeswith 2.6 ,ugof28K-EBNA perml,
and thepercent DNAboundwasdeterminedbythenitrocellulose filterbindingassay.(B) The2,751-bplinearpRl (D)plasmidand the 236-bp PvuII-HindIII-cleaved pRl (A) fragment were incubated with 2.6 ,ug of 28K-EBNA per mlforthe indicated times,and the percent DNA unboundwasdeterminedbythenitrocellulose filter-bindingassay.
80
60
40
20
0
0.v'I I0 A
0 10 20 30 40 50 60
Timrl e (Miri)
FIG. 3. Factorsaffectingtherateof 28K-EBNAbinding. Linear 32P-labeledplasmid pRl was incubated at24°C (*) and 37°C (-) with 2.1 ,ugof 28K-EBNA per ml and0°C (A)and24°C(V)with 13.2 ,ug of 28K-EBNA per ml, and the percent DNA bound at the indicated times wasdetermined bythe nitrocellulose filter-binding assay.
VOL.61, 1987 467
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.52.302.252.644.2] [image:3.612.315.556.479.668.2]468 MILMAN AND HWANG
100 high concentrations of DNA, suggesting that this bandarises
from interaction between two DNAmolecules.
28K-EBNA cross-links DNA molecules
containing binding
I 0
80
/sites. 32P-labeled PvuII fragments from pUC8 and
pRl
were
/ _ mixed with excess unlabeledPvuII
fragments
frompUC8,
0 pRl, pR2, orpR3. The DNA mixtureswereincubated with
60
28K-EBNA,
and theresulting
DNAspecies
wereanalyzed
by
agarosegel
electrophoresis (Fig. 6). InFig. 6A,the small unlabeled PvuII fragments for pUC8, pRl, pR2, and pR34 40 appear as heavily staining bands in lanes H, G, F, and E. The
32P-labeled
pRl
andpUC8
PvuIIfragments
are visible as lightly staining bands in the same lanes. Incubation with u$4 20 _ _ 28K-EBNA causedadecrease in the intensities of the bands
X) containing binding sites. The 28K-EBNAcaused nochange
in the heavily staining pUC8 PvuII band in lane D or the
o _ I I lightly staining pUC8 bands in lanes A through C. Binding of
28K-EBNA caused the clearappearance of two new bands
0 2 4 6 8 10 12 14 only in lane C. The autoradiograph of the gel in Fig. 6B
EBNA Coriceritratiora (Gg/rml) demonstrates that incubation with 28K-EBNA caused no
[image:4.612.66.305.71.263.2]changeinthemobilityof thepUC8 small PvuII fragment, but FIG. 4. 28K-EBNA binds withhigh concentrationdependence. greatly decreased the intensity of the
pRl
small PvuIILinear32P-labeled plasmids pRl (K), pR2 (*), andpR3 (A) were fragment in lanes A through D from that in the absence of incubated for 30 min at 24°C with the inciated concentrations of 28K-EBNA in lanes E through G. When the 28K-EBNA
28K-EBNA, and the percent DNAbound was determinedby the bound tothe
pRl
PvuIIfragment,
afast-migrating complex
nitrocellulose filterbindingassay. (at an apparentmobility of 468bp)was produced in all four
DNA mixtures (lanes A through D), but a slow-migrating
results at 24°C, binding at0°C was barely detectable even
after 60 min. Ata slightly lower 28K-EBNAconcentration
(2.1
pug/ml),
the rate ofbindingdecreaseddramatically.
Thebinding did not differgreatly between 37 and 24°C, and at both temperatures the apparent half-time of binding was
approximately 70 min. The percent DNA bound at 24°C
continued to increase for 120min, and at0°C there wasno
detectablebinding (data not shown).
Binding of 28K-EBNA ishighlyconcentrationdependentat a single binding site. Figure 4 illustrates the sharp depen-dence ofbinding on the concentration of28K-EBNA
pro-tein.Different concentrations of28K-EBNAwereincubated
for 30 min at24°Cwith linearplasmids pRl, pR2, pR3, and
pUC8.NobindingtopUC8DNAwasobserved. Thebinding
of 28K-EBNA to the plasmids with one, two, or three repeats was virtually identical.
