0022-538X/90/0301271-07$02.00/0
CopyrightC)1990, American SocietyforMicrobiology
Localization
on
the
Herpes
Simplex Virus
Type
1
Genome
of
a
Region Encoding
Proteins Involved in Adsorption
to
the
Cellular Receptor
NINA LANGELAND,12* ANNE M. 0YAN,1 HOWARD S. MARSDEN,3 ANN CROSS,3
JOSEPH C. GLORIOSO,4'5 LINDSEY J. MOORE,1 AND LARSHAARR`6
DepartmentofBiochemistryl* and the GadeInstitute, DepartmentsofPathology2 andMicrobiology,6 Universityof
Bergen, Bergen, Norway; Medical Research Council Virology Unit, Glasgow GIl 5JR, Scotland3; and DepartmentofMicrobiology and Immunology4 and the Unitfor Laboratory AnimalMedicine,5
Universityof Michigan Medical School, AnnArbor, Michigan 48109
Received 7August1989/Accepted 11 November 1989
We havepreviouslyshownthataminoglycosidessuch asneomycinand thepolyamino acids polylysine and
polyarginine selectively inhibit the binding ofherpes simplex virus type 1 (HSV-1) to thecellular receptor,
whereas HSV-2 infection is unaffected.In thepresentstudywetook advantageofthisdifference between HSV-1
andHSV-2byusingHSV-1-HSV-2intertypic recombinantstolocatearegionontheHSV-1genomeencoding
proteins affecting the binding of the virion to the cellular receptor. The results were consistent with those
obtainedby markerrescueexperiments. Theidentifiedregion,which mappedbetweencoordinates0.580 and
0.687, containstwo partialandeight completegenes, includingthe glycoprotein C (gC) geneandtwoothers withpotentialtransmembranesequences.Various gC monoclonalantibody-resistantmutantsof HSV-1aswell as amutantcompletely lackinggCwerefoundtobefullysensitivetoneomycin, suggesting that gC isnotthe
site ofdrugsensitivityandisnotessentialfor adsorptionofvirustothe cellularreceptor.However, therateof
adsorption was reduced in the absence of gC, indicating a facilitating function of the glycoprotein. The
universal nature of this HSV-1 receptorbindingwasrevealedbythesimilarityindrug sensitivityofinfectivity
infour differentcell lines fromvarioustissuesandspecies.
The mechanism by which HSV adsorbs to cells is still
poorly understood. It has been generally assumed that because glycoproteins reside on the outside of infectious virions, they are involved in the adsorption process. Seven
herpes simplex virus (HSV) glycoproteins (glycoprotein B
[gB], gC, gD, gE, gG, gH, and gI) have been identified in
HSV-infected cells, and all except gI are present on virions
(26, 34, 38). gIisalsolikely to be presenton virions, since
they possess affinity for the Fc end ofimmunoglobulin G
(48), andthisactivity dependson acomplexof bothgE and
gI (27). Aputative eighth glycoprotein, theproduct ofgene US5 (43),remains tobe identified.
Ofthese glycoproteins, gC, gE, gG, gI, and the putative US5gene product are notessential for infectivity intissue culture andtherefore cannotbe essential foradsorption (9,
13,
20-23,
25, 35, 36, 46, 62, 66). Ofthethreeglycoproteins
essential forinfectivity,gB (33, 53), gD (32), and gH (12, 19,
42, 63), neither gB norgD isrequired foradsorption, since
mutantshavebeen isolatedwhich do not produce gB (6) or gD (32) and which are able to adsorb to cells. Thus, gH remains the only candidate for a glycoprotein essential in
adsorption.
None of the above results exclude the involvement of
HSVglycoproteins in adsorption: morethan one
glycopro-tein may becapable ofinteractingwiththecellular receptor,
so that in the absence of any particular one, others may mediate the process. Moreover, any of the glycoproteins
could play a facilitating role, as we show for gC in this
communication. Itis alsopossible, however,that the virion
protein(s) responsible for adsorption is not aglycoprotein,
*Correspondingauthor.
since adsorption canoccur in the absenceofnormal glyco-sylation (7, 28, 55, 57).
We have shownearlier that neomycin selectivelyinhibits theadsorptionof HSV-1 but notHSV-2to BHK cells (30).
Polyaminoacids suchaspolylysineandpolyarginineshowed similar effects, most probably by interference with the receptor (31, 64). We now report experiments in which we
have taken advantage of the difference in drug sensitivity
between HSV-1 and HSV-2 to locate a region of the HSV genome encodingoneor moreproteinsof the virion
attach-mentcomponent.
MATERIALS ANDMETHODS
Materials. Eagle minimal essential medium and newborn
calf serum were purchased from Flow Laboratories, Inc.
Neomycin and poly-L-lysine (Mr 3,700) were from Sigma
Chemical Co. Cell culture dishes were from Nunc.
[35S]methionine
(SJ204;1,270Ci/mmol)wasfrom AmershamCorp.
Virus stocks and antibodies. When not otherwise stated,
the wild-type strains used were HSV-1 17 syn+ (5) and HSV-2 HG52 (59).
