0022-538X/08/$08.00⫹0 doi:10.1128/JVI.02720-07
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Reduction in Severity of a Herpes Simplex Virus Type 1 Murine
Infection by Treatment with a Ribozyme Targeting the
U
L
20 Gene RNA
䌤
Jia Liu,
1Alfred S. Lewin,
1Sonal S. Tuli,
2Steven C. Ghivizzani,
3Gregory S. Schultz,
4and David C. Bloom
1*
Departments of Molecular Genetics and Microbiology,1Ophthalmology,2Orthopedics,3and Obstetrics and Gynecology,4
University of Florida College of Medicine, Gainesville, Florida 32610-0266
Received 21 December 2007/Accepted 16 May 2008
Hammerhead ribozymes were designed to target mRNA of several essential herpes simplex virus type 1 (HSV-1) genes. A ribozyme specific for the late gene UL20 was packaged in an adenovirus vector (Ad-UL20 Rz) and evaluated for its capacity to inhibit the viral replication of several HSV-1 strains, including that of the wild-type HSV-1 (17synⴙand KOS) and several acycloguanosine-resistant strains (PAAr5, tkLTRZ1, and ACGr4) in tissue culture. The Ad-UL20 Rz was also tested for its ability to block an HSV-1 infection, using the mouse footpad model. Mouse footpads were treated with either the Ad-UL20 Rz or an adenoviral vector expressing green fluorescent protein (Ad-GFP) and then infected immediately thereafter with 104PFU of HSV-1 strain 17synⴙ. Ad-UL20 ribozyme treatment consistently led to a 90% rate of protection for mice from lethal HSV-1 infection, while the survival rate in the control groups was less than 45%. Consistent with this protective effect, treatment with the Ad-UL20 Rz reduced the viral DNA load in the feet, the dorsal root ganglia, and the spinal cord relative to that of the Ad-GFP-treated animals. This study suggests that ribozymes targeting essential genes of the late kinetic class may represent a new therapeutic strategy for inhibiting HSV infection.
Herpes simplex virus type 1 (HSV-1) is a neurotropic virus that establishes a lifelong latent infection within the sensory nerve ganglia of humans, after it has been acquired by infection of the oral mucosa or eye. Reactivation from latency occurs sporadically and can result in either asymptomatic shedding or a lesion at the primary site of infection. While the majority of HSV-1 infections are subclinical, in some individuals, reacti-vation of the latent virus can cause recurrent keratitis and corneal scarring.
HSV keratitis is the most common cause of corneal blind-ness in the United States (17), emphasizing the necessity for improved treatment strategies, especially since traditional vac-cines are not beneficial in preventing initial HSV infection or recurrences (21). While topical nucleoside analogue-based an-tiviral medications are effective in treating acute epithelial infections, they can be toxic to corneal and conjunctival cells, and some strains of HSV develop resistance. In the present study, we investigated the use of virally vectored ribozymes (Rz) engineered to cleave essential HSV genes as an alterna-tive therapeutic strategy.
Ribozymes are RNA enzymes that promote a variety of reactions, usually involving cleaving or splicing of RNA mole-cules (5, 22). While ribozymes are generally not as potent or convenient as RNA interference using small interfering RNAs (siRNAs), they are not known to initiate “off-target” effects through the binding of short sequences in the 3⬘untranslated region of unintended mRNAs, a common hazard with siRNAs.
Effective ribozyme inhibitors can be identified through a com-bination of cell-free kinetic testing and cell-based functional assays.
HSV-1 carries more than 80 genes, which are organized into three temporal expression classes, immediate early (alpha), early (beta), and late (gamma) (30). The goal of this study was to evaluate the effects of targeting essential HSV-1 lytic gene RNAs for degradation, using hammerhead ribozymes. Since even the most efficient ribozyme would not be predicted to cleave 100% of the targeted viral RNA, we chose to identify the viral lytic gene mRNAs whose cleavage by ribozymes would result in a significant reduction in viral yield. We therefore designed hammerhead ribozymes to cleave mRNAs of four HSV-1 essential genes (ICP4, UL20, UL30, and UL54), and these ribozymes were tested for their abilities to cleave the RNA targets in vitro and in vivo. Based on these initial cleav-age analyses, the ribozyme that was ultimately selected tar-geted the gene product for the HSV late gene UL20. An
ad-enoviral vector was then used to express this ribozyme in cell culture or in mice to assess the ability of this ribozyme to inhibit HSV-1 replication.
Although the mechanism of HSV-1 virus maturation and egress to the extracellular space is not fully understood, it has been shown that UL20 protein, an essential gene product, plays
an important role in this aspect of viral replication in cell culture (1). The HSV-1 UL20 gene is highly conserved in
al-phaherpesviruses, e.g., varicella-zoster virus (4), bovine her-pesvirus 1 (29), and pseudorabies virus (15), as well as in the gammaherpesvirus Marek’s disease virus type 2 (12). The UL20 open reading frame, which is positionally conserved
among alphaherpesvirus genomes, encodes a 222-amino-acid nonglycosylated membrane protein and is regulated as a ␥1
* Corresponding author. Mailing address: Department of Molecular Genetics and Microbiology, Box 100266, University of Florida College of Medicine, Gainesville, FL 32610-0266. Phone: (352) 392-8520. Fax: (352) 392-3133. E-mail: [email protected].