Essentially
nobinding
wasobserved below2.1 ,ugof28K-EBNA permlevenafterlong time periods, and complete binding occurred at
concentra-tions of2.6 ,ug/mland higher. Binding at2.1 ,ug/ml hadnot
reached equilibriumduring the 30-min periodofincubation,
and the twofold difference inbinding between Fig. 3 and 4 resultedfrom the large rate differences for small variations in 28K-EBNAconcentrationin thishighlysensitive range. The results are similar to those previously observed for
28K-EBNAbindingtoauthenticoriPsitesinpHEBO (15;
unpub-lished observations).
Interactionof28K-EBNA withsynthetic repeat sequences.
Figure 5 illustrates the binding of 28K-EBNA to a single
syntheticrepeat. Amixture ofPvuII-cleavedpUC8 and pRl
was incubated for 60 min with 28K-EBNA, and the DNAs
were separated by electrophoresis in 1.5% agarose and stained with ethidium bromide. The 308-bp fragment is
derivedfrom pUC8, and the 337-bp fragment is the homol-ogous fragment from pRl containing the 29-bp synthetic
insert. The 28K-EBNA bound only to the pRl 337-bp
fragment, shiftingits mobility to the two new positions with apparent mobilities of 370 and 820 bp (Fig. 5, lane B). The relativeintensitiesof the two new bands did not change with
28K-EBNAconcentration. The upper band appeared only at
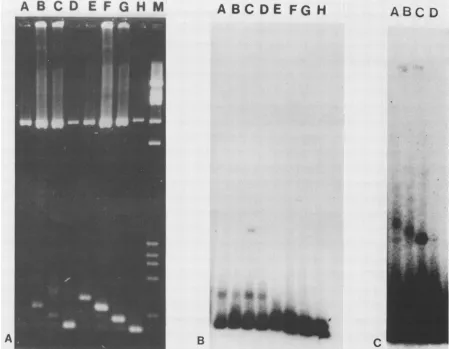
FIG. 5. Binding of 28K-EBNA toasingle repeatinpRl. Lane A containedPvuII-cleaved lambda DNAformolecular weight markers (sizes: 343, 468, 532, 636, 1,708, and 2,296 bp). Lanes B and C contained 110 fmoleach ofPvuII-cleaved pUC8 and pRl DNAs. Lane Balso contained 13.3 pmolof28K-EBNA. The bottom right arrowindicates thepUC8 308-bp fragment, and the top right arrow indicates the pRlhomolgous 337-bp fragment containing the 29-bp EBNA-binding site. The two left arrows indicate the two new positions (apparent mobilities of370 and820bp) of the pRl fragment boundto28K-EBNA.
J. VIROL.
9
r,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.405.483.352.631.2]HIGHLY CONCENTRATION DEPENDENT EBNA BINDING TO DNA
A B
CD E F
G H
M
____--
ABC DE FG H
A
B
C
FIG. 6. 28K-EBNA cross-links DNA molecules containing binding sites. (A) Ethidium bromide-stained 1.5% agarose gel. Lane M contained PvuII-cleaved lambdaDNAfor molecular weight markers. Lanes A through D contained 13.3 pmol of 28K-EBNA. Lanes A through G contained approximately6.5fmol each of32P-labeled(300 cpm) 308 and 337-bpfragments fromPvuII-cleavedpUC8 andpRl.In addition, 55 fmolof unlabeled PvuII-cleaved pUC8 DNA was included in lanes D and H, PvuII-cleaved pRl DNA in lanes C and G,
PvuII-cleavedpR2DNAinlanes B and F; andPvuII-cleavedpR3 DNA in lanes A and E. (B) Autoradiograph of gel in (A) exposed for 18 h.(C) Autoradiograph ofgel in (A)exposed for 120 h.
complex (at 870 bp apparent mobility) was formed only in
the presence ofexcess unlabeled pRl. An autoradiograph exposedforalonger time (Fig. 6C) reveals that the labeled
pRl PvuII fragment formed slower-migrating complexes
withunlabeled pR2 and pR3 PvuII fragments (lanes A and
B), but formed no complex with the pUC8 PvuII fragment
(lane D).