[35S]methionine-labeled
viruses weregrownfromthese strainsas described
previously
(31). Theintertypic HSV-1-HSV-2 recombinants used have been characterized earlier (40, 51). The
monoclonal-antibody-resistantgC mutantsofHSV-1, thegC-39mutant, and their
parental strain HSV-1 KOS 321 have been described and
characterized
previously
(23-25). The monoclonal antibod-ies 1001, 1105, and 2454 raisedagainst
gC have also been describedpreviously (47).ThegCantibodies2063,4901,
and 4916werepreparedasdescribedbyCrosset al.(11),except that BALB/c mice were immunized with HSV-1 17 syn+-infected cells.1271
on November 10, 2019 by guest
http://jvi.asm.org/
Cell lines. When not otherwise stated, the cell line used
was BHK 21 clone 13 (37). The other celllines tested were
rabbit skin cells (61), the D-54 humangliomacell line (4), and normalhumanglia cells. The humangliacellswereobtained from nonmalignantglial tissue whichwasremovedaspartof the routine operation on a hematoma. The specimens ob-tained at surgery were cut with scalpels into ca. 0.5-mm3
pieces and placed in 25-cm2 tissue culture flasks together with 2.5 ml of growth medium (Eagle minimal essential medium supplemented with 10% newborn calf serum, four
times the prescribed concentration of nonessential amino
acids, 2%L-glutamine, 100IUofpenicillin perml,100
jig
ofstreptomycin per ml). The earliest outgrowth of cells was seen after 7 days in culture. Further cell growth was slow andcontinued for 5 to 7 weeks untilamonolayercovered the
bottom of the flask. The cells were subcultured nine times
during a 7-month period. At this time they were used in infection experiments. For further details onthe growth of normal humanglia, see reference 50.
Infection and virus growth inthepresenceofdrugs. Plaque
assayswereperformedwithorwithoutdrugspresentfrom 5 min before the addition of virus andthroughout the
adsorp-tionperiod (1h). Medium and drugswerethen replacedby
freshmedium, and the number ofplaques wascounted 48 h later. Adsorption kinetics were studied by removing
un-bound viruses and washing them with phosphate-buffered
saline(pH 7.0) atvarious times after infection,asindicated in the legend toFig. 3.
Antibody assays. A mixture of virus and antibodies was
incubated at 37°C for 30 min prior to dilution of virus, adsorption for 1 h, removal of unbound virus, and further incubation forplaqueassay.Incubationswerecarriedoutin
Eagleminimal essential mediumcontaining 10% heat-inacti-vated serum. Routinely,25
,dl
of ascitic fluidwasused per ml when antibodies were incubated with virus; higher doses gavenofurther inhibition ofadsorption. Bindingof radiola-beled virus was measured similarly, except that after theadsorption periodthe cells wereextensivelywashed and the
remaining bound radioactivity was measured in a liquid scintillation counter. Unspecific binding has not been sub-tractedin the presentationof the data.
Superinfectionexperiments.
[35S]methionine-labeled
viruswas purified on aPercoll gradient as previously described
(31). Superinfection experiments were performed at 4°C.
The cellswerepreincubatedfor2hwith cold HSV-1at200 PFU percell,beforebindingof radiolabeled recombinantsor
wild-type virus for 1 h. Subsequent binding ofhomologous virus was reduced by approximately 50%. Unspecific
bind-ing is not subtracted in thepresentation of the data.
Marker rescue. Purified BamHI a fragment of HSV-2
DNA was cotransfected with HSV-1 (17 syn+) DNA into BHK cells. Thefragment wasfrom strain HVD2577, which
has a BamHI map identical to that of strain HG52.
Trans-fection conditionswereessentiallyasdescribedby Milleret
al. (45). An approximately ninefold excess offragmentwas
used. Plaques were purified and individually tested for
neomycin sensitivity.Twoneomycin-resistantrecombinants
were obtained, which were designated T2(35) and T2(45).
Theywere assayed for incorporation of the BamHI a
frag-ment of HSV-2 DNA into the HSV-1 genome by Southern
blot and hybridization (58) with
32P-labeled
(14) HSV-2 BamHI-a as aprobe.RESULTS
Mapping of a drug-sensitive region on the HSV-1 genome. ElevenHSV-1-HSV-2intertypic recombinantswereusedto
0 0.2 0.4 0.6 0.8 1.0
HSV genome O Sensitivity to
neo polylysine
Bx6(17-1)
Bx 1(28-1-1) U - 1
17+x 11r
Bx1(24) 1
Fx9(5-8) 1 1 . Bx 5(7-2) 1
R E 6
R H6
IFILLL
LLa
R 12-5 =
Dx 1 (34-1)
Dx 1(48)
(0.574) 0.580 0.687 (0.682)
FIG. 1. Drug sensitivity of various intertypic HSV-1-HSV-2 recombinants. The genomic composition of the recombinants is indicated by the upper and lower lines derived from HSV-1 and HSV-2, respectively. A crossoverregion is shown bytwo vertical lines betweenthe horizontal lines. The distances between thetwo vertical lines indicate the region of uncertainty of the crossover event. Where the uncertainty is small, the crossover appears as a single vertical line. The ability of eachrecombinanttoformplaques on BHK cells after exposure to5mMneomycin or 10 ,uMpolylysine was measured as described in the text.
map the genomic region on HSV-1 associated with drug sensitivity. The assay was performed as described
previ-ously for theparental strains of HSV-1 and HSV-2 (31). The sensitivity of each recombinant was tested at various
con-centrations of polylysine between 1 and 15 p.M (results not shown). When the recombinant was sensitive, a maximal effect was observed at 10 p.M or less. Therefore, 5 mM neomycin or 10 F.M polylysine was chosen asthe standard concentration for assay. From previous experiments (31), thesedoseswereknowntoinhibitHSV-1 infection, whereas HSV-2 infection was only slightly affected. Sensitivity and resistance were therefore defined as a reduction in plaque numberbymorethan70%orless than30%, respectively. By these criteria, all recombinants tested were either sensitive
orresistant; noneshowed intermediate sensitivity. Sensitive recombinants were even more strongly affected than wild-type HSV-1, whereas all resistant recombinants were less
strongly affectedthanthe parent HSV-2. The results of these experiments are summarized inFig. 1and Table 1. Correla-tion ofthese data with the genome structures of the recom-binants (Fig. 1) gives a map location for the drug sensitivity delimited on the left by the HSV-2 HpaI d-e restriction
enzyme site and on the right by theHSV-1 Kpn z-u restric-tion enzyme site. Thecoordinates for these siteswere 0.574 and 0.682, until determination of the DNA sequence (41) allowed amoreprecise location to 0.580 and 0.687 (49). The
formersetof coordinatesare indicatedatthefoot ofFig. 1in
parentheses.