䌤Published ahead of print on 28 May 2008.
7467
on November 8, 2019 by guest
http://jvi.asm.org/
gene. Deletion of the HSV-1 UL20 and the pseudorabies virus UL20 genes has been shown to reduce infectious virus
produc-tion by up to 100-fold compared to that of wild type (1, 6, 9). In the absence of the UL20 protein, virions are trapped in the
perinuclear space, as well as in cytoplasmic vesicles of the host cell, and therefore, infectious virions are not released to the extracellular space. Although it has been recognized as a mem-brane protein, the UL20 protein is also involved in Golgi
ap-paratus-dependent glycosylation and cell surface expression of glycoprotein K; both glycoprotein K and UL20 are required for
syncytium formation during HSV-1 infection. Therefore, UL20 is also involved in virus-induced cell fusion.
Previous attempts to inhibit HSV-1 replication by gene tar-geting have focused largely on inhibiting immediate early gene expression, e.g., that of ICP4 (27, 28). In this study, the impact of knocking down a late gene of HSV-1 was addressed. Using an adenoviral vector to express a hammerhead ribozyme tar-geting UL20 RNA (Ad-UL20 Rz), expression of the UL20 gene
was significantly reduced, leading to an inhibition of HSV-1 viral replication in vitro and in vivo. These findings suggest a possible new approach for therapy for HSV-1 infections.
MATERIALS AND METHODS
Ribozyme kinetics in vitro.We selected several genes as ribozyme targets, based on their requirements for viral replication (18). Cleavage sites for ham-merhead ribozymes were selected as previously described (23), and two ri-bozymes were tested for each target mRNA. Synthetic RNA oligonucleotides comprising hammerhead ribozymes and corresponding target sequences were purchased from Dharmacon, Inc. (Lafayette, CO). The cleavage activity of each ribozyme was studied kinetically, as described previously (8).
Ribozyme cloning and packaging in an adenoviral vector.Ribozymes were cloned into the pAdlox shuttle plasmid (GenBank accession number R62024) for packaging into an adenoviral vector. The hammerhead ribozyme was cloned between the cytomegalovirus promoter sequence and the simian virus 40 poly(A) signal. Following cloning of the hammerhead ribozyme, we inserted a hairpin ribozyme that separates the antiviral hammerhead ribozyme from a downstream internal ribosome entry site-green fluorescent protein (IRES-GFP) element. The
shuttle plasmid was then inserted into the5 adenoviral vector backbone by a
cis-acting replication element (CRE)-loxrecombination. The recombinant
vec-tors were then propagated in cre8 293 cells (11, 20). Adenoviral purification was conducted using a Vivapure AdenoPACK 100 (Vivascience AG, Hannover, Germany). The virus particle concentration and the 293 cell infectious titer of the adenovirus stock were determined as described previously (20). Two
adenovi-ruses,5, the parental vector for transgene packaging (11), and Ad-GFP, which
expresses GFP from the5 backbone, were used as controls in this study.
HSV-1 preparation.A low-passage stock of HSV-1 strain 17syn⫹was pre-pared from a master stock obtained from J. Stevens (UCLA). The drug-resistant strains of HSV-1, PAAr5 (2, 13), tkLTRZ1 (3, 14), and ACGr4 (2), as well as their parental strain, KOS, were generously provided by D. Coen (Harvard Medical School). The HSV-1 virus was propagated and titrated on rabbit skin cells grown in minimal essential medium supplemented with 5% calf serum and antibiotics (26).
Ribozyme inhibition of multiple-step HSV-1 replication.All infections were performed in triplicate. Three groups were included to test the effect of the
ribozyme in a wild-type HSV-1 infection: mock treatment,5 (backbone
ade-novirus), and Ad-UL20 Rz. Each experiment was performed in triplicate. Rabbit
skin cells were seeded at a density of 2⫻105cells per well (3.5 cm2), followed
by an adenoviral inoculation at a dose of 7.5⫻104
virus particles per cell for 15 h.
The infection of HSV-1 (17syn⫹) was conducted at a multiplicity of infection
(MOI) of 10⫺3
for 24 h before cell lysates were harvested for titration. Titration was conducted by serial dilution of cell lysates, and PFU were counted on rabbit skin cells, in triplicate for each dilution. After the yields were calculated, average infectious yield values and standard deviations were calculated from the three separate replicates of each experiment.
Four groups were included to test the impact of the vector-expressed ribozyme
on the replication of drug-resistant HSV-1 strains: mock infection and Ad-UL20
Rz treatments were performed as described before; an adenovirus expressing
GFP was used instead of5, and an acyclovir (ACV) treatment (0.1M) was
included. The dosage of acyclovir was determined by the threshold dose needed to distinguish drug resistance of HSV-1 (25). Forty-eight hours after infection with HSV-1, cell lysates were harvested for titration. Five HSV-1 strains were
tested: the drug-sensitive strains 17syn⫹and KOS and the drug-resistant isolates
PAAr5, tkLTRZ1, and ACGr4.