DISCUSSION
Thefacilitatedtransferorslidingofabinding protein along
aDNA molecule is themechanism proposedfor thelocation
of
binding
sitesbymanyregulatory proteins (e.g.,E. coli lacrepressor) (2). The rate-limiting step is the
low-affinity,
nonspecificassociation ofthebinding proteintoDNA.After
nonspecific binding, the protein rapidly migrates along the DNAto locate a high-affinity specific binding site. A facili-tated transfer mechanism is
required
to accountfor observedrate constants
(1010
Ms-')
which are 100- to1,000-fold
greater than diffusion-controlled reactions. A calculatedone-dimensional diffusion coefficient of10-9 cm2s-1 forthe lac repressor enables the
protein
to scan 1,000bp
pers.Two kinetic experiments suggest that the 28K-EBNA
polypeptidelocatesits
binding
siteby
facilitated transfer. Inthe firstexperiment (Fig. 2A), therateof 28K-EBNA binding isindependent ofthe numberofhigh-affinitybinding sites on
aDNAmolecule. Asinglesite in pRl boundby28K-EBNA issufficienttoretain DNAonnitrocellulose.The presence of additionalbinding sitesinpR2orpR3 has little influenceon
the time required for binding. All three plasmids bind to
28K-EBNA with apparent first-order kinetics and with the
same rateconstant. Theseobservations are consistentwith the rate-limitingstep beingthe low-affinity bindingtoDNA
predicted by the facilitated transfermechanism.Then,even
though the time required for28K-EBNA to scanthe DNA
forhigh-affinitysites maydecrease with increaseddensityof
sites,nosignificant
change
would beexpected
inthe overallrate of DNAbinding by 28K-EBNA.
In the second experiment (Fig. 2B), the rate ofbinding dependsuponthelengthofthe DNAcontainingthebinding
site. The timerequiredfor 28K-EBNAtobind halfthe DNA
is 5 min for a 2,751-bp DNA
(linear
HindIII-cleavedpRl
DNA) and 8 min for a 236-bp DNA (the same DNA also
cleaved withPvuII).These observationsarealso consistent
with a facilitated transfer mechanism. The 10-fold lower
concentration for the shorter DNA causes alower rate for
the low-affinity, nonspecific binding. The rate may not be
ABC D
VOL.61, 1987 469
. .,.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.78.527.75.424.2]10-fold lower because the nonspecific 2,515-bp DNA frag-ment in the reaction could facilitate the transfer of the binding protein from one DNA moleculeto another.
The 28K-EBNA concentration is 2.6 ,ug/ml in the rate
studies shown in Fig. 3. This concentration is slightly above the threshold required for binding. A fivefold increase in 28K-EBNAconcentration from 2.6 to 13.2
jig/ml
producedacorresponding fivefold increase in the rate of binding to
DNA (Fig. 3). However, a small decrease in 28K-EBNA concentration from 2.6 to 2.1
jig/ml
causes a 14-fold de-crease inbinding rate. Therateofbindingisapproximatelythe same at 24 and 37°C degrees, but is greatlyinhibited or
totallyblocked at0°C, suggestingthataggregation does not
take place at0°C.
The sharp transition fromboundtounbound DNAwithin a narrowrangeof28K-EBNAconcentrationdoesnotresult fromdepletionofproteinin thereactionbybindingtoDNA. At a 28K-EBNA concentration of 2.1 ,ug/ml in the binding
reactions, the molar amount of protein (374 fmol) is more
than 100-fold in excess of that of the DNA (3 fmol). The
excess 28K-EBNA protein is active and capable ofbinding
atleast 0.1fmol of DNA per fmolof28K-EBNAat2.6
jLg/ml
(datanot shown). Once thebinding complexis formed,it is very stable (Table 1).