The assumption that the neomycin-sensitive regioncodes forproteins involved in specific receptor binding was
con-firmed in superinfection assays not involving the use of
on November 10, 2019 by guest
http://jvi.asm.org/
GENOMIC LOCALIZATION OF HSV-1 ADSORPTION PROTEINS TABLE 1. PlaqueformationbyHSV-1-HSV-2 intertypic
recombinants inthe presence of drugs
Plaqueformation (% of control)
Recombinant in presence of:
Neomycin Polylysine
Bx6(17-1) 30 13
Bx1(28-1-1) 36 29
17+x 11r 24 23
Bx1(24) 30 27
Fx9(5-8) 28 28
Bx5(7-2) 19 18
RE6 9 25
RH6 13 29
R12-5 83 80
Dx1(34-1) 79 82
Dx1(48) 105 85
drugs. BHK cells were preincubated with unlabeled
wild-type HSV-1 (17+ strain) at a multiplicity ofmore than 200
PFU per cell. After 2 h ofpreincubation at 4°C, the cells
were superinfected with [35S]methionine-labeled wild-type
HSV-1, wild-type HSV-2, Bx1(24), and R12.5. The bound radioactivitywas compared with that obtained without
pre-incubation withunlabeled HSV-1 and was47, 110, 54, and
100% of this maximal binding, respectively. The results indicate that HSV-1 blocks subsequent binding of HSV-1
and recombinant Bx1(24), whereas the binding ofHSV-2 and recombinant R12.5 was unaffected. These data are
consistent withthe dataobtained with neomycin and poly-lysine.
Marker rescue. Experiments were performed in which
DNA from neomycin-sensitive HSV-1 was cotransfected
with the BamHI a fragment of HSV-2 into BHK cells. The
BamHIa fragment of HSV-2 has coordinates 0.604 to0.69
close to those of the neomycin-sensitive region on the
HSV-1 genome. Two neomycin-resistant viruses were
ob-tained (Table 2). DNA preparations from these were di-gested with BamHIand testedbySouthern blotanalysisfor
incorporation of BamHI-a HSV-2 DNA into the HSV-1 genome. DNAsfrom parentalHSV-1 and HSV-2aswellas
fromtwoneomycin-sensitive viruses isolatedafter transfec-tion were used as controls. The probe, which was
radiola-beledBamHIafragment,gave a stronghybridization signal
withanHSV-2-specific band migratingmoreslowly thantwo
moreweaklylabeled bands from HSV-1. Thetwo
neomycin-sensitive viruses T2(22) and T2(3) gave signals similar to those of the HSV-1 17 syn+ strain, whereas the
neomycin-resistant viruses T2(45) and T2(35) contained a slowly
[image:3.612.59.298.96.238.2]mi-grating radiolabeled band consistent with incorporation of HSV-2 sequences (data not shown). It is thus clear that
TABLE 2. Conversiontoneomycin resistance by incorporation
ofsequencesfrom the BamHIafragmentof HSV-2
into the HSV-1genomea
Virusisolated % Inhibition of plaque Presence of HSV-2 aftertransfection formation by 5 mM BamHI-a sequences
neomycinq
T2(3) 82
T2(22) 83
T2(35) 11 +
T2(45) 14 +
[image:3.612.312.558.611.715.2]aCotransfectionwithcompleteHSV-1 DNA andthe BamHIafragmentof HSV-2.
TABLE 3. Effect of5 mMneomycinonplaque formation bygC mutantsandwild-type HSV-1 and HSV-2
% Plaqueformation with: Virus
Noneomycin 5mMneomycin
17syn+ 100 <10
KOS 321 100 36
MAR C 11.1 100 29
MAR C 4.3 100 17
MAR C 9.6 100 39
MAR C 16.1 100 22
gC-39 100 34
HSV-2 HG-52 100 90-100
presence of theHSV-2BamHIafragment confers neomycin resistance (Table 2).
Role ofgC in adsorption and drug sensitivity. The only glycoprotein yet established to be present in the
drug-sensitive region of HSV-1 is gC. We therefore tested the potential role of gC inadsorption and drug sensitivity. First, we examined the neomycin sensitivity of various HSV-1
mutants resistant to monoclonal antibodies against gC, as
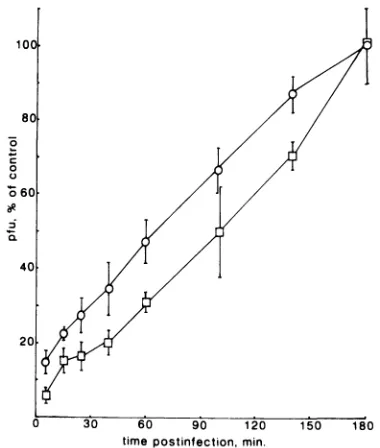
well as a deletion mutant (gC-39)which did not synthesize gC (Table 3). Under the experimental conditions used, the parentalwild-type strain HSV-1 KOS 321 showed somewhat lowerneomycin sensitivity thandid strain 17 syn+, used in the previous experiment. However, all mutants, gC-39 in-cluded,showed similardrug sensitivitytothat of the paren-talwild-typestrain. Wealsocompared adsorptionkineticsof gC-39and itsparentKOS 321 (see Fig.3). The data show a
delay in adsorption of 20 to 30 min for the gC-negative
mutantcompared with theparentvirus. Finally, theeffects of various monoclonal antibodies directed against gCwere
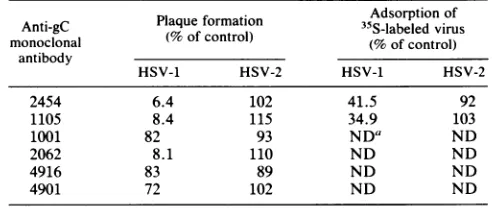
tested with regard to infectivity and adsorption ofHSV-1 andHSV-2(Table 4). Of the six monclonal antibodies tested,
three strongly inhibited HSV-1 infection, whereas HSV-2 infectionwasunaffectedbyallof them. Two of the
antibod-ies (2454 and 1105) inhibiting HSV-1 infection were also tested for their ability to inhibit the adsorption of radiola-beled virus. Again, HSV-2 adsorption was unaffected,
whereas there was a marked reduction in adsorption of HSV-1.