Ribozyme inhibition of UL20 expression.We evaluated the effect of the
vector-expressed UL20 ribozyme on single-step growth of HSV-1 (strain 17syn⫹). Each
experiment was conducted in triplicate, and each experiment included three
treatments: mock infection, Ad-GFP, and Ad-UL20 Rz. After Ad-UL20 Rz was
inoculated as described above, HSV-1 infection was conducted at an MOI of 3 for 8 h before cell lysates were harvested using TRIzol reagent (Invitrogen, Carlsbad, CA). Total RNA and DNA were separated by following the manufac-turer’s recommendation, and total RNA was treated with RNase-free DNase (10) to remove the DNA contamination. Reverse transcription was conducted using a First-Strand cDNA synthesis kit (Amersham Biosciences, Piscataway, NJ), followed by real-time PCR (16). Specific primers and probes for either the viral genes (Table 1) or the rabbit GAPDH (glyceraldehyde-3-phosphate dehy-drogenase) gene were synthesized by Applied Biosystems (Foster City, CA).
Testing the ribozyme inhibition of HSV-1 encephalitis in mice.Six-week-old female ND4 Swiss mice (Harlan-Sprague Dawley, Inc.) were infected via the rear footpads as previously described (16). Three groups of 10 mice were employed,
and two independent experiments were conducted. First, 1.4⫻1010
virus
par-ticles of adenovirus (expressing either UL20 Rz or GFP) or phosphate-buffered
saline (PBS) in a volume of 10l per footpad was combined with 40l of 10%
sterile saline solution and injected into both rear footpads. After 3 to 4 h, mice
were inoculated again with 1.4⫻1010
virus particles of adenovirus (UL20 Rz or
GFP) or PBS applied topically to each rear footpad following light scarification of the footpad with an emery board. This second application of adenovirus or
PBS was immediately followed by a topical application of HSV-1 (17syn⫹) at a
dose of 104
PFU per footpad.
RESULTS
Kinetic analysis of HSV-1 ribozymes.Initially, four essential HSV-1 genes were chosen as ribozyme targets, ICP4, UL20,
UL30/DNA polymerase, and UL54/ICP27. Two ribozyme
[image:2.585.44.543.80.156.2]cleavage sites were selected for each gene, based on their moderate G⫹C content and their predicted accessibility within the mRNA (33) and on other established design and validation parameters for therapeutic ribozymes (7). Sequences of ri-bozymes and their cleavage sequences are shown in Table 2. The ribozymes were evaluated for their catalytic efficiency in vitro on synthetic target RNAs, using time course and multi-ple-turnover kinetic analysis. The time course of cleavage was
TABLE 1. Real-time PCR primers and probes
DNA target Sequence (5⬘33⬘) Accession no. (nucleotide no.)
HSV-1 UL20 CCA TCG TCG GCT ACT ACG TTA C (forward) X14112 (41118–41187)
CGA TCC CTC TTG ATG TTA ACG TAC A (reverse) CCC GCA CCG CCC AC (probe)
HSV DNA polymerase (UL30) AGA GGG ACA TCC AGG ACT TTG T (forward) X14112 (65880–65953) CAG GCG CTT GTT GGT GTA C (reverse)
ACC GCC GAA CTG AGC A (probe)
on November 8, 2019 by guest
http://jvi.asm.org/
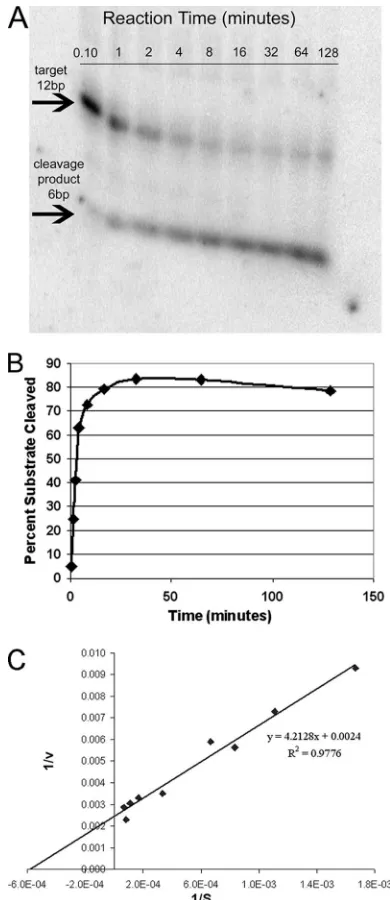
established at a 10-fold excess of substrate relative to the amount of ribozyme and at a Mg2⫹concentration of 5 mM, unless the rate of cleavage was low, in which case 20 mM Mg2⫹ was also employed. The UL20 Rz154, for example, led to 75% cleavage of its RNA target by 8 min in the presence of low Mg2⫹(Fig. 1). The dependence of the initial rates of cleavage on substrate concentration was then determined and analyzed using Lineweaver-Burk double-reciprocal plots to determine
Km, kcat(Vmax/[Rz]), andkcat/Kmvalues (Fig. 1C). The value forkcat/Km, which is an estimate of catalytic efficiency, ranged from 0.01 for UL20 Rz135 to 15.9 for UL20 Rz154 (Table 3). For each HSV-1 target gene, the most efficient ribozyme was chosen for analysis as an inhibitor of viral replication.