Previous studies (15; unpublished observations) of 28K-EBNAbindingtoauthenticoriPsitesin theplasmid pHEBO
(20) indicated a step function dependence on 28K-EBNA
concentration. The apparent cooperativity of 28K-EBNA
binding topHEBO couldresultfrominteraction among the
multiplebinding sites. Anexaminationof therateofbinding
toasinglesite(Fig. 3) indicatesalargeincrease in therateof
the binding reaction for a small increase in 28K-EBNA
concentration.The same sharpbinding dependenceon 28K-EBNA concentration is foundforplasmids containing one, two, or three binding sites (Fig. 4). The concentration of
28K-EBNA required for binding in these experiments is almost identical to thatobserved forbinding to pHEBO in
earlier studies. Therefore, if the step function
dependence
results from cooperativity, the interactions must require only asingle binding site. Cooperativity at a singlebindingsite suggests that the 28K-EBNA forms a complex either beforebinding oratthebinding site.
Thespecificity ofEBNA-1as aregulatoryprotein depends
upon the fidelity of the EBNA-DNA interaction. High fi-delity requires EBNA-1 recognition of EBV DNA-binding sites in the presence of human genomic sequences. If it occurredrandomly, a12-bpbindingsitewould be presentat
approximately 60 copies in a
109-bp
genome. An evengreater number of EBNA-binding sites probably exist
be-causethereisambiguity in the consensusbindingsequence. EBNA-1 may achievebinding specificity in the presence of
these pseudo-binding sites by forming complex structures which link clusters of binding sites. Because these clusters
areextremelyunlikely to occur by chance, the nucleoprotein
structuresformed can provide high-fidelity recognition (5). There are many examples of procaryotic complex nucleoprotein structures. The lambda phage Int protein interacts with approximately seven sites distributed over 230 bp and condenses the linear sequence of 800 A (ca. 80 nm) into a compact three-dimensional structure of 140 A in diameter (3, 4). Similarly, lambda phage 0 protein binds to
eight sites within 105 bp at the lambda origin of replication
(6),andE.coli DnaA protein binds to four sites in 240 bp in the oriC origin of replication (7). Clusters of binding sites which probably form nucleoprotein complexes also are
foundin manyeucaryotic viruses (21).
The EBV oriP contains a cluster of 20 tandem repeat EBNA-1 binding sequences.Twoother
regions
in the EBV genome contain clustered arrays ofEBNA-1-binding
sites. Four sites occur in the dyad symmetryregion
oforiP, andtwo additional sitesoccur in the BamHI
Q
restrictionfrag-ment (15). Binding ofEBNA to these clustered sites could result inorganized
nucleoprotein
structures.A consequence of a 28K-EBNA complexbinding
to asingle
site is that additionalunoccupied
DNA-binding
sites would be presenton the
complex.
These empty sites should then be able tobind additional
copies
of thebinding
sequence. If theseadditional
binding
sequences were on different DNAmole-cules,
binding
would cross-link the DNA. Thefollowing
experiments
demonstrate that 28K-EBNA can cross-linktwoDNA strands
containing
binding
sites.The agarose
gel
electrophoresis mobility
retardationex-periment
exhibited inFig.
5 demonstrates that 28K-EBNAbinds
specifically
tothe337-bp fragment
ofpRl
and not tothe
homologous
308-bp fragment
ofpUC8.
Theonly
differ-encebetween these
fragments
isthe inserted29-bp
synthetic
binding site. The
binding
of 28K-EBNA to the337-bp
fragment causes adecrease in the
intensity
ofthis band and the appearance oftwo new bands ofslowermobility.
The upper band is not observed when low concentrations of32P-labeled
DNA are used forbinding,
whichimplies
thatthis band results from interactions between two DNA
strands.
Proof that the
slower-migrating
band inFig.
5 representscross-linking
ofpRlPvuIIfragments
isprovided
by
the datain
Fig.