Drug sensitivity of HSV-1 17 syn+ infection invarious cell lines.Todetermine whether thedrugsensitivityof virus-cell interaction was specificto infection of BHKcells, we also
performed similarplaqueassays in rabbit skincells, human
gliomacells(D-54),and normal humanglialcells.Sensitivity
to neomycin (5 mM) was observed in all the cells tested
(15.3, 18.7, 31.6, and 14.5% of control values for BHK,
TABLE 4. Inhibitionof infection andadsorption by monoclonal
antibodiesagainst gC
Adsorption of Anti-gC Plaqueformation 35S-labeledvirus monoclonal (%of control) (%ofcontrol)
antibody
HSV-1 HSV-2 HSV-1 HSV-2
2454 6.4 102 41.5 92
1105 8.4 115 34.9 103
1001 82 93 NDa ND
2062 8.1 110 ND ND
4916 83 89 ND ND
4901 72 102 ND ND
aND,Notdetermined.
VOL. 1990 1273
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.58.297.635.706.2]rabbit, glioma, and glial cells, respectively). Experimental conditions were identical to those described for Fig. 1.
DISCUSSION
This study is concerned with HSV adsorption: the first step in the infectious process. Using intertypic HSV-1-HSV-2 recombinants, we have identified a region on the HSV-1 genome encoding a protein(s) responsible for sensi-tivity to neomycin and polylysine. Previous results (31) indicate that this sensitivity is due to an interaction between the drug and the cellular receptor which, in turn, inhibits virus attachment. Mapping of drug sensitivity therefore indirectly maps one or more proteins in the virion attach-ment component, a term originally proposed by Fuller and Spear (17) for the virus structure which mediates binding to cells. Our results demonstrate that this protein(s) is encoded within the same map units, 0.580 and 0.687, on both the HSV-1 and HSV-2 genomes. If this were not so, one would expect virions of one or more of the recombinants to contain both type 1 and type 2 attachment component protein(s). Such arecombinant would be insensitive to the drugs despite thepresence of HSV-1 genome between map units 0.580 and 0.687. However, none of the eight recombinants with type 1
sequences inthis region (Fig. 1) were insensitive.
These findings do not rule out the possibility of more than one type of attachment site on each virus type. Any site encoded outside the mapped region would be insensitive to the drugs. This could, for instance, explain the fact that HSV-1 infection is never inhibited 100% by neomycin or
polylysine. Thepresence of high- and low-affinity receptors, for instance, would thus explain our results. This may, however, not be very likely, since increasing the doses of neomycin beyond 5 mM also increased the inhibitory activ-ity of the drug, and this therefore argues against an alterna-tive binding mechanism insensitive to neomycin. There is,
correspondingly, also the theoretical possibility that such a
low-affinity attachmentis to the other type-specific receptor. Again, there is no evidence for such high- and low-affinity binding, even though the possibility may not have been studied closely enough.
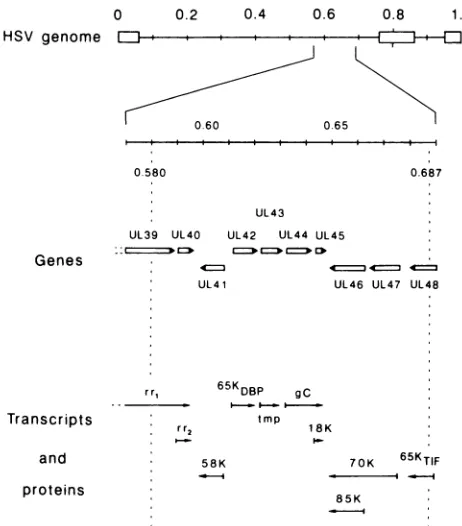
The region between map units 0.580 and 0.687 (Fig. 2) contains eight complete genes (UL40 to UL47) and the 3' portion of two other genes (UL39 and UL48) (41). Of these 10genes, 5 encode identified proteins: the large and small subunits ofribonucleotide reductase (UL39 and UL40, re-spectively [2, 3, 10, 15, 52]); a 65-kilodalton double-stranded
DNA-binding protein (UL42 [18, 39]); virion gC (UL44 [13,
16]); and the 65-kilodalton virion-transinducing factor (8). Theremaining genes potentially encode a virion component responsible for the shutoff of host protein synthesis (UL41 [29,41]),transmembraneproteins (UL43 [41] and UL45 [16, 41]), andgenesinvolved in modulation of the virion-transin-ducingfactor (UL46 and UL47 [44]). The proteins encoded by this second group of five genes remain to be identified. gC isnot theproteinassociated with sensitivity to neomycin and
polylysine, as demonstrated by experiments with the gC mutants (Table 3). These results are consistent with reports that gC isnotessential for infectivity. Although not directly involved, gC seems somehow to facilitate receptor binding, since a gC-negative mutant attached more slowly to cells than itsparental virus did (Fig. 3). It must be noted that these experiments are limited by the lack of rescued-virus exper-iments andshouldtherefore be interpreted with caution. The observed reduction in attachment rate is, however, consis-tentwith thedataobtained by Stannard et al. (56), who used
0 HSVgenome
EJ-0.2 0.4 0.6 0.8 1.0
0 60 0.65
0.580
UL43
UL39 UL40 UL42 UL44 UL45
Genes
UL41 ULt
rr, 65KDBP gC
*- * l_F ""-|
Transcripts tmp
rr2 18K and
proteins
58K
0.687
46 UL47 UL48
70K 65KTIF
-_
85K
FIG. 2. Gene organization in the drug-sensitive region of HSV-1. The scale of the genome has been expanded to allow individual transcripts and protein mapping in the neomycin-sensitive region to be depicted. Vertical dotted lines indicates the limits of the drug-sensitive region. The genes are designated by the system of McGeoch et al. (41).