The HSV-1 UL20 ribozyme was the most efficient at inhib-iting HSV-1 replication in vitro.Synthetic ribozymes for each of the four genes were then transfected into rabbit skin cells and tested for their abilities to reduce viral yields following infection with HSV-1, as described in Materials and Methods. These analyses demonstrated that the UL20 ribozyme consis-tently resulted in the greatest reduction in viral yields (data not shown). Ribozymes targeting immediate early genes (ICP4 and UL54) did not lead to a significant inhibition in viral replica-tion, but ICP4-Rz885 mRNA displayed low catalytic efficiency (kcat/Km⫽0.3), which may explain its low inhibition of repli-cation.
[image:3.585.43.547.80.174.2]The vector-expressed ribozyme targeting the HSV-1 UL20 mRNA significantly reduced wild-type viral replication in rab-bit skin cells. Based on a screening of the above-described results, the UL20 Rz154 ribozyme was determined to be the most efficient at inhibiting HSV-1 infection following transfec-tion. Therefore, a DNA cassette expressing UL20 Rz154 was cloned into an adenoviral vector in order to provide a more
[image:3.585.63.258.207.657.2]FIG. 1. In vitro kinetic analysis of hammerhead ribozyme UL20-154. (A) Autoradiogram of the time course of cleavage of an RNA target (end labeled with [␥-32P]ATP) by ribozyme UL20-154 at a mag-nesium concentration of 5 mM. (B) The percentage of target RNA cleavage in each time point was calculated by quantification of cut and uncut target bands shown in panel A. (C) Lineweaver-Burk plot of UL20 Rz154 cleavage of the synthetic HSV RNA target.
TABLE 2. Ribozyme sequences and sequences of their respective targets
Ribozymea Ribozyme sequence Target sequence
ICP4-885 ACGAACUGAUGAGCGCUUCGGCGCGAAAGGAUG CAUCCUCUUCGU
ICP4-533 UCGAUCUGAUGAGCGCUUCGGCGCGAAACGCCG CGGCGUCAUCGA
UL20-135 GAACUCUGAUGAGCGCUUCGGCGCGAAACAAAA UUUUGUCAGUUC
UL20-154 CGGAACUCAUGAGCGCUUCGGCGCGAAACGCGA UCGCGUCUUCCG
UL30-933 AAGGUCUGAUGAGCGCUUCGGCGCGAAACGAAC GUUCGUCACCUU
UL30-1092 CACAUCUGAUGAGCGCUUCGGCGCGAAAGCUUG CAAGCUCAUGUG
UL54-233 UUCUGCUGAUGAGCGCUUCGGCGCGAAACGAGA UCUCGUCCAGAA
UL54-825 UGCAUCUGAUGAGCGCUUCGGCGCGAAACCUGU ACAGGUCAUGCA
aThe ribozyme name is composed of the name of the target gene (e.g., ICP4) and the cleavage site nucleotide (e.g., 885).
TABLE 3. Summary of in vitro kinetic analyses of hammerhead ribozymes designed against HSV-1a
HSV target
gene Mg
2⫹(mM) k
cat(min⫺1) Km(M)
kcat/Km(M⫺1
min⫺1)
ICP4–885 20 15.87 52.83 0.3
ICP4–533 5 and 20b NA NA NA
UL20-135 20 0.08 5.64 0.01
UL20-154 5 27.78 1.75 15.9
UL30-933 20 9.26 2.57 3.6
UL30-1092 5 22.99 23.59 1.0
UL54-233 5 0.91 8.58 0.1
UL54-825 5 51.28 4.44 11.7
a
NA, no activity detected.
b
No activity was detected at either Mg2⫹concentration.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.301.543.606.707.2]efficient means of delivering this ribozyme for subsequent anal-yses. This adenoviral vector (Ad-UL20 Rz154) was then tested for its ability to inhibit HSV-1 replication in rabbit skin cells. HSV-1 infection was conducted at a low MOI (10⫺3), and the
ribozyme was used to inhibit multistep HSV-1 viral replication. Ad-UL20 Rz154 reduced HSV-1 wild-type viral replication by 83% compared with that of the control treatment (5) (P ⬍
0.001), and no significant differences were observed between the groups that received no treatment and those that received
5 treatments (Fig. 2).
Ad-UL20 treatment blocked the UL20 gene expression dur-ing HSV-1 infection of rabbit skin cells.Ad-UL20 Rz154 was also tested with a one-step replication assay of HSV-1 follow-ing infection at an MOI of 3. As UL20 is a late gene, an 8-h incubation was used to allow completion of late gene expres-sion. The effect of UL20 Rz154 on UL20 expression at the mRNA level was assayed by reverse transcription, followed by
real-time PCR. A 68% reduction in UL20 mRNA level was
detected by comparing results of the Ad-ribozyme treatment with that of the Ad-GFP treatment (P⬍ 0.0005) (Fig. 3B). There was also a 50% reduction in UL20 mRNA level using Ad-GFP treatment relative to that of mock infection (P ⬍
0.0004). It is therefore likely that preinfection of the cells with adenoviral vectors reduces the replication of HSV-1. DNA was also extracted from each infected sample, and the viral DNA level normalized to that of the endogenous cellular control (GAPDH) was also determined using real-time PCR (Fig. 3A). A 54% reduction in viral DNA level was observed for the ribozyme treatment group compared to that of the Ad-GFP group (P ⬍ 0.004), but there were no significant differences between the DNA levels of the mock infection and those of the Ad-GFP treatments.