6. 32P-labeled PvuIIfragments
frompRl
form dif-ferentmobility complexes
with 28K-EBNAdepending
upon the sizeoftheunlabeledfragments
in thereaction.The threedifferent-sized
species
observed areexplained by
cross-linking of the labeledpRl
fragment
toPvuIIfragments
frompRl, pR2, or pR3 and no
cross-linking
with the PvuIIfragmentfrom
pUC8.
These DNAcross-linking
dataprovide
confirming
evidence for amultimeric 28K-EBNAcomplex.
These studies of
28K-EBNA-binding
kinetics andassoci-ation with DNA suggest mechanisms for
regulation by
full-length
native EBNA-1. If an EBNA-1complex
cross-links DNA in
vivo,
thenonefunction ofEBNA-1 could beto link the tworegions
essential forreplication-the
DNA sequences in the tandem repeats with those in thedyad
symmetryregion. Cross-linking
would formaloop
structure whichmight
be animportant
feature for theregulatory
function. Forexample, topoisomerase activity
couldunwind the DNA in theloop
andprovide
active sites forreplication
or
transcription.
This process could be mediatedby
theEBNA-1 amino-terminal domain
directly
orby
proteins
which interact with this domain.The EBNA-1 amino-terminal domain is linked to the
carboxyl-terminal DNA-binding
domainby
apoly(Gly-Gly-Ala) region of variable length in different EBV isolates.
X-ray diffraction studies ofpoly(Gly-Gly-Ala) indicate that
the
polymer
aggregates into long fibers where the repeatsequences are
fully
extended (9.3 A pertripeptide)
(1, 12, 22). In a multimeric EBNA-1 complex, the 230-amino-acidpoly(Gly-Gly-Ala) region
of eachmonomercould associateinto
a rope structure over 2,000 A in length which could extend over 600 bp in distance. An extended EBNA-1 structure would enable the protein complex to interact in processes at considerable distances from the DNA-bindingsite,
agenerally
accepted property of a trans-activatingeucaryotic
regulatory protein. In this regard, EBNA-1 maybesimialrtothe GAL4proteinofSaccharomycescerevisiae
(11).
on November 10, 2019 by guest
http://jvi.asm.org/
HIGHLY CONCENTRATION DEPENDENT EBNA BINDING TO DNA ACKNOWLEDGMENTS
This workwasaidedby grant MV-287fromtheAmerican Cancer Society, and Public Health Service grants ES03131 from the Na-tional Institute of Environmental Health Sciences and GM32950 from the National Instituteof General Medical Sciences.
LITERATURE CITED
1. Andries, J. C., J. M. Anderson, and A. G. Walton. 1971. Morphological and structural studies of poly(gly-gly-ala). Biopolymers10:479-485.
2. Berg,0.G., R. B. Winter,and P. H. von Hippel. 1982. How do genome-regulatory proteins locate their DNA target sites. TrendsBiochem. Sci. 7:52-55.
3. Better, M., S. Wickner, J. Auerbach, and H. Echols. 1982. Site-specificDNAcondensation andpairing mediatedby the Int protein of bacteriophage lambda. Proc. Natl. Acad. Sci. USA 79:5837-5841.
4. Better, M., S. Wickner, J. Auerbach, and H. Echols. 1983. Role oftheXis protein of bacteriophagelambdainaspecificreactive complexatthe attRprophage attachment site. Cell 32:161-168. 5. Echols, H. 1985. Specialized nucleoprotein structures in
high-fidelityDNAtransactions. BioEssays 1:148-152.
6. Echols, H., M. Dodson, M. Better, J. D. Roberts, and R. McMacken. 1984. The roleof specialized nucleoprotein struc-turesinsite-specific recombination andinitiation of DNA rep-lication. Cold Spring Harbor Symp. Quant. Biol. 49:727-733. 7. Fuller, R. S., B. E.Funnell, and A. Kornberg. 1984. The dna A
protein-DNA complex atoriC and other sites: structure and specificity. Cell 38:889-900.