immunoelectron microscopy to examine the distribution of glycoproteins on virions. They found gC in long, slender structures (up to 24 nm in length). Such projections could well contribute to the initial interactions between virions and cells and enhance the adsorption rate. Interestingly, it has been reported that the gC homolog in pseudorabies virus,
gIll,
although not essential for virus growth, is important forefficient virus growth in culture (64), and
gIll-mutants
showed a reduced rate of adsorption (54).Three different monoclonal antibodies directed against gC inhibited infection with HSV-1 by more than 90%, and two of these reduced the binding of radioactive virus (Table 4). Such blocking might be produced by antibodies which bind to a protein outside of but close to the attachment compo-nent, since they would sterically hinder interaction between the component and the receptor.
A second protein which can be excluded from our consid-erations as a candidate for the virion attachment component is the 65-kilodalton DNA-binding protein, since it is not found in virions (39; H. S. Marsden, unpublished data).
Nonglycosylated proteins must also be considered as candidates for the virion attachment component, since HSV-1 virions, produced when glycosylation is inhibited, adsorb to cells (28, 57). The putative transmembrane proteins (en-coded by genes UL43 and UL45) and nontransmembrane proteins (encoded by UL39,UL40,UL41, UL46, UL47, and UL48) all fall into this category. Although it is likely that proteins in the virion attachment component will be mem-brane anchored, the possibility that a nontransmemmem-brane virion protein linked to a protein on the virion surface is involved in adsorption should not be excluded.
Our observation that infectivity is sensitive to neomycin in
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.318.549.71.334.2]100X
80
0
060
40
20
0 30 60 90 120 150 180
time postinfection, min.
FIG. 3. Adsorption kineticsofthe gC-negativemutantgC-39and
wild-type parent KOS 321. Cells were infected at time zero with
KOS 321 (0) orgC-39
(O).
At the times indicted, the cells werewashed with phosphate-buffered saline(pH 7.2)toremoveunbound
virusand incubated further for the plaque assay. Eachpoint is the average ofsixindividual determinations fromtwo different
experi-ments.
several cell types indicates that HSV-1 binds to the same
receptorin the different celltypes, butdoes notexcludethe existence of additional binding mechanisms varying from
one cell line to another. Ofparticular interest is the drug sensitivity of the normal human glial cells, indicating that human nerve tissue shares receptor-binding mechanisms
with cells from other species and with transformed and
permanentcelllines.
Recently,heparansulfate has beenidentified as theinitial
receptor for both HSV-1 and HSV-2 (65). The authors proposed that the interaction might be thefirst inacascade
of virion-cell interactions,culminatingin fusionof the virion with theplasma membrane. Wepresumethat thecomponent
we have mapped interacts with heparan sulfate either
di-rectly or via some other protein. WuDunn and Spear (65)
also used polylysine to block adsorption of virus. They found that although polylysine was type selective in thatit blocked adsorption ofHSV-1 but not HSV-2, infections of bothserotypeswereequallyinhibited in plaque assay
exper-iments. This contrasts with our previous data (31), which
showed type selectivity also in plaque assays. This was,
however,dose dependent,and theuseofahigherpolylysine
dose (or longer lysine polymers) in the experiments of WuDunn and Spear (65) could explain the discrepancy in results. Also importantisthatdifferentcelllines wereused.
There are apparently conflicting reports concerning the existence of serotype-specific receptors. Two provide evi-dence in favor of their existence. In the first set of experi-ments,UV-irradiated viruses ofone serotypewereadsorbed
to cells, and the ability of viruses ofthe other serotype to subsequently adsorbtothose cells wasinhibited (60). Inthe
second report a temperature-sensitive mutant of HSV-1, ts1204, was used; this mutant is blocked at the adsorption
stage at the nonpermissive temperature. If this mutant was
first adsorbed to cells at the nonpermissive temperature,
expressionofsuperinfectingwild-typeHSV-1, butnot HSV-2, in those cells was reduced (1). On the other hand, WuDunnand Spear (65) reported competitive-binding
exper-iments with fully infectious virus which showed that adsorp-tion ofone serotype interfered with that ofthe other. Our results favor the existence of serotype-specific receptors,
butdonotexclude thepossibilityofsome mechanisms in the cascade of events leading to uptake common to the two serotypes.
ACKNOWLEDGMENTS
We aregratefulto RolfBjerkvig, theGade Institute, Department ofPathology at Haukeland Hospital for providing the normalhuman glia cells and for establishing theirgrowth conditions. The HSV-2
BamHI A fragment inserted into the pBR322 vector was the generous gift ofValerie Preston, Medical Research Council Virol-ogy Unit, Glasgow, Scotland. We thank Johan Lillehaug forfruitful discussions, particularly concerningthe Southern blot work.