Ad-UL20 Rz154 inhibits viral replication of HSV-1 drug-resistant strains.Treatment with Ad-UL20 Rz154 was com-pared with that with acyclovir, using multistep infection at an MOI of 10⫺3. Ad-U
L20 Rz154 significantly inhibited wild-type
HSV-1 viral production (by 98% against 17syn⫹and by 95% against KOS), and acyclovir treatment led to similar levels of inhibition (99% and 80%, respectively). However, acyclovir
treatment was not effective against drug-resistant strains of HSV-1 (no significant differences in HSV yields), while the UL20 ribozyme maintained its inhibitory effect. In the case of
PAAr5 infection, the UL20 ribozyme reduced viral production by 99%; reductions were 76% for tkLTRZ1 infection and 70% for ACGr4 infection (Fig. 4). Therefore, knocking down the expression of the UL20 gene led to a significant inhibition of
HSV-1 viral replication in viral strains that were resistant to a conventional antiviral drug. It should be noted that the 70- to 100-fold reduction of viral yields shown in Fig. 4A and B are greater than those shown in Fig. 2 because the infectious virus was harvested at 48 h in the experiments shown in Fig. 4 as opposed to 24 h as shown in Fig. 2.
The vector-expressed UL20 ribozyme protects mice following footpad infection with HSV-1.To evaluate the impact of block-ing the expression of the UL20 gene in vivo, Ad-UL20 Rz154 was delivered to the mouse footpad prior to infection with HSV-1. Viral particles (1.4⫻1010) of either Ad-U
L20 Rz154
or Ad-GFP were injected into both rear footpads. Three hours later, mice were challenged with HSV-1 (104PFU of 17syn⫹
[image:4.585.45.284.68.226.2]per footpad). Beginning at day 6 postinfection, mice from the
FIG. 2. Inhibitory effect of Ad-UL20 Rz154 on wild-type HSV-1 viral replication. At day 1 (24 h) after infection with HSV-1 at an MOI of 10⫺3, Ad-U
[image:4.585.305.537.69.392.2]L20 Rz154 inhibited wild-type HSV-1 viral replication by 83% compared with that of the adenovirus vector (without ribozyme) control treatment (5).
FIG. 3. Ad-UL20 Rz led to a significant reduction in UL20 RNA and viral DNA levels. (A) Real-time PCR was used to detect viral DNA relative to that of total recovered cellular DNA. A ratio of the viral DNA polymerase gene to the cellular GAPDH gene was used to determine the relative abundance of viral DNA. (B) Reverse transcrip-tion followed by real-time PCR was conducted to study the UL20 mRNA level. A ratio of the amount of viral UL20 mRNA to the cellular GAPDH RNA was used to indicate the abundance of UL20 mRNA.
on November 8, 2019 by guest
http://jvi.asm.org/
control groups (receiving Ad-GFP and PBS treatments) showed signs of encephalitis, including hind-limb paralysis, hunched posture, ruffled fur, ataxia, and weakness. Mice that eventually became nonresponsive and could not ambulate were euthanized. However, mice from the ribozyme treatment group remained healthy and active overall (though at a much later time point, two deaths were observed, and one mouse showed mild paralysis). With the HSV ribozyme treatment, 89% of animals survived, while 45% and 40% survival rates were observed for PBS- and Ad-GFP-treated controls, respec-tively (Fig. 5). Kruskal-Wallis nonparametric analysis showed significant differences among the three groups (P⫽0.04).
To further investigate the ribozyme’s impact on viral repli-cation in vivo, an independent experiment was performed in which four or five mice per group (receiving the Ad-ribozyme or Ad-GFP control treatment, respectively) were sacrificed at 6 days postinfection, and the feet, dorsal root ganglia, and spinal cords were dissected. Real-time PCR was conducted to compare viral DNA levels from different tissues. While Ad-ribozyme treatment resulted in decreased HSV-1 viral DNA recovery from all tissues, the decrease did not reach statistical significance. In the spinal cord assay, where day 6 represents the peak of viral DNA loads, Ad-ribozyme treatment reduced the amount of viral DNA by sixfold (Fig. 6).
DISCUSSION
In this study, hammerhead ribozymes were designed to limit the expression of several HSV-1 essential genes (ICP4, UL54, UL30, and UL20). In vitro kinetic analysis led to the selection
of ribozymes with the greatest potential for efficient target cleavage. Because the UL20 Rz154 exhibited the most robust
inhibition of viral replication following transfection, it was cloned into an Ad5 vector and tested further in vitro and in vivo.