8. Halprin, J., A. L.Scott, L. Jacobson, P. H. Levine, J. H. Ho, J. C. Niederman, S. D. Hayward, and G. Milman. 1986. En-zyme-linked immunosorbent assay of antibodies to Ep-stein-Barr virus nuclear and early antigens in patients with infectious mononucleosis and nasopharyngeal carcinoma.Ann. Intern. Med. 104:331-337.
9. Hearing, J. C., and A. J. Levine. 1985.TheEpstein-Barrvirus nuclear antigen (BamHI K antigen) is a single-stranded DNA binding phosphoprotein. Virology 145:105-116.
10. Hummel, M., M. Arsenakis, A. Marchini, L. Lee, B. Roizman, and E. Kieff. 1986. Herpes simplex virus expressing Ep-stein-Barrvirus nuclearantigen1. Virology 148:337-348. 11. Keegan, L., G. Gill, and M. Ptashne. 1986.Separation ofDNA
binding from thetranscription-activation function ofa eukary-oticregulatory protein. Science231:699-704.
12. Lotz, B., and H. D. Keith. 1971. The crystal structures of
poly(L-ala-gly-gly-gly)IIandpoly(L-ala-gly-gly)II.J.Mol. Biol.
61:195-200.
12a.Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: a laboratory manual, p. 97-148. Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.
13. Messing, J., and J. Vieira. 1982. The pUC plasmids, an M13mp7-derivedsystemforinsertion mutagenesis and sequenc-ing withsynthetic universal primers. Gene19:259-268. 14. Milman, G., A. L.Scott,M.S. Cho, S. C. Hartman, D. K. Ades,
G. S. Hayward, P. F. Ki, J. T. August, and S. D. Hayward. 1985. Carboxyl-terminal domain ofthe Epstein-Barr virus nu-clearantigen ishighly immunogenic inman. Proc. Natl. Acad. Sci. USA 82:6300-6304.
15. Rawlins, D. R., G.Milman, S.D.Hayward, and G. S.Hayward.
1985.Sequence-specificDNAbinding of the Epstein-Barr virus nuclear antigen (EBNA-1) to clustered sites in the plasmid maintenance region. Cell42:859-868.
16. Reisman, D., J. Yates, and B.Sugden.1985.Aputativeorigin of replication of plasmids derived from Epstein-Barr virus is
composed of two cis-acting components. Mol. Cell. Biol. 5: 1822-1832.
17. Robert, M. F., D. Shedd, R. J. Weigel, D. K. Fischer, and G. Miller. 1984. Expression in COS-1 cells of Epstein-Barr virus nuclearantigen from acomplete gene and adeleted gene. J. Virol.50:822-831.
18. Sculley, T. B., T. Kreofsky, G. R. Pearson, and T. C. Spelsberg. 1983. Partial purification of the Epstein-Barr virus nuclear antigen(s). J. Biol. Chem. 258:3974-3982.
19. Sugden, B. 1982.Epstein-Barr virus: ahuman pathogen induc-inglymphoproliferation in vivo andin vitro. Rev. Infect. Dis. 4:1048-1061.
20. Sugden, B., K. Marsh, and J. Yates. 1985. A vector that replicates as a plasmid and can be efficiently selected in B-lymphoblasts transformed by Epstein-Barr virus. Mol. Cell Biol. 5:410-413.
21. Tooze, J. (ed). 1980. DNA tumorviruses, 2nd ed. ColdSpring HarborLaboratory, ColdSpring Harbor,N.Y.
22. Traub, W.1969. Polymers of tripeptidesascollagen model. V. An x-ray study ofpoly(L-prolyl-glycyl-glycine). J. Mol. Biol. 43:479-485.
23. Yates, J., N. Warren, D. Reisman, and B. Sugden. 1984. A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci. USA81:3806-3810. 24. Yates, J.L.,N.Warren, and B.Sugden.1985.Stablereplication
ofplasmids derived fromEpstein-Barrvirus in various
mamma-liancells.Nature (London) 313:812-815.
25. Zur Hausen, H. 1981.Oncogenicherpesviruses, p. 747-795. In J. Tooze(ed.),DNAtumorviruses, 2nd ed. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
471 VOL.61, 1987