Thisworkwassupported by grants from the NorwegianResearch Council for Science and the Humanities, from the Norwegian Society for Fighting Cancer, and from L. Meltzers H0yskolefond.
LITERATURE CITED
1. Addison, C., F. J. Rixon, J. W. Palfreyman, M. O'Hara, and V. G. Preston. 1984. Characterisation of a herpes simplex virus type 1 mutant which has a temperature-sensitive defect in penetration of cells and assembly of capsids. Virology 138:
246-259.
2. Baccetti, S., M. J. Eveleigh, and B. Muirhead. 1986. Identifica-tion and separaIdentifica-tion of the two subunits of the herpes simplex virus ribonucleotide reductase. J. Virol. 57:1177-1181. 3. Baccetti, S., M. J. Eveleigh, B. Muirhead, and C. S. Sartori.
1984. Immunological characterization of herpes simplex type 1 and 2 polypeptide(s) involved in viral ribonucleotide reductase activity. J. Virol. 49:591-593.
4. Bigner, D. D., S. H. Bigner, J. Ponten, B. Westermark, M. S. Mahaley, E. Ruoslahti, H. Herschman, L. F. Eng, and C. J. Wikstrand. 1981. Heterogeneity ofgenotypic and phenotypic characteristics of 15 permanent cell lines derived from human gliomas. J. Neuropathol. Exp. Neurol. 40:201-229.
5. Brown, S. M., D. A. Ritchie, and J. H. Subak-Sharpe. 1973. Genetic studies with herpes simplex virustype1. Theisolation of temperature-sensitive mutants, their arrangement into com-plementation groups and recombination analysis leading to a linkage map. J. Gen. Virol. 18:329-346.
6. Cai, W., B. Gu, and S. Person. 1988. Role of glycoprotein B of herpes simplex virus type 1 in viral entry and cell fusion. J. Virol. 62:2596-2604.
7. Campadelli-Fiume, G., L. Poletti, F. Dall'Olio, and F. Serafini-Cessi. 1982. Infectivity and glycoprotein processing of herpes simplex virus type 1 grown in a ricin-resistant cell linedeficient in N-acetylglucosaminyl transferaseI. J. Virol. 43:1061-1071. 8. Campbell, M. E. M., J. W. Palfreyman, and C. M. Preston.
1984. Identification of herpes simplex virus DNA sequences which encode a trans-acting polypeptide responsible for stimu-lation of immediate early transcription. J. Mol. Biol. 180:1-19. 9. Cassai, G., R. Manservigi, A. Corallini, and M. Terni. 1975/76. Plaque dissociation of herpes simplex viruses: biochemical and biological characters of the viral variants. Intervirology 6: 212-223.
10. Cohen, E. A., J. Charron, J. Perret, and Y. Langelier. 1985. Herpes simplex ribonucleotide reductase induced in infected
BHK-21/C13 cells: biochemical evidence for the existence of two non-identical subunits, Hi and H2. J. Gen. Virol. 66: 733-745.
11. Cross, A. M., R. G. Hope, and H. S. Marsden. 1987. Generation and properties of the glycoprotein E-related 32K/34K/35K and
55K/57Kpolypeptides encoded by herpes simplex virus type 1. J. Gen. Virol. 68:2093-2104.
12. Desai, P. J., P. A. Schaffer, and A. C. Minson. 1988. Excretion of non-infectious virus particles lacking glycoprotein H by a
VOL.64, 1990 1275
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.81.271.72.296.2]temperature-sensitive mutant of herpes simplex virus type 1:
evidencethatgH is essential forvirioninfectivity. J. Gen.Virol.
69:1147-1156.
13. Draper, K. G., R. H. Costa, G. T.-Y. Lee, P. G. Spear, and E.K.
Wagner. 1984. Molecular basis of the glycoprotein-C-negative phenotype of herpes simplex virus type1macroplaque strain. J.
Virol. 51:578-585.
14. Feinberg, A. P., and B. Vogelstein. 1984. A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 137:266-267. (Addendum.)
15. Frame, M. C., H. S. Marsden, and B. M. Dutia. 1985. The
ribonucleotide reductaseinducedby herpes simplexvirus type1 involves minimally a complex oftwo polypeptides (136Kand
38K).J. Gen. Virol. 66:1581-1587.
16. Frink, R.J.,R.Eisenberg, G. Cohen, andE.K. Wagner. 1983. Detailedanalysis of the portion oftheherpes simplex virustype
1genomeencoding glycoproteinC. J. Virol. 45:634-647.
17. Fuller,A.O., and P. G. Spear. 1985. Specificities of monoclonal
and polyclonal antibodies that inhibit adsorption of herpes simplex virus to cells and lack of inhibition bypotent neutral-izing antibodies. J. Virol. 55:475-482.
18. Gallo,M.L., D. H. Jackwood,M.Murphy,H.S. Marsden, and
D.S.Parris. 1988. Purification of the herpes simplex virustype
165-kilodalton DNA-binding protein: properties of the protein
and evidence of its association with the virus-encoded DNA polymerase. J. Virol.62:2874-2883.
19. Gompels, U., andA.Minson. 1986.Theproperties andsequence of glycoprotein H of herpes simplex virus type 1. Virology
153:230-247.
20. Harland, J., and M. Brown. 1988. Generation of a herpes simplexvirus type 2variat devoid ofXbaIsites: removal of the 0.91mapcoordinate site resultsinimpaired synthesis of glyco-protein G-2. J. Gen. Virol. 69:113-124.