Our best target and best ribozyme was the UL20 gene, a␥1
gene, encoding a membrane protein that is essential for viral intra- and extracellular egress, as well as intracellular transport of viral glycoproteins. The ribozyme targeting UL20 mRNA, which also likely disrupts UL20.5 mRNA (32), not only
re-duced the replication of wild-type HSV-1 (in strains 17syn⫹
[image:5.585.68.261.67.715.2]and KOS) but also inhibited the replication of drug-resistant HSV-1 strains (PAAr5, tkLTRZ1, and ACGr4). Data pre-sented here confirm that the UL20 gene product of HSV-1 is
FIG. 4. Ad-UL20 Rz is effective in inhibiting acyclovir-resistant strains of HSV-1. After treatments, cells were infected with the indi-cated HSV strain at an MOI of 10⫺3for 48 h. (A) Ad-ribozyme treatment led to a significant reduction (by 98%) in 17syn⫹ viral replication compared with Ad-GFP treatment (P⬍0.002), and acy-clovir treatment had very similar inhibitory effect (99% reduction;P⬍ 0.02). (B) Ad-ribozyme inhibited the viral replication of HSV-1 strain KOS by 95% compared with Ad-GFP treatment (P ⬍ 0.05), while ACV inhibited KOS by 80% (P⬍0.02). (C) The HSV-1 drug-resistant strain PAAr5 could be inhibited by ribozyme (99% reduction;P ⬍ 0.005) but not by ACV. (D) The drug-resistant strain tkLTRZ1 viral replication was inhibited by ribozyme by 76% (P⬍0.05), while there was no effect from ACV. (E) The double-mutant strain ACGr4 viral replication was inhibited by ribozyme by 70% (P⬍0.006), while ACV had no effect.
on November 8, 2019 by guest
http://jvi.asm.org/
essential for the virus life cycle in rabbit skin cells and that decreasing the expression of this gene leads to a reduction of viral production. Since UL20 is a late gene involved in virion maturation, our results indicate that these steps, including viral egress, may be suitable targets for antiviral intervention.
A control adenovirus, Ad-GFP, was included to assess vec-tor effects. When UL20 mRNA levels were evaluated for the ribozyme effect, Ad-GFP treatment led to an approximately 50% reduction (Fig. 3B), and a similar reduction was observed
at the level of mRNA for the viral DNA polymerase (data not shown). The Ad vectors we employed contained deletions in the E1 and E3 regions but still contained viral late genes. It is possible that Ad-GFP competed with HSV-1 for the usage of cellular machinery, e.g., RNA polymerase II, indirectly leading to a lower level of viral gene expression in the Ad-GFP treat-ment group. Overall, in spite of the nonspecific effect of ade-novirus, UL20 Rz154 reduced viral target gene expression and
inhibited virus replication significantly compared with the Ad-GFP treatment.
An interesting finding from this study was the significant reduction in viral DNA yields observed when the UL20
ri-bozyme was tested with a single-step infection assay (Fig. 3A). It has been previously shown that UL20 expression is dimin-ished when HSV-1 DNA replication is blocked, consistent with the designation of UL20 as a late gene (31). It was therefore surprising that the inhibition of this late gene by the ribozyme following a high MOI resulted in a decrease in viral DNA accumulation. These results suggest that UL20 expression pro-vides a feedback signal to viral DNA synthesis.
[image:6.585.123.463.68.271.2]The Ad-UL20 Rz154 ribozyme was capable of inhibiting the replication of acyclovir-resistant strains of HSV-1 (PAAr5, tkLTRZ1, and ACGr4) (Fig. 4), which were chosen because they represent two general drug resistance mechanisms: mu-tations in thymidine kinase and in DNA polymerase. Most of the antiviral drugs currently approved for treatment of HSV-1 infection are nucleotide analogs, which are either substrates of thymidine kinase that indirectly disrupt viral DNA synthesis or are incorporated into the elongated DNA strand and lead to premature termination. The treatment of patients, especially those with immune deficiency and repeated reactivations, with antivirals has led to the selection of drug-resistant viruses that results in the uncontrolled spread of HSV-1 infection. In some cases, these infections are lethal to patients. It is encouraging that a nucleic acid-based approach such as a ribozyme targeting
FIG. 5. Ad-UL20 Rz limits the lethality of HSV-1 in the mouse footpad model. Mouse rear footpads were pretreated with subcutaneous injections of PBS, Ad-GFP, or Ad-UL20 Rz and received a second topical application 4 h later, followed immediately by infection with HSV-1 at a dose of 104PFU per footpad. Kruskal-Wallis nonparametric analysis shows significant differences among the three group, with aPvalue of 0.04; n⫽10 mice per group.
FIG. 6. Adenovirus (expressing either Ad-UL20 Rz or Ad-GFP) was inoculated on both rear footpads of mice, which were then infected with HSV-1. At 6 days postinfection of HSV-1, five mice from the control group and four from the ribozyme treatment group were sac-rificed to collect tissue samples. Viral DNA levels (relative to that of GAPDH) in the spinal cord were assessed in these two groups by real-time PCR. Squares represent relative viral DNA levels from the control group; circles represent relative viral DNA levels from the Ad-ribozyme treatment group; bars indicate the mean of each group.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.44.285.463.641.2]mRNA of HSV-1 UL20 might overcome this resistance. Although nucleotide changes can cause the emergence of resistant escape mutants for the UL20 ribozyme, ribozymes targeting different essential genes of HSV-1 can be combined to guarantee their inhibition, e.g., by combining ribozyme-targeting immediate early genes and early and late essential genes. The use of a ribozyme-based therapy may be particularly applicable to HSV infections of the cornea, where the toxicity of the currently available antivirals poses a problem.