21. Heine,J. W., R.W. Honess,E.Cassai, and B. Roizman. 1974.
Proteins specified by herpes simplex virus. XII. The virion
polypeptidesof type 1 strains. J.Virol. 14:640-651.
22. Hoggan, M. D., and B. Roizman. 1959. The isolation and propertiesofavariantofherpes simplex producing multinucle-atedgiant cells in monolayer culture in the presence of anti-body.Am. J. Hyg.70:208-219.
23. Holland, T. C., F. L. Homa, S. D.Marlin,M. Levine, and J.
Glorioso. 1984. Herpes simplex virus type 1 glycoprotein C-negativemutants exhibitmultiple phenotypes, including secre-tionof truncatedglycoproteins. J. Virol. 52:566-574.
24. Holland,T.C.,S. D.Marlin, M. Levine, and J. Glorioso. 1983. Antigenicvariants ofherpes simplexvirusselected with glyco-protein-specific monoclonalantibodies. J. Virol. 45:672-682.
25. Homa,F. L.,D.J. M. Purifoy, J. C. Glorioso, andM.Levine. 1986. Molecular basis of the glycoprotein C-negative
pheno-typesofherpes simplex virus type 1 mutants selected with a
virus-neutralizing monoclonal antibody. J.Virol. 58:281-289. 26. Johnson,D. C.,and V.Feenstra. 1987. Identification ofanovel
herpes simplex virus type 1-induced glycoprotein which
com-plexes with gE and binds immunoglobulin. J. Virol.
61:2208-2216.
27. Johnson, D. C., M. C. Frame,M. W. Ligas,A. M.Cross, and N. D. Stow. 1988. Herpessimplex virus immunoglobulin G Fc
receptoractivity dependson acomplex oftwo viral glycopro-teins,gEandgI.J. Virol. 62:1347-1354.
28. Kuhn, J. E., B. R. Eing, R. Brossmer, K. Munk, and R. W. Braun. 1988.Removal ofN-linked carbohydrates decreases the infectivity of herpes simplex virus type 1. J. Gen. Virol.
69:2847-2858.
29. Kwong, A. D., J. A. Kruper, and N. Frenkel. 1988. Herpes simplexvirus virionhostshutofffunction. J. Virol.62:912-921.
30. Langeland, N.,H.Holmsen, J.R.Lillehaug, and L. Haarr. 1987. Evidencethatneomycin inhibitsbinding ofherpessimplexvirus type 1tothecellularreceptor. J. Virol. 61:3388-3393.
31. Langeland, N.,L. J. Moore, H. Holmsen, and L. Haarr. 1988. Interaction ofpolylysine with the cellular receptorforherpes
simplexvirustype 1. J. Gen. Virol. 69:1137-1145.
32. Ligas,M.W.,and D. C. Johnson.1988. A herpessimplex virus mutant in which glycoprotein D sequences are replaced by
,B-galactosidase sequences bindsto but is unable to penetrate into cells. J. Virol. 62:1486-1494.
33. Little, S. P., J. T. Jofre, R. J. Courtney, andP. A. Schaffer.
1981. Avirion-assosiated glycoproteinessential for infectivity ofherpes simplex virus type 1. Virology 115:149-160. 34. Longnecker, R., S.Chatterjee, R.J. Whitley, and B. Roizman.
1987. Identification of a herpes simplex virus 1 glycoprotein gene within a gene cluster dispensable forgrowth in cell culture. Proc. Natl. Acad. Sci. USA 84:4303-4307.
35. Longnecker, R., and B. Roizman. 1986.Generation of an invert-ing herpes simplex virus 1 mutant lacking the L-S junction a sequences, an origin of DNA synthesis, and several genes includingthosespecifying glycoprotein E and the a 47 gene. J. Virol. 58:583-591.
36. Longnecker, R., and B. Roizman. 1987. Clustering of genes dispensible forgrowth in culture in the S component of the HSV-1genome. Science 236:573-576.
37. Macpherson, I., and M. Stoker. 1962. Polyomatransformation of hamster cell clones-an investigation of genetic factors affectingcell competence. Virology16:147-151.
38. Marsden, H. S. 1987. Herpes simplexvirus glycoproteins and pathogenesis, p. 259-288. In W. C. Russell and J. W. Almond (ed.), Molecularbasis of virus disease. Cambridge University Press,Cambridge.
39. Marsden,H.S., M. E. M.Campbell, L. Haarr, M. C. Frame,
D.S. Parris,M.Murphy,R.G.Hope, M. T. Muller, and C. M. Preston. 1987. The 65,000-Mr DNA-binding and virion trans-inducing proteins of herpes simplex virus type 1. J. Virol. 61:2428-2437.
40. Marsden,H.S., N.D.Stow,V.G.Preston, M. C.Timbury, and N. M.Wilkie. 1978. Physical mappingofherpes simplex virus-inducedpolypeptides.J. Virol.28:624-642.
41. McGeoch, D. J., M. A. Dalrymple, A. J. Davison, A. Dolan,
M.C. Frame, D. McNab,L.J. Perry, J.E.Scott, and P. Taylor. 1988. Thecomplete DNA sequenceof the long unique region in the genome of herpes simplex virus type 1. J. Gen. Virol. 69:1531-1574.
42. McGeoch,D.J., and A.J. Davison. 1986.DNAsequenceof the herpessimplexvirus type 1geneencoding glycoproteingH, and identification ofhomologuesin the genomes of varicella-zoster virus andEpstein-Barr virus.NucleicAcidsRes.14:4281-4292. 43. McGeoch, D. J., A. Dolan, S. Donald, and F. J. Rixon. 1985. Sequence determinationandgeneticcontentofthe short unique region in the genome of herpessimplex virus type-1. J. Mol. Biol. 181:1-13.
44. McKnight, J.L.C.,P. E.Pellett,F.J.Jenkins,and B.Roizman. 1987. Characterization and nucleotide sequence of twoherpes simplex virus1geneswhoseproducts modulateatrans-inducing factor-dependent activation ofagenes. J.Virol.61:992-1001. 45. Miller, G., P. Wertheim, G. Wilson, J. Robinson, J. L.M. C.
Geelen, J. van der Noordaa, and A. J. van der Eb. 1979. Transfection ofhuman lymphoblastoid cells with herpes sim-plexviral DNA. Proc. Natl. Acad. Sci. USA 76:949-953. 46. Neidhardt, H., C. H. Schroeder, and H. C. Kaerner. 1987.
Herpessimplexvirus type 1glycoproteinEis notindispensible forviralinfectivity.J. Virol. 61:600-603.
47. Palfreyman,J. W., L. Haarr, A.Cross, R. G. Hope, and H. S. Marsden. 1983. Processing of herpes simplex virus type 1 glycoproteins: two dimensional gel analysis using monoclonal antibodies. J. Gen. Virol. 64:873-886.
48. Para,M.F.,R. B.Baucke, and P. G. Spear. 1980. Immunoglob-ulinG(Fc)bindingreceptors onvirions of herpes simplex virus type 1 and transfer ofthese receptors to the cell surface by infection. J. Virol. 34:512-520.
49. Parris, D. S., A. Cross, L. Haarr, A. Orr, M. C. Frame, M. Murphy, D. J. McGeoch, and H. S. Marsden. 1988. Identifica-tionof thegeneencodingthe 65-kilodalton DNA-bindingprotein ofherpessimplex virustype 1. J. Virol.62:818-825.
50. Ponten, J., and E. H. Macintyre. 1968. Long term culture of normal and neoplastic human glia. Acta Pathol. Microbiol. Scand. 74:465-486.
51. Preston,V. G.,A.J. Davison,H. S.Marsden, M. C. Timbury, J. H. Subak-Sharpe, and N. M. Wilkie. 1978. Recombinants
on November 10, 2019 by guest
http://jvi.asm.org/
betweenherpes simplexvirustypes 1 and 2:analyses of genome structures and expression ofimmediate early polypeptides. J. Virol. 28:499-517.
52. Preston, V. G., J. W. Palfreyman, and B. M. Dutia. 1984. Identification of a herpes simplex virus type 1 polypeptide which is a component ofthe virus-induced ribonucleotide re-ductase. J.Gen. Virol. 65:1457-1466.
53. Sarmiento, M., M. Haffey, and P. G. Spear. 1979. Membrane proteins specified by herpes simplex viruses. III. Role of glycoprotein VP7(B2) in virion infectivity. J. Virol. 29:1149-1158.
54. Scheurs, C., T. C. Mettenleiter, F.Zuckermann, N. Sugg, and T. Ben-Porat. 1988. Glycoprotein glll of pseudorabies virus is multifunctional. J. Virol. 62:2251-2257.
55. Spivack, J. G., W. H. Prusoff, and T. R.Tritton. 1982. A study of the antiviral mechanism of action of 2-deoxy-D-glucose: normally glycosylated proteins are not strictly required for herpes simplexvirus attachment but increase viral penetration andinfectivity. Virology 123:123-138.
56. Stannard, L. M., A. 0. Fuller, and P. G.Spear. 1987. Herpes simplex virus glycoproteins associated with different morpho-logical entities projecting from the virion envelope. J. Gen. Virol. 68:715-725.
57. Svennerholm, B., S. Olofsson, R. Lunden, A. Vahlne, and E. Lycke. 1982. Adsorption and penetration of enveloped herpes simplex virus particles modified by tunicamycin or 2-deoxy-D-glucose. J.Gen. Virol. 63:343-349.
58. Thomas, P.S. 1980.Hybridization of denaturedRNAand small
DNAfragments transferred to nitrocellulose. Proc. Natl. Acad. Sci. USA 77:5201-5205.
59. Timbury, M. C. 1971.Temperature-sensitive mutantsofherpes simplexvirus type 2. J. Gen. Virol. 13:373-376.
60. Vahlne, A., B. Svennerholm, and E. Lycke. 1979. Evidence for herpessimplexvirustype-selectivereceptors on cellularplasma membranes. J.Gen. Virol.44:217-225.
61. Watson, K., J. G.Stevens, M. L. Cook, and J. H.Subak-Sharpe. 1980. Latency competence of thirteen HSV-1 temperature-sensitive mutants. J. Gen. Virol. 49:149-159.
62. Weber, P. C., M. Levine, and J. C. Glorioso. 1987. Rapid identification of nonessentialgenesofherpes simplexvirus type 1 byTnSmutagenesis. Science236:576-579.
63. Weller, S. K., D. P. Aschman, W. R. Sacks, D. M. Coen, and P. A.Schaffer. 1983. Genetic analysis of temperature-sensitive mutantsof HSV-1: thecombined useofcomplementation and physical mapping for cistron assignment. Virology 130:290-305. 64. Whealy, M. E., A. K. Robbins, and L. W. Enquist. 1988. Pseudorabies virus glycoprotein glll is required for efficient virusgrowth in tissue culture. J. Virol. 62:2512-2515.
65. WuDunn, D., and P. G.Spear.1989.Initialinteraction ofherpes simplex virus with cellsisbindingtoheparan sulfate. J. Virol. 63:52-58.
66. Zezulak, K.M., and P. G. Spear. 1984. Mappingof the
struc-tural gene for the herpes simplex virus type 2 counterpart of herpessimplexvirus type 1glycoprotein Candidentificationof atype 2 mutant which does not express this glycoprotein. J. Virol.49:741-747.