The mouse footpad infection model offers an efficient ap-proach with which to study HSV-1 viral neuroinvasion, neuro-virulence, and latency. We applied Ad-UL20 Rz154 to block the initial replication in footpad epithelium, thereby reducing the retrograde transport of HSV-1 to the dorsal root ganglion (24). At an inoculum of 104PFU of wild-type HSV-1, the viral
replication in the footpad epithelium led to severe damage in the central nervous system, which caused death in 60% of our control animals. Pretreating the mouse footpad with Ad-ri-bozyme consistently led to a significant level of protection (an overall 89% survival rate) against the HSV-1 challenge. De-spite the HSV challenge, most animals in this group remained healthy throughout the study; one mouse showed mild paral-ysis of the hind limbs but remained active after the study’s end point. In contrast, in both of the control groups, death and the indication of severe damage in the central nervous system were observed.
Interestingly, the ribozyme targeting a late essential gene, UL20, achieved the most significant therapeutic effect against HSV-1 infection in vitro and in vivo, in this study. Palliser et al. designed siRNAs against HSV-2 UL27 and UL29, which
en-code an envelope glycoprotein and a DNA binding protein, respectively (19). Their results indicate that suppressing the expression of early/late genes of HSV has a greater impact on the virus lytic life cycle than inhibiting the expression of im-mediate early genes. Our results here demonstrated that tar-geting a late gene was more effective than tartar-geting genes of the other kinetic classes. We hypothesize that significantly in-hibiting viral protein production of genes encoding structural proteins or virion maturation might produce a more profound effect than inhibiting regulatory genes, whose products are required in smaller and nonstoichiometric amounts. We have extended this finding to demonstrate that a nucleic acid-based therapeutic agent (UL20 Rz-154) targeting a late gene could inhibit the viral replication of drug-resistant HSV-1 strains and could limit the lethal effects of an HSV-1 infection in vivo. Our next step is to develop a delivery system that will permit long-lasting expression of such therapeutics in the corneal epithe-lium.
ACKNOWLEDGMENTS
This work was supported by an Investigators in Pathogenesis award from the Burroughs Wellcome Fund (D.C.B.), the Shaler-Richardson Professorship in Ophthalmological Sciences endowment, pilot funding from the UF Research Foundation (A.S.L.), an award from Research to Prevent Blindness (S.S.T.), and grants AR48566 (S.C.G.) and EY05587 (G.S.S.) from the National Institutes of Health.
We thank N. Giordani for helpful comments on the manuscript.
REFERENCES
1.Baines, J. D., P. L. Ward, G. Campadelli-Fiume, and B. Roizman.1991. The
UL20 gene of herpes simplex virus 1 encodes a function necessary for viral
egress. J. Virol.65:6414–6424.
2.Coen, D. M., and P. A. Schaffer.1980. Two distinct loci confer resistance to acycloguanosine in herpes simplex virus type 1. Proc. Natl. Acad. Sci. USA
77:2265–2269.
3.Davar, G., M. F. Kramer, D. Garber, A. L. Roca, J. K. Andersen, W. Bebrin, D. M. Coen, M. Kosz-Vnenchak, D. M. Knipe, and X. O. Breakefield.1994. Comparative efficacy of expression of genes delivered to mouse sensory
neurons with herpes virus vectors. J. Comp. Neurol.339:3–11.
4.Davison, A. J., and J. E. Scott.1986. The complete DNA sequence of
varicella-zoster virus. J. Gen. Virol.67:1759–1816.
5.Fedor, M. J., and J. R. Williamson.2005. The catalytic diversity of RNAs.
Nat. Rev. Mol. Cell Biol.6:399–412.
6.Foster, T. P., J. M. Melancon, J. D. Baines, and K. G. Kousoulas.2004. The herpes simplex virus type 1 UL20 protein modulates membrane fusion events during cytoplasmic virion morphogenesis and virus-induced cell fusion. J.
Vi-rol.78:5347–5357.
7.Fritz, J. J., M. Gorbatyuk, A. S. Lewin, and W. W. Hauswirth.2004. Design and validation of therapeutic hammerhead ribozymes for autosomal
domi-nant diseases. Methods Mol. Biol.252:221–236.
8.Fritz, J. J., D. A. White, A. S. Lewin, and W. W. Hauswirth.2002. Designing and characterizing hammerhead ribozymes for use in AAV vector-mediated
retinal gene therapies. Methods Enzymol.346:358–377.
9.Fuchs, W., B. G. Klupp, H. Granzow, and T. C. Mettenleiter.1997. The UL20 gene product of pseudorabies virus functions in virus egress. J. Virol.
71:5639–5646.
10.Gorbatyuk, M., V. Justilien, J. Liu, W. W. Hauswirth, and A. S. Lewin.2007. Preservation of photoreceptor morphology and function in P23H rats using
an allele independent ribozyme. Exp. Eye Res.84:44–52.
11.Hardy, S., M. Kitamura, T. Harris-Stansil, Y. Dai, and M. L. Phipps.1997.
Construction of adenovirus vectors through Cre-loxrecombination. J. Virol.
71:1842–1849.
12.Hatama, S., H. K. Jang, Y. Izumiya, J. S. Cai, Y. Tsushima, K. Kato, T. Miyazawa, C. Kai, E. Takahashi, and T. Mikami.1999. Identification and DNA sequence analysis of the Marek’s disease virus serotype 2 genes ho-mologous to the herpes simplex virus type 1 UL20 and UL21. J. Vet. Med.
Sci.61:587–593.
13.Jacobson, J., M. Kramer, F. Rozenberg, A. Hu, and D. M. Coen.1995. Synergistic effects on ganglionic herpes simplex virus infections by mutations or drugs that inhibit the viral polymerase and thymidine kinase. Virology
206:263–268.
14.Jacobson, J. G., S. H. Chen, W. J. Cook, M. F. Kramer, and D. M. Coen.
1998. Importance of the herpes simplex virus UL24 gene for productive
ganglionic infection in mice. Virology242:161–169.
15.Klupp, B. G., H. Kern, and T. C. Mettenleiter.1992. The virulence-deter-mining genomic BamHI fragment 4 of pseudorabies virus contains genes corresponding to the UL15 (partial), UL18, UL19, UL20, and UL21 genes
of herpes simplex virus and a putative origin of replication. Virology191:
900–908.
16.Kubat, N. J., A. L. Amelio, N. V. Giordani, and D. C. Bloom.2004. The
herpes simplex virus type 1 latency-associated transcript (LAT) enhancer/rcr
is hyperacetylated during latency independently of LAT transcription. J.
Vi-rol.78:12508–12518.
17.Liesegang, T. J.2001. Herpes simplex virus epidemiology and ocular
impor-tance. Cornea20:1–13.
18.MacLean, C. A., S. Efstathiou, M. L. Elliott, F. E. Jamieson, and D. J. McGeoch.1991. Investigation of herpes simplex virus type 1 genes encoding
multiply inserted membrane proteins. J. Gen. Virol.72:897–906.
19.Palliser, D., D. Chowdhury, Q. Y. Wang, S. J. Lee, R. T. Bronson, D. M. Knipe, and J. Lieberman.2006. An siRNA-based microbicide protects mice
from lethal herpes simplex virus 2 infection. Nature439:89–94.
20.Palmer, G. D., E. Gouze, J. N. Gouze, O. B. Betz, C. H. Evans, and S. C. Ghivizzani.2003. Gene transfer to articular chondrocytes with recombinant
adenovirus. Methods Mol. Biol.215:235–246.
21.Pepose, J. S., T. L. Keadle, and L. A. Morrison.2006. Ocular herpes simplex: changing epidemiology, emerging disease patterns, and the potential of
vac-cine prevention and therapy. Am. J. Ophthalmol.141:547–557.
22.Saldanha, R., G. Mohr, M. Belfort, and A. M. Lambowitz.1993. Group I and
group II introns. FASEB J.7:15–24.
23.Shaw, L. C., P. O. Whalen, K. A. Drenser, W. Yan, W. W. Hauswirth, and A. S. Lewin.2000. Ribozymes in treatment of inherited retinal disease.
Methods Enzymol.316:761–776.
24.Stevens, J. G., and M. L. Cook.1973. Latent infections induced by herpes
simplex viruses. Cancer Res.33:1399–1401.
25.Thi, T. N., C. Deback, I. Malet, P. Bonnafous, Z. Ait-Arkoub, and H. Agut.
2006. Rapid determination of antiviral drug susceptibility of herpes simplex
virus types 1 and 2 by real-time PCR. Antivir. Res.69:152–157.
26.Tran, R. K., P. T. Lieu, S. Aguilar, E. K. Wagner, and D. C. Bloom.2002. Altering the expression kinetics of VP5 results in altered virulence and
pathogenesis of herpes simplex virus type 1 in mice. J. Virol.76:2199–2205.
27.Trang, P., A. Kilani, J. Kim, and F. Liu.2000. A ribozyme derived from the catalytic subunit of RNase P from Escherichia coli is highly effective
in inhibiting replication of herpes simplex virus 1. J. Mol. Biol. 301:
817–826.
on November 8, 2019 by guest
http://jvi.asm.org/
28.Trang, P., J. Lee, A. F. Kilani, J. Kim, and F. Liu.2001. Effective inhibition of herpes simplex virus 1 gene expression and growth by engineered RNase
P ribozyme. Nucleic Acids Res.29:5071–5078.
29.Vlcek, C., V. Benes, Z. Lu, G. F. Kutish, V. Paces, D. Rock, G. J. Letchworth, and M. Schwyzer.1995. Nucleotide sequence analysis of a 30-kb region of the bovine herpesvirus 1 genome which exhibits a colinear gene arrangement with the UL21 to UL4 genes of herpes simplex virus.
Virology210:100–108.
30.Wagner, E. K., J. F. Guzowski, and J. Singh.1995. Transcription of the herpes simplex virus genome during the productive and latent infections, p.
123–168.InW. E. Cohen and K. Moldave (ed.), Progress in nucleic acid
research and molecular biology. Academic Press, San Diego, CA. 31.Ward, P. L., G. Campadelli-Fiume, E. Avitabile, and B. Roizman.1994.
Localization and putative function of the UL20 membrane protein in cells
infected with herpes simplex virus 1. J. Virol.68:7406–7417.
32.Ward, P. L., B. Taddeo, N. S. Markovitz, and B. Roizman.2000. Identifica-tion of a novel expressed open reading frame situated between genes U(L)20
and U(L)21 of the herpes simplex virus 1 genome. Virology266:275–285.
33.Zuker, M.2003. Mfold web server for nucleic acid folding and hybridization
prediction. Nucleic Acids Res.31:3406–3